










Servicios Personalizados
Revista

Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO  uBio
uBio
Compartir

 Permalink
PermalinkInVet
versión On-line ISSN 1668-3498
InVet vol.20 no.2 Ciudad Autónoma de Buenos Aires jun. 2018
ARTÃCULO DE INVESTIGACIÃN
Comparación morfométrica de la columna vertebral del ocelote (Leopardus Pardalis), puma (Puma Concolor) y gato doméstico (Felis Catus)
Sánchez, HL1, 3*; Barbeito-Andrés, J2,5*; Nishida, F3,5; Barbeito, CG4,5; Portiansky, EL3,5
1Departamento de Anatomía. Facultad de Ciencias Veterinarias (FCV). Universidad Nacional de La Plata (UNLP), Argentina.
2Unidad Ejecutora de Estudios en Neurociencias y Sistemas Complejos (ENyS). CONICET.
3Laboratorio de Análisis de Imágenes (LAI). Facultad de Ciencias Veterinarias (FCV). Universidad Nacional de La Plata (UNLP), Argentina.
4Laboratorio de Histología y Embriología Descriptiva, Experimental y Comparativa (LHYEDEC). Facultad de Ciencias Veterinarias (FCV). Universidad Nacional de La Plata (UNLP), Argentina.
5Consejo Nacional de Ciencia y Tecnología (CONICET). Argentina.
*Igual contribución.
Recibido: 07/09/2018
Aceptado: 15/03/2019
Correspondencia e-mail: Hilda Liliana Sánchez lilianasanchez@fcv.unlp.edu.ar
Resumen
El ocelote y el puma son felinos salvajes nativos de América, particularmente distribuidos en áreas montañosas y boscosas. El conocimiento de la estructura ósea de la columna vertebral es de gran importancia, ya que estas especies se caracterizan por su flexibilidad, lo que les da la agilidad para trepar y saltar con gran ligereza, y moverse rápidamente mientras capturan a su presa. Nuestro objetivo fue realizar un estudio morfométrico comparativo entre la columna vertebral del ocelote, del puma y del gato doméstico. Los estudios morfométricos basados en distancias lineales se realizaron tanto sobre las piezas anatómicas aisladas como sobre su representación digital, en tanto que los análisis de morfometría geométrica se realizaron sobre estas últimas. En este estudio se encontraron similitudes anatómicas entre el puma y el gato, que pueden relacionarse con su asociación filogenética.
Palabras clave: Felinos salvajes; Felinos americanos; Locomoción; Raquis.
Comparative morphometry of the vertebral column of ocelot (Leopardus Pardalis), puma (Puma Concolor) and domestic cat (Felis Catus)
Summary
Ocelot and puma are native American wild cats especially distributed in mountainous and forested areas. Knowledge of the bone structure of the spine is of great importance since these species are characterized by their flexibility, which gives them the agility to climb and jump with great lightness and move quickly while capturing the prey. Our goal was to perform a comparative morphometric study between the spine of ocelot, puma and domestic cat (Felis catus). Morphometric studies (linear distances) were carried out both on the isolated anatomical pieces and on their digital representation, while the geometric morphometric analysis was performed on the latter. In this study, anatomical similarities between puma and cat that may be related to their major phylogenetic relationship were found.
Key words: Wild cats; American felines; Locomotion; Spine.
Introducción
El ocelote (Leopardus pardalis) y el puma (Puma concolor) son felinos nativos de América que se distribuyen, particularmente, en zonas montañosas y boscosas. El ocelote, también conocido como el leopardo enano, es un felino neotropical de tamaño mediano, que alcanza una longitud total del cuerpo de 70 a 90 cm, con una cola fina de 27 a 45 cm de largo, y un peso de 7 a 16 kg. Es un depredador oportunista que consume pequeñas presas (usualmente inferiores a 1 kg de peso). Se distribuye desde el sur de Estados Unidos hasta el norte de Argentina5,6.
Por su parte, el puma también conocido como león de montaña, es un felino grande, con una longitud corporal de 2 m y una cola de 66-78 cm de largo. El peso de esta especie oscila entre 22-80 kg 18. Se distribuye desde el norte de Canadá al sur de los Andes. Este gran depredador permanece al acecho entre los arbustos, rocas y árboles durante un tiempo corto y caza mediante emboscadas. Particularmente, las presas de estos animales consisten en mamíferos de tamaño pequeño a mediano 3.
La evolución ha modelado a estos felinos salvajes como cazadores y depredadores. El estudio de la locomoción es esencial para comprender la adaptación y subsistencia de estos animales. Los pumas son grandes depredadores, con miembros pelvianos musculosos, más largos y más fuertes que los miembros torácicos. Estas características les permiten saltar y desarrollar velocidades de hasta 65-80 km/h durante la persecución de la presa. Parte de esta función es ejercida por los músculos del antebrazo y la mano, cuyos datos anatómicos para el puma y otros felinos fueron descriptos previamente 26. Por su parte, los ocelotes presentan dos estrategias de caza: perseguir a su presa muy lenta y sigilosamente (0,3 km/h) o simplemente sentarse a esperar, para luego desplazarse a otra área a una velocidad de 0,8-1,4 km/h y recomenzar el acecho 27.
El conocimiento de la estructura ósea de la columna vertebral de los felinos es de gran importancia, ya que estas especies se caracterizan por su flexibilidad y agilidad para escalar y saltar 1,17. Estos felinos utilizan la cola como contrapeso del cuerpo durante los movimientos rápidos y la escalada. Al enderezarla, estos animales logran el equilibrio necesario que les garantiza un aterrizaje seguro 25. El rango de masa corporal ha inspirado discusiones centradas en la postura, la captura de las presas y la capacidad locomotora 4,17.
El gato doméstico presenta algunas similitudes con estos felinos salvajes, ya que son cazadores naturales e instintivos. Sus preferencias están relacionadas con la caza de pequeños roedores, aves o insectos 15.
Anatómicamente, la columna vertebral de los gatos se compone de siete vértebras cervicales, trece torácicas, siete lumbares, tres sacras y veinte a veintitrés caudales. Entre los cuerpos vertebrales, se insertan discos muy elásticos para amortiguar el salto 8,24.
Dadas las pocas referencias anatómicas sobre la columna vertebral de los felinos sudamericanos, nuestro objetivo fue realizar un estudio morfométrico comparativo del raquis del ocelote y del puma, dos felinos arbóreos, de masas corporales distintas. Incluimos en nuestros análisis al gato doméstico (Felis catus) como referencia comparativa con las especies silvestres. En conjunto, nuestros resultados brindarán algunas bases morfológicas para posibles estudios funcionales que interpretan los patrones de locomoción de diferentes representantes de la familia Felidae.
Materiales y Métodos
Piezas anatómicas
Se utilizaron dos gatos domésticos hembras adultas, provenientes del servicio de necropsias de la Facultad de Ciencias Veterinarias de la Universidad Nacional de La Plata. El zoológico de la Municipalidad de La Plata contribuyó con dos pumas adultos, un macho y una hembra, y dos ocelotes hembra, una adulta y otra púber, todos ellos muertos naturalmente en diferentes momentos. El peso de los pumas varió entre 40 y 45 kg mientras que el de los ocelotes fue de 10 kg para el adulto y 7 kg para la hembra púber.
Remoción de la columna vertebral
La columna vertebral de cada uno de los animales fue extraída, descarnada, hervida, macerada en agua y posteriormente sumergida en un baño de hipoclorito de sodio al 5 %, durante 6 horas, para eliminar todos los restos correspondientes a músculos y articulaciones. Las vértebras separadas fueron posteriormente reensambladas para su exhibición (Fig. 1A-C). Aunque se midió la totalidad de las vértebras de todos los esqueletos, solo se consideraron piezas representativas de cada región. Para este estudio, se compararon todas las vértebras cervicales (C1-C7), primera torácica (T1), primera lumbar (L1) y el hueso sacro.

Fig. 1. Columna vertebral de los felinos y vértebras aisladas. A. Ocelote (A1: vista lateral y A2: dorsal); B. Puma; C. Gato doméstico. Las vértebras escaneadas del ocelote (D), el puma (E) y el gato doméstico (F) se usaron para describir, medir y comparar todos los patrones morfométricos. Barra = 20 mm.
Morfometría lineal
El espesor del cuerpo vertebral fue registrado en todas las piezas anatómicas por medio de un calibre convencional.
Cada vértebra fue escaneada a 600 ppp, mediante un escáner plano (Genius HR6X, EE. UU.). Cada pieza anatómica se colocó sobre la base del escáner, de manera tal que toda la luz de su canal central se pudiera observar en la imagen final sin la generación de sombras (perpendicular a la base del escáner) (Fig. 1D-F). De esta manera, el primer plano de las imágenes contendría la región craneal de la vértebra, dejando en un segundo plano a la región caudal, junto con los detalles de los recesos laterales (apófisis transversas). A nuestro entender, a excepción del trabajo descripto por Nishida et al. 19, no se utiliza el escáner plano, de manera regular para este tipo de estudios, ya que, por lo general, el estudio de las vértebras se realiza mediante un escáner tridimensional 20. Las imágenes escaneadas (bidimensionales) se analizaron utilizando un analizador digital de imágenes (ImagePro Plus v6.3 - Media Cybernetics, EE. UU.). Para fines morfométricos, las imágenes fueron divididas en tres regiones: arco, cuerpo y canal vertebral, en las que se determinaron la altura y el ancho (Fig. 2A). Para determinar las características morfométricas del canal vertebral, las imágenes fueron umbralizadas (transformación a imagen binaria - blanco y negro - a partir de un valor umbral de intensidad de píxel de la imagen color o monocromática) y posteriormente filtradas mediante algoritmos de esqueletonización (Thinning y Prunning), para establecer los puntos medios de las estructuras vertebrales completas y evaluar la variación de la masa ósea (Fig. 2B-D). La región central de la imagen esqueletonizada, correspondiente al canal vertebral, fue utilizada para calcular su área, altura, ancho, perímetro y redondez. Este último parámetro determina la similitud del objeto con respecto al círculo, según lo determinado por la siguiente fórmula: (perímetro2) / (área 4 * Ï *); los objetos circulares tendrán una redondez = 1, mientras que otras formas tendrán una redondez > 1. Todas las mediciones fueron realizadas de manera automatizada.

Fig. 2. Morfometría lineal de las vértebras. A. Dimensiones morfométricas utilizadas para describir cada vértebra. Fusión digital entre la vértebra C2 y su esquema de esqueletonización para el ocelote (B), puma (C) y gato doméstico (D). E: datos morfométricos basados en las dimensiones descritas en A. En todos los casos también se expresa la relación ancho:longitud. Particularmente, para el canal vertebral, se agregaron medidas de área, perímetro y redondez.
Morfometría Geométrica
El análisis se llevó a cabo en base al diseño previamente publicado19. Las coordenadas 2D de 2 puntos de referencia (landmarks) y 20 semi-puntos de referencia (semi-landmarks) se digitalizaron en el contorno interno y externo del canal vertebral de cada una de las vértebras, para describir la curva del canal vertebral (Fig. 3A, B). Luego de desplazar los semi-landmarks, se obtuvo una variable de tamaño (tamaño del centroide) y un conjunto de variables de forma (coordenadas superpuestas), utilizando el Análisis de Procrustes Generalizado (GPA) como técnica de superposición 23. Como resultado, se obtuvo una variable de tamaño global para cada vértebra (tamaño de centroide) y nuevas coordenadas de puntos ajustadas, que representan las variables de la forma. Este procedimiento se realizó para obtener los datos de forma que resultan luego de controlar el tamaño y los efectos de posición.

Fig. 3. Análisis de morfometría geométrica. Delimitación del canal externo (A) e interno (B) de una vértebra con puntos de referencia (landmarks) y semi-puntos de referencia (semilandmarks). Tamaño promedio del centroide del canal externo (C) e interno (D) de todas las vértebras de la especie analizada. E y F representan el centroide de cada vértebra.
Análisis estadísticos
Las mediciones correspondientes a la morfometría lineal fueron analizadas mediante el Análisis de Componentes Principales (ACP), como técnica estadística, que permite sintetizar las variables lineales (relación alto:ancho del arco, cuerpo y canal vertebral, y la redondez) y establecer las relaciones entre estas variables. De este modo, permite establecer la similitud entre especies. A su vez, se realizó un ACP sobre las coordenadas superpuestas para obtener ejes que retoman una gran cantidad de variación. Los análisis se realizaron usando el software NCSS (v07.1.20. www.NCSS.com) y R (www.r-project.org).
Las diferencias de tamaño de los centroides obtenidas mediante la morfometría geométrica se utilizaron para describir el canal central de cada vértebra de las tres especies de felinos analizadas. Para este propósito, la recopilación de datos y el análisis posterior se llevaron a cabo mediante el uso de los programas TPS series 13 y MorphoJ 22.
Dado el escaso tamaño muestral relativo a la procedencia de las muestras, este trabajo debe ser considerado como un estudio descriptivo que pretende establecer las relaciones que existen entre las variables cuantitativas de las vértebras, y no como un estudio analítico.
Resultados
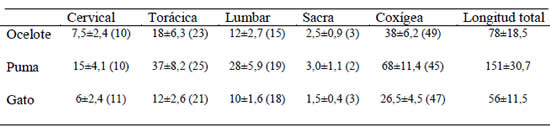
La medición de la longitud total de la columna vertebral reveló que el 10-11% correspondía a la región cervical, mientras que el 45-49% correspondía a la región caudal, en las tres especies estudiadas (Tabla 1). En términos generales, se observaron diferencias leves en el porcentaje de las regiones restantes. Así, la región torácica del gato tenía el menor porcentaje; la región lumbar más corta correspondía al ocelote, y el porcentaje mínimo de la región caudal le correspondió al puma.
Tabla 1. Longitud de los diversos sectores raquídeos. Las medidas se expresan en cm ± SEM. Entre paréntesis, se indica el porcentaje que representa el sector con respecto a la longitud total del raquis.
Morfometría lineal
El hueso sacro en el gato y el ocelote estaba formado por tres vértebras fusionadas formando un hueso compacto, mientras que en el puma estaba formado también por tres vértebras, pero poco fusionadas, dejando abierto hacia dorsal el canal vertebral (Fig. 1 A,B y C).
El Atlas (C1) es una vértebra atípica que carece de cuerpo y apófisis espinosa (aunque con fines morfométricos se consideró como tal). Está formada por un arco dorsal y otro ventral. Este último representa el cuerpo de la vértebra. En todas las especies estudiadas, el arco ventral (cuerpo) de C1 tenía un ancho mayor que su altura en comparación con el arco dorsal correspondiente (Fig. 2E). Además, el ancho de ambos arcos fue siempre mucho mayor que su altura, como se observa en la relación ancho:altura de las tres especies. De acuerdo con nuestras mediciones, el arco dorsal es más estrecho, pero más alto que su arco ventral correspondiente (Fig. 2E).
Cuerpo vertebral. La medición del ancho del cuerpo vertebral mostró valores relativamente constantes de C2 a C7, con la excepción de una leve reducción en C3 para el ocelote. Estos valores se mantuvieron con el mismo tenor para las vértebras T1 y L1 del ocelote y del gato, pero no así para las del puma, en el que a partir de T1, el ancho de las vértebras se incrementó, al menos un tercio más que en la región cervical. En todas las especies se detectó un incremento craneocaudal del tubérculo ventral. En general, el espesor del cuerpo de las vértebras del puma duplicó al de las otras dos especies (Tabla 2).
Tabla 2. Espesor craneocaudal del cuerpo vertebral. Las mediciones se tomaron mediante un calibre y se expresan en cm. Los valores se expresan como media ± SEM. * Corresponde al espesor craneocaudal del arco ventral; & Incluye el receso odontoideo; # Espesor craneocaudal total del cuerpo sacro.

La altura del cuerpo en el ocelote era mayor, incluso que la del puma, en las vértebras C4, C5 y C6, pero se reducía a la mitad con respecto a esa especie desde C7 a S1. En T1, los cuerpos vertebrales del ocelote y el gato eran cilíndricos y cortos. La relación ancho:altura fue casi similar en todas las especies. En el gato y el puma se observaron las mayores similitudes en C3, C4, C5, C6 y T1. El cuerpo vertebral lumbar del puma era relativamente largo y aplanado dorso-ventralmente. El ancho del cuerpo del sacro del puma y del gato era relativamente más grande que el del ocelote.
Arco vertebral. Los datos morfométricos correspondientes a los parámetros medidos en el arco de cada vértebra se pueden ver en la figura 2E. En el ocelote y el gato, el ancho del arco aumentaba su tamaño de C2 a T1, pero estos incrementos no eran pronunciados. Sin embargo, las diferencias más relevantes se observaron entre C2 y T1 en el ocelote y entre C2 y C3 en el gato. En el puma, el ancho del arco aumentaba de manera constante desde C2, alcanzando su máximo en C5 y disminuyendo hacia S1. En el gato y el puma, el arco aumentaba su altura desde C3 hacia T1. En el ocelote, aumentaba hacia C7 y luego disminuía.
En todas las especies, la relación ancho:altura del arco disminuía de C3 a T1, excepto para el ocelote, en el que dicha relación aumentaba en T1 a un valor similar a la de C5, pero luego disminuía hacia L1.
Los procesos articulares caudales de L1 a L7 del puma presentaban una muesca triangular, lo que inducía a una ligera, pero notable, diferencia en el ancho de los arcos vertebrales. En el puma, observamos un proceso accesorio robusto y prominente, orientado de manera horizontal, mientras que en el ocelote y el gato doméstico era muy delgado y pequeño (Fig. 1). Los procesos transversos orientados cráneo-ventralmente son muy desarrollados en el puma mientras que, los procesos espinosos mantienen una altura constante de craneal a caudal en los tres felinos estudiados (Fig. 1 y 2).
Canal vertebral. La relación ancho:altura del canal vertebral se mantuvo constante en todas las especies, aún en los sectores correspondientes a las expansiones de la médula espinal (intumescencia cervical y lumbar). Sin embargo, la relación ancho:altura del canal vertebral del gato y del ocelote era mayor que la del puma a lo largo de toda la columna vertebral. Por otro lado, esta relación era mayor en el cuerpo de las vértebras del gato, en comparación con las otras dos especies analizadas.
El área, el perímetro y la redondez del canal vertebral fueron constantes en todas las especies estudiadas. Las proporciones entre los rasgos vertebrales se analizaron mediante la reducción de la variación de la relación ancho:altura y redondez en los principales ejes de diferenciación.
Los segmentos vertebrales se pueden agrupar en un modelo de dos componentes (PC1 y PC2) que representa el 88,10% de toda la variación de datos (Fig. 4A-B). El Atlas de las tres especies analizadas se distribuyó por separado del resto de los segmentos. Por otro lado, el resto de las vértebras se distribuyeron centralmente, en las que las vértebras del ocelote y del
gato mostraron dos grupos definidos, excepto para S1 del gato. Las vértebras del puma, a diferencia de las del ocelote y del gato, estaban más dispersas a lo largo de PC2. Considerando la similitud de las formas entre las tres especies estudiadas, C2 se mostró ligeramente cercana entre el puma y gato, mientras que C3 estuvo más cercana entre el ocelote y el puma. Las vértebras de C4 a S1 mostraron una distancia equivalente entre las especies consideradas (Tabla 3).

Fig 4. Análisis de componentes principales (PCA). A. PCA en variables lineales de las vértebras (valores de la relación ancho: altura del arco, cuerpo y canal vertebral y de la redondez del canal), se muestra la comparación PC1 vs. PC2. B. PC1 (obtenido de las variables de forma) vs. el tamaño del centroide para el contorno del canal interno y externo. CS = tamaño de centroide. PCA en las coordenadas de la forma del canal externo (C) e interno (D). Se muestra la distribución de muestras y vértebras a lo largo de PC1 y PC2. Los contornos deformados (líneas oscuras) representan cambios de forma en los extremos de las PC, mientras que las líneas claras indican el promedio de la forma para el mismo eje.
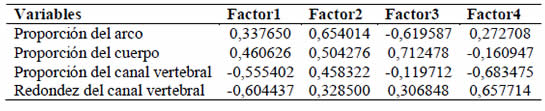
Tabla 3. Autovectores.
Los valores indican la importancia de las variables para cada uno de los factores.
Morfometría geométrica
La variación en el contorno del canal vertebral interno y externo también se analizó mediante morfometría geométrica. Los resultados obtenidos a partir de la digitalización de landmarks y semilandmarks en relación con el tamaño del centroide reflejan que el tamaño de los contornos del canal vertebral es mayor en promedio para el puma que para las otras especies comparadas (Fig. 3C-D). En algunos segmentos vertebrales, esa diferencia resulta más marcada, como se observa para C2, mientras que, en otros segmentos, como se registró para T1, esa diferencia es más sutil (Fig. 3E,F). A su vez, encontramos que el tamaño del canal vertebral del ocelote tiende a ser mayor que en el gato doméstico para algunos segmentos, mientras que esa relación se invierte en algunas vértebras, como la C6 (Fig. 3E,F).
En cuanto a las diferencias de forma, se observó que la variación principal estuvo dada por el tipo de vértebra. Para el contorno externo de las tres especies, S1 (relativamente más ancha) y C2 (más redondeada) se representan en los extremos opuestos de PC1 (Fig. 4C), excepto para S1 del puma, que estaba ubicada hacia el extremo positivo de P1, relativamente separada de las de los otros felinos (Fig. 4C). Esto indicaría que, en general, las vértebras del puma tenían una altura mayor en comparación con el ancho. A lo largo de PC2, la separación más marcada correspondía al Atlas, que ocupaba el extremo negativo con una morfología redondeada (Fig. 4C).
Con respecto al contorno interno, se encontró un patrón similar al del contorno exterior, aunque se observaba un mayor solapamiento de los puntos que correspondían a las vértebras (Fig. 4D). En general, se observó que el Atlas, el Axis y S1 tienen una forma de canal similar en las tres especies, mientras que el canal vertebral de C3 a L1 del ocelote y el gato también tienen una configuración similar (Fig. 4C-D). La relación entre la variación del tamaño y la forma se evaluó mediante la regresión de PC1 de las configuraciones internas y externas en función de los tamaños del centroide. En general, la asociación de estas dos variables tuvo menos dispersión en el interior (r2 = 0,45) que en el exterior (r2 = 0,22). Podemos afirmar que una gran parte de la variación de la forma no se predice por los cambios de tamaño (Fig. 4).
Discusión y conclusiones
En los felinos, la columna vertebral tiene una gran flexibilidad, lo que les permite realizar los movimientos de escalada y salto mientras capturan a la presa. De esta manera, se puede considerar que la columna vertebral funciona como un todo dinámico y no como una mera estructura estática de soporte 7,12. Ciertos autores describen la regionalización morfológica y funcional de la columna vertebral en felinos a través de módulos. Sin embargo, estos módulos no necesariamente están de acuerdo con las regiones vertebrales tradicionalmente descriptas (cervical, torácica y lumbar) 28. Los módulos morfológicos vertebrales reflejan grupos principales de vértebras que organizan al raquis de acuerdo con el desarrollo o la función y comparten patrones alométricos comunes 2.
En nuestro estudio comparativo observamos que la mayoría de las vértebras se ajustan a un patrón común de construcción anatómica en los felinos estudiados. En la región cervical observamos que el cuerpo vertebral de C2 es de mayor tamaño y que disminuye en sentido cráneo-caudal. La relación ancho:alto fue similar entre las tres especies de felinos. En cuanto al arco vertebral la relación ancho: alto disminuye de C3 a T1. La región cervical de la columna vertebral tiene una dirección en forma de S con una inflexión alrededor de la unión de C7 y T1.
En base a los análisis multivariados sobre las medidas lineales, C1 (Atlas) mostró una marcada diferenciación con respecto a la de las otras vértebras. En general, se mantuvo una similitud de las morfología de las vértebras, aunque se observaron diferencias puntuales en alguna de las especies estudiadas. C2 mostró diferencias en el ocelote con respecto a las otras especies, mientras que en el puma y en el gato doméstico se ubicaron más cercanas. Se puede inferir que este patrón se corresponde con la regionalización funcional y no refleja simplemente las regiones vertebrales principales 28. En los cuadrúpedos, la orientación vertical de la columna cervical y la modificación de C1 y C2 se interpreta como consecuencia de la necesidad de proporcionar equilibrio y estabilidad a la cabeza 7.
El tamaño del canal vertebral fue estudiado en diferentes especies como ratas y humanos 9, ciervo rojo 14, ratón, musaraña, conejo, ardilla gris 12 y varios primates 16. Más recientemente, se comparó el canal vertebral de ratas jóvenes y envejecidas 19. En felinos, diversos estudios muestran el análisis morfométrico de vértebras de diferentes especies de felinos agrupados en
linajes, que incluyen técnicas morfométricas lineales y geométricas 20. El presente estudio complementa estos resultados con una descripción morfológica exhaustiva.
Particularmente en el puma, parecería ser necesario un canal vertebral más grande para alojar los componentes nerviosos capaces de inervar su gran masa muscular, entre otras funciones. Más allá de estas diferencias de tamaño, la variación de la forma requiere una interpretación más detallada. A primera vista, las principales diferencias en la forma del canal se corresponden con particularidades de los segmentos vertebrales y no con la variación interespecífica. Esto significa que, a pesar de las variaciones más sutiles, la forma del canal sigue los patrones conservados que pueden rastrearse en todos los felinos o incluso en todos los mamíferos 11. Sin embargo, cuando los resultados se observan en detalle, encontramos diferencias entre las especies de felinos que se estudiaron en el presente trabajo. El puma se diferencia claramente por tener un canal vertebral más estrecho, lo que es evidente tanto por los patrones de la relación ancho:altura, como por la variación de la forma analizados mediante técnicas de morfometría geométrica. Esto puede ser un patrón alométrico que resulta del cambio de tamaño y/u otros factores, que pueden estar modelando la forma del canal, en el que este tipo de cambios se relaciona con los patrones de locomoción 16.
La evidencia genética y paleontológica respalda la idea de que Felis catus y Puma concolor están más estrechamente relacionados entre sí que con el ocelote, con un ancestro común que vivió hace 7.000.000 de años; mientras que el ancestro común de las tres especies se localizó hace alrededor de 8.000.000 años 29. Otros autores consideran que la separación entre los taxones ocurrió previamente; sin embargo, coinciden en que el puma y el gato doméstico son las especies más relacionadas 11. La mayor relación entre el puma y el gato doméstico puede explicar algunos de nuestros resultados, como las similitudes entre estas dos especies con respecto a las proporciones (ancho:altura) del cuerpo vertebral y la altura relativamente mayor en C7 y T1. En el ocelote, observamos algunas particularidades propias de esta especie, ya que no se encontraron en los otros felinos estudiados, tales como la tendencia al estrechamiento en el cuerpo y el canal central de las vértebras cervicales.
Observamos que las proporciones de ancho:altura del cuerpo vertebral en las regiones lumbar y sacra son similares entre el ocelote y el puma. Dado que estas regiones tienen un papel clave en términos biomecánicos para funciones como la escalada y el salto, podemos formular la hipótesis de que la forma de sus vértebras sobrevino a las demandas funcionales similares.
Conclusiones
Estudios funcionales comparativos entre animales con una anatomía musculoesquelética similar son relevantes para comprender la relación forma-función 20. La división en compartimientos que utilizamos en nuestro estudio y medición ha sido utilizada por otros autores 20. Esta división no necesariamente coincide con las regiones tradicionales (cervical, torácica y lumbar), pero sí parecen estar relacionadas con la función y adaptaciones al hábitat. Las principales diferencias encontradas en la columna vertebral de los felinos estudiados parecen estar relacionadas con los diferentes patrones de locomoción y las adaptaciones al tamaño de la presa 4. Los aumentos evolutivos en el tamaño corporal han promovido la estabilización de la región cervical y la modificación generalizada de C1 y C2 en las tres especies de felinos estudiados, mientras que las diferencias halladas en la región lumbar y sacra demuestran cómo los felinos americanos han adaptado su esqueleto axial para la locomoción, la escalada y el desplazamiento en tierra. La mayor relación entre el puma y el gato doméstico en nuestros resultados probablemente se deba a características ontológicas de los linajes que se encuentran más relacionados11.
Dada la escasa cantidad de muestras analizadas se espera que en futuros estudios podamos contar con un mayor número de individuos de distinto sexo y edades, así como comparar especímenes de vida silvestre y en cautiverio. Asimismo, esperamos poder realizar estudios comparativos del esqueleto axial con el esqueleto apendicular para poder determinar, con mayor precisión, la especialización locomotora de estos felinos.
1. Abreu, K.C.; Moro-Rios R.F.; Silva-Pereira J.E.; Miranda J.M.D.; Jablonski E.F.; Passos F.C. Feeding habits of the Ocelot (Leopardus pardalis) in Southern Brazil. Mamm Biol. 2008; 73:407-11. [ Links ]
2. Clune, J.; Mouret, J.B.; Lipson, H. The evolutionary origins of modularity. Proc Biol Sci. 2013; 280(1755):2012-863. [ Links ]
3. Culver, M.; Johnson, W.E.; Pecon-Slattery J.; O'Brien S.J. Genomic ancestry of the American puma (Puma concolor). J Heredity. 2000; 91:186-97. [ Links ]
4. Day, L.M.; Jayne B.C. Interspecific scaling of the morphology and posture of the limbs during the locomotion of cats (Felidae). J Exp Biol. 2007; 210(4):642-54. [ Links ]
5. De Villa-Meza, A.; Martínez-Meyer, E.; López-González, C.A. Ocelot (Leopardus pardalis) food habits in a tropical deciduous forest of Jalisco, México. Am Midl Nat. 2002; 148:146-54. [ Links ]
6. Di Bitetti, M.S.; Paviolo, A.; De Angelo, C. Density, habitat use and activity patterns of ocelots (Leopardus pardalis) in the Atlantic forest of Misiones, Argentina. J Zool (1987). 2006; 270:153-63. [ Links ]
7. Done, S.H.; Goody, P.C.; Evans, S.A.; Stickland, N.C. Atlas en color de Anatomía Veterinaria. El perro y el gato. 2ª Edición. Elsevier, Barcelona, 2010. [ Links ]
8. Dyce, K.M.; Sack, W.O.; Wensing, C.J.G. Textbook of veterinary anatomy. 4th Edition Saunders/Elsevier. St. Louis, Mo, 2010. [ Links ]
9. Flynn, J.R.; Bolton, P.S. Measurement of the vertebral canal dimensions of the neck of the rat with a comparison to the human. Anat Rec (Hoboken). 2007; 290:893-9. [ Links ]
10. Johnson, W.E.; Eizirik, E.; Pecon-Slattery, J.; et al. The Late Miocene radiation of modern Felidae: A genetic assessment. Science. 2006; 311:73-7. [ Links ]
11. Jones, K.E.; Pierce S.E. Axial allometry in a neutrally buoyant environment: effects of the terrestrial-aquatic transition on vertebral scaling. J Evol Biol. 2016; 29:594-601. [ Links ]
12. Kida, M.Y.; Johnson, D.R.; McAndrew, T.J.; O'Higgins, P. Adaptation in the vertebral column: a study of patterns of metameric variation in seven species of small mammals. J Anat. 1999; 194:207-14. [ Links ]
13. Klingenberg, C.P. MorphoJ: an integrated software package for geometric morphometrics. Mol Ecol Resour. 2011; 11:353-7. [ Links ]
14. Kumar, N.; Kukreti, S.; Ishaque, M.; Sengupta, D.K.; Mulholland, R.C. Functional anatomy of the deer spine: an appropriate biomechanical model for the human spine? Anat Rec (Hoboken). 2002; 226:108-17. [ Links ]
15. Lyonsc, L.A.; Murphyb, W.J.; Warrena, W.C. Comparative analysis of the domestic cat genome reveals genetic signatures underlying feline biology and domestication. Proc Natl Acad Sci U S A. 2014; 111:17230-5. [ Links ]
16. Manfreda, E.; Mitteroecker P.; Bookstein F.L.; Schaefer K. Functional morphology of the first cervical vertebra in humans and nonhuman primates. Anat Rec B New Anat. 2006; 289:184-94. [ Links ]
17. Meachen-Samuels, J.; van Valkenburgh, B. Forelimb indicators of prey-size preference in the Felidae. J Morphol. 2009; 270(6):729-44. [ Links ]
18. Moreno, R.R.; Kays, R.; Samudio, R. Jr. Competitive release in the diet of ocelots (Leopardus pardalis) and puma (Puma concolor) after jaguar (Panthera onca) decline. J Mammal. 2006; 87(4):808-16. [ Links ]
19. Nishida, F.; Barbeito J.; Barbeito C.G.; Portiansky E.L. Is the vertebral canal prepared to host the aged spinal cord? A morphometric assessment. Zoomorphology. 2014; 133:219-25. [ Links ]
20. Randau, M.; Goswani, A.; Hutchinson, J.R.; Cuff, A.R.; Pierce, S.E. Cryptic complexity in felid vertebral evolution: shape differentiation and allometry of the axial skeleton. Zool J Linn Soc. 2016; 178:183-202. [ Links ]
21. Randau, M.; Goswami, A. Morphological modularity in the vertebral column of Felidae (Mammalia, Carnivora). BMC Evol Biol. 2017; 17(133):1-12. [ Links ]
22. Rohlf, FJ. Morphometrics at SUNY Stony Brook. Thin-plate spline. Disponible en http://life.bio.sunysb.edu/morph/. Consultado 3 de noviembre 2016. [ Links ]
23. Rohlf, F.J.; Slice, D.E. Extensions of the Procrustes method for the optimal superimposition of landmarks. Syst Zool. 1990; 39:40-59. [ Links ]
24. Rosenzweig, LJ. Dubuque. Anatomy of the Cat: Text and Dissection Guide. W.C. Brown Publishers. Dubuque, 1990. [ Links ]
25. Saggese, M.D. Felinos Autóctonos. (1999). Disponible en http://www.oocities.org/heartland/garden/4105/felaut.html. Consultado 15 de octubre 2016. [ Links ]
26. Sánchez, H.L.; Silva, L.B.; Rafasquino, M.E.; et al. Anatomical study of the forearm and hand nerves of the domestic cat (Felis catus), puma (Puma concolor) and jaguar (Panthera onca). Anat Histol Embryol. 2013; 42(2):99-104. [ Links ]
27. Sunquist, M.; Sunquist, F. Wild Cats of the World. University of Chicago Press. Chicago, 2002. [ Links ]
28. Vidal, P.P.; Graf, W. y Berthoz, A. The orientation of the cervical vertebral column in unrestrained awake animals. Exp Brain Res. 1986; 61:549-59. [ Links ]
29. Werdelin, L.; Nobuyuki, Y.; Johnson, W.; O'Brien, S. Biology and Conservation of Wild Felids. Oxford University Press. Oxford, 2010. [ Links ]

