










Services on Demand
Journal

Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkInVet
On-line version ISSN 1668-3498
InVet vol.20 no.1 Ciudad Autónoma de Buenos Aires 2018
ARTÃCULO DE INVESTIGACIÃN
Estudio anatómico y funcional del aparato masticatorio del antílope negro, (Antilope Cervicapra, Artyodactyla, Bovidae)
Lozano, D.A.1; Blanco, C.J.1
1Universidad de Buenos Aires, Facultad de Ciencias Veterinarias, Cátedra de Anatomía. Chorroarín 280. CABA. Argentina
Recibido: 11/04/2018
Aceptado: 03/12/2018
Correspondencia e-mail: Diego Lozano diegoalozano@gmail.com
Resumen
El aparato digestivo de la Familia Bovidae está especialmente adaptado a la digestión fermentativa de la fibra vegetal, la bibliografía en base a las características etológicas y morfológicas del sistema digestivo establece tres grupos: ramoneadores (browsers), forrajeadores de pastos (grazers) y tipos intermedios. El presente trabajo se propone el estudio del aparato masticatorio del Antílope negro (A. cervicapra) centrado en sus características macroscópicas. Se estudió en dos cabezas de la especie la anatomía macroscópica de los músculos de la masticación, la articulación témporo-mandibular y las glándulas salivales. Especial atención se prestó a la morfología de la mandíbula y los dientes. Se determinaron las características morfométricas y macroscópicas de estas estructuras. Concluimos que en A. cervicapra las características anatómicas de su aparato masticador pueden ubicarlo como una especie intermedia.
Palabras clave: Antílope cervicapra; Ramoneadores; Forrajeadores; Anatomía veterinaria; Aparato masticador.
Anatomical and functional study of the masticatory apparatus of the black antelope, (Antilope Cervicapra, Artyodactyla, Bovidae)
Summary
The digestive system of Bovidae Family is specially adapted to the fermentative digestion of vegetable fiber; the bibliography based on ethological and morphological characteristics of digestive system establishes three groups: browsers, grazers and intermediate types. The present work proposes the study of the masticatory apparatus of the Blackbuck antelope (A. cervicapra) focused on its macroscopic characteristics. The macroscopic anatomy of the masticatory muscles, the temporo-mandibular joint and the salivary glands was studied in two heads of the species. Special attention was paid to the morphology of the jaw and teeth. The morphometric and macroscopic characteristics of these structures were determined. We conclude that in A. cervicapra the anatomical characteristics of its chewing apparatus can locate it as an intermediate species.
Key words: Antílope cervicapra; Browsers; Grazzers; Veterinary anatomy; Masticatory apparatus.
Introducción
El antílope negro (Antilope cervicapra) es uno de los tantos mamíferos exóticos introducidos en el país en el último siglo. Se encuentra distribuido en varias provincias del territorio nacional, utilizándose principalmente para la producción de carne y para caza deportiva. Se trata de una especie originaria de Asia, más puntualmente del territorio de la India, Pakistán, Nepal y Bangladesh. Su hábitat natural ha disminuido considerablemente durante el siglo XX, básicamente reduciéndose a reservas naturales e incluso extinguiéndose en los tres últimos países nombrados. Fueron introducidos con éxito en EE.UU. (especialmente el estado de Texas) y Argentina (principalmente en las regiones centro y norte) (8).
Es un rumiante diurno. Presenta dimorfismo sexual marcado: Los machos tienen manto pardo oscuro/negro en la zona dorsal del torso, laterales de las manos y los pies y la cara, mientras que la hembra presenta un pelaje marrón claro-amarillento. Los machos pesan entre 20 y 57 kg, y las hembras entre 20 y 33 kg. Son animales bicornes, de cuernos rectos y anillados que miden entre 35 y 75 cm, generalmente sólo presentes en los machos. Conforman grupos pequeños, con un macho dominante, hembras que pueden incluir a uno o dos machos periféricos. Habitan planicies semidesérticas y matorrales, incluyendo llanuras costeras pantanosas y bosques abiertos, aunque vive principalmente en zonas de pastizales templados y relativamente áridos. Tienen, sin embargo, un alto requerimiento de agua, por lo que suelen preferir estar en zonas con fuentes perennes de agua. Tienden al sedentarismo, aunque (generalmente en verano) pueden movilizarse grandes distancias en busca de agua (14). Su alimentación se constituye principalmente por pastos y vegetales con alto valor de fibra (9).
Los rumiantes han sido caracterizados acorde a sus hábitos alimentarios y el desarrollo adaptativo del sistema digestivo, dado por las preferencias y disponibilidad de las distintas especies forrajeras al momento de la alimentación. Hofmann y Stewart establecieron una clasificación al respecto dividida en tres categorías (-browser- -o selectores blandos, -grazer- - de alimentación dura y pastos, e -Intermediate-, o intermedios, especies que adaptan su dieta según la disponibilidad de alimento por temporadas) (6). Los rumiantes de estos tres grupos se diferencian evolutiva y adaptativamente por distintas características de las diferentes porciones del sistema digestivo. Sin embargo, los autores advierten que estas diferenciaciones en la alimentación y morfología no son estrictas y que las distintas especies adaptan su dieta según disponibilidad, pudiendo ser esta una razón de las fallas al ingresar animales a nuevos ecosistemas.
El presente trabajo se propone el estudio del aparato masticatorio del antílope negro, centrado en la estructura de la cavidad bucal y la dentadura, las glándulas salivales y los músculos de la masticación. El objetivo es entender las adaptaciones de esta especie profundizando en el conocimiento de la misma. De esta manera se espera brindar información a los productores nacionales para mejorar los hábitats en los que estos animales se encuentran, permitiendo alcanzar estándares aceptables de bienestar animal.
Materiales y Métodos
El estudio fue realizado en dos cabezas de A. cervicapra adultos machos provenientes de un criadero ubicado en General Belgrano, Provincia de Buenos Aires, Argentina. La conservación durante el transporte y disección se realizó por congelación. Las piezas se donaron a la Cátedra de Anatomía de la Facultad de Ciencias Veterinarias de la Universidad de Buenos Aires ya congeladas y seccionadas a la altura de la articulación atlanto-occipital. Se trabajó congelando los ejemplares entre sesiones de disección para retrasar la descomposición, evitando la retracción de los tejidos frente a una posible fijación química. Se realizó una disección capa por capa de la musculatura, glándulas salivales, vasos y nervios. El aparato hioideo se seccionó y conservó por separado. La lengua y los cartílagos auriculares de uno de los ejemplares se conservaron para estudios posteriores.
Luego de la disección de componentes blandos se limpiaron las estructuras óseas y se realizaron placas radiográficas de la dentadura y cinco mediciones de la mandíbula para comprender mejor la biomecánica del aparato masticatorio. Las variables determinadas fueron -altura de la rama mandibular- (desde el ángulo mandibular hasta el extremo del proceso coronoides); -altura del proceso coronoides- (desde la cabeza mandibular hasta el extremo del mismo); -altura efectiva- (entre la cabeza mandibular y la superficie oclusal de los molares mandibulares); -ancho de la rama mandibular- (entre el foramen mandibular y el borde caudal de la rama; -largo total- (desde el ángulo mandibular hasta el extremo incisivo de la mandíbula); -largo efectivo- (desde el borde caudal al foramen mentoniano) (Fig. 1 y 2).
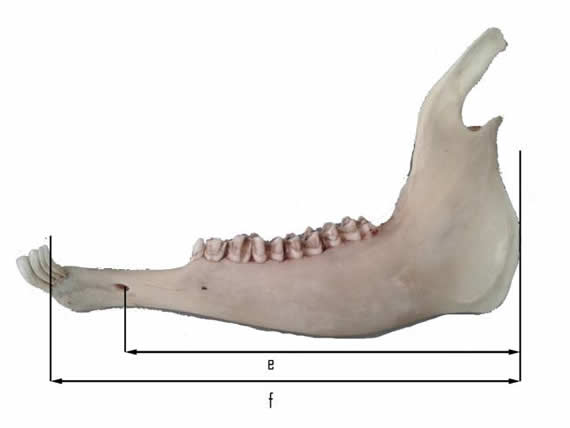
Figura 1. Vista lateral de la mandíbula izquierda con las mediciones realizadas: e) largo efectivo y f) largo total.

Figura 2. Vista medial de la mandíbula izquierda con mediciones: a) altura de la rama mandibular, b) altura del proceso coronoides, c) altura efectiva y d) ancho de la rama mandibular.
Los resultados obtenidos fueron comparados con la bibliografía existente (2) y material del Museo de Anatomía de la Facultad de Ciencias Veterinarias de la Universidad de Buenos Aires.
Los términos utilizados, se hallan traducidos del latín y se encuentran contenidos en la sexta edición revisada de la Nómina Anatómica Veterinaria (NAV) (7).
El análisis estadístico debido al pequeño tamaño de las muestras, las medias y su error fueron estimadas por bootstrap.
Resultados
Osteología
La -altura de la rama mandibular- fue de (9,66±0,37) cm, la -altura del proceso coronoides- (2,55±0,17) cm, y la -altura efectiva- (3,26±0,22) cm, el -ancho de la rama mandibular- (2,34±0,22) cm, el -largo total- (16,44±0,25) cm y el -largo efectivo- (13,81±0,34) cm. En todos los casos se indica la media ± error estándar estimado por bootstrap.
Mandíbula: (Fig. 1 y 2) La rama mandibular forma un ángulo recto con el cuerpo mandibular (más agudo que el de bovinos, ovinos y caprinos). El cuerpo es relativamente alargado y aplanado. Presenta un diastema relativamente largo. El borde ventral es menos convexo que el del bovino y caprino, con la convexidad hacia caudal (2). El ángulo mandibular es redondeado hacia caudal y presenta crestas rugosas para las inserciones musculares. Por medial, la cresta para el músculo pterigoideo lateral es pronunciada. La apófisis coronoides nace con un ángulo marcado y se extiende caudalmente en forma recta. Tiene un pequeño engrosamiento hacia la mitad del borde caudal. La apófisis condílea es angosta y aplanada, sus bordes ovalados.
La articulación intermandibular es una sincondrosis, si bien estudios histológicos futuros deben confirmar esta presunción.
Dentición. Esta especie es heterodonte y difiodonte. Ambas piezas estudiadas corresponden a machos adultos jóvenes (de aproximadamente 4-6 años), con dentición definitiva completa. La misma responde a la fórmula:(I 0/4;C 0/0;PM 3/(2(3)) ;M 3/3)*2.
Incisivos. Son braquiodontes, la corona se adelgaza en el eje transversal desde mesial a distal (Fig. 1). El largo de las piezas permanece constante. No hay incisivos superiores como ocurre en toda la familia Bovidae. El rodete dentario es comparativamente delgado. Se encuentran laxamente fijados al alvéolo. Las caras labiales son convexas y las linguales cóncavas. Se proyectan oblicuos en dirección dorso-rostral. Sólo el I1 tiene la característica forma de pala.
De I2 a I4
son más angostos, de superficie decreciente y progresivamente más curvos hacia distal. El cuello es marcado sólo en los primeros dos incisivos. De I1 a I4 aumentan gradualmente su curvatura (convexa hacia distal y la cara vestibular).
Premolares. Son hipsodontes y ocupan la mitad de espacio que los molares (Fig. 1, 2, 3 y 5). P1 y P1 están ausentes (característico de la familia Bovidae) (4). P2 también puede estar ausente. Presentan dos raíces y una corona. Crecen y se complejizan hacia distal. Las coronas de los premolares inferiores se tornan alargadas y más complejas en sus plegamientos en la misma dirección. Los superiores tienen cara lingual convexa y una bucal en forma de -w-. Poseen invaginaciones de esmalte y cemento en forma de medialuna (selenodontes) que también se complejizan y completan hacia distal.
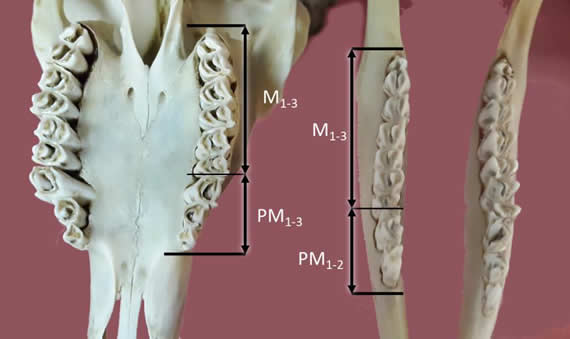
Figura 3. Vista de la cara oclusal de molares y premolares: Se aprecia la dentición selenodonte y el menor espesor linguo-bucal de la superficie de las muelas inferiores.

Figura 4. Vista lateral de la disección media del aparato masticador. Se seccionaron el nervio facial, arteria facial transversa, glándula parótida y conducto parotídeo, músculos malar y de la mímica y glándula periorbitaria. Se observan en: a) M. masetero superficial, b) M. Masetero profundo, c) M. temporal, d) M. buccinador y e) glandulas bucales.

Figura 5. Vista lateral izquierda, disección profunda con alvéolos expuestos. Se observa: b) M. masetero profundo, c) M. temporal, e) Arco cigomático, f y f-) raíces de los molares y premolares superiores, g y g-) raíces de los molares y premolares inferiores.
Emergen perpendiculares a la superficie alveolar. Las raíces de los superiores son tres, pero las dos rostrales se encuentran fusionadas. Tanto los premolares como los molares superiores tienen mayor superficie oclusal y sobresalen por lateral de la arcada inferior, con crestas laterales prominentes. Los molares y premolares (especialmente los inferiores) son delgados, se posicionan formando una hilera de cúspides a lo largo de toda la dentadura.
Molares. Son sensiblemente mayores que los premolares (Fig. 1, 2, 3 y 5). Sus raíces (generalmente tres con dos fusionadas) son voluminosas y se extienden más en profundidad en los respectivos alvéolos. Tienen dos o tres cúspides (paracono, protocono y metacono) posicionadas linealmente de distal a mesial respectivamente. Presentan crestas medial y lateral. Ãstas últimas sobresalen del borde lateral de los molares inferiores, proyectándose por lateral a éstos. Emergen de los alveolos oblicuos en dirección dorso-rostral, a diferencia de los premolares.
Sindesmología
Articulación temporomandibular: La superficie articular del hueso temporal es amplia, alargada cráneo-caudalmente y aplanada. En conjunto con el disco articular y la forma del cóndilo de la mandíbula muestran capacidad de circunducción y protracción-retracción marcada. La distancia (altura) entre la articulación y la dentadura es relativamente baja (Fig. 2 y 3). El disco articular está bien desarrollado. No se observaron ligamentos articulares más que el ligamento capsular.
Miología
Músculo masetero (Fig. 4 y 5): Es relativamente plano en comparación con bovinos. Las fibras de la porción superficial se dirigen desde la totalidad del ángulo de la mandíbula en dirección oblicua dorso-rostral. Está cubierto por una lámina gruesa de fascia, en la que también se inserta (en rostral) un tendón del esterno-mandibular (Esterno-cigomático). Las fibras de la porción profunda discurren con dirección más recta, dorso-ventral, desde la cresta facial y arco cigomático, hacia una leve escotadura en la superficie lateral de la rama de la mandíbula y el borde dorsal de la porción caudal del cuerpo. Sólo se ven las fibras de la capa profunda en el extremo dorso-caudal del músculo. Hacia rostral no son claramente divisibles los límites entre la porción profunda y la superficial, ya que se encuentran fusionadas.
Relaciones: hacia rostral y superficial discurren la arteria y vena faciales y el conducto parotídeo. La glándula parotídea y arteria y vena maxilares limitan al músculo por caudal. A la altura de la terminación de la porción superficial se encuentra el linfonódulo parotídeo. También sobre su cara lateral se observan dorsalmente el tendón de origen del músculo cigomático y más ventralmente el nervio facial y la arteria y vena faciales transversas.
Músculo temporal (Fig. 4 y 5): El músculo ocupa la totalidad de la fosa temporal (de forma relativamente circular), por superficial de la cápsula articular mandibular. Las fibras se distribuyen rostro-caudalmente, abriéndose en forma de abanico hacia caudal. Tiene una aponeurosis en la que se unen las fibras dorsales y ventrales a la apófisis coronoides de la mandíbula. La forma (recta y alargada), y dirección en la que se desarrolla esta apófisis, en coincidencia con el espesor del músculo temporal muestran que éste tiene un importante rol en la masticación de esta especie.
Músculos pterigoideos. Fuertes de aspecto tendinoso, con concordancia con la forma de la articulación témporo-mandibular. Sus lugares de inserción mandibulares son amplios y están relativamente marcados (Fig. 6).

Figura 6. Vista medial de la mandíbula derecha. Se seccionó la mandíbula izquierda, músculos intrínsecos y extrínsecos de la lengua y aparato hioideo. Se observa: a) Cuerpo de la mandíbula derecha, b) Proceso ganchoso del hueso pterigoideo, c) Músculo pterigoideo medial, d) (recuadro): se seccionó el músculo pterigoideo medial para visualizar el músculo pterigoideo lateral.
Músculo pterigoideo lateral. Es plano y recto. Sus fibras discurren más paralelas y dorso-ventralmente que las del pterigoideo medial. Una delgada capa de fascia lo separa de este músculo. Su inserción a la mandíbula es mediante una aponeurosis, y tiene una escotadura de forma similar a la del ángulo mandibular.
Músculo pterigoideo medial. Es un músculo plano, sus fibras se dispersan en forma de abanico de rostro-dorsal a caudo-ventral. Está cubierto por una fascia en casi toda la superficie, desde el tendón de origen en la porción perpendicular del hueso palatino y proceso pterigoideo del basiesfenoides hasta el ángulo de la mandíbula, en el que se distribuye por toda su extensión.
Músculo digástrico. Se origina en la apófisis yugular del hueso occipital y se inserta en la superficie medial del cuerpo mandibular. El vientre rostral se inserta desde caudal al tercer molar, hacia rostral hasta el primer molar. Este vientre es relativamente ancho, mayor al caudal, el que se encuentra en relación con la glándula salivar mandibular.
Glándulas
Glándula parótida. En relación con el músculo masetero, su tamaño relativo es menor que en otras especies forrajeras de pastos, no se extiende sobre el músculo limitándose al borde caudal del músculo como ocurre en el ovino (1). La misma se extiende sobre el linfocentro parotídeo. Su conducto sigue el mismo trayecto que en el resto de los pequeños rumiantes (10).
Glándula Mandibular. Más pequeña en proporción que en Carnivora, se presenta como en los pequeños rumiantes, parcialmente cubierta por la parótida entre el ángulo mandibular y el ala del atlas. El aspecto es alargado y su conducto se extiende entre los músculos estilogloso y geniogloso hasta confundirse en parte con la glándula sublingual.
Glándula bucal. Se encuentra entre las fibras del músculo buccinador, malar y depresor del labio superior (fig.4). Se observa como pequeños nódulos dispersos sobre la cara lateral de los vientres musculares.
Glándula sublingual. Es delgada y delicada, se continúa rostralmente al músculo estilogloso y en el mismo plano. La porción monostomática cubre parcialmente al conducto parotídeo para desembocar con él en la carúncula sublingual como ocurre en oveja y cabra (1).
Conclusiones
Del posicionamiento de los molares y premolares (oblicuos en dirección opuesta y la arcada superior significativamente más ancha que la inferior) se evidencia un mecanismo masticatorio adaptado para realizar un doble trabajo de trituración de pasturas: Cierra la boca con la mandíbula protraccionada, y en una segunda acción la retrae. Esto se condice con la superficie articular del temporal alargada, el gran tamaño del músculo temporal y el ángulo de proyección de la apófisis coronoides (11). El posicionamiento de las fibras superficiales y profundas del masetero y pterigoideos (3), por su parte, dan muestra de la capacidad de circunducción.
Las diferencias de tamaño y grado de adhesión alveolar de los incisivos, el tamaño relativo pequeño de la glándula parótida y mandibular se condicen con los rumiantes de alimentación intermedia de acuerdo a los estudios de Shipley (13) y no coinciden con uno estrictamente forrajeador (5).
Sin embargo la delgadez del hocico y los molares son característicos de animales seleccionadores de forraje. El ángulo de los incisivos no se presentó marcadamente inclinado hacia rostral como ocurre en los forrajeadores. Estas características, acordando con la bibliografía (5) (13) , muestran un desarrollo evolutivo del sistema de prehensión y masticación de un animal de dieta intermedia. Coincidimos, entonces con Sauer y cols. (2016), en que si bien la especie se comporta etológicamente como un forrajeador, el aparato masticador presenta características intermedias (12).
1. Barone R. Anatomie Comparée des Mammifieres Domestiques. Tomo III Paris: Vigot; 2009. [ Links ]
2. Choudhary OP, Singh I, Kumar N. Morphological Studies on Mandible of Indian Blackbuck (Antelope cervicapra). Indian Veterinary Journal. 2016; 93(01): p. 79-81. [ Links ]
3. Fletcher TM, Janis CM, Rayfield EJ. Finite Element Analisys of ungulate jaws: can mode of digestive physiology be determined? Palaeontologia Electronica. 2010; 13(3): p. 21-15. [ Links ]
4. Getty R, Grossman J, Sisson S. Anatomía de los animales domésticos Barcelona, España: Masson Editores; 2000. [ Links ]
5. Hoffman RR, Axemacher H. Morphologycal characteristics of the maseter muscle of 22 ruminant species. 1988. [ Links ]
6. Hofmann RR, Stewart DRM. Grazer or browser: A classification based on the stomach structure and feeding habits of East African ruminants. Mammalia. 1972;(36): p. 226-240. [ Links ]
7. International Committee on Veterinary Gross Anatomical Nomenclature. Nomina Anatomica Veterinaria. 5th ed. World Association of Veterinary Anatomist, editor. Hannover; 2012. [ Links ]
8. IUCN. http://www.iucnredlist.org/details/1681/0. [Online].: IUCN; 2017 [cited 2017 November 11. Available from: http://dx.doi.org/10.2305/IUCN.UK.2017-2.RLTS.T1681A50181949.en. [ Links ]
9. Naik R, Solanki G. Grazing Interactions between wild and domestic herbibores. Elsevier Science B. V. 1998: p. Volumen 27, Issue 3, Pags 231-235. [ Links ]
10. Nickel R, Schummer A, Seiferle E. Lehrbuch der Anatomie der Haustiere Berlin: Verlag Paul Parey; 1979 [ Links ]
11. Pérez-Barberia FJ, Gordon IJ. The functional relationship betwen feeding type and jaw and cranial morphology in ungulates. Oecología. 1999; 118: p. 157-165. [ Links ]
12. Sauer C, Bertelsen MF, Hammer S, Lund P, Weisbjerg MR, Clauss M. Macroscopic digestive tract anatomy of two small antelopes, the blackbuck (Antilope cervicapra) and the Arabian sand gazelle (Gazella subgutturosa marica). Anatomia, Histologia, Embryologia. 2016 Oct; 45(5): p. 392-398. [ Links ]
13. Shipley L. Grazers and Browsers: How Digestive Morphology Affects Diet Selection. Grazing Behavior Of Livestock and Wildlife #70. 1999. [ Links ]
14. Vats R, Bhardwaj C. A Study of Reproductive Behaviour of Indian Black Buck (Antilope cervicapra) Linn. with Reference to Courtship, Breeding, Fawning and Colouration. Ambala, India; 2009. [ Links ]

