










Serviços Personalizados
Journal

Artigo
 Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares em
SciELO
Similares em
SciELO
Compartilhar

 Permalink
PermalinkCuadernos de la Facultad de Humanidades y Ciencias Sociales. Universidad Nacional de Jujuy
versão On-line ISSN 1668-8104
Cuad. Fac. Humanid. Cienc. Soc., Univ. Nac. Jujuy no.43 San Salvador de Jujuy jun. 2013
ARTÍCULO ORIGINAL
Madres wichí: Cuando ser y cuantos niños tener
(Wichi mothers: When to be and how many children to have)
Marta Graciela Méndez* y Stella Ofelia Ferrarini**
* CONICET.
** Facultad de Ciencias Naturales y Museo. Universidad Nacional de La Plata. 122 y 60 s/nº - CP 1900 - La Plata - Buenos Aires - Argentina. Correo Electrónico: gmendez@fcnym.unlp.edu.ar
RESUMEN
Las estrategias reproductivas están enmarcadas por las limitaciones ecológicas específicas ante las que cada grupo humano se enfrenta. Los patrones naturales de la reproducción de los Wichí se exploran tomando en consideración la energía disponible que subyace a su ancestral patrón de cazadores-recolectores. En la pubertad, cuando se frena la inversión en crecimiento, las mujeres comienzan la inversión en reproducción. Afrontan, entonces, una negociación entre continuar su crecimiento un tiempo más o comenzar su ciclo reproductivo. Una vez comenzada la etapa reproductiva el equilibrio debe establecerse entre la energía reservada a la reproducción y la dirigida a mantenerse y poder llegar a una nueva fase reproductiva. Se analizan las historias reproductivas de mujeres Wichí para indagar acerca de las decisiones sobre cuándo comienzan la reproducción y cuál es el tamaño de la progenie resultante. La edad media de la primera maternidad fue 18,23 años y la longitud media de los intervalos entre nacimientos de 3,2 años. Se demuestra que las madres que comenzaron antes su reproducción tuvieron historias reproductivas más rápidas. La fx arrojó un valor estimado de la reproducción individual de 7,5, valor que se discute y compara con el de otras poblaciones aborígenes chaqueñas y del mundo. La edad temprana a la primera maternidad de las mujeres Wichí y la tasa reproductiva alta con intervalos entre nacimientos de longitud moderada delinean el modelo de maximización de la distribución de energía destinada a la reproducción a lo largo del curso de sus vidas, en contextos socio-ecológicos de recursos limitados.
Palabras Clave: Edad al Primer Nacimiento; Estrategia Reproductiva; Fecundidad; Wichí.
ABSTRACT
Reproductive strategies are framed by the specific ecological limitations that each human group faces. Wichí natural patterns of reproduction are explored taking in consideration the available energy that underlies to their ancestral hunter-gatherers pattern. In the puberty, investment in growth declines and women begin the investment in reproduction. They confront a tradeoff among continuing their growth for a time or begin their reproductive cycle. Once the reproductive stage begun the tradeoff will be among allocated energy to reproduction or to maintenance and be able to arrive to a new reproductive phase. Wichí women’s reproductive histories are analyzed to investigate about the decisions on when they begin the reproduction and how many children they have. The mean age of the first maternity was 18.23 years and the mean length of intervals among 3.2 year-old births. The mothers that began their reproduction before had quicker reproductive histories. The fx threw a value of individual reproduction of 7.5, value that we discuss and compare with that of other populations. The early age at first birth of the Wichí women and the high reproductive rate with moderate birth intervals delineate a pattern of maximization of the energy distribution dedicated to the reproduction along the lifetime, in a ecological contexts of limited resources.
Key Words: Age to the First Birth; Fecundity, reproductive strategy, Wichí.
INTRODUCCIÓN
Tener hijos es un imperativo biológico para la supervivencia de cualquier especie, incluida la humana. Los niveles de fertilidad en los humanos varían entre las poblaciones, y aún dentro de las mismas, ya que existen interactuando componentes biológicos, culturales y ecológicos que intervienen en dicha diferenciación. Los perfiles demográficos de una población surgen de los planes reproductivos individuales condicionados por los factores antes mencionados, que actúan como modeladores de los dos elementos más importantes que conforman una vida: tiempo y energía (Kaplan y Gurven, 2008). En las sociedades en las que los recursos energéticos son limitados, y aún escasos en algunas épocas del año, la regulación temporal de los eventos reproductivos en las mujeres tenderá a maximizarlos para lograr la optimización de su éxito reproductivo (Ellison, 2003, Crognier, 2003).
La administración de estos recursos energéticos en el curso de una vida enfrenta dos negociaciones principales que afectan la selección natural sobre la fertilidad: 1) la reproducción presente versus la futura; y 2) la cantidad de descendientes versus su calidad (Kaplan y Lancaster, 2004, Vitzthum, 2008). En el primer caso, el individuo invierte sus tasas energéticas en el futuro para crecer e incrementar así su futura fertilidad como en las etapas infantil y juvenil, en las cuales la fecundidad es igual a cero y los beneficios energéticos se vuelcan completamente al crecimiento (Bogin, 1999, Kaplan et al., 2003). En la pubertad, cuando se frena la inversión en crecimiento comienzan la inversión en reproducción y las mujeres son el sexo que paga los costos directos más altos en ella. Afrontan, entonces, una negociación entre continuar su crecimiento un tiempo más o comenzar su ciclo reproductivo. Debido a que ambas funciones son energéticamente costosas, una vez que han comenzado la etapa reproductiva raramente tienen crecimientos adicionales. Cambiar del crecimiento a la reproducción en edades juveniles puede conferir beneficios en la eficacia al incrementar la cantidad de tiempo que las mujeres disponen para la reproducción (Käär y Jokela, 1998). Pero, retrasarla e invertir más energía en crecimiento puede también ser evolutivamente ventajoso al disminuir los riegos de mortalidad infantil y materna (Walker et al., 2006, Sear, 2007). En los humanos se ha demostrado que las edades tempranas a la primera reproducción están fuertemente favorecidas por la selección natural, como el componente más importante del fitness femenino tanto en las poblaciones históricas como en las contemporáneas (Kirk et al., 2001, Helle et al., 2005, Pettay et al., 2007).
Igualmente, una vez comenzada la etapa reproductiva el equilibrio debe establecerse entre la energía reservada a la reproducción y la dirigida a mantenerse y poder llegar a una nueva fase reproductiva. Por esta razón, la edad a la primera maternidad y la tasa reproductiva de las mujeres adultas delinearán el modelo de maximización de la distribución de energía destinada a la reproducción a lo largo del curso de la vida, en contextos socio-ecológicos determinados (Stearns y Koella, 1986, Hill y Hurtado, 1996).
En la historia reproductora de las mujeres, el intervalo fecundo es el período comprendido entre el primer y el último nacimiento. Este período se abre con el primer alumbramiento y la edad a la que este evento se produce influye sobre su longitud definitiva. En las poblaciones que no utilizan métodos contraconceptivos el final de este intervalo se funde con el del período fértil, causado por el cese definitivo de los ciclos menstruales. La menopausia es un parámetro biológico inexorable mientras el inicio de la vida reproductiva es más flexible y da lugar a elecciones que modelarán las estrategias individuales. Otro factor importante que contribuirá a conformar estas estrategias está dado por la frecuencia de los eventos reproductivos que determinará la utilización fecunda del período fértil y el número de hijos nacidos vivos que tendrá cada mujer (Livi-Bacci, 2011).
En sociedades como la Wichí, donde los recursos alimenticios son limitados, la regulación temporal tanto del primer nacimiento como de los sucesivos partos es un elemento crítico en términos de la supervivencia tanto de las madres como de sus descendientes y de ahí surge la importancia evolutiva de este indicador.
Numerosos trabajos realizados sobre sociedades que conservan en algún grado atributos tradicionales de cazadores-recolectores-pescadores han ofrecido distintas versiones acerca de las estrategias utilizadas para delinear su historia vital. Todas ellas son eficaces a la hora de la subsistencia enfrentando ambientes ecológicos disímiles y generalmente poco amigables (Blurton Jones et al., 1992, Yamauchi et al., 2000, Valeggia y Ellison, 2003, Sánchez Ocasio, 2003, Crognier, 2003, Gurven y Kaplan, 2006, Mendez y Ferrarini 2011, Ferrarini y Mendez, 2011, entre otros)
A su vez, las poblaciones con fertilidad natural, que no han experimentado una transición demográfica, proveen datos valiosos para investigar esquemas reproductivos desde una perspectiva evolutiva (Hill y Hurtado, 1996, Sear et al., 2000).
Los datos utilizados en este estudio integran un protocolo de investigación más amplio que intenta analizar la situación demográfica de poblaciones aborígenes chaqueñas, con un enfoque evolutivo y ecológico que centra su atención en la reproducción y la fertilidad, tomando en cuenta rasgos bioculturales. En este trabajo se exploran los patrones naturales de la reproducción de los Wichí tomando en consideración la energía disponible que subyace a su patrón ancestral de cazadores-recolectores. Las madres pueden acarrear sólo un niño por vez, entonces importa en qué momento se comienza la reproducción y la longitud de los intervalos entre nacimientos incluidos en el período fecundo, alcanzable en función de los límites energéticos. Se analizan las historias reproductivas de mujeres Wichí para indagar acerca de las decisiones sobre cuándo comienzan la reproducción y cual es el tamaño de la progenie resultante.
MATERIALES Y MÉTODOS
La población Wichí que aquí se analiza se asienta en una comunidad próxima al río Pilcomayo, en el municipio de Santa Victoria Este del departamento Rivadavia Banda Norte de la provincia de Salta. La comunidad Kanohis se fundó en el año 1985 con familias de la etnia Wichí provenientes en su mayoría de las comunidades Pozo El Tigre y La Paz. Habitan en ella 424 personas, distribuidas en 87 familias. Unas pocas de ellas pertenecen a la etnia Chorote, pero no serán tomadas en cuenta en este estudio. Disponen de servicios hospitalarios y escuela en el poblado más próximo, Santa Victoria Este, a unos 2,5 km.
Se relevaron las historias reproductoras de 45 mujeres con al menos un hijo. Se registraron las edades de las madres en el momento de la encuesta, al momento de cada nacimiento y el número de hijos nacidos vivos. Se calculó la edad al primer nacimiento, su estadística descriptiva y se analizó la distribución de frecuencias en grupos de edad. Se determinaron las edades de las madres en los distintos órdenes de descendencia y se compararon con las obtenidas en otras poblaciones de fertilidad natural y controlada. Se computaron las longitudes de los intervalos entre nacimientos y sus estadísticas descriptivas; la información se desglosó para cada orden de paridad con los respectivos valores estadísticos. Se estimó la variación de la fecundidad de las mujeres a lo largo de su vida reproductora. Se calculó el coeficiente de correlación (r) y el de determinación (R2) entre la edad al primer nacimiento y los intervalos intergenésicos medios. Las longitudes medias de los intervalos por paridad se analizaron en función de la edad al primer nacimiento. Se determinó la edad media de las madres y la distribución numérica por intervalos de edad. También se analizó la distribución porcentual de las madres según el número de hijos. Se calcularon las tasas específicas de fecundidad fx y se analizaron los patrones de fecundidad con las respectivas probabilidades en todas las edades y grupos de edad. A su vez, se estimaron las contribuciones de cada grupo de edad al valor de la tasa de fecundidad F y, por derivación, el tamaño medio de la descendencia que una mujer teóricamente tendrá.
RESULTADOS
La edad media de la primera maternidad en esta población fue 18,23 años (Tabla 1), que es temprana al compararla con las de otras poblaciones mundiales, incluida otra población aborigen chaqueña de diferente etnia (Chorote) que comparte el hábitat con la aquí estudiada (Tabla 2).

Tabla 1. Estadística descriptiva de la edad al primer nacimiento.
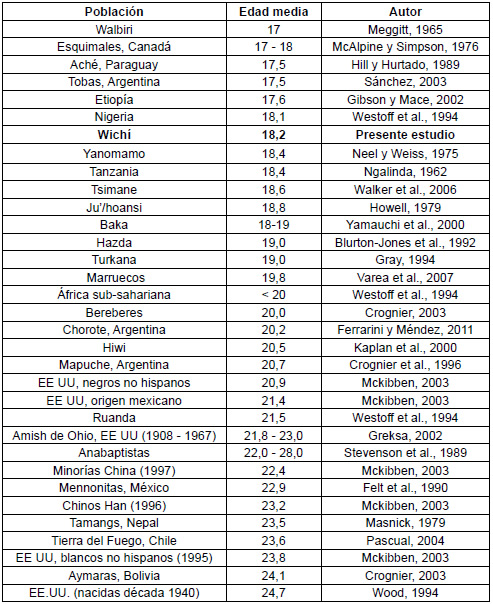
Tabla 2. Edad al primer nacimiento en distintas poblaciones mundiales.
En el análisis de la distribución de frecuencias de edades al primer nacimiento, considerando grupos de edad, se observa que el 75,6% de las madres tuvieron su primera maternidad antes de los 20 años. La Figura 1 expresa gráficamente este resultado así como explica los valores alcanzados por los estadísticos edad mediana y edad modal de la Tabla 1.

Figura 1. Distribución de frecuencias de la edad al primer nacimiento.
Al investigar las edades de las madres en los distintos ordenes de paridades fue posible indagar cómo se han regulado temporalmente los eventos reproductivos en ellas. En la Tabla 3 se presentan los descriptivos correspondientes. A medida que aumenta el orden de paridad, lógicamente aumenta la edad de las madres. Pero, como el comienzo se registra a edades tempranas, altas paridades como es el caso de la séptima, ocurre a edades relativamente tempranas (34,5 años) cuando a las mujeres aún les restan unos diez años de vida fecunda. La edad media de maternidad considerando todos los órdenes de paridad es de 30,9 años.

Tabla 3. Edad de la madre en los distintos órdenes de la descendencia (en años)

Figura 2. Edad de las madres en los distintos órdenes de paridad para la población Wichí y otras poblaciones.
En la Figura 2 se compara el comportamiento reproductivo de la población Wichí con dos poblaciones de fertilidad natural: Chorote (Ferrarini y Mendez, 2011) y Mapuche de Río Negro (Crognier et al., 1996) y una controlada: población chilena de Tierra del Fuego (Pascual, 2004). Se observa que el comienzo más temprano se da entre los Wichí y el más tardío entre los Fueguinos. Las pendientes son similares en todas hasta la paridad 3, desde allí las curvas difieren entre las poblaciones de fertilidad natural y la controlada. Esta última presenta una meseta con ascenso muy suave en tanto que las restantes continúan con la misma pendiente y con ligeras diferencias a partir de las paridades 7 y 8, correspondientes a los efectos de la infertilidad secundaria que comienza alrededor de los 35 años.
La distribución de los nacimientos según la edad de la madre en el momento del nacimiento aporta información que permite conocer cómo varía la fecundidad de las mujeres de una población a lo largo de su vida reproductora. La distribución de los nacimientos, según la edad de la madre en el momento del nacimiento, está representada en la Figura 3. La curva asume la típica forma de U invertida con un pico máximo en los grupos de edad 15-19 y 20-24 años. Se registra además una lenta y progresiva disminución hasta edades tardías.
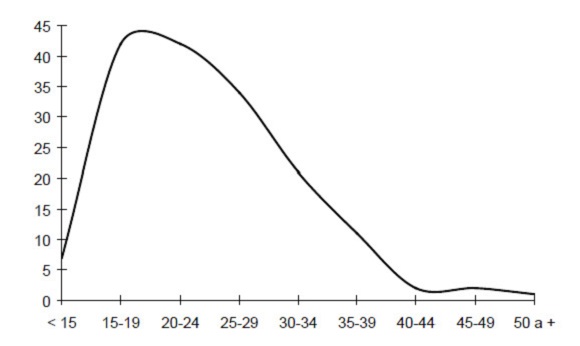
Figura 3. Distribución de los nacimientos según la edad de la madre al nacimiento.
En este punto interesa analizar las modalidades de desarrollo de las carreras reproductivas individuales. Para la muestra en general los intervalos entre nacimientos asumen un valor de 38,05 meses, es decir 3,2 años. No obstante existen diferencias en los valores de los estadísticos de posición tal como se presentan en la Tabla 4. Entonces, se evaluó si la edad al primer nacimiento tuvo alguna influencia sobre las longitudes de los intervalos y, por lo tanto, algún efecto sobre la cantidad de hijos que las madres Wichí tuvieron en su período fecundo. A este fin, se calculó el promedio de los intervalos para cada mujer tomada individualmente, más allá del número de hijos que tuvo, y se lo comparó con la edad al primer alumbramiento de esa mujer. Esta relación se testeó a través del coeficiente de correlación (r) que arrojó un valor de 0,69 demostrando que la edad en la que comienza la reproducción está relacionada positiva y marcadamente con la longitud de los intervalos que cada mujer tuvo. El coeficiente de determinación (R2) indica que la fracción de la variabilidad de una de las medidas depende en un 47% de la otra. Estos hallazgos concuerdan con los expuestos por Bongaarts y Potter (1983) que consideran a la edad materna al primer nacimiento como uno de los determinantes de la longitud de los intervalos intergenésicos. Así, en las poblaciones en las que se calculó la existencia de correlación entre ambos parámetros se encontró una relación positiva, poniendo en evidencia que las mujeres que tienen su primer hijo a edades más avanzadas tienden a tener intervalos más largos.

Tabla 4. Longitud media del intervalo entre nacimientos (en meses)
Para una cuantificación de los valores de las longitudes de los intervalos a medida que aumenta la edad al primer nacimiento se reagrupó a la muestra según tres categorías de edad de la madre al primer alumbramiento. El procedimiento utilizado se funda en la consideración de todos los intervalos de todas las madres agrupadas según la edad a la que tuvieron su primer hijo, a saber antes de los 18 años, entre 18,01 y 22 años y aquellas que lo hicieron después de los 22, sin tomarlas en forma individual. Los resultados se presentan en la Tabla 5 y muestran un aumento progresivo de más de seis meses entre las dos primeras categorías y de más de tres meses entre la segunda y la tercera. Estos resultados confirman lo hallado en el análisis de las madres tomadas en sus carreras reproductivas individualmente e indican que se trata de un comportamiento del grupo. Así las madres que comienzan antes tienen espacios intergenésicos más cortos y en consecuencia más hijos al final de su período fecundo.

Tabla 5. Longitud media de los intervalos entre nacimientos según la edad al primer nacimiento.
Ahora bien, las 45 madres estudiadas tuvieron un total de 162 hijos. Dado que no todas han completado su período fecundo, el resultado en un análisis transversal al momento de la toma de los datos es de 3,6 hijos por madre. La edad media de las madres es de 29,9 años; es decir, se trata de un conjunto que incluye muchas madres a quienes todavía les resta por cumplir tiempo para cerrar su intervalo fértil. Un análisis de la distribución de las madres por grupos de edad arroja que casi el 70% tiene menos de 35 años y el 30 % restante corresponde a todos los grupos de edad superiores. Esto se muestra en la Figura 4.
Al considerar la distribución de las madres según el número de hijos se tiene que casi el 70% tienen entre 1 y 4 hijos (Figura 5). Cabe consignar que las mujeres que tienen 7 o más hijos están próximas o superan los 40 años de edad, es decir cercanas al fin de su período reproductor.

Figura 4. Distribución de frecuencias porcentuales de las madres según grupos de edad.

Figura 5. Distribución porcentual de las madres según el número de hijos.
Por tratarse, entonces, de una población de mujeres que, en su mayoría, no ha completado su período reproductor se calculó la tasa de fecundidad F, que es una medida de fecundidad pura no influenciada ni por la estructura de la población ni por la mortalidad (Livi-Bacci, 2011). Esta tasa permite obtener una estimación del número esperado de descendientes de un grupo de mujeres si estas sobrevivieran hasta el final del período reproductor y estuvieran expuestas a las tasas de fecundidad general, específicas para cada edad (Campbell y Wood, 1988).
Los resultados del cálculo de las tasas de fecundidad a edad específica se muestran en la Tabla 6.

Tabla 6. Tasas de fecundidad a edad específica según la paridad (x 1000)
La tasa F de fecundidad resulta de la sumatoria de las tasas específicas para cada edad, según la expresión:
| β ω |
|
| F = ∑ ∑ o f x | 7483,2 |
| x=α о=1 |
|
|
|
|
Donde, x son las distintas edades de la madre al tener el recién nacido, desde la edad más temprana α a la más tardía β y o es el orden de nacimiento del hijo (desde 1 hasta el más elevado ω).
La tasa F estimada para esta población es de 7483,2 hijos por mil mujeres. De esta manera, la estima realizada a través de esta tasa es de 7,5 hijos por mujer si cada una de ellas siguiera con el mismo ritmo reproductivo hasta completar su período fértil.

Tabla 7. Tasas específicas de fecundidad (fx) por grupos de edad de las madres.
Cuando las tasas específicas de fecundidad (fx) se agrupan por períodos etarios y se relacionan con la del grupo 20-24 (fx / fx 20-24 * 100), para facilitar su comparación con las registradas para otras poblaciones, se observa que los grupos etarios 25-29 y 30-34 alcanzan los valores máximos, aunque próximos al valor de referencia. También se advierten valores elevados en el intervalo 15-19 y un descenso progresivo en los grupos de mayor edad adjudicables a factores biológicos como la declinación de la fertilidad, el aumento de abortos naturales y la menopausia. Al compararlo con los patrones observados en otras poblaciones de fertilidad natural y controlada la curva asume una forma convexa con valores comprendidos entre los de las poblaciones Amish (Greksa, 2002) y Hutteritas (Nonaka et al., 1994) de fecundidad natural y se diferencia claramente de la población testigo de fecundidad controlada (Pascual, 2004) (Figura 6).

Figura 6. Patrones de fecundidad por grupos de edad de poblaciones de fecundidad natural y controlada y de la población Wichí

La fila Fecundidad natural incluye la media de trece poblaciones de fecundidad natural (Henry, 1961).
Tabla 8. Número medio de hijos nacidos vivos (HNV) esperado.
Tomando en cuenta las tasas específicas de fecundidad, se estimó la contribución de cada grupo de edad al valor final de la tasa de fecundidad F y, por consiguiente, el tamaño teórico medio de la descendencia de una mujer si hubiera iniciado su reproducción a una edad determinada. De esta forma, si una mujer Wichí tuviera su primer hijo a los 20 años tendría, 6,35 hijos nacidos vivos por término medio al final de su período fecundo; es decir, reduciría su fecundidad teórica un 13,37% con relación a las que tuvieran su primer nacimiento a los 15 años (Tabla 8).
DISCUSIÓN Y CONCLUSIONES
El perfil reproductivo de las madres Wichí de la comunidad Kanohis, tomando varios parámetros y tasas relacionados con la fertilidad, permite comprender sus decisiones acerca de cuándo convertirse en madres y cuántos hijos tener. También brinda una excelente oportunidad para conocer las estrategias de mujeres de sociedades tradicionales de pequeña escala para conseguir el éxito reproductivo y la supervivencia en un marco de constreñimiento ecológico.
El comienzo de la maternidad sucede en promedio a los 18,23 años. Desde un punto de vista evolutivo, la edad al primer nacimiento es uno de los eventos que modelan las estrategias reproductivas de los grupos humanos. Dado que a lo largo de su historia vital las madres deben administrar los recursos que afectarán su desempeño reproductivo, las decisiones que tomen son especialmente cruciales en las mujeres en las cuales el embarazo, parto y amamantamiento requieren importantes inversiones tanto de tiempo como de energía. El acceso temprano a la gestación, el mantenimiento de la reproducción hasta etapas finales del período fértil y una intensa natalidad, son los principales factores que afectan la salud y la supervivencia de la madre y su hijo. Un gran número de estudios han relacionado la edad al primer nacimiento con la fertilidad, sobre todo en poblaciones con fertilidad natural (Bumpass et al., 1978, Gaisie, 1984, Konogolo, 1985, Westoff et al., 1994). Desde la perspectiva demográfica, varios trabajos han mostrado que las edades jóvenes al primer nacimiento tienden a estimular el número de niños que una mujer tendrá, en aquellas poblaciones que no utilizan métodos de control de la natalidad (Menken, 1980, Senderowitz y Paxman, 1985, Voydanoff y Donnelly, 1990, Wulf y Singh, 1991). Aún en las que la planificación familiar está ampliamente distribuida, la regulación temporal de los primeros nacimientos puede afectar el tamaño de la familia completada si la contraconcepción es utilizada para espaciar, pero no para limitar la fertilidad (Ngalinda, 1998).
La edad media a la primera maternidad Wichí es temprana comparada en un ranking de 33 poblaciones de distintos continentes y épocas. En general el grupo de sociedades cazadores recolectores se distribuyen en dos conjuntos: uno con edades de inicio muy tempranas (Walbiri, Esquimales, Aché, Toba, Tsimane, Ju’/hoansi y Baka) y otro con edades un poco más tardías (Hazda, Chorote y Hiwi). Los Toba, etnia que habita similares ambientes chaqueños y comparte con los Wichí numerosos aspectos de cultura material, social e ideológica, presentan una edad de 17, 5 años para el primer nacimiento (Sánchez Ocasio, 2003). Otro etno-grupo chaqueño, los Chorote, con similares relaciones ecológicas y culturales, manifiestan un comienzo dos años más tardío que los Wichí (Ferrarini y Mendez, 2011). Se dibujan así dos elecciones diferentes frente a la decisión de canalizar la energía: los Toba y los Wichí privilegiando la reproducción y los Chorote postergándola en favor de completar su crecimiento somático. Después de la menarca deberá establecerse un equilibrio entre la reproducción presente y la futura, es decir entre inversiones en el propio capital corpóreo versus inversiones inmediatas en reproducción (Kaplan y Lancaster, 2004). Las jóvenes Wichí, después de la menarca establecen relaciones sexuales libres hasta que se formaliza una pareja. También es común que, aún sin vínculo matrimonial, muchas jóvenes tengan niños que llevan el apellido materno y que se crían en el ámbito de la familia extensa, aunque todos conocen de quienes son hijos (Barúa et al., 2008). Este comienzo temprano del período fecundo pareciera ir en contra de la biología, ya que se produce durante la etapa de esterilidad adolescente cuando la probabilidad de concebir es baja debido a razones hormonales y anatómicas (Apter et al., 1978, Apter et al., 1987). Sin embargo, es una estrategia asumida por una porción importante de las mujeres, ya que el 75% de las madres Wichí tuvieron su primera maternidad antes de los 20 años. Para las mujeres Chorote, en cambio, posponer casi dos años el inicio de la vida reproductiva les ha permitido invertir en su propio crecimiento con la ventaja de que quienes alcanzaron estaturas más altas tuvieron mayor cantidad de hijos y también mayor número de sobrevivientes (Mendez y Ferrarini, 2009). Allal et al. (2004) y Sear et al. (2004) hallaron que una dilación de un año resultaba en un incremento entre 0,33 y 0,41cm en las mujeres en contextos rurales de Gambia y que los beneficios de postergar la primera reproducción se experimentaban a través de una menor mortalidad de los hijos de las madres más altas. Helle (2008) encuentra que por cada año de postergación de la primera maternidad las jóvenes finlandesas contemporáneas aumentan 0,20cm en su estatura adulta, sin embargo esta ganancia resulta irrelevante en términos del desempeño reproductivo. Varios estudios también han mostrado que las mujeres que tienen un comienzo tardío de su reproducción tienen mortalidades más bajas en períodos posteriores de la vida, comparadas con mujeres que comenzaron su reproducción más temprano (Koperlainen, 2000, Doblhammer, 2000, Smith et al., 2002, Grundy y Tomassini, 2005). Si bien los mecanismos por los cuales se producen estas relaciones aún no son claras, la mayor longevidad de las mujeres con nacimientos tardíos parece reflejar un mejor estado de salud y un lento envejecimiento (Sear, 2007).
Al igual que el comienzo de la reproducción, la intensidad puede también ser importante. Podría presumirse que un comienzo temprano conducirá a un espaciamiento más amplio entre los futuros nacimientos, de manera de permitir que las madres puedan recomponer su condición fenotípica antes de emprender otro embarazo. En la continuidad de las carreras reproductivas, el intervalo entre nacimientos promedio en las mujeres Wichí tiene una duración de 38,05 meses (3,2 años) valor moderado que permitiría la adquisición de energía suficiente para enfrentar un nuevo esfuerzo reproductivo. En realidad la intensidad del esfuerzo reproductivo ha sido menos estudiado que el esfuerzo acumulativo (paridades) o el esquema de la reproducción, pero varios autores han encontrado que las tasas de mortalidad se incrementan con cortos intervalos entre nacimientos (Grundy y Tomassini, 2005), el nacimiento de mellizos (Helle et al., 2004) y de hijos varones (Helle et al., 2002). Los tres factores citados se piensa que son indicadores de reproducción particularmente intensiva.
Las mujeres Wichí que comenzaron antes de los 18 años tuvieron intervalos con longitud media de 33,46 meses, más de seis meses más cortos que aquellas que lo hicieron entre los 18 y 22 años. A su vez, éstas tuvieron intervalos tres meses más breves que las que comenzaron después de los 22 años. Además, la edad a la primera maternidad mostró estar correlacionada positiva y marcadamente con la longitud de los intervalos entre partos, es decir, las madres que comienzan más temprano tienen intervalos más cortos. Resultados similares fueron hallados por Sánchez Ocasio (2003) entre los Toba para los cuales una baja edad al primer nacimiento se relaciona con intervalos cortos y tasas de nacimiento altas. En tanto en los Chorote, las longitudes de los intervalos no están relacionados con la edad al primer nacimiento y éstos se acortan a medida que las mujeres avanzan en sus intervalos fecundos buscando completar un tamaño elevado de la progenie (Mendez y Ferrarini, 2011).
La distribución de los eventos reproductivos según la edad de la madre en el momento del nacimiento permite conocer cómo varía la fertilidad de las mujeres a lo largo de su período fecundo. En los Wichí los valores máximos se alcanzan a los 15-19 y 20-24 años. La curva de fecundidad según edad de la madre tiene, como en la mayoría de las poblaciones, forma de U invertida con los valores máximos entre los 15 y los 24 años y una progresiva y lenta declinación en las edades tardías. En la mayoría de las poblaciones humanas el máximo se ubica entre los 20 y 30 años y las diferencias se dan en los primeros y los últimos años por la presencia o ausencia de control de la natalidad, las incidencias de las separaciones y la nupcialidad repetida (DeJong, 1972).
El comienzo temprano de la maternidad y la duración moderada de los intervalos intergenésicos resultan en que altas paridades ocurren a edades relativamente tempranas, cuando a las mujeres aún les restan años de vida fecunda. Tal es el caso de las mujeres que alcanzan la séptima paridad a una edad media de 34,5 años. Esto contrasta, por ejemplo con el esquema de las Aymará cuyo patrón reproductivo está marcado por un inicio tardío de la maternidad, asociado con grandes intervalos entre nacimientos y un período reproductivo bastante breve. Los intervalos entre partos, más prolongados que en muchas sociedades tradicionales, podrían ser el resultado de una restauración más lenta de la fecundabilidad inducida por el arduo estilo de vida del Altiplano, agravado por la hipoxia (Crognier et al., 2002).
La edad media de las madres Wichí en todos los órdenes de paridad es de 30,9 años, valor relativamente alto que concuerda con el de poblaciones de fecundidad natural donde las mujeres tienen sus últimos nacimientos a edades elevadas. La fecundidad natural significa simplemente que no existe control premeditado de los eventos reproductivos. El comportamiento reproductivo Wichí, regular e intenso, se acerca al de otras poblaciones con fertilidad natural como los Mapuche de Río Negro (Crognier et al., 1996) y los Chorote (Ferrarini y Mendez, 2011), pero con comienzo más temprano y difiere de las de fecundidad controlada como el caso de la población chilena de Tierra del Fuego (Pascual, 2004).
La edad media de las mujeres-madres de la comunidad es de 29,9 años. Es decir, se trata de una población joven y muchas de ellas no han completado aún su período fecundo. La estimación realizada del nivel de fecundidad que alcanzarían las mujeres Wichí una vez finalizada su carrera reproductiva y estando expuestas a las tasas de fecundidad general, específicas para cada edad, da un número esperado de 7,5 hijos por mujer. Es decir, un nivel elevado si lo comparamos con el valor medio de 5,6 hijos por mujer calculado para las sociedades cazadoras-recolectoras (Bentley et al., 1993), valor similar al calculado para los Chorote de 5,5 (Ferrarini y Mendez, 2011).
La tasa de fecundidad a edad específica, a su vez, permite inferir que al término de su período fértil la fx más elevada se instalará en el intervalo 25-29 años, lo que se ajusta al comportamiento de las poblaciones con fecundidad natural. Sin embargo, se podría esperar que ésta varíe según las condiciones ecológicas y la disponibilidad de recursos, lo que explica que las sociedades con este tipo de fecundidad muestren niveles diferentes entre sí y también por debajo del máximo fisiológico teórico (Low, 1993).
La estimación de las reducciones que la fecundidad asumiría si las mujeres Wichí, con idéntico esquema reproductivo, comenzaran más tarde su carrera fecunda ponen en evidencia una estrategia reproductora de comienzo más temprano y final más tardío, con valores algo inferiores en las edades medias y superiores en las edades mayores con respecto al esquema medio de las poblaciones de fecundidad natural de Henry (1961).
En síntesis, se desglosa una estrategia reproductiva de comienzo muy temprano y final tardío, con intervalos intergenésicos de longitud moderada que resultan en una fecundidad elevada de 7,5 hijos por madre al final del período fértil. Las mujeres Wichí maximizan la potencialidad reproductiva en relación a las limitantes ambientales en las cuales se desarrollan.
Agradecimientos
Varias personas e instituciones intervinieron para hacer posible este estudio. Agradecemos a la comunidad Kanohis del Departamento Rivadavia, Provincia de Salta, Argentina, que nos brindaron su tiempo y paciencia para acceder a la información empírica. También al Sr. Francisco Pérez por su inestimable ayuda y al Instituto Provincial del Aborigen del Ministerio de Acción Social de la Provincia de Salta, Argentina por su apoyo. Este trabajo fue subsidiado por el Consejo Nacional de Investigaciones Científicas y Técnicas de Argentina (PIP 6493) y por la Facultad de Ciencias Naturales y Museo de la Universidad Nacional de La Plata, Argentina (11/N624).
1 ALLAL, N, SEAR, R, PRENTICE, AM, and MACE, R (2004) An evolutionary model of stature, age at first birth and reproductive success in Gambian women. Proceeding of the Royal Society of London B, 271: 465-470. [ Links ]
2 APTER, D, RAISANEN, I, YLOSTALO, P, and VIHKO R (1987) Follicular growth in relation to serum hormonal patterns in adolescents compared with adult menstrual cycles. Fertility and Sterility, 47: 82-88. [ Links ]
3 APTER, D, VIINKA, L, and. VIHKO, R (1978) Hormonal patterns of adolescent menstrual cycles. Journal and Clinical Endocrinology and Metabolism, 47: 944-954. [ Links ]
4 BARÚA, G, DASSO, MC y FRANCESCHI, ZA (2008) El papel femenino en la convivencia Wichí del Chaco Central. En: HIRSCH, S (Coord.) Mujeres indígenas en la Argentina. Cuerpo, trabajo y poder. Editorial Biblos, pp. 117-151. Buenos Aires. [ Links ]
5 BENTLEY, GR, GOLDBERG, T, and JASIENSKA, G (1993) The fertility of agricultural and non-agricultural traditional societies. Population Studies, 34: 778–785.
6 BLURTON JONES N, SMITH L, O’CONNELL J, HAWKES K, and KAMUSORA CL (1992) Demography of the Hadza, an increasing and high density population of savanna foragers. American Journal of Physical Anthropology, 89:159–181
7 BOGIN B (1999) Patterns of human growth. Cambridge, Cambridge University Press. [ Links ]
8 BONGAARTS, J, and POTTER, R (1983) Fertility, biology and behavior. An analysis of the proximates determinants. New York, Academic Press. [ Links ]
9 BUMPASS, L, RIFFUS, R, and JONOSIK, R (1978) Age and marital status at first birth and pace of subsequent fertility. Demography, 15 (1): 75-80. [ Links ]
10 CAMPBELL, BC, and WOOD, JW (1988) Fertility in traditional societies. En: DIGGORY P et al., (Eds) Natural human fertility. Social and biological determinants. MacMillan Press and The Eugenics Society, pp. 39-69. London. [ Links ]
11 CROGNIER, E (2003) Reproductive Success: Which Meaning? American Journal of Human Biology, 15: 352-360. [ Links ]
12 CROGNIER, E, ELIZONDO, S, CARATINI, A, ZUBIETA, N, NIBORSKY, R y CARNESE, F (1996) Los comportamientos reproductivos de la población Mapuche de la provincia de Río Negro. Revista Argentina de Antropología Biológica, 1 (1): 86-97. [ Links ]
13 CROGNIER, E, VILLENA, M, and VARGAS, E (2002) Reproduction in high altitude Aymara: physiological stress and fertility planning? Journal of Biosocial Science 34 (4): 463-473. [ Links ]
14 DEJONG, GF (1972) Patterns of human fertility and mortality. En: HARRISON GA, and BOYCE, AJ (Eds) The structures of human populations. Clarendon Press, pp. 32-56. Oxford. [ Links ]
15 DOBLHAMMER, G (2000) Reproductive history and mortality later in life: A comparative study of England and Wales and Austria. Population Studies, 54:169-176. [ Links ]
16 ELLISON, PT (2003) Energetics and reproductive effort. American Journal of Human Biology, Vol. 15: 342-351. [ Links ]
17 FELT, JC, RIDLEY, JC, ALLEN, G, and REDEKOP, C (1990) High fertility of old colony Mennonites in Mexico. Human Biology, 62 (5): 689-700. [ Links ]
18 FERRARINI, SO y MENDEZ, MG (2011) Historia reproductora y análisis de la fecundidad de las mujeres Chorote, Argentina. Memorias de la Convención Internacional de Antropología Anthropos 2011. II Congreso Iberoamericano de Antropología, pp. 2941-2958. La Habana, Cuba. [ Links ]
19 GAISIE, SK (1984) The proximate determinants of fertility in Ghana. WFS Scientific Reports, 53 [ Links ]
20 GIBSON, M, and MACE, R (2002) The Impact of Labor-saving Technology on First birth Intervals in Rural Ethiopia. Human Biology, 74(1): 111-128. [ Links ]
21 GRAY, SJ (1994) Comparison of effect of breast-feeding practices on birth-spacing in three societies: Nomadic Turkana, Gainj and Quechua. Journal of Biosocial Science, 26 (1): 69-90. [ Links ]
22 GREKSA, LP (2002) Population growth and fertility patterns in an Old Order Amish settlement. Annals of Human Biology, 29 (2): 192-201. [ Links ]
23 GRUNDY E, and TOMASSINI, C (2005) Fertility history and health in later life: a record linkage study in England and Wales. Social Science and Medicine 61: 217-228. [ Links ]
24 GURVEN M, and KAPLAN HS (2006) Determinants of time allocation to production across the lifespan among the Machiguenga and Piro Indians of Peru. Human Nature, 17: 1–49.
25 HELLE, S (2008) A tradeoff between reproduction and growth in contemporary Finnish women. Evolution and Human Behavior, 29: 189–195.
26 HELLE, S, LUMMAA, V, and JOKELA, J (2002) Sons reduced maternal longevity in preindustrial humans. Science, 296: 1085-1085. [ Links ]
27 HELLE, S, LUMMAA, V, and JOKELA, J (2005). Are reproductive and somatic senescence coupled in humans? Late, but not early, reproduction correlated with longevity in historical Sami women. Proceeding of the Royal Society of London B, 272: 29–37.
28 HENRY, L (1961) Some data on natural fertility. Eugenics Quarterly, 8: 81-91. [ Links ]
29 HILL, K, and HURTADO, AM (1989) Hunther-gatherers of the New World. American Scientist, 77: 437-443. [ Links ]
30 HILL, K, and HURTADO, AM (1996) Ache life history. New York. Aldine de Gruyter [ Links ]
31 KÄÄR, P, and JOKELA, J (1998) Natural selection on age-specific fertilities in human females: comparison of individual-level fitness measures. Proceeding of the Royal Society of London B, 265: 2415-2420. [ Links ]
32 KAPLAN, HS, LANCASTER, JB, and ROBSON, A (2003) Embodied capital and the evolutionary economics of the human life span. En: CAREY, JR, and TULJAPURKAR, S (Eds.) Life span: evolutionary, ecological, and demographic perspectives. Population and Development Review, Supplement Vol. 29. Population Council, pp. 152-182. New York. [ Links ]
33 KAPLAN, HS, and LANCASTER, JB (2004) An Evolutionary and ecological analysis of human fertility, mating patterns, and parental investment. En: WACHTER, KW, and BULATAO, RA (Eds) Offsprings. Human fertility behavior in biodemographic perspective. The National Academies Press, pp. 170-223. Washington, D.C. [ Links ]
34 KAPLAN, HS, and GURVEN, M (2008) Top-down and bottom-up research in biodemography. Demographic Research, 19, Art. 44: 1587-1602. [ Links ]
35 KIRK, KM, BLOMBERG, SP, DUFFY, DL, HEATH, AC, OWENS, IPF, and MARTIN, NG (2001) Natural selection and quantitative genetics of life-history traits in western women: A twin study. Evolution, 55: 423–435.
36 KONOGOLO, L (1985) Variation in entry into matherhood and length of effective reproductive life among women in Kenya. Studies in African and Asian Demography. CDC Annual Seminar. [ Links ]
37 KORPELAINEN, H (2000) Fitness, reproduction and longevity among European aristocrats and rural Finnish families in the 1700s and 1800s. Proceeding of the Royal Society of London B, 267: 1765-1770. [ Links ]
38 LIVI-BACCI, M (2011) Introducción a la demografía. Barcelona. Editorial Ariel. [ Links ]
39 LOW, BS (1993) Ecological demography. A synthetic focus in evolutionary anthropology. Evolutionary Anthropology, 1 (5): 177-187. [ Links ]
40 MASNICK, GS (1979) The demographic impact of breastfeeding: a critical review. Human Biology, 51 (2): 109-125. [ Links ]
41 MCALPINE, PJ, and SIMPSON, NE (1976) Fertility and other demographic aspects of the Canadian Eskimo communities of Igloolik and Hall Beach. Human Biology, 48 (1): 113-138. [ Links ]
42 MCKIBBEN, SL (2003) The social construction of adulthood: menarche and motherhood. Tesis. Texas A&M University.
43 MENDEZ, MG y FERRARINI, SO (2009) Rasgos fenotípicos y desempeño reproductivo en hombres y mujeres Chorote. En: MÉNDEZ, MG (Comp.) Terra Incognita. Estudios Antropológicos en el Chaco Meridional. Editorial Universitaria de La Plata, pp. 35-55. La Plata, Argentina. [ Links ]
44 MENDEZ, MG y FERRARINI, SO (2011) Intervalos entre nacimientos e inversión parental en aborígenes Chorote de Argentina. Memorias de la Convención Internacional de Antropología Anthropos 2011. II Congreso Iberoamericano de Antropología, pp. 2959-2976. La Habana, Cuba. [ Links ]
45 MENKEN, J (1980) The health and demographic consequences of adolescent pregnancy and childbearing. En: CHILMAN, CS (Ed) Adolescent pregnancy and childbearing; findings from research. Washington DC. US Department of Health and Human Services. [ Links ]
46 NGALINDA, I (1998) Age at First Birth, Fertility, and Contraception in Tanzania. Tesis. Humboldt University of Berlin. [ Links ]
47 NONAKA, K, MIURA, T, and PETER, K (1994) Recent fertility decline in Dariusleut Hutterites: an extension of Eaton and Mayer’s Hutterite fertility study. Human Biology, 66 (3): 411-420.
48 PASCUAL, J (2004) Caracterización de la historia reproductora y análisis de la fecundidad de las mujeres de Tierra del Fuego. Tesis. Universidad de Barcelona. [ Links ]
49 PETTAY, JE, HELLE, S, JOKELA, J, and LUMMAA, V (2007). Natural selection on female life-history traits in relation to socio-economic class in preindustrial human populations. PLoS ONE. 2 (7): e606. [ Links ]
50 SÁNCHEZ OCASIO, K (2003) A Fork in Fertility: The Demographic Transition and Factors Mediating a Fertility Decline among the Indigenous Toba of Argentina. Tesis. Harvard University, Cambridge, Massachusetts. [ Links ]
51 SEAR, R (2007) The impact of reproduction on Gambian women: does controlling for phenotypic quality reveal costs of reproduction? American Journal of Physical Anthropology, 132 (4): 632-641. [ Links ]
52 SEAR, R, MACE, R, and McGREGOR, IA (2000) Maternal grandmothers improve the nutritional status and survival of children in rural Gambia. Proceeding of the Royal Society of London B, 267: 1641-1647. [ Links ]
53 SEAR, R, ALLAL, N, and MACE, R (2004) Height, marriage and reproductive success in Gambian women. Research in Economic Anthropology, 23: 203-224. [ Links ]
54 SENDEROWITZ, J, and PAXMAN, JM (1985) Adolescent Fertility: Worldwide Concerns. Population Bulletin, 40 (2): 3-49. [ Links ]
55 SMITH, KR, MINEAU, GP, and BEAN LL (2002) Fertility and post-reproductive longevity. Social Biology, 49: 185-205. [ Links ]
56 STEARNS, SC, and KOELLA, JC (1986) The evolution of phenotypic plasticity in life history traits: predictions of reaction norms for age and size at maturity. Evolution, 40: 893–913.
57 STEVENSON, JC, EVERSON, P, and CRAWFORD, M (1989) Changes in completed family size and reproductive span in Anabaptist populations. Human Biology, 61 (1): 99-115. [ Links ]
58 VALEGGIA, C, and ELLISON, PT (2003) Impact of breastfeeding on anthropometric changes in peri-urban Toba women (Argentina). American Journal of Human Biology, 15: 1–8
59 VAREA, C, BERNIS, C, CROGNIER, E, BAALI, A y HILALI, MK (2007) Tendencias contrapuestas en los determinantes de la fertilidad en poblaciones tradicionales: el ejemplo de Marruecos. Antropología Física Latinoamericana, 5: 63-78. [ Links ]
60 VITZTHUM, VJ (2008) Evolutionary models of women’s reproductive functioning. Annual Review of Anthropology, 37: 53-73.
61 VOYDANOFF, P, and DONNELLY, BW (1990) Adolescent Sexuality and Pregnancy. Newbury Park. Sage Publications. [ Links ]
62 WALKER, R, GURVEN, M, HILL, K, MIGLIANO, A, CHAGNON, N, DE SOUZA, R, DJUROVIC, G, HAMES, R, HURTADO, AM, KAPLAN, H, KRAMER, K, OLIVER, WJ, VALEGGIA, C, and YAMAUCHI, T (2006) Growth Rates and Life Histories in Twenty-Two Small-Scale Societies. American Journal of Human Biology, 18: 295–311
63 WESTOFF, CF, BLANC, AK, and NYBLADE, L (1994) Marriage and entry into parenthood. DHS Comparative Studies Nº 10. Calverton, Maryland. Macro International Inc. [ Links ]
64 WOOD, JW (1994) Dynamics of human reproduction. Biology, biometry, demography. New York. Aldine de Gruyter. [ Links ]
65 WULF, D and SINGH, S (1991) Sexual activity, union and childbearing among adolescent women in the Americas. International Family Planning Perspectives, 17 (4): 137-144. [ Links ]
66 YAMAUCHI, T, SATO, H, and KAWAMURA K. 2000. Nutritional status, activity pattern, and dietary intake among the Baka hunter-gatherers in the village camps in Cameroon. African Study Monograph 21:67–82.

