










Services on Demand
Journal

Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO
Related links
-
 Similars in
SciELO
Similars in
SciELO
Share

 Permalink
PermalinkCuadernos de la Facultad de Humanidades y Ciencias Sociales. Universidad Nacional de Jujuy
On-line version ISSN 1668-8104
Cuad. Fac. Humanid. Cienc. Soc., Univ. Nac. Jujuy no.22 San Salvador de Jujuy May 2004
Diversidad biológica humana en la provincia de Salta
Human biological diversity in Salta
Noemí Acreche *, **, *** - María Virginia Albeza *, **, *** - Graciela B. Caruso **, *** - Viviana G. Broglia **, *** - Rebeca Acosta **, ***
* Facultad de Humanidades - Universidad Nacional de Salta - Buenos Aires N° 177 - CP 4400 - Salta - Argentina.
** Facultad de Ciencias Naturales - Universidad Nacional de Salta - Av. Bolivia Castañares - CP 4400 - Salta - Argentina.
*** Consejo de Investigación - Universidad Nacional de Salta.
Noemí Acreche: Correo Electrónico: acreche@unsa.edu.ar
RESUMEN
La Provincia de Salta cuenta con una marcada heterogeneidad geoestructural, distinguiéndose básicamente las siguientes unidades: Puna, Valle Calchaquí, Valle de Lerma y Chaco, habitadas por el hombre desde hace al menos 9000 años.
Las características ambientales y topográficas pueden determinar el aislamiento y la adaptación de las poblaciones provocando su diferenciación a nivel genético. Para poner a prueba esta relación, se consideraron variables genéticas y parámetros demográficos.
En la Puna se analizaron los grupos eritrocitarios (Santa Rosa de los Pastos Grandes, Tolar Grande y Cobres) y leucocitarios (Cobres), heterocromatina constitutiva (Tolar Grande y Cobres) y STRs (San Antonio de los Cobres y Cobres) además de indicadores de Demografía Genética (Santa Rosa de los Pastos Grandes, Tolar Grande, Olacapato, San Antonio de los Cobres y Cobres). En el valle de Lerma se estimaron los parámetros demográfico-genéticos (San Agustín y Chicoana) y en el Chaco se analizó la heterocromatina constitutiva. Asimismo, se incorporaron al análisis los datos de Matson et al (1969) y Pagés Larraya (1978) referentes a grupos eritrocitarios del Valle Calchaquí y el Chaco.
Los procesos microevolutivos estudiados (Selección Natural, Deriva Génica y Flujo Génico) a partir de medidas de variabilidad genética intra e interpoblacional, Indices de Oportunidad para la Selección Natural y Coeficientes de Endogamia y Aislamiento Reproductivo evidencian un fuerte y continuado contacto entre las poblaciones, moderada diferenciación y en algunos casos alta endogamia.
Palabras Clave: Diversidad humana; Variabilidad genética; Parámetros demográficos; Procesos microevolutivos; Diferenciación poblacional; Endogamia.
ABSTRACT
The study area is highly heterogeneous, presenting the following ecological units: Puna, Valle Calchaquí, Valle de Lerma and Chaco. Environment and topography can determine isolation and adaptation of human population, inducing genetic differentiation. In order to consider this fact, several genetic and demographic variables were studied.
In Puna, eritrocitary variants were analized in Santa Rosa de los Pastos Grandes, Tolar Grande and Cobres, leucocitary in Cobres, while Constitutive Heterochromatine was studied in Tolar Grande and Cobres and STRs in San Antonio de los Cobres and Cobres. On the other hand, Genetic Demography was used to study Opportunity for Selection, Reproductive Isolation and Endogamy in Santa Rosa de los Pastos Grandes, Tolar Grande, Olacapato, San Antonio de los Cobres and Cobres. In Lerma Valley, parameters for Genetic Demography were estimated in San Agustín and Chicoana; in the Calchaquí Valley in Pichao and La Alumbrera and in Chaco Constitutive Heterochromatine was analized. Data published by Matson et al (1969) and Pagés Larraya et al (1978) were included.
Microevolutive processes studied (Natural Selection, Genetic Drift and Gene Flow), variability measures show strong and continued contact among populations, moderate differentiation and, in some cases, high endogamy.
Key Words: Human diversity; Genetic variability; Demographic parameters; Microevolutive process; Popultion differentiation; Endogamy.
INTRODUCCIÓN
POBLACIÓN Y ÁREA DE ESTUDIO
La región del Noroeste Argentino (NOA) cubre una superficie de 466.740 Km2, el 16,71% de la del país. Por su ubicación geográfica, franja subtropical y la influencia de factores climáticos es muy diversa presentando variedad de unidades estructurales: Puna, Cordillera Oriental, Sierras Subandinas, Sierras Pampeanas y Llanura Chaqueña.
Las poblaciones humanas presentan estrategias diferentes vinculadas a la diversidad ambiental que determina diferencias en los recursos disponibles para su establecimiento y supervivencia.
Así, la diversidad humana es evidente desde tiempos prehispánicos. Tanto en la Puna, en las zonas de Valles Intermontanos como en el norte de Chile y el sur de Bolivia, durante ese período tuvo lugar el desarrollo de un verdadero mosaico de culturas (Krapovickas, 1983) locales con un permanente y constante intercambio entre ellas. Indicios arqueológicos señalan un aumento de población hacia fines de los siglos XIV y XV probablemente por corrientes migratorias, revirtiéndose dicho proceso en el período previo a la expansión incaica, con el abandono de centros poblados.
Los Incas ingresaron en el actual territorio argentino hacia 1480, aproximadamente, utilizando vías de acceso naturales que transformaron en caminos, comunicando al Cuzco con Bolivia, el NOA y Chile (Martínez Sarasola, 1992). A la llegada de los españoles, el NOA, cuya población estimada era de 200.000 habitantes, constituía la región más poblada del actual territorio argentino (75% del total), (Martínez Sarasola, 1992). En 1800 poseía el 40% de la población nacional; sin embargo, hacia 1869 (Primer Censo Nacional) representaba el 26%. A partir de 1970 las tasas de crecimiento total de la región fueron siempre mayores que las registradas en el país (aproximadamente el 18%), (Boleda, 1993).
Durante el período de organización nacional, comenzó a ocuparse más la zona de climas subtropicales como Valles Calchaquíes, por ejemplo, permaneciendo con escasa población las regiones chaqueña y andina. Actualmente, la población del NOA representa el 11% de la población total de Argentina, con una distribución poco homogénea (aproximadamente el 34% de su superficie despoblada) como algunas áreas de la Puna y Chaco (Pontussi, 1993).
En general, la región norte ha sufrido el deterioro de sus economías y ha acentuado sus diferencias con el resto del país. Los menores ingresos por habitante los tienen las regiones NOA y NEA, con los más bajos índices de educación, salud y vivienda y altas tasas de desnutrición (Pontussi, 1993). Estos índices reflejan de manera directa el marginamiento no sólo geográfico sino social, político y económico de un gran número de áreas de la región manifestado a través de la insatisfacción de necesidades básicas de subsistencia y la importante dificultad de producción.
La provincia de Salta es la de mayor extensión en ese contexto, representando el 33,16% de su superficie, con 154.775 Km2. En cuanto a su población, en el período 1980 -1991, se registró la tasa de crecimiento mayor dentro de la región con un valor de 23,2% frente a la media nacional de 15,80% (Pontussi, 1993). Cabe destacar que, aún cuando en los primeros lustros del siglo XX el NOA registró tasas de migración neta negativas (sobre todo después de 1970), Salta y Jujuy mantuvieron siempre un saldo positivo de migración debido principalmente a que en estas provincias existe una inmigración importante, proveniente principalmente de Bolivia (Boleda, 1993).
Por su extensión, presenta una variedad de unidades geoestructurales: Puna, Valles Intermontanos y Chaco, con marcadas variaciones tanto en clima y topografía, como en flora y fauna. Así, las temperaturas varían ampliamente desde las máximas que se registran en la región chaqueña a las mínimas en la Puna, mientras que las precipitaciones decrecen en sentido este - oeste. Estas características permiten afirmar que Salta es una provincia diversa. Así, la variedad de ambientes que presenta, configuran y condicionan los asentamientos humanos.
La Puna es la prolongación austral del altiplano peruano - boliviano ubicada a una altitud media de 3.200 msnm. En Salta abarca los departamentos de Los Andes, parte del de La Poma y parte del de Rosario de Lerma. El clima es frío y seco. Según Cabrera (1971) comprende las provincias fitogeográficas de la Puna, Altoandina y Patagónica.
La evidencia arqueológica sugiere que la región de la Puna, es habitada desde, en el caso del altiplano peruano hace unos 12.000 años y el chileno más de 8.000 años (Rothhammer y Schull, 1980). En Yavi, Jujuy se han hallado restos de hace 9.000 años (Kulemeyer, comunicación personal). Esta región ha presentado marcadas fluctuaciones poblacionales producto de diversos eventos históricos. En la actualidad se caracteriza por un marcado predominio de niños sobre adultos y ancianos debido, principalmente a las migraciones estacionales, fundamentalmente masculinas, a las zonas subtropicales, salvo en aquellos sectores como los centros mineros que atraen mano de obra (Albeza, 1995).
Las características de la región puneña con un clima riguroso y condiciones de hipotermia e hipoxia, inducen al desarrollo de una sociedad cuya base productiva es la unidad doméstica que en conjunto representan una economía de subsistencia. Así, esta sociedad ha desarrollado un complejo adaptativo en el que incluye la transhumancia cíclica como forma de resolver la escasez estacional de los recursos necesarios para la actividad pastoril, lo que resulta en un patrón de poblamiento disperso, cuya característica más notable es la ausencia de centros densamente poblados y la estacionalidad en la ocupación de poblados y puestos de montaña (Albeza, 1995; Caruso, 1995).
Los Valles Intermontanos como el Valle de Lerma y los Valles Calchaquíes presentan clima de tipo subtropical.
Los Valles Calchaquíes, están formados por los valles del río Calchaquí y el extremo norte del río Santa María, ocupando una faja de 200 Km aproximadamente de largo y extendiéndose en las provincias de Tucumán y Catamarca. Se ubican en sentido norte-sur, con una altitud que varía entre los 1.680 msnm hasta los 3.015 msnm. Políticamente, en la Provincia de Salta abarcan los departamentos de Cachi, La Poma, Molinos, San Carlos y Cafayate. El clima es templado y según Cabrera (1971) la región pertenece a la provincia fitogeográfica del monte.
Las poblaciones humanas están desigualmente distribuídas, estimándose alrededor de 25.000 habitantes en total. Las localidades más pobladas son Cafayate, con 9.264 habitantes y Cachi, con 6.157 habitantes (Censo Nacional 1991). Asimismo, se registran desplazamientos poblacionales hacia los valles más bajos en época de cosecha. La producción vitivinícola es una de las más importantes y los pequeños productores se dedican al cultivo de pimiento para pimentón (Bertoncello y García, 1995).
El Valle de Lerma, tiene una extensión de 115 Km, en sentido norte-sur, una superficie de 10.500 Km2 y una altitud promedio de 1.200 msnm. Incluye los departamentos de La Caldera, Capital, Cerrillos, sur este del de Rosario de Lerma, Chicoana, La Viña y Guachipas. El clima es Subtropical Serrano con una estación seca. Sin embargo, sus características fisiográficas le imponen particularidades que permiten definir verdaderos climas locales (Baumgartner y Cozzi, 1998). Según Cabrera (1971) la zona de estudio se encuentra comprendida en la Provincia Chaqueña. Registra 448.180 habitantes, totalizando el 51,74% de la población provincial. Cabe destacar que el 92,15% de la población es urbana (Baumgartner y Cozzi, 1998).
La unidad geoestructural del Chaco se ubica entre los 500 y 250 msnm, aquí se encuentra la isoterma de 47ºC, aunque la temperatura media en verano es de 26ºC y en invierno de 16ºC. Desde el punto de vista fitogeográfico corresponde a la provincia Chaqueña. Políticamente, en la provincia de Salta, lo conforman los departamentos de San Martín, Rivadavia, la región oriental de los departamentos de Orán y Anta y el oeste del de Metán y Rosario de la Frontera.
La ocupación de la zona del Chaco data, según fechados arqueológicos de al menos 9.000 años. Se consideran varias hipótesis de su poblamiento, como el arribo de oleadas migratorias de grupos amazónidos que se asentaron sobre un sustrato paleochaqueño. Así, se plantea al chaco como un embudo cultural, donde confluyeron diversas culturas en distintas épocas o como un corredor por donde se movieron diversos grupos de noroeste a sur y de este a oeste y visceversa. En la actualidad diversos grupos aborígenes habitan esta unidad geoestructural, Mataco, Chorote, Chulupi, Toba, Pilagá, Mocoví, Chiriguano, Chané, junto a la población caracterizada como criollos. Históricamente los españoles realizaron esfuerzos de ocupación, pero ésta no se completó sino hacia 1.700 con el sometimiento de las poblaciones aborígenes. Las zonas centro y norte de la región se mantuvieron como territorios exclusivamente habitados por comunidades aborígenes, disminuyendo este espacio lentamente hasta la invasión final hacia fines del siglo XIX (Karlin et al, 1994).
Se registran en esta unidad altos índices tanto de analfabetismo (59,45%), como de NBI (81,9%). La situación anteriormente mencionada influye negativamente en el estado sanitario de la población. La presencia de enfermedades endémicas como Chagas-Mazza, y Leishmaniasis está asociada a la ausencia de una dieta adecuada, registrándose además enfermedades vinculadas principalmente a la deficiente calidad y disponibilidad de agua y a la falta de infraestructura para el tratamiento de las excretas y desperdicios.
Así, el complejo conjunto de características físicas de la región, las fluctuaciones y movimientos poblacionales que tuvieron lugar en ella y las condiciones actuales de relativo aislamiento y tamaño pequeño de muchas de sus poblaciones, la señala como de alto interés para realizar estudios microevolutivos.
MARCO GENÉTICO-POBLACIONAL
Se entiende por Evolución el cambio en las frecuencias génicas de una población. Estos cambios, en nuestra especie, estudiados entre dos generaciones se conocen como cambios microevolutivos.
Así, una población es un conjunto de individuos que comparten un pool génico en una misma dimensión espacial y temporal. Para que una población evolucione deben existir fuerzas o factores que promuevan la variación de las frecuencias génicas y genotípicas de la misma. Los factores deterministas: Flujo Génico, Selección Natural, Mutación, entre los más estudiados, tienden a cambiar las frecuencias de manera predecible tanto en magnitud como en dirección. En los estocásticos, Deriva Génica, es posible medir la magnitud pero no así la dirección del cambio.
La definición de los límites poblacionales en nuestra especie es generalmente arbitraria. Por ejemplo, en el caso de la Puna, la población, por razones geográficas y socio económicas, se encuentra distribuida en pequeñas localidades, puestos o poblados parcialmente aislados, en muchos casos a considerables distancias entre sí, resultando difícil definir los límites de lo que se considera una población determinada.
El Flujo Génico es la expresión genética del fenómeno demográfico denominado migración. Así, la migración posibilita el ingreso o salida de genes de una población variando por tanto la frecuencia de los mismos.
La Selección Natural opera por muerte o nacimiento diferencial y es producida por la capacidad diferencial de los distintos tipos hereditarios presentes en una población de dar descendientes en un medio determinado.
La Mutación es la fuente de variabilidad en una población, pero considerando su baja frecuencia, generalmente se la excluye de estudios poblacionales en humanos ya que requiere de muestras poblacionales excesivamente grandes para su detección.
La Deriva Génica, ocurre en poblaciones pequeñas debido a los efectos de muestreo, ajenos a la intervención del observador. Este proceso consiste en la fluctuación al azar de las frecuencias génicas en una población que se acumulan de generación en generación hasta llegar a la fijación de un alelo, disminuyendo la variabilidad intrapoblacional.
Estos factores que determinan la divergencia genética podrían variar conjuntamente con factores culturales. La relación entre divergencia cultural y genética ha sido estudiada en numerosas poblaciones (Acreche et al, 1996; Albeza, 1995; Broglia, 1998; Caruso, 1995). Tradicionalmente se ha intentado delimitar subdivisiones en la especie humana, considerando el concepto especie desde la perspectiva realista. Así, se han utilizando ciertos rasgos morfológicos, descriptivos y métricos, como pigmentación, caracteres faciales, entre otros, escogidos por su aparente facilidad de reconocimiento generándose en consecuencia una variedad de clasificaciones raciales. Sin embargo, estos caracteres como muchos otros son morfológica y genéticamente complejos y aún cuando su heredabilidad es alta, están sujetos a fuertes influencias ambientales por lo que los resultados incluyen un amplio margen de error (Caruso, 1995).
Es importante asentar en este punto que no se han encontrado caracteres biológicos que permitan definir inequívocamente entidades biológicas tales como razas. De esta manera no es posible, por ejemplo, asignar un alelo determinado a una entidad o subentidad biológica ya que para ello sería necesario reconocer situaciones de aislamiento (Deriva Génica) y/o procesos de adaptación (Selección Natural) prolongados en el tiempo para poder realizar esta asunción. No se busca una justificación biológica a-priori de las diferencias culturales observadas. Sí es posible observar algún tipo de diferencias biológicas vinculadas a las diferentes estrategias de utilización del espacio y el tiempo en la amplia variación de ambientes por parte de las poblaciones humanas asentadas en ellos.
La variabilidad genética es el sustrato sobre el que actúan los procesos evolutivos. Su estudio requiere del conocimiento de las propiedades de los genes y de la descripción de la población en términos de frecuencias fenotípicas, genotípicas y génicas, parámetros que permiten cuantificar la variabilidad intra e interpoblacional.
En las poblaciones humanas la mayor parte de los polimorfismos estudiados pueden agruparse en alguna de las siguientes categorías principales:
¨ Polimorfismos detectables mediante técnicas inmunológicas: antígenos eritrocitarios (sistemas ABO, Rh, MNSs, etc.), antígenos leucocitarios (polimorfismo HL-A) y antígenos plaquetarios (sistemas Zwa y Kua).
¨ Polimorfismos proteínicos detectables por electroforesis: variantes de hemoglobina, proteínas plasmáticas (haptoglobinas, sistema Gm, grupos Gc, etc.) e isoenzimas plasmáticas y celulares (grupos Xm, Gc, G-6PD, etc.).
¨ Secuenciación de aminoácidos.
¨ Polimorfismos de ADN (nuclear y mitocondrial), estudiados mediante técnicas de hibridación, secuenciación, fragmentos de restricción, etc.
¨ Otra aproximación al conocimiento de la diversidad genética en poblaciones humanas es a través de estudios citogenéticos como la Heterocromatina Constitutiva.
¨ A partir de parámetros demográfico-genéticos se puede estimar la variabilidad.
Las variaciones bioquímicas, que dependen de la estructura de las proteínas o de diferencias inmunológicas son los fenotipos resultantes de la expresión directa o muy próxima de los genes. La probabilidad de que efectos ambientales o epistáticos modifiquen esta expresión es baja, ofreciendo numerosos casos de variación discontinua. La mayor parte de los polimorfismos humanos se encuentran en casi todas las poblaciones, aunque las frecuencias de los distintos alelos pueden diferir considerablemente entre grupos.
El factor evolutivo conocido como Flujo Génico mide los efectos de la migración de individuos o conjuntos de individuos sobre la estructura genética de las poblaciones.
El flujo de genes puede cambiar la estructura genética introduciendo nuevos alelos desde otras poblaciones, por pérdida de variabilidad en algunos casos de emigración o simplemente por cambio en las frecuencias por el contacto entre grupos con diferencias en este nivel.
La acción del Flujo, en el caso de la introducción de nuevos alelos, incrementa la variabilidad siendo semejante a la de la mutación. Puede ocurrir aleatoriamente o bien presentarse en algunos casos vinculada a la Selección Natural. Sin embargo, el factor con el que se encuentra más estrechamente vinculado es la Deriva Génica ya que aparecen como fuerzas de efectos contrapuestos, al menos en los resultados que producen.
En toda población se encuentran individuos portadores de diferentes genotipos, esto puede provocar una supervivencia y reproducción diferencial, lo que significa que algunos de los individuos de la población portan combinaciones de genes que les permiten estar mejor adaptados al ambiente en el que se encuentran que otros. Cuando una combinación particular de genes confiere una ventaja a sus portadores con respecto a aquellos que llevan combinaciones alternativas, actúa la Selección Natural. La fuerza relativa de ésta varía con la ventaja que proporciona, es decir con la probabilidad de sobrevivir y dejar descendencia (eficacia biológica).
Es muy difícil probar que un carácter es adaptativo, ya que esto implica afirmar que la Selección Natural está favoreciendo una variante determinada en un ambiente dado para lo que es necesario estimar las diferencias de fertilidad y viabilidad, es decir los nacimientos y muertes diferenciales asociados a ese carácter determinado. Para ello, deberían analizarse la supervivencia y la media y varianza del número de hijos de los portadores de las distintas variantes considerando la acción conjunta de otros genes y teniendo presente que en nuestra especie estos parámetros están sujetos a factores culturales, sociales y económicos.
Además es importante tener en cuenta en el análisis que las fuerzas selectivas que actúan en los diferentes loci pueden tener distinta intensidad y dirección. Es posible también que en los casos en los que se observa correlación entre una variante y el ambiente no haya una relación causa efecto y se puede pensar que el gen analizado esté ligado a un locus que sí se encuentre bajo la acción de la Selección Natural.
El aislamiento entre dos o más poblaciones es de gran importancia como fenómeno evolutivo ya que si se mantiene durante largos períodos de tiempo, la acumulación de mutaciones en cada una de ellas puede conducir a su diferenciación. (Acreche, 1994; Dobzhansky et al, 1983). Las características climáticas, topográficas y los sistemas culturales y productivos son factores que de manera directa pueden incidir en el aislamiento de una población.
Estudios realizados en poblaciones pequeñas explican la divergencia genética entre éstas como un efecto del proceso dispersivo que conocemos como Deriva Génica ya que al ocurrir independientemente en diferentes subpoblaciones, conduce a la diferenciación genética entre ellas (Azofeifa y Barrantes, 1991; Bertranpetit y Cavalli-Sforza, 1991; Cavalli-Sforza, 1969; Cross, 1976; Glass, 1953). Tal diferenciación se ve afectada por el grado de contacto que mantengan las subpoblaciones (Flujo Génico), que a su vez puede ser función de la separación geográfica entre ellas (Wright, 1943).
Existe un alto grado de intercambio entre sitios registrado como migración o Flujo Génico. Sin embargo, el límite asignado a la población está directamente vinculado a la estimación de su grado de sujeción a la Deriva por medio de métodos biodemográficos, dependiente del número de habitantes y sobre todo de los que efectivamente se reproducen en ellas.
Las comunidades pequeñas y aisladas o semiaisladas presentan un particular interés que alcanza aspectos tanto teóricos como prácticos. Si bien las consideraciones teóricas permiten prever que hay algún grado de consanguinidad en estas poblaciones, su evaluación y seguimiento es particularmente importante, sobre todo considerando la tendencia secular a la disolución de los aislados.
Los sistemas de apareamiento se definen por cruzamientos sistemáticos entre individuos y constituyen un fenómeno complejo. Para analizarlos, es necesario tener en cuenta factores físicos, demográficos y socio-culturales.
Hay especies que se aparean tan cercanamente al azar que puede asumirse la condición panmíctica para el equilibrio. Sin embargo, esto es probablemente poco común. Las desviaciones pueden deberse a apareamientos selectivos por diferentes motivos tales como la elección de la pareja en función de relaciones de parentesco o la similitud fenotípica, pero también pueden ocurrir desviaciones de la panmixia por aislamiento espacial más o menos completo de poblaciones locales.
El grado de aislamiento tiene relación a su vez con el grado de endogamia y la consanguinidad. En aislados poblacionales de baja densidad, el grado de consanguinidad es mayor, no necesariamente porque se alejan de la panmixia, sino como consecuencia de la pequeña magnitud de sus efectivos. En efecto, aún manteniendo cruzamientos al azar, la probabilidad de que los progenitores de un individuo estén emparentados, es alta (Magalhaes y Arce-Gómez, 1987a).
METODOLOGÍA
Desde esta perspectiva y, considerando los procesos que pueden estar involucrados, los estudios realizados por este grupo de trabajo han sido abordados desde diferentes metodologías y con diferentes cuerpos de datos independientes entre sí.

Tabla 1. Factores estudiados a través de diferentes metodologías.
Desde la Demografía Genética, a partir del análisis de parámetros demográficos se pueden estimar y evaluar sujeción a Deriva Génica, efectos de Selección Natural y Flujo Génico en las poblaciones.
Desde la Citogenética se analiza la influencia de las condiciones ambientales sobre características de posible valor adaptativo. Se estudia la variabilidad genética intra e interpoblacional en polimorfismos de Heterocromatina Constitutiva en diferentes ambientes.
Desde la Genética de Poblaciones, se intenta establecer relaciones genéticas y efectos de Deriva Génica en poblaciones a partir de las frecuencias génicas de los sistemas de grupos sanguíneos y antígenos leucocitarios.
Finalmente la perspectiva molecular a través del análisis de STRs nos permite evaluar los cambios de las frecuencias génicas a través de fuerzas como el Flujo Génico y la Deriva Génica.

Tabla 2. Poblaciones estudiadas por unidad geoestructural y metodologías aplicadas.
Fuente: 1 Matson et al, 1969; 2 Pagés Larraya et al, 1978; 3 Acreche et al, 1999; 4 Caruso et al, 1999; 5 Caruso, 1995; 6 Vullo, Comunicación personal; 7 Broglia, 1998; 8 Albeza et al, 2002; 9 Albeza, 1995; 10 Acreche y Albeza, 2001; 11 Acreche y Albeza, ms; 12 Acreche et al, 1997.
Las dos medidas más usadas para expresar numéricamente la cantidad de variabilidad en una población son la Cantidad de Polimorfismo (P) y la Heterozigosis Media (H) (Doolitle, 1986).
Se consideró un locus polimórfico cuando la frecuencia del alelo más común no es superior a 0,99. Este criterio se adoptó por considerarse que permite distinguir los loci con alelos cuyas frecuencias son tan altas que no pueden ser explicadas solamente por mutación recurrente.
Para evaluar la acción del Flujo Génico se consideró que existe correlación entre la distancia genética y la geográfica. Los migrantes se clasifican de acuerdo a criterios establecidos en función de esta última y atendiendo a otra información pertinente para la evaluación del grado de aislamiento entre la o las poblaciones. El supuesto subyacente es el que grupos geográficamente más cercanos, son también más parecidos o cercanos genéticamente ya que la distancia geográfica impone una limitación física al flujo de genes. Factores culturales tales como la lengua, organización social, prácticas religiosas o uso del medio, deben también ser considerados en la delimitación de grupos entre los que se espera hipotéticamente un mayor o menor contacto.
Una correcta aproximación a la inclusión de estas consideraciones en la evaluación del efecto de la migración en la estructura genética de las poblaciones consiste en clasificar los migrantes en migrantes de corta y migrantes de larga distancia, ponderando de manera diferencial estos dos grupos en el cómputo del aislamiento y su consecuencia, la sujeción a la endogamia o a la Deriva Génica.
Para el caso de las poblaciones de la Puna estudiadas, se consideró que teniendo en cuenta factores históricos, geográficos y físicos, los migrantes de corta distancia serían aquellos que, sin importar la distancia real, provinieran de localidades situadas en la misma unidad geomorfológica. Se consideraron de larga distancia a aquellos individuos provenientes de la zona de valles u otras unidades, sin considerar como determinante la distancia real y absoluta (Albeza, 1995). Similar criterio se siguió con las poblaciones del Valle Calchaquí, considerando de corta distancia a los migrantes provenientes del mismo Valle (Acreche y Albeza, 2001).
En el Valle de Lerma, tradicional zona de contacto, por otra parte, se consideraron migrantes de corta distancia a aquellos provenientes de localidades situadas a distancias menores de 60 Km (Acreche et al, 1997), significativo para las dimensiones del Valle.
Desde el punto de vista genético-poblacional, no todos los migrantes, sea que ingresen o salgan de una población son importantes. En este sentido, la edad y el sexo de los individuos es un punto a considerar ya que, en ausencia de otro tipo de información puede aportar a la comprensión del sentido, objeto y duración en el tiempo del fenómeno.
Debe tenerse en cuenta que no todos los migrantes que ingresan a una población dejan descendencia en ella, por lo que debe calcularse la migración efectiva, me, que representa la fracción de individuos que efectivamente contribuyen con sus genes a la siguiente generación.
Los valores de la tasa de migración efectiva pueden estar afectados en el área de estudio en particular por los desplazamientos masculinos y femeninos vinculados con la disponibilidad de empleo influyendo en la reducida población reproductora. Del mismo modo, la salida de individuos pertenecientes a la fracción reproductora, reduce su número.
A pesar de las dificultades para estudiar la acción de la Selección Natural en poblaciones humanas, es posible estimar su acción a través del análisis demográfico genético que permite calcular el Indice de Oportunidad para la Selección a partir de la fracción de oportunidad para la selección debida a la mortalidad y el potencial de selección debido a la fecundidad diferencial (Magalhaes y Arce Gómez, 1987b). Con métodos citogenéticos, se estudió la relación entre la cantidad de Heterocromatina Constitutiva en los cromosomas 1, 9, 16 e Y y la altitud a la que habitan las diferentes poblaciones, siguiendo la hipótesis de Ibraimov (1993) que supone bandas menores a mayor altitud.
El tamaño de la población, el aislamiento, la formación de parejas de modo no aleatorio, la endogamia y la consanguinidad son de gran influencia cuando se estudia la estructura genética de una población y se la compara entre dos generaciones. Los cambios que se producen cuando el tamaño es pequeño, el aislamiento y por ende la endogamia y consanguinidad grandes y la reproducción no panmíctica se miden desde la Demografía Genética a través del Coeficiente de Aislamiento Reproductivo y del Coeficiente de Endogamia. Ambos parámetros incluyen la consideración de Número Efectivo de indivivuos que dejan descendencia y Número Efectivo de migrantes (en el que se incluyen sólo aquellos que dejan descendencia) (Magalhaes y Arce Gómez, 1987a).
Estos parámetros se calcularon en nueve poblaciones del NOA, cinco de la Puna, dos del Valle de Lerma y dos del Valle Calchaquí (Tabla 2).
Se considera una población estructurada a una población subdividida que está sujeta a efectos de Deriva. F como coeficiente de endogamia o como índice de fijación, tiene como función primaria la de proveer un parámetro en la especificación de la estructura poblacional.
Wright (1978) formula un modelo en el que no se asume nada con respecto al grado de aislamiento de las subdivisiones (S) de la población total (T) o su arreglo en el espacio. Pueden estar completamente aisladas en un extremo o ser limitadas arbitrariamente de un continuo.
Con el objeto de estimar la acción de la Deriva, se calcularon los Índices de Fijación de Wright FIS, FST y FIT a partir de la variabilidad genotípica en ocho loci de grupos sanguíneos (ABO, Dd, Cc, Ee, MN, Ss, Di, P) en tres poblaciones de la Puna, once del Chaco, una de Valles Calchaquíes, uno de HLA (DRB1) en una muestra de Cobres y doce STR-s (D3S1358, VWA, FGA, D8S1179, D21S11, D18S51, D5S818, D13S317, D7S820, D4S243, D18S535, HUMTHO1) en una muestra de Cobres y una de San Antonio de los Cobres. En materia de frecuencias génicas, los resultados de la endogamia y la Deriva son los mismos, de ahí que el Coeficiente Promedio de Endogamia de una población (F), sea una medida del proceso dispersivo (Cavalli-Sforza y Bodmer, 1981; Doolittle, 1986).
Se hizo un análisis jerárquico incluyendo las categorías intermedias Zona y Etnia. En el caso de Zona, se trata de estudiar el grado de diferenciación entre la Puna y el Chaco, cuyas características son definidamente diferentes, tanto en el aspecto ambiental como en el cultural.
A partir de las matrices de frecuencias alélicas, se estimaron los coeficientes de distancias genéticas de acuerdo con Nei (1987). Las distancias genéticas así obtenidas, fueron analizadas y posteriormente incluídas en la construcción de un fenograma de los grupos de estudio, mediante agrupaciones secuenciales, aglomerativas, jerárquicas y anidadas.
RESULTADOS Y DISCUSIÓN
El análisis de numerosos sistemas con determinación genética ha permitido estimar que en el hombre, alrededor del 28% de los loci son polimórficos, con una Heterozigosis Media del 6,7% (Dobzhansky et al, 1983). Sin embargo, deben suponerse valores superiores, ya que se considera que las técnicas más sensibles de la electroforesis con las que se calculan estos valores, sólo detectan un tercio de la variabilidad real (Lewontin, 1979).
De los 9 loci de grupos sanguíneos estudiados en las tres poblaciones de la Puna, 3 son monomórficos en Santa Rosa y Tolar Grande (Dd, Diego y Kell) y en Cobres únicamente Dd. En el resto de las poblaciones del NOA, uno o más de uno son monomórficos.
P tiene igual valor en Santa Rosa y Tolar Grande P=0,667 y más elevado en Cobres P=0,889. Sin embargo Cobres muestra semejante H que Tolar Grande (0,1858 ± 0,0563; 0,1895 ± 0,0640 respectivamente) y H es menor para Santa Rosa (0,136 ± 0,050), contribuyendo en forma diferencial los diferentes loci en cada población. Los valores más elevados de H se encuentran entre los Chiriguanos y los Chanés (0,348 ± 0,049 y 0,362 ± 0,06. Se observan diferencias en la h de los diferentes loci, contribuyendo cada uno de manera diferente a H.
Comparando estos resultados con otras poblaciones del NOA, observamos que las tres localidades puneñas son las que presentan menor variabilidad medida como H.
Las diferencias manifestadas con ambos tipos de medidas de la variabilidad se deben a que la proporción de loci polimórficos no es sensible a la frecuencia de individuos heterocigotos sino que destaca la presencia de uno o más alelos en un locus a una frecuencia apreciable (entre 0,01 y 0,99) mientras que H mide la variabilidad intrapoblacional a partir de las frecuencias de los genotipos.
En las localidades analizadas, el porcentaje de migrantes de corta distancia es mayor en todos los casos que el de larga. Sin embargo, en las poblaciones Puneñas y de los Valles Calchaquíes, se observa una diferencia entre ambos tipos de migraciones mucho mayor que en las localidades del Valle de Lerma en las que no se registran diferencias notables.
Para las localidades de la Puna, se observa mayor número de migrantes femeninos de corta distancia, lo que sugiere la posibilidad de que se fije el domicilio familiar en alguna medida en función de la residencia del varón. Esta situación se invierte cuando se consideran los migrantes de larga distancia, pero con muy pequeñas diferencias.
La tasa de migración efectiva (me) presenta valores muy variables, que oscilan entre 0,12 en Cobres y 0,77 en Tolar Grande.

Tabla 3: Tasa de migración efectiva y porcentanjes de migrantes
Tradicionalmente se consideró que las poblaciones aborígenes americanas eran exclusivamente de grupo sanguíneo O. Si bien es cierto que se encuentra a frecuencias muy elevadas, se han detectado también en el continente, los alelos A y B a una frecuencia promedio de 0,016 y 0,004 respectivamente (Caruso, 1995). Entre 43 poblaciones de Perú, Bolivia, Noroeste y Patagonia de Argentina, el grupo O es el más frecuente pero sólo está fijado en 10 de ellas. Salaberry et al (1999) en tejidos momificados del sitio Las Pirguas ratificaron la presencia del antígeno A y la ausencia del O.
Al analizar otros sistemas, también es notable la frecuencia a la que se presentan algunos alelos, hasta hace algunas décadas considerados marcadores americanos. En el sistema Diego, si bien el alelo Dia es característico de poblaciones americanas, Dib es el más frecuente según datos de las poblaciones estudiadas. El locus Dd del sistema Rh-hr presenta al alelo D fijado en 35 de 39 poblaciones estudiadas. Kell-Cellano, al igual que Dd y Diego, presenta un único locus k; entre las poblaciones del NOA, K se encuentra a frecuencias muy bajas. Tanto el alelo K como el d se consideran característicos de poblaciones no americanas. Con respecto al otro sistema del que se tiene información, el Duffy, el alelo a es el que se presenta a mayor frecuencia (35 de 39 poblaciones estudiadas).
Generalmente se adujo que la presencia de determinados alelos no americanos detectados por diferentes técnicas serológicas en el continente eran consecuencia de la incidencia de factores contaminantes, no obstante, en la última década, estudios del sistema ABO a nivel molecular confirmaron la presencia de los tres antígenos (Salaberry et al, 1999).
Frente a estos resultados, si bien el sistema ABO es el más estudiado y existe un importante número de investigaciones paleoserológicas realizadas en restos sudamericanos que detectaron la presencia de los tres antígenos, sería necesario reconsiderar antiguas posiciones respecto a la presencia y/o fijación no sólo del alelo O sino la de otros alelos dentro de las poblaciones precolombinas como característicos de ellas.
Estudios a nivel molecular que incluyen marcadores característicos tanto del mtDNA como del cromosoma Y postulan la contribución europea y africana al pool génico de varias poblaciones amerindias como resultado de Flujo Génico (Dipierri et al, 1999). Si bien la presencia de determinados marcadores ha sido interpretada como consecuencia de migraciones no americanas al continente, es imprescindible considerar y evaluar la acción de otros factores en las poblaciones.
El Flujo Génico es uno de los factores evolutivos de gran peso en los procesos microevolutivos de la región ya que existe suficiente evidencia de intercambio desde hace varios milenios. Por lo tanto desde el punto de vista estrictamente genético-poblacional es necesario un replanteo metodológico que permita analizar la variabilidad sin partir de supuestos que hasta el momento no han sido suficientemente validados.
Estudios realizados por Acreche et al (1997), Acreche et al (1999), Acreche y Albeza (1998), Albeza (1995) Caruso et al (1999) muestran que la Selección Natural estaría operando sobre todo sobre la componente de fecundidad.

Tabla 4: Indice de Oportunidad para la Selección
El ambiente para las poblaciones humanas no suele ser un factor limitante, sin embargo éste debe tener algo de peso cuando las condiciones son extremas, como en los ambientes de altura donde la radiación, la temperatura y la hipoxia permiten suponer que es posible que a través de procesos evolutivos las poblaciones humanas allí existentes hayan desarrollado estrategias adaptativas.
Según Salzano y Callegari-Jacques (1988), las poblaciones de altura podrían diferenciarse de aquellas de zonas bajas por ciertas características morfológicas, fisiológicas y bioquímicas, por acción de la Selección Natural.
Sobre las características que presentan estas poblaciones relacionadas con la adaptación a las condiciones de altura, hay pocos estudios de anatomía y fisiología (Brooke Thomas, 1977; Frisancho, 1996; Paulotti, 1949; Salzano y Callegari-Jacques, 1988; Rothhammer y Schull, 1980), siendo difícil determinar el rol que juegan los factores culturales y biológicos (genéticos, fisiológicos y de desarrollo) y por lo tanto, obtener conclusiones sobre la acción de la Selección Natural, dado que los caracteres analizados son poligénicos y a pesar de su alta heredabilidad están sujetos a influencias ambientales.
Con respecto a la hipoxia, es difícil modificarla a través de respuestas comportamentales o culturales, por lo que se deben generar respuestas fisiológicas para superar el stress provocado por esta condición. Rothhammer y Schull (1980) hallaron individuos portadores de variantes enzimáticas asociadas al ciclo glicolítico y la vía de las pentosas, que podrían constituir alternativas más eficientes para metabolizar glucosa en ambientes hipóxicos.
Estudios realizados por Frisancho (1996) en habitantes de zonas altas de Perú, indican que tienen presión arterial pulmonar más alta que los nativos de zonas ubicadas a nivel del mar, lo que permite optimizar el proceso de hematosis. Aún no se ha dilucidado si estos mecanismos de acomodación fisiológica son o no adaptativos en el sentido genético.
Con respecto al metabolismo intermedio en eritrocitos de Aymaras de zonas altas se observa que éste presenta varios cambios, que llevan a un significativo aumento de metahemoglobina; estas alteraciones ocurren también en personas del mismo grupo étnico que residen en zonas bajas. Este y otros estudios hematológicos indicarían que se trata de procesos determinados genéticamente más que de acomodación fisiológica (Salzano y Callegari-Jacques, 1988).
Las respuestas asociadas al stress provocado por el frío son más complejas que aquellas provocadas por las altas temperaturas y requieren la sincronización de la actividad cardiovascular con la del sistema circulatorio y lo más importante, la activación de procesos metabólicos. Frisancho (1996) ha realizado estudios en diversas poblaciones sobre las adaptaciones fisiológicas y culturales al frío. Concluye que en algunas poblaciones que llevan habitando zonas frías desde hace al menos 6.000 años, como Esquimales y Algonkians, las respuestas estarían mediadas por factores genéticos.
Dulout et al (1996) y Vahter et al (1995) han realizado estudios clínicos, bioquímicos y citogenéticos a habitantes de San Antonio de los Cobres e informan de la posible existencia de un polimorfismo genético en el control de la actividad de la metiltransferasa involucrada en la metilación del arsénico, requerida para convertir el arsénico inorgánico en MMA (ácido metilarsénico) y en DMA (ácido dimetilarsénico). Este polimorfismo sería una respuesta adaptativa a la presencia de altos niveles de este elemento carcinógeno en las aguas de la zona.
A pesar de la presencia de arsénico en el agua y los alimentos ingeridos por estos pobladores, no se han detectado síntomas de hidroarsenisismo. Además excretan arsénico principalmente en su forma inorgánica o como DMA y con bajos niveles de MMA a diferencia de individuos de otras poblaciones que también utilizan aguas contaminadas.
Otro polimorfismo asociado a adaptación a ambientes hipóxicos y fríos es la presencia de bandas de menor tamaño de Heterocromatina Constitutiva. De acuerdo con la hipótesis de Ibraimov (1993) la menor cantidad de heterocromatina implica menor gasto energético en la duplicación del ADN, lo que representaría una ventaja selectiva. Broglia (1998), Broglia et al (1999), han realizado entre otros, un análisis cuantitativo de polimorfismos de Heterocromatina Constitutiva en el que se observa que son significativamente menores las medias de las longitudes de las bandas C de pobladores de las regiones de la Puna que las de los del Chaco. Siendo posible explicar las diferencias de tamaño de las bandas C entre las dos regiones como resultado de la acción selectiva.
A partir de estos datos y de los valores de Indices de Oportunidad para la Selección puede interpretarse que los procesos selectivos han configurado en alguna medida la estructura genética de estas poblaciones.

Tabla 5: Parámetros demográficos de Aislamiento Reproductivo.
Ne: Número efectivo poblacional
me: Migración efectiva
CAR: Coeficiente de Aislamiento Reproductivo
CE: Coeficiente de Endogamia
En la mayoría de los casos, las características de la población evidenciadas a través del Coeficiente de Aislamiento Reproductivo (pequeña y relativamente aislada) confirman que la Deriva Génica es el principal factor que está actuando.
El Coeficiente de Aislamiento Reproductivo señala muy fuerte sujeción a deriva en Cobres, Santa Rosa de los Pastos Grandes, Olacapato y Chañarcito, en la Puna y en La Alumbrera en Catamarca, moderada sujeción en Pichao en el Valle Calchaquí, San Agustín en el Valle de Lerma y Tolar Grande en la Puna y ninguna en Chicoana, Valle de Lerma.
El Coeficiente de Endogamia, F, estimado a partir de la migración efectiva (me) y el tamaño efectivo de población (Ne) es muy cercano a 0 en Tolar, Chicoana y La Alumbrera con valores máximos en Pastos Grandes, Cobres, Olacapato y Chañarcito entre las poblaciones estudiadas y concuerda con lo propuesto por Cavalli-Sforza y Bodmer (1981) para poblaciones aisladas de tamaño pequeño.
En el caso de Chile, la situación es conocida a través de estudios como el del Valle de Elqui (Zúñiga, 1980), en el que se pudo comprobar un gradiente geográfico de endocruzamiento medido a partir de registros de dispensas otorgadas por la iglesia católica para realizar uniones consanguíneas. Ese gradiente revela mayores índices de endogamia en la montaña que en el valle y en esta última región que en la costa. Asimismo, se registran mayores coeficientes de endogamia en el medio rural que en el urbano y mayores tasas de consanguinidad en medios de menor densidad, coincidente con los resultados aquí consignados.
A pesar de no haberse detectado explícitamente uniones consanguíneas, los resultados obtenidos en el presente trabajo, señalan que los reducidos efectivos poblacionales tienen como consecuencia una alta tasa de endogamia.
En términos poblacionales, Wright (1978) llamó la atención sobre la posible importancia evolutiva derivada de la subdivisión de una población en grupos parcialmente aislados (cada uno endogámico pero sujeto a cruzamientos con los otros por migraciones), sugiriendo que la subdivisión puede acelerar el proceso evolutivo:
Favoreciendo la adaptación a nichos ecológicos locales.
Incrementando la variabilidad genética al permitir el establecimiento de genotipos con distinta adecuación darwiniana en diferentes nichos.
Favoreciendo nuevas combinaciones de genes.
En una población panmíctica, no hay correlación entre los genes homólogos de las gametas que se unen con sus respectivas frecuencias en la población total. Si hay una subdivisión entre líneas que se reproducen entre sí, se puede esperar una cierta correlación. Esto sugiere la descripción de la estructura de la población y de los efectos del endocruzamiento por medio de las correlaciones.
Los valores de F obtenidos, indican en la población de Santa Rosa en los loci MN, Ss y Ee efectos de Deriva y/o endogamia (F altamente significativo), mientras que los loci ABO y Cc, no muestran diferencias significativas de cero. En la localidad de Tolar Grande, sólo el locus Ss presenta valores significativos de F, mientras que para Cobres tales efectos son evidentes en los loci Cc, Ee y Ss.
En el locus ABO, se obtuvieron F-s no significativamente diferentes de 0 y negativos en las tres localidades para los tres alelos, reflejando un exceso de heterocigotos que sabemos obedece a Flujo Génico reciente por la información biodemográfica de que disponemos.
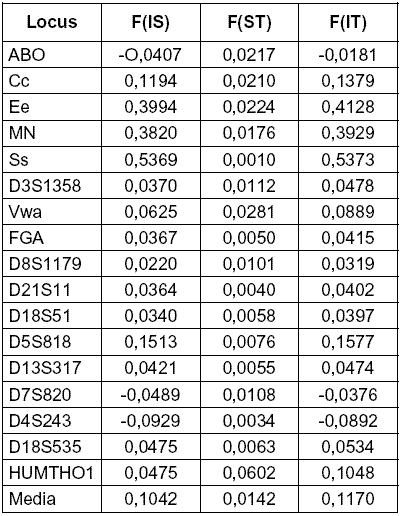
Tabla 6: Resumen de F-s en todos los loci
El valor promedio de FST que indica la relación entre subpoblaciones, en los doce STR-s es de 0,013 y de 0,022 considerando conjuntamente los STR-s y los cinco loci de grupos sanguíneos. El promedio general, 0,0142 muestra el bajo nivel de diferenciación entre las poblaciones.
Según Harpending (1995), para la mayoría de los marcadores genéticos neutros, el valor de FST entre grupos regionales humanos está alrededor del 10 %. El estadístico equivalente computado a partir de DNA mitocondrial es el mismo. Aún computado de grandes conjuntos de datos de medidas de cráneos es 10 %. Por otra parte, el valor estimado para el color de la piel es seis veces mayor, lo que lleva a la obvia conclusión de que este carácter está sujeto a selección local. IQ está entre el 10 común a las características neutras y el 60 % del color de piel.

Tabla 7: Estadísticos F-s calculados mediante análisis jerárquico, 1 categoría, 14 poblaciones.
La categoría Zona muestra menor grado de diferenciación. Las contribuciones al índice pasan en gran medida por la diferenciación entre poblaciones. Los loci Pp y Ee son los que más contribuyen, aunque presentan F-s negativos para la contribución Zona-Total indicando posible exceso de heterocigotos.

Tabla 8: Estadísticos F calculados mediante análisis jerárquico, 2 categorías.
La categoría Etnia se estudia con el objeto de agrupar la información sobre los mismos grupos del Chaco en diferentes localizaciones por un lado y los de la Puna por el otro. Se agruparon de esta manera las tres poblaciones de la Puna junto a los grupos: Mataco (Pagés Larraya et al, 1978), Chorote (Matson et al, 1969; Pagés Larraya et al, 1978), Chulupi (Matson et al, 1969; Pagés Larraya et al, 1978) Toba (Matson et al, 1969; Pagés Larraya et al, 1978), Pilagá (Pagés Larraya et al, 1978), Mocoví (Pagés Larraya et al, 1978), Chiriguano (Matson et al, 1969), Chané (Matson et al, 1969).
Incluyendo la categoría intermedia Etnia en el análisis jerárquico, se obtienen resultados semejantes a los observados en el análisis anterior con sólo una categoría intermedia, indicando la escasa contribución de Etnia a la diferenciación. Esto indica que la diferenciación es en gran medida debida a las diferencias locales entre poblaciones. Se puede concluir por lo tanto, escasa estructuración en las poblaciones estudiadas.
La agrupación de poblaciones obtenida corrobora los resultados de escasa estructuración y baja diferenciación por Zona y Etnia.
ARBOL DE CONSENSO:
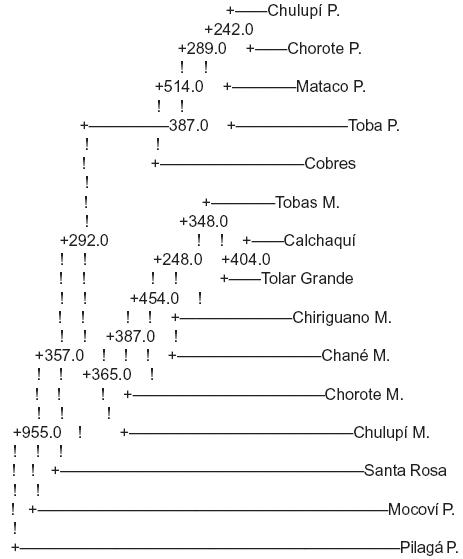
P. Pagés Larraya et al, 1978
M. Matson et al, 1969
Se realizaron mil repeticiones obteniendo un árbol de consenso (Biosys 2, Phillyp) en el que se observa que las poblaciones se agrupan por proximidad geográfica y no por Zona o grupo étnico.
CONCLUSIONES
· El complejo conjunto de características físicas de la región NOA configura el patrón de distribución de las poblaciones humanas, determinando a su vez las relaciones que se establecen entre éstas.
· En las poblaciones estudiadas la variabilidad, medida como Heterozigosis Media, es relativamente baja, siendo las poblaciones puneñas las que muestran menores valores.
· En ambientes extremos, se evidencia la acción de la Selección Natural, evaluada a través de Heterocromatina Constitutiva y del Indice de Oportunidad para la Selección, en el que incide con mayor fuerza el componente de fecundidad.
· Tanto los Coeficientes de Aislamiento Reproductivo como los Indices de Fijación indican aislamiento y sujeción a la Deriva aunque baja estructuración.
· Si bien se observa la acción de Selección Natural y Deriva Génica, la alta y sostenida incidencia del Flujo Génico tiene como consecuencia la baja diferenciación genética de las poblaciones estudiadas.
· El análisis jerárquico indica baja diferenciación tanto entre zonas como entre grupos étnicos, confirmado esto mediante un fenograma de distancias genéticas.
· La acción de la Selección Natural y la Deriva Génica explica la baja variabilidad observada, mientras que la acción del Flujo Génico, la falta de estructuración entre las poblaciones.
AGRADECIMIENTOS
Agradecemos la desinteresada colaboración de todas aquellas personas de las diferentes localidades de la Puna, Valle de Lerma y Valle de Santa María de la provincia que generosamente contribuyeron a la realización de este trabajo.
BIBLIOGRAFÍA
1. ACRECHE, N. (1994) La Evolución del Hombre. Notas de Cátedra. C.E.F.I.Sa, Salta. [ Links ]
2. ACRECHE N y ALBEZA, MV. (2001) El Pichao: Población Actual - Parámetros Biodemográficos. En: Investigations at El Pichao. Introduction to studies in the Santa Maria Valley, North-Western Argentina. BAR International Series 978, Oxford. pp: 195 - 203. [ Links ]
3. ACRECHE, N, ALBEZA, MV y CARUSO, GB. (1997) San Agustín (Salta): Estructura Poblacional, Deriva y Selección. Cuadernos de Humanidades Nº 9, Fac. de Humanidades, Universidad Nacional de Salta. [ Links ]
4. ACRECHE, N, BROGLIA, V, CARUSO, G y ALBEZA, MV. (1999) Chicoana (Salta): Genética y Población. Rev. Arg. de Antropología Biológica 2(1):257-266. [ Links ]
5. ACRECHE, N, CARUSO, GB y ALBEZA, MV. (1996) Distancias genéticas en poblaciones del NOA. Rev. de la Sociedad Arg. de Antropología Biológica 1(1):139-152. [ Links ]
6. ALBEZA, MV. (1995) Deriva Génica: Estructura Poblacional en Poblaciones Andinas. Tesis Profesional. UNSa. FCN. [ Links ]
7. ALBEZA MV, PICORNELL, A, ACRECHE, N, TOMÀS MAS, C, CASTRO OCÓN, JA and RAMÓN, MM. (2002) Genetic Variability at 14 STR loci in the Puna population of north western Argentina. Int J Legal Med 116: 126-132. [ Links ]
8. AZOFEIFA, J y BARRANTES, R. (1991) Genetic variation in the Bribri and Cabecar Amerindians from Talamanca, Costa Rica. Rev. Biol. Trop., 39 (2):249-253. [ Links ]
9. BAUMGARTNER, A y COZZI, EA. (1998) Planificación Territorial del Valle de Lerma (Provincia de Salta) orientada al sector agropecuario, mediante el uso de Sistemas de Información Geográfica. Alternativas de Inserción en el nuevo orden económico transnacional. Tesis de Licenciatura en Recursos Naturales, Facultad de Ciencias Naturales, Universidad Nacional de Salta. [ Links ]
10. BERTRANPETIT, J y CAVALLI-SFORZA, LL. (1991) A genetic reconstruction of the history of the population of the Iberian Peninsula. Am. J. Hum. Genet., 55: 51-67. [ Links ]
11. BERTONCELLO, R y GARCÍA, P. (1995) Geografía Argentina. Ed. Santillana. [ Links ]
12. BOLEDA, M. (1993) La Población del Noroeste Argentino. Historia y Actualidad. Ed. Legasa / Gredes. Argentina. [ Links ]
13. BROGLIA, V. (1998) Variabilidad Genética Poblacional de Heterocromatina Constitutiva en ambientes de altura. Tesis Profesional. Facultad de Ciencias Naturales. UNSa. Salta. [ Links ]
14. BROGLIA, V, de la FUENTE, S, ALBEZA, MV, CARUSO, G y ACRECHE, N. (1999) Heterocromatina constitutiva en ambientes de altura. IV Jornadas Nacionales de Antropología Biológica. S. S. de Jujuy. [ Links ]
15. BROOKE - THOMAS, RB. (1977) Adaptación Humana y ecología de la Puna. Pastores de Puna. Uywamichiq punarunakuna. (3): 87-111. IEP Ediciones. Lima. Perú. [ Links ]
16. CABRERA, AL. (1971) Fitogeografía de la República Argentina. Boletín de la Sociedad Argentina de Botánica, Vol. XIV, (1-2). [ Links ]
17. CARUSO, G. (1995) Deriva Génica y Polimorfismos Hematológicos en Poblaciones Andinas. Tesis Profesional, U.N.Sa, F. C. N. [ Links ]
18. CARUSO, G, ALBEZA, MV, ACRECHE, N y BROGLIA, V. (1999) Grupos sanguíneos y demografía en localidades puneñas de la Provincia de Salta. Rev. Arg. de Antropología biológica 2(1): 243-256. [ Links ]
19. CAVALLI-SFORZA, LL. (1969) Deriva genética en una población italiana. Scientific American, 3: 26-33. [ Links ]
20. CAVALLI-SFORZA, LL y BODMER, WF. (1981) Genética de las Poblaciones Humanas. Ed. Omega. Barcelona. [ Links ]
21. CROSS, H. (1976) Population studies and the old Order Amish. Nature, 262: 17-20. [ Links ]
22. DOBZHANSKY, T, AYALA, FJ, STEBBINS, GL y VALENTINE, JW. (1983) Evolución. Ed. Omega, Barcelona. [ Links ]
23. DOOLITTLE, DP. (1986) Population Genetics. Basic Principles. Advanced Series in Agricultural Sciences. Vol. 16. [ Links ]
24. DULOUT, FC, GRILLO, A, SEOANE, A, MADERNA, C, NILSSON, R, VAHTER, M, DARROUDI, F and NATARAJAN, A. (1996) Chromosomal aberrations in peripherial blood lymphocytes from native Andean women and children from Northwestern Argentina exposed to arsenic in drinking water. Mutation Research. Genetic Toxicology Testing, 370: 151-158. [ Links ]
25. GLASS, BH. (1953) Los Dunkers: un caso de Deriva Genética. Scientific American. [ Links ]
26. FELSENSTEIN, J. (1993) Phylip (Philogeny inference package) Version 3.5c. Distributed by the author, Department of genetics, Washington University Seattle. [ Links ]
27. FRISANCHO, AR. (1996) Human Adaptation and Accommodation. Enlarged and revised edition of Human Adaptation. De University of Michigan. USA. [ Links ]
28. HARPENDING, H. (1995) Human Biological Diversity. Evolutionary Anthropology, 4(3): 99-103. [ Links ]
29. BRAIMOV, AI. (1993) The origin of modern humans: A cytogenetic model. Human Evolution. 8 (2): 81-91. [ Links ]
30. INDEC. (1991) Censo Nacional de Población y Vivienda. Ministerio de Economía y Obras y Servicios Públicos. Secretaría de Programación Económica. Serie B, N 17. Salta. [ Links ]
31. KARLIN, UO, CATALÁN, L y COIRINI, R. (1994) La naturaleza y el hombre del Chaco. Proyecto GTZ. [ Links ]
32. KRAPOVICKAS, P. (1983) Las poblaciones indígenas históricas del Sector Oriental de la Puna. Relaciones, XV, Nueva Serie: 7-24. [ Links ]
33. LEWONTIN, R. (1979) Las Bases Genéticas de la Evolución. Ed. Omega. [ Links ]
34. MAGALHAES, JCM & ARCE-GOMEZ, B. (1987 a) Study on a brazilian isolate. I. Population structure and random genetic drift. Hum. Hered., 37: 278-284. [ Links ]
35. MAGALHAES, JCM & ARCE-GOMEZ, B. (1987 b) Study on a brazilian isolate. II. Opportunity for selection. Hum. Hered., 37: 381-383. [ Links ]
36. MARTINEZ SARASOLA, C. (1992) Nuestros Paisanos los Indios. Vida, Historia y Destino de las Comunidades Indígenas de la Argentina. Emecé. Buenos Aires. [ Links ]
37. MATSON, GA, SUTTON, HE, SWANSON, J and ROBINSON, A. (1969) Distribution of hereditary blood groups among indians in South America. VII: In Argentina. Am. J. Phys. Anthrop., 30: 61-84. [ Links ]
38. NEI, M. (1987) Molecular Evolutionary Genetics. Library of Congress Cataloging. USA. [ Links ]
39. PAGES LARRAYA, F, WYLLER de CONTARDI, N y SERVY, E. (1978) Marcadores genéticos de la población aborigen del Chaco Argentino. Universidad Nacional de Córdoba, Rev. del Inst. de Antropología, Tomo VI: 217-241. [ Links ]
40. PAULOTTI, O. (1949) Los Nativos de la Puna de Jujuy. Rev. Inst. Antrop. Vol 4: 5-83. UNJ. [ Links ]
41. PONTUSI, EP. (1993) Geografía del Noroeste Argentino. Fac. de Ciencias Naturales, Universidad Nacional de Salta. [ Links ]
42. ROTHHAMMER, F y SCHULL, WJ. (1980) Adaptación genética de poblaciones andinas. Actas IV Congr. Latinoam. Genética, Vol. 2: 357-368. [ Links ]
43. SALABERRY, MT; DEJEAN; CB, GOICOECHEA; AS, BAFFI, EI y CARNESE, FR. (1999) Grupos sanguíneos en restos momificados de una población prehispánica: Las Pirguas (Salta, Argentina). Rev. De Antropología Biológica 2(1): 267-280. [ Links ]
44. SALZANO, F & CALLEGARI-JACQUES, SM. (1988) South American Indians. A Case Study in Evolution. Clarendon Press. Oxford. [ Links ]
45. SWOFFORD and SELANDER. (1997) Biosys2 Department of Genetics and Development. University of Illinois at Urbana-Champaigne. [ Links ]
46. VAHTER, MG, CONCHA, G, NERMELL, B, NILSSON, R, DULOUT, F and NATARAJAN, A. (1995) A unique metabolism of inorganic arsenic in native Andean women. European Journal of Pharmacology. Environmental Toxicology and Pharmacology Section, 293: 455-462. [ Links ]
47. WRIGHT, S. (1978) Evolution and the Genetics of Populations, Vol. 2. The Theory of Gene Frequencies. Chicago University Press. [ Links ]
48. WRIGHT, S. (1943) Isolation by distance. Genetics, 28: 114-138. [ Links ]
49. ZUÑIGA, IJ. (1980) La Consanguinidad en el Valle de Elqui. Un estudio de Genética de Poblaciones Humanas. Ed. de la Universidad de Chile, La Serena. [ Links ]

