










Servicios Personalizados
Revista

Articulo
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO  uBio
uBio
Compartir

 Permalink
PermalinkInVet
versión On-line ISSN 1668-3498
InVet vol.18 no.1 Ciudad Autónoma de Buenos Aires jun. 2016
ARTÃCULO DE INVESTIGACIÃN
Efectos de la restricción alimentaria sobre el desarrollo de los vasos sanguÃneos placentarios en cabras
Coniglio, MV1.; Merkis, CI2.; Diaz, T3.; Romanini, MC3.; Turiello, MP1.; Bozzo, AA3.; Cots, DS3. y Rolando, AN3.
1Departamento de Producción Animal, FAV- UNRC,
2Ãrea de MicroscopÃa Electrónica, FAV-UNRC,
3Departamento de AnatomÃa Animal FAV-UNRC. Ruta 36 Km 601. 5800 RÃo Cuarto.
Correspondencia e-mail: vconiglio@ayv.unrc.edu.ar
Recibido: 1/08/2015
Aceptado: 18/05/2016
Resumen
El objetivo fue estudiar los efectos de la restricción alimentaria sobre la angiogénesis placentaria en cabras Anglo nubian. Se utilizaron 19 placentas a término de tres grupos de estudios: control, restringido y restringido con monensina. Los cortes placentarios se procesaron con técnicas histológicas de rutina e inmunomarcaron con anticuerpo anti-VEGF y se cuantificó el área y perÃmetro de los vasos sanguÃneos. El perÃmetro de los vasos de los grupos restringidos fue mayor estadÃsticamente con respecto al control. Mientras que el área de los mismos no mostró diferencias estadÃsticamente significativas en ninguno de los tres grupos. El VEGF se localizó en las células epiteliales trofoblásticas de los grupos control y restringido, además en éstos últimos también se localizó en el endotelio vascular. El número de células inmunomarcadas con el VEGF fue significativamente mayor en los grupos restringidos coincidiendo con el mayor perÃmetro de los vasos, lo que sugiere que el VEGF estimula el crecimiento endotelial. Las diferencias encontradas entre los grupos estudiados, indicarÃan que hubo una adaptación compensatoria en las cabras que fueron sometidas a una dieta restringida, ya que el área vascular se mantuvo sin modificaciones a expensas del aumento del perÃmetro de los vasos.
Palabras clave: Placenta; Angiogénesis; VEGF; Cabra.
Effects of nutritional restriction on placental blood vessels development in goats
Summary
The aim was to study the effects of nutritional restriction on placental angiogenesis in Anglo nubian goats. 19 term placenta of three groups were used: control, restricted and restricted with monensin. Tissue sections were processed by routine histological techniques and immunostained with the anti-VEGF antibody and the area and perimeter of the blood vessels were quantified. The perimeter of vessels of the restricted groups was statistically higher compared to the control. While the area of the vessels do not show significant differences in any of the 3 groups. VEGF was localized in trophoblastic epithelial cells control and restricted, also in the latter was also localized to the vascular endothelium. The number of immunolabeled VEGF cells was higher in the restricted groups, coinciding in these groups with the larger perimeter of the vessels, so VEGF would stimulate the growth of the vascular endothelium. The differences between the restricted and the control groups, suggest that there was a compensatory adaptation of goats subjected to a restricted diet, because the vascular area remained unchanged due to the increase of the perimeter of the blood vessels.
Key words: Placenta; Angiogenesis; VEGF; Goat.
Introducción
En Argentina la producción caprina, en su mayorÃa, se desarrolla utilizando el pastizal natural, el cual presenta variaciones anuales y un marcado descenso de la oferta forrajera en el periodo invernal. Ante esta situación, los animales no alcanzan a cubrir sus requerimientos nutritivos en esta época del año afectando la tasa de crecimiento de las hembras en el perÃodo prepuberal, puberal y postpuberal3.
Investigaciones realizadas con cabras jóvenes prepúberes, Anglo nubian, establecieron que una restricción alimentaria de un 30 % del consumo de materia seca afecta el crecimiento de las mismas, pero se permite mantener un nivel de metabolitos y hormonas compatibles con el desenlace de la pubertad2,14,15. En los sistemas caprinos el uso de monensina tendrÃa, como en otros rumiantes, un efecto compensador ante las restricciones energéticas en el manejo nutricional5. Rivera et al. (2003), demostraron que el uso de monensina provocó una tendencia favorable en el diámetro del folÃculo ovulatorio, en la ovulación y en la tasa de preñez de cabras.
La nutrición materna condiciona el desarrollo placentario, el tamaño de la placenta y el correcto intercambio nutricional maternofetal16. Asimismo, el mantenimiento de la preñez depende en gran parte del intenso crecimiento y formación de nuevos vasos sanguÃneos o angiogénesis, asociados con el desarrollo de una eficiente interfase maternofetal12. Un compromiso significativo de la vascularización de la placenta, ocasionado por una mala nutrición materna, puede menoscabar el intercambio de nutrientes, de gases y residuos entre el sistema materno y fetal8, teniendo un gran impacto en la supervivencia neonatal y el desarrollo fetal10.
La densidad del área capilar (DAC) y la densidad del número de capilares (DNC) son parámetros usados como medidas del desarrollo vascular. En este sentido, estudios realizados en ovejas por Vonnahme (2006) demostraron que la DAC y la DNC aumentan exponencialmente tanto en la parte caruncular como en la cotiledonaria desde el dÃa 40 de preñez hasta el final de la gestación. No obstante, si bien el crecimiento es exponencial en ambos tejidos, proporcionalmente es mayor el desarrollo en la parte cotiledonaria. El crecimiento en el tejido caruncular se acompaña por un pequeño aumento del número de capilares y un gran incremento del diámetro capilar, mientras que en el tejido cotiledonario hay aumento del número de capilares con una disminución en el diámetro capilar7. Para una misma área de sección transversal, muchos vasos de pequeño calibre permiten una mayor eficiencia de intercambio capilar en comparación con pocos vasos de gran diámetro, porque el área superficial (perÃmetro o área de superficie de los capilares por unidad de tejido) disponible para el intercambio es mayor11.
Numerosos factores están implicados en la angiogénesis, siendo uno de los más importantes el factor de crecimiento vascular endotelial (VEGF)9 , que es un estimulador del proceso de angiogénesis6. En las placentas de cerdos se demostró que la expresión del VEGF aumenta progresivamente durante la gestación en asociación con el marcado incremento en el número de vasos sanguÃneos en la interfase materno-fetal17.
Teniendo en cuenta las condiciones de alimentación a las que son sometidas las cabras por las variaciones estacionales de las pasturas naturales y que los estudios en placenta de las mismas son escasos o nulos en Argentina, se propuso estudiar el desarrollo de los vasos sanguÃneos y la localización del VEGF en placentas de cabras sometidas a restricción alimentaria en el perÃodo prepuberal.
Materiales y métodos
Se utilizaron 19 cabras prepúberes de la raza Anglo nubian con un peso promedio de 12 kg, y aproximadamente 4 meses de edad. Los animales fueron asignados en tres grupos: control (C), (n=5) animales con consumo de materia seca a voluntad; restringido (R), (n=7) animales sometidos a una restricción del 30% del consumo potencial logrado en las hembras C y restringido con monensina sódica al 20% (laboratorio Elanco®) (M), (n=7) animales sometidos a restricción con la adición de 12,5 mg. de monensina/animal/dÃa. La dieta estuvo compuesta por una mezcla de 70:30 de heno de alfalfa picado y grano de maÃz molido, logrando una concentración energética de 2.4 Mcal/ KgMS. A los grupos M y R se los suplementó con urea para lograr una restricción solo energética. El alimento se ofreció en jaulas individuales durante 8 horas diarias (8 a.m a 4 p.m) a fin de controlar el consumo diario. El resto del tiempo los animales permanecieron libres con la finalidad que pudieran establecer interacción social y comportamiento reproductivo. Durante este tiempo tuvieron libre acceso a la provisión de agua. El aporte de minerales fue igual para todos los grupos a través de una suplementación con un núcleo vitamÃnico-mineral comercial.
El experimento se desarrolló durante 592 dÃas. El perÃodo de restricción alimentaria se prolongó por 250 dÃas, a partir de este periodo y coincidiendo con el comienzo de la etapa reproductiva (aproximadamente 12 meses de edad) los grupos R y M fueron alimentados igual que el grupo C hasta el final de la experiencia. Se realizó la detección de celo una vez por dÃa, por la mañana, antes de la entrega de alimento, por observación directa durante 45 minutos, utilizando un macho adulto provisto de delantal. Luego de comprobar la ciclicidad de todas las hembras, se realizó el servicio por monta natural. A los 25â30 dÃas posteriores al servicio se realizó el diagnóstico de la gestación utilizando un ecógrafo Berger®. Después del parto se recolectaron 19 placentas provenientes de los diferentes grupos de estudio establecidos y se registró el peso de las mismas. El protocolo experimental fue evaluado y aprobado por el Comité de Ãtica de la UNRC. Las muestras de tejido placentario se fijaron en formol al 10% tamponado y se procesaron con la técnica histológica de rutina. Algunos cortes se colorearon con hematoxilina-eosina y otros se usaron para inmunohistoquÃmica usando un anticuerpo primario anti-VEGF (1/100) (Santa Cruz, Biotechnology®), un anticuerpo secundario biotilinado, el complejo Avidina- Biotina-Peroxidasa (ABC) y se revelaron con diaminobenzidina (DAB) (Kit SK-41 Vector Lab. USA). En todos los casos se realizó un control negativo omitiendo el anticuerpo primario.
Análisis de los vasos sanguÃneos
En tres cortes de cada placenta se midieron, en 5 campos tomados al azar, el área y el perÃmetro de los vasos y se los clasificaron de acuerdo a su calibre utilizando el software Axiovision Release 4.6.3 (Carl Zeiss®). Para el estudio del área de los vasos sanguÃneos se dividieron los datos en tres intervalos de acuerdo al tamaño de los mismos: 1- vasos de pequeño calibre (10 a 100 μm2); 2- vasos de mediano calibre (100 a 400 μm2) y 3- vasos de gran calibre (400 a 1000 μm2).
Análisis inmunohistoquÃmico
Se contaron las células inmunomarcadas con el VEGF de 5 campos ópticos tomados al azar con un aumento de 400X en tres cortes de cada una de las 19 placentas y se obtuvo un Ãndice de VEGF calculado como el cociente entre las células inmunomarcadas con el VEGF y células totales por 100.
Análisis EstadÃstico
Los datos fueron analizados mediante análisis de Varianza y las comparaciones de medias se realizaron con la diferencia media mÃnima (LSD) de Fisher (alfa=0.05) bajo el modelo extendido: Yij = u + Ti +eij, siendo Yij la observación de la cabra j bajo el tratamiento i, u= la media total, Ti= el efecto del tratamiento i. Los análisis fueron realizados mediante la versión 2009 del Software InfoStat4 que permite el ajuste de modelos mixtos.
El peso promedio de las placentas fue de 457 ± 140,6 g (C: 539,4 ± 143,13; R: 417,9 ± 100,50; SD: 138,855) y no se encontraron diferencias significativas entre grupos.
La observación macroscópica de las placentas obtenidas luego del parto no demostró anomalÃas en ninguno de los grupos en estudio.
Análisis morfométrico de los vasos sanguÃneos
La mayor parte de los vasos de mediano y gran calibre se ubicaron en el tejido conectivo de las vellosidades corioalantoideas, siendo su distribución muy variable. Los vasos de pequeño calibre, en especial los capilares, se observaron cercanos al epitelio trofoblástico (Figura 1).

Figura 1. Corte histológico de placenta de cabra del grupo control con coloración de Hematoxilina/Eosina (200x).
Medición del área y perÃmetro de los vasos sanguÃneos.
El perÃmetro de los vasos sanguÃneos fue mayor significativamente (pâ¤0,01) en las placentas de cabras del grupo R y M, con respecto a las controles (Gráfico 1).
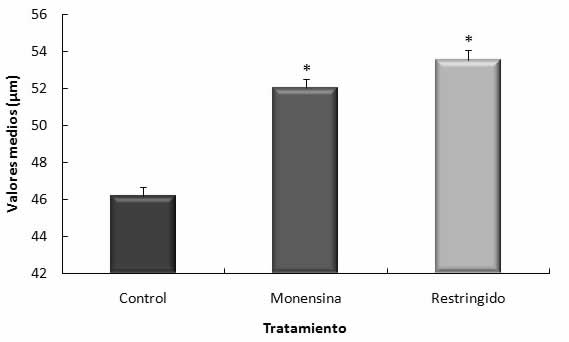
Gráfico 1 PerÃmetros de vasos sanguÃneos en el tejido placentario
El área de los vasos, dividida en tres intervalos de acuerdo al calibre, no mostró diferencias significativas en ninguno de ellos, entre los grupos R, M y C (Gráfico 2).

Gráfico 2. Ãrea de los vasos sanguÃneos en el tejido placentario
Análisis inmunohistoquÃmico de la expresión del factor de crecimiento endotelial vascular (VEGF) en placentas
Se determinó la presencia del VEGF en el epitelio trofoblástico de placentas C, R y M. El mismo se localizó en el citoplasma tanto de las células mononucleadas como de las binucleadas, observándose en estas últimas una intensa inmunomarcación. En las placentas de cabras R y M, el VEGF también fue identificado en el endotelio de los vasos sanguÃneos (Figura 2).

Figura 2. FotografÃa de corte histológico de placenta de cabra de grupo control inmunomarcadas con VEGF (400x).
ET: epitelio trofoblástico; CM: células mononucleadas; CB: células binucleadas.
Los resultados obtenidos demostraron que hubo diferencias estadÃsticamente significativas entre los 3 grupos en estudio, siendo mayor el Ãndice del VEGF en las placentas R (73.58 ± 3.65), con respecto a C (42.51 ± 3.04) y M (32.40 ± 3.98) (p â¹ 0.001) (Gráfico 3).

Gráfico 3. Ãndice de células inmunomarcadas con VEGF.
Discusión
Actualmente no hay evidencias del efecto que ejerce la restricción alimentaria realizada en hembras prepúberes, sobre el desarrollo placentario en cabras. Este estudio demostró que no hubo diferencias significativas entre los pesos de las placentas de las cabras sometidas a restricción alimentaria con respecto a sus controles. Las placentas de cabras restringidas y suplementadas con monensina, tampoco presentaron diferencias significativas con los controles, lo que indicarÃa que el uso del ionóforo no incide en el peso placentario. Se conoce que para lograr el mantenimiento de la preñez es fundamental el intenso crecimiento y formación de nuevos vasos sanguÃneos, asociados con el desarrollo de una eficiente interfase maternofetal12. El estudio de medidas morfométricas de los vasos sanguÃneos en las placentas de cabras de cada uno de los grupos establecidos y la expresión del VEGF, permitió evidenciar el grado de desarrollo de la vasculatura en placentas a término de cabras, presentando diferencias estadÃsticamente significativas entre los grupos estudiados, en concordancia con lo observado en placentas de ovinos por Vonnahme (2006).
El perÃmetro de los vasos sanguÃneos placentarios fue significativamente mayor en las placentas de cabras de los grupos con dieta restringida con respecto a sus controles. Mientras que, el área de los vasos no presentó diferencias significativas entre los grupos y para ninguno de los tres intervalos de tamaños establecidos. En este estudio el área de los vasos sanguÃneos no presentó cambios en las placentas de los grupos restringidos, por lo que se podrÃa inferir que no se afectarÃa la angiogénesis. El perÃmetro de los vasos sanguÃneos presentó un aumento significativo en las placentas de los grupos restringidos, lo que podrÃa indicar la presencia de un mecanismo compensatorio que mantendrÃa el área vascular a expensas de un incremento del tamaño de los vasos y no, mediante la formación de nuevos vasos sanguÃneos. Reynolds et al. (2004) demostraron que es más eficiente el intercambio placentario materno-fetal, cuando hay mayor cantidad de vasos sanguÃneos de pequeño calibre que escasos vasos de mayor tamaño.
En placentas de ovejas normales la arquitectura microvascular de la placenta fetal es altamente ramificada y compuesta de abundantes capilares pequeños que aseguran un transporte sanguÃneo rápido para mejorar el intercambio feto-materno10-11. En este estudio, el incremento del perÃmetro de los vasos en las placentas de los grupos restringidos, indicarÃa que la velocidad de transporte serÃa más baja, pudiendo afectar el adecuado intercambio materno-fetal.
En las placentas del grupo con dieta restringida, el Ãndice del VEGF fue mayor con respecto a las controles, coincidiendo con el mayor perÃmetro de sus vasos. De este modo, el VEGF estarÃa estimulando el crecimiento del endotelio vascular, propiedad demostrada por Choi et al., (2005) en las placentas de ratas. Sin duda, otros factores de crecimiento contribuyen en la formación y desarrollo de los vasos sanguÃneos, tal lo demostrado por Regnault et al. (2003) y por Vonnahme et al. (2001), siendo la angiogénesis el resultado de la influencia de numerosos factores.
Se puede concluir que las diferencias encontradas entre los grupos estudiados, indican que hubo una adaptación compensatoria en las cabras que fueron sometidas a una dieta restringida, ya que el área vascular se mantuvo sin modificaciones a expensas del aumento del perÃmetro de los vasos.
1. Barinaga, M. Designing therapies that target tumor blood vessels. Cancer Res. 1997; 275: 482 - 484. [ Links ]
2. Cufré, G.; Chaves, M.; Godio, L.; Vasquez, M.; Forchetti, O.; Martinez, R. Efecto de la restricción en el consumo de energÃa sobre parámetros sanguÃneos y pubertad en cabrillonas. Rev Arg Prod Anim. 2007; 27(3): 169 - 177. [ Links ]
3. De la Rosa Carbajal, S. Manual de producción caprina. 1a ed. Formosa. 2011; 1 - 4. ISBN 978-987-33-0421-7. [ Links ]
4. Di Rienzo J.; Casanoves, F.; Balzarini, M.; Gonzalez, L.; Tablada, M.; Robledo, C. Grupo InfoStat, 2009; FCA, Universidad Nacional de Córdoba, Argentina [ Links ]
5. Di Rienzo J.; Casanoves, F.; Balzarini, M.; Gonzalez, L.; Tablada, M.; Robledo, C. Grupo InfoStat, 2009; FCA, Universidad Nacional de Córdoba, Argentina Biblioteca Universidad Nacional de RÃo Cuarto. 2012; 118 p. [ Links ]
6. Garrett, J. Influence of prepartum and postpartum supplementation of a yeast culture and monensin, or both, on ruminal fermentation and performance of multiparous dairy cows. Anim Feed Sci Technol. 2005; 12: 129 - 239. [ Links ]
7. Fraser, S.; Ogawa, M.; Yu, R.; Nishikawa, S.; Yoder, M.; Nishikawa, S. Definitive hematopoietic commitment within the embryonic vascular endothelial-cadherin population. Exp Hematol. 2002; 30: 1070 - 1078. [ Links ]
8. Hafez, S.; Borowicz, P.; Reynolds, P.; Redmer, D. Maternal and fetal microvasculature in sheep placenta at several stages of gestation. J Anat. 2010; 216: 292 â 300. [ Links ]
9. Kelly, R. Nutrition and placental development. Proc Nutr Soc Aust. 1992; 17: 203 -211. [ Links ]
10. Regnault, T.; Vrijer, B.; Galan, H.; Davidsen, M.; Trembler, K.; Battaglia, F.; Wilkening, R.; Russell, V. The relationship between transplacental O2 diffusion and placental expression of PlGF, VEGF and their receptors in a placental insufficiency model of fetal growth restriction. J Physiol. 2003; 550: 641 - 656. [ Links ]
11. Reynolds, L.; Redmer, D. Growth and microvascular development of the uterus during early pregnancy in ewes. Biol Reprod. 1992; 47: 698 - 708. [ Links ]
12. Reynolds, L.; Borowicz, P.; Vonnahme, K.; Johnson, M.; Grazul-Bilska, A.; Wallace, J.; Caton, J.; Redmer, D. Animal Models of Placental Angiogenesis. Placenta. 2004; 26: 689 - 708. [ Links ]
13. Rivas, P.; RodrÃguez-Márquez, J.; Hernández, A. Número de vasos sanguÃneos y expresión del factor de crecimiento del endotelio vascular y de la óxido nÃtrico sintetasa endotelial e inducida en la alantoides ovina. Rev Cien FCV-LUZ. 2006; 16(4): 347 - 352. [ Links ]
14. Rivera, G.; Alanis, G.; Chaves, M.; Ferrero, S.; Morillo, H. Seasonality of estrus and ovululation in Creole goats of Argentina. Small Ruminant Res. 2003; 48: 109 â 117 [ Links ]
15. Spencer, T.; Bazer, F. Uterine and placental factors regulating conceptus growth in domestic animals. J Anim Sci. 2004; 82: 4 - 13. [ Links ]
16. Turiello, P. Incidencia del nivel nutricional sobre el estado metabólico y hormonal de cabrillonas y su repercusión sobre la eficiencia reproductiva. Tesis de doctorado. [ Links ]
17. Van Rens, B.; Koning, G.; Bergsma, R.; Lende, T. Preweaning piglet mortality in relation to placental efficiency. J Anim Sci. 2005; 83: 144 - 151. [ Links ]
18. Vonnahme, K.; Ford, S. Placental vascular endothelial growth factor receptor system mRNA expression in pigs selected for placental efficiency. J Physiol. 2004; 554: 194 - 201. [ Links ]
19. Vonnahme, K.; Hess, B.; Nijland, M.; Nathanielsz, P.; Ford, S. Placentomal differentiation may compensate for maternal nutrient restriction in ewes adapted to harsh range conditions. J Anim Sci. 2006; 84: 3451 - 3459. [ Links ]

