










Servicios Personalizados
Revista

Articulo
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkCuadernos de la Facultad de Humanidades y Ciencias Sociales. Universidad Nacional de Jujuy
versión On-line ISSN 1668-8104
Cuad. Fac. Humanid. Cienc. Soc., Univ. Nac. Jujuy no.49 San Salvador de Jujuy jun. 2016
ARTÍCULO ORIGINAL
La reproducción chorote en números
(Chorote reproduction in numbers)
Marta Graciela Méndez* - Stella Ofelia Ferrarini*
* CONICET - Facultad de Ciencias Naturales y Museo - Universidad Nacional de La Plata. Calles 122 y 60 s/nº - CP 1900 - La Plata - Argentina. Correo Electrónico: gmendez@fcnym.unlp.edu.ar
RESUMEN
La expresión numérica de las decisiones reproductivas reflejan las estrategias evolutivas de un grupo humano. Se procura comprender los mecanismos biodemográficos, con centro en la reproducción y la fertilidad, dentro del contexto biocultural Chorote. Se estudiaron las historias reproductoras de 71 mujeres con al menos un hijo. Se registraron nacimientos, edades de las madres y sexo de los hijos. Se calcularon los estadísticos descriptivos de las variables y se las relacionaron a través de pruebas de significación. Se computaron las tasas específicas de fecundidad y el tamaño medio de la descendencia que una mujer teóricamente alcanzará. La edad media al primer nacimiento es de 20,22 años. La longitud media de los intervalos intergenésicos es 3,2 años y se acorta a medida que las mujeres avanzan en su intervalo fecundo. Se manifiesta una tendencia a espaciamientos más cortos en mujeres con inicio rezagado de la maternidad. La tasa de fecundidad fx es de 5,55 de comienzo temprano y finalización tardía. La iniciación de la reproducción muestra una tendencia a prolongar la etapa de crecimiento y mejorar las condiciones de salud y nutrición para enfrentar sucesivos embarazos. La edad media a las distintas paridades concuerda con el de poblaciones de fertilidad natural con un tamaño final de la progenie moderado en relación a otros grupos cazadores-recolectores. La longitud del intervalo intergenésico es significativa en relación al sexo del hijo que lo abre, favoreciendo a las hijas. Esto se interpreta como una toma de decisión reproductiva sesgada por la matrilocalidad y con fundamento evolutivo.
Palabras Clave: Chorote; historia reproductiva, primer nacimiento, intervalo entre nacimientos, sexo de los hijos.
ABSTRACT
The numeric expression of reproductive decisions reflects the evolutionary strategies of a human group. We tried to understand the bio-demographic mechanisms, with center in reproduction and fertility, inside the bioculturalChorote context. The reproductive histories of 71 women were studied with at least a son. It was registered births, mothers' ages and the children's sex. Descriptive statistical of the variables were calculated and they were related through significance tests. The specific rates of fecundity and the average number of the descendant that a woman theoretically will reach were computed. The mean age at the first birth is of 20,2 years. The average length of the intergenesic intervals is 3,2 years and it shortens as the women advance in its fertile interval. A tendency is manifested to shorter spacing in women with beginning straggler of the maternity. Fertility rate fx is 5,5, with early starting and late ending. The initiation of the reproduction shows a tendency to prolong the stage of growth and to improve health and nutrition conditions to face successive pregnancies. The mean age of the different parities agrees with that of populations of natural fertility with a final size of the offspring moderated in relation to other group's hunter-gatherers. The longitude of the intergenesic interval is significant in relation to the child's sex that opens it, favoring the daughters. It is interpreted as a reproductive decision slanted by the matrilocality with evolutionary foundation.
Key Words: Chorote, reproductive history, first birth, interbirth intervals, sex of the children.
INTRODUCCIÓN
Los principales factores que afectan la salud y la supervivencia de la madre y su hijo derivan de los patrones reproductivos asumidos por el grupo social: el acceso temprano a la gestación, el mantenimiento de la reproducción hasta etapas finales del período fértil y la intensidad de la natalidad.
La edad a la que una mujer accede a la maternidad es altamente variable y depende tanto de factores biológicos como sociales. Un gran número de estudios han relacionado la edad al primer nacimiento con la fertilidad, sobre todo en poblaciones con fertilidad natural (Bumpass et al., 1978; Gaisie, 1984; Konogolo, 1985; Westoff et al., 1994). El nacimiento del primer hijo es un evento de gran significación social y su importancia es reconocida en todas las sociedades humanas. Expresa la transición de una pareja dentro de un nuevo estatus social, la paternidad, con nuevas expectativas y responsabilidades relacionadas. También es el último peldaño en el proceso que conduce a una mujer a la madurez sexual y que comienza en la pubertad con la menarca. Desde un punto de vista evolutivo, la edad al primer nacimiento es uno de los eventos que modelan las estrategias vitales de los grupos humanos, dado que la energía y el tiempo son recursos finitos que deben ser administrados para el crecimiento, el mantenimiento y la reproducción. A lo largo de su historia vital los individuos deben repartir estos recursos de maneras que terminarán afectando su desempeño reproductivo. Esto es crucial en las mujeres en las cuales el embarazo, parto y amamantamiento requieren importantes inversiones tanto de tiempo como de energía. Después de la menarca deberá establecerse un equilibrio entre la reproducción presente y la futura, es decir entre inversiones en el propio capital corpóreo versus inversiones inmediatas en reproducción (Kaplan y Lancaster, 2003). En las sociedades cazadoras-recolectoras donde los recursos alimenticios son limitados, y aún escasos en algunas épocas del año como sucede entre los Chorote, estas negociaciones son críticas en términos de la supervivencia tanto de las madres como de sus descendientes. Cuando la nutrición es pobre durante el embarazo, las mujeres deben retirar reservas acumuladas a través de la niñez. De esta manera la postergación de la edad del comienzo de la reproducción aumenta la probabilidad de supervivencia propia y de sus hijos (Allal et al., 2004).
La edad de la última maternidad determina el final del período temporal que la mujer destina para tener su descendencia y por lo tanto la longitud temporal del período fecundo. Cuando los nacimientos se suceden sin restricciones la edad a la última maternidad se aproxima a la edad natural de la declinación de la fertilidad. El término fecundidad natural no hace referencia a la máxima fecundidad potencialmente alcanzable por los individuos de una población, sino a la fecundidad caracterizada por la no existencia de mecanismo alguno de control de la fertilidad dependiente del tamaño familiar alcanzado en cada momento de la etapa reproductiva (Henry, 1961).
El período de tiempo entre dos nacimientos sucesivos es un determinante importante de los niveles de fecundidad y en especial en las poblaciones con fertilidad natural, dado que se ha demostrado que la fecundidad está inversamente relacionada con el intervalo cerrado promedio entre nacimientos.
Por otra parte, el intervalo entre nacimientos es una medida de la inversión parental en un hijo. Un intervalo largo mejora el desarrollo del hijo ya nacido y toma tiempo del intervalo fecundo durante el cual la madre podría tener más hijos. Se ha demostrado en muchos estudios que los intervalos cortos generan mayor morbilidad y mortalidad infantil.
En este trabajo se procura comprender, dentro del contexto cultural Chorote, cómo los mecanismos biodemográficos, centrados en la reproducción y la fertilidad, afectan a la descendencia presente y futura.
MATERIALES Y MÉTODOS
CARACTERÍSTICAS DE LA COMUNIDAD
La población estudiada habita el norte de la provincia de Salta, sobre el río Pilcomayo, en dos poblados de alrededor de 385 personas que conservan el patrón de asentamiento arcaico circular. El grupo doméstico se organiza alrededor de una pareja, sus hijas y respectivos yernos y los hijos de estos; es decir, una familia compuesta matrilocal. Las familias emparentadas habitan en viviendas contiguas lo que asegura un esquema de convite y reciprocidad de bienes alimentarios. Tradicionalmente los Chorote han sido cazadores, recolectores y pescadores con nomadismo estacional. En la actualidad conservan en parte su modo de subsistencia ancestral, a pesar de haberse incorporado al trabajo rural como peones asalariados hace ya más de seis décadas. Subsiste entre ellos la división sexual del trabajo con las mujeres dedicadas a la recolección y a las tareas hogareñas y con los hombres entregados a la caza, la pesca y a trabajos ocasionales como obreros rurales. El nomadismo estacional subsiste bajo las apariencias de visitas a familias emparentadas que residen en otros asentamientos, con acceso a recursos diferentes. Entre los Chorote el matrimonio es una pauta universal y la norma es la monogamia. Es la mujer la que elige a su pareja en función de concebir al hombre como un productor de alimentos. Así, se asegura a través del marido la posibilidad de evitar el hambre al insertarse en el intercambio de bienes. También es la mujer la que distribuye los productos obtenidos por el marido y quien recibe los bienes que provienen de otro hombre, a través de su esposa. En la crianza de los niños la lactancia a demanda es la práctica social prescripta y las mujeres emplean una manta a manera de cabestrillo para acarrear a los niños y facilitar este proceso. (Siffredi, 1977; Mendez y Ferrarini, 2009; Ferrarini y Mendez, 2009).
METODOLOGÍA
Se relevaron las historias reproductoras de 71 mujeres Chorote con al menos un hijo. Se registraron las historias de nacimientos, edades de las madres y sexo de los hijos. Se calcularon los estadísticos descriptivos de la edad de la madre al primer nacimiento, las edades en los distintos órdenes de paridad y las longitudes de los intervalos entre nacimientos. Se examinó la distribución de las frecuencias por clases de intervalo. La información se desglosó para cada orden de paridad. Las longitudes medias de los intervalos por paridad se analizaron en función de la edad al primer nacimiento y se indagó la relación entre ambas variables a través del coeficiente de correlación. Se exploró a través de la prueba “t”, el efecto del tamaño de la progenie sobre la longitud del intervalo. Para indagar la inversión parental en algún sexo se compararon las longitudes de los intervalos según el sexo del hijo consecutivo, el del que abre y el que cierra el intervalo, y se investigó su significación estadística a través del análisis de la varianza. Se calcularon las tasas específicas de fecundidad fx y se analizaron los patrones de fecundidad con las respectivas probabilidades en todas las edades y grupos de edad. A su vez, se estimaron las contribuciones de cada grupo de edad al valor final de la tasa de fecundidad y, consecuentemente, el tamaño medio de la descendencia que una mujer teóricamente tendría. Sobre una sub-muestra de mujeres que ya completaron su período fecundo se estudiaron las distribuciones de la edad a la última maternidad y de la longitud del intervalo fecundo, se compararon las longitudes del intervalo fecundo y de los intervalos entre nacimientos y el número medio de hijos nacidos vivos con las longitudes de los períodos entre nacimientos.
RESULTADOS
La edad media del primer alumbramiento abre el período fecundo de la mujer. En las poblaciones de pequeña escala es un indicador clave del tiempo que, en términos de inversión, se ha dedicado al crecimiento. En muchos casos, un comienzo excesivamente temprano pone en riesgo la salud de la madre y/o de su hijo.
La edad media de las mujeres Chorote encuestadas es de 20,22 años (Tabla 1), edad intermedia cuando se compara con las de otras poblaciones. Así, por ejemplo, otras sociedades cazadoras recolectoras como los Esquimales, Aché y Toba tienen una media a la primera maternidad de 17,5 años, mientras los Aymara de Bolivia registran una edad media de 24, 1 (McAlpine y Simpson, 1976; Hill y Hurtado, 1989; Sánchez Ocasio, 2003; Crognier, 2003). Los Chorote comparten el valor de este parámetro con los Mapuche (Crognier et al., 1996).
En la Tabla 1 se registran las edades de las madres en las distintas paridades para observar cómo se han regulado temporalmente estos eventos en las historias reproductoras de las mujeres. Obviamente, las edades se incrementan en el devenir reproductivo. También se observa que las mujeres continúan teniendo hijos hasta la finalización de sus períodos fecundos y que el número de los mismos es alto, una manifestación clara de la ausencia de contraconcepción. Un promedio general de las edades a las distintas paridades, o sea la edad media de la maternidad, alcanza el valor de 32,3 años, que es relativamente alto y concuerda con el de poblaciones de fecundidad natural, por cuanto las mujeres tienen sus últimos nacimientos a edades elevadas.

EUM: Edad de la madre a la última maternidad
Tabla 1. Descriptivos de la edad de la madre en las distintas paridades y de la edad a la última maternidad (en años)
La Tabla 1 también expone la distribución de la edad media de la última maternidad en una sub-muestra de mujeres que ya experimentaron la menopausia. El valor medio es una edad tardía y cercana a la tradicionalmente aceptada como propia de poblaciones con fecundidad natural (Huteritas de Norteamérica y Mennonitas de México: 40,9 años; Canadá, siglo XVII: 40, 1; Amizamiz de Marruecos: 38,8) (Bongaarts y Potter, 1983; Felt et al., 1990; Bernis, 1990). Los valores registrados para las mujeres Chorote atestiguan que la edad de última maternidad depende más del calendario biológico que de factores externos.
La longitud del intervalo fértil teórico de una mujer es una fuente significativa de la fecundidad diferencial entre las poblaciones humanas. No obstante, las distintas poblaciones pueden utilizar de manera muy diferente su potencial por diversas causas, sean estas de tipo biológico, económico o socio-cultural. Por ello, el análisis de la longitud del intervalo fecundo de las mujeres de una población tiene mayor interés debido a su carácter más informativo. El intervalo fecundo es el período temporal que transcurre entre las edades de la madre al nacimiento del primer y del último hijo, por lo que la estimación de su longitud informa sobre la fracción de la vida reproductora que las mujeres destinan efectivamente a tener su descendencia. Las características principales de la distribución de este intervalo se muestran en la Tabla 2. La longitud media se revela larga al comparar con poblaciones con fertilidad controlada (Tierra del Fuego, Chile: 8,9) y aún en el cotejo con poblaciones con fertilidad natural (Mapuche, Argentina: 14,9; Bereberes, Marruecos: 17,2) (Pascual, 2004; Crognier et al., 1996; Crognier et al., 2001).
Al analizar la influencia que sobre la longitud del intervalo fecundo tiene el inicio de la maternidad queda explícito que a medida que aumenta la edad materna al primer nacimiento se reduce la longitud media del intervalo fecundo. Comenzar antes de la edad media de la primera maternidad para las mujeres Chorote implica una ganancia de cinco años en el intervalo fecundo (Tabla 2).

Tabla 2. Descriptivos de la distribución de la longitud del intervalo fecundo (LIF) y longitud media según la edad al primer nacimiento (EPN)(en años)
La Tabla 3 muestra la longitud media del intervalo entre nacimientos para la población Chorote.

Tabla 3. Descriptivos de la longitud intergénesica media (IIM) y por paridades (en años)
En los estudios realizados en varias poblaciones sobre longitudes medias de los intervalos entre nacimientos, estas se alargan a medida que se incrementa la paridad. El análisis en esta población muestra que las longitudes van disminuyendo gradualmente, poniendo en evidencia que las mujeres Chorote buscan completar un tamaño de progenie elevado, acortando los intervalos entre nacimientos a medida que avanzan en su intervalo fecundo. Esto se demuestra en la Tabla 3, no sólo en los valores de la longitud media sino también por el descenso en el coeficiente de variabilidad. Para la paridad 11, el intervalo es amplio por la razón biológica de disminución de fertilidad que acontece sobre el final de la vida reproductiva.
Bongaarts y Potter (1983) han descrito la edad materna al primer nacimiento como uno de los determinantes de la longitud de los intervalos entre nacimientos. En las poblaciones para las que se calcularon relaciones de correlación entre ambos parámetros se encontró que era positiva, indicando que las mujeres que tienen su primer hijo a edades más avanzadas tienden a tener intervalos más largos. En los Chorote, el cálculo del coeficiente de correlación para las mujeres que han completado su reproducción arroja un valor negativo (-0,054), lo cual indica que la relación es inversa. Es decir, cuanto más elevada es la edad al primer nacimiento la tendencia es tener hijos a intervalos más cortos. Al examinar la relación entre la longitud del intervalo fecundo según la duración del intervalo intergenésico se observa que longitudes medias de intervalo fecundo más bajas se asocian con intervalos intergénesicos medios más cortos (Tabla 4).

Tabla 4. Descriptivos de la longitud del intervalo fecundo según la duración del intervalo intergenésico medio (IIM) (en años)
Cuando se considera conjuntamente la paridad y la edad de la madre al primer nacimiento, corroboran la débil influencia que parece tener el inicio del período fecundo sobre la longitud de los intervalos entre nacimientos posteriores. Aquellas mujeres que comenzaron su reproducción antes de los 20 años mantienen la longitud hasta la paridad 4 y disminuyen ligeramente en las posteriores. Las que tuvieron su primer hijo entre los 20 y 24 años acortan sus intervalos a partir de la paridad 3. Por último, aquellas que iniciaron tardíamente su período fecundo, más allá de los 25 años, comienzan con un intervalo más corto que el resto y tienen los subsiguientes más largos. Adviértase que son pocas y que pueden estar comprendidas en el porcentaje de mujeres con dificultades relacionadas con la fertilidad o con la estabilidad conyugal, como se manifiestan en todas las sociedades (Tabla 5).
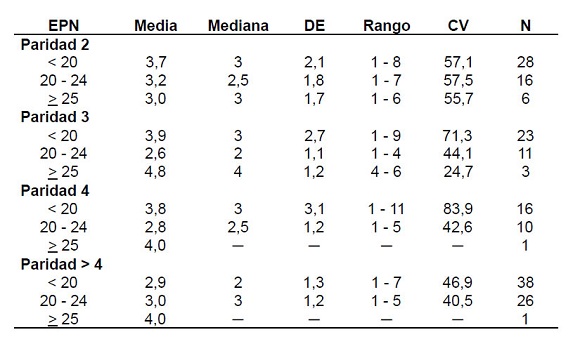
Tabla 5. Descriptivos de la longitud de los intervalos entre nacimientos según la paridad y la edad de la primera maternidad (en años)
Se examinó, luego, el efecto del tamaño de la progenie sobre la longitud de los intervalos entre nacimientos, considerando las diferencias entre madres con menos y más de 6 hijos. Se tomó este valor como punto de corte debido a que el número medio de hijos nacidos vivos, para mujeres con su período reproductor completado, es 6,6. Se encontró que el intervalo medio para las primeras es de 4,4 años y de 3,1 para las segundas. Esta diferencia no resultó significativa al ser testeada estadísticamente a través de la prueba “t”, a pesar de que las mujeres con mayor descendencia tienen sus hijos más próximos.
Varias investigaciones han indicado que los intervalos entre nacimientos que siguen a un varón son más largos que los que siguen a una mujer (Greenberg y White, 1967; Wyshak, 1969; Nath et al., 2000). Se ha resaltado que cuanto mayor es el costo energético de un niño para la madre y más largo el período sobre el cual ese costo es extraído, más largo será el intervalo. Se indagaron, entonces, las longitudes medias de los intervalos en todas las paridades teniendo en cuenta el sexo del hijo consecutivo (Tabla 6). Se encontró que el intervalo más largo es el que separa el nacimiento de dos hijas mujeres y el más corto el correspondiente al de varón seguido por mujer. La diferencia entre ambos es de 1 año.

Tabla 6. Descriptivos de la longitud del intervalo entre nacimientos según el sexo del hijo consecutivo en todas las paridades (en años)
Cuando se aplicó el análisis de la varianza sobre la longitud de los intervalos entre nacimientos según el sexo del niño previo y el actual se halló que existen diferencias significativas entre mujer-mujer con varón-mujer (F = 6,437; p = 0,013) y entre varón-mujer con mujer-varón (F = 6,368; p = 0,013). Las restantes diferencias no lo fueron. Las diferencias significativas están entre las díadas que presentan las mayores distancias temporales.
Se exploró, a continuación, si estas diferencias estaban presentes en las diversas paridades (Tabla 7) y se vio que las mayores longitudes se presentaban cuando las mujeres abrían los intervalos. Las menores, en cambio, se daban cuando los intervalos eran abiertos por varones.
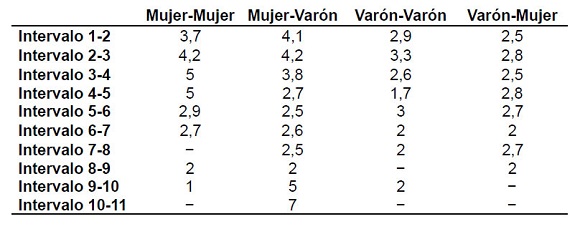
Tabla 7. Longitud del intervalo entre nacimientos según el sexo del hijo consecutivo y las diferentes paridades (en años)
Esto condujo a examinar el efecto del sexo que abre el intervalo. Los resultados se muestran en la Tabla 8.

Tabla 8. Longitud del intervalo entre nacimientos abierto y cerrado por mujer o varón sin tener en cuenta el sexo del hijo consecutivo en las distintas paridades (en años)
La longitud de los intervalos que siguen al nacimiento de una niña son más largos que los que siguen al de un niño. Al aplicar el análisis de la varianza sobre estos datos dio resultados significativos para la diferencia (F = 9,330; p = 0,002). También se analizó el efecto del sexo que cierra el intervalo (Tabla 8). En este caso no se halló significación (F = 0,064; p = 0,799). De manera que los Chorote muestran una clara diferencia en la inversión parental sobre el sexo femenino.
El número medio de hijos nacidos vivos (HNV) permite ponderar el resultado final de las historias reproductoras de las mujeres en una población y, por lo tanto, debe hacerse sobre la población femenina que ha completado su ciclo reproductivo. Como ya se ha dicho, entre los Chorote el número medio HNV es 6,6, moderado en relación a otras poblaciones de fertilidad natural aunque se encuentra comprendido entre los obtenidos para los grupos cazadores-recolectores (Hill y Hurtado, 1996).
Si en el grupo de mujeres algunas aún no ha completado su vida reproductiva –como es el caso aquí estudiado- se estila estimar la fecundidad esperada a través de la tasa F que brinda una medida de fecundidad pura, no influenciada ni por la estructura de la población ni por la mortalidad y puede interpretarse en términos de la reproducción esperada individual (Campbell y Wood, 1988). Por otra parte, la distribución de los nacimientos según la edad materna al momento del nacimiento informa sobre las variaciones de fecundidad de las mujeres a lo largo de su vida reproductora. A partir de esta distribución pueden calcularse las tasas específicas de fecundidad a una edad o intervalo determinado. Esta medida de la fecundidad permite un análisis más detallado, ya que la probabilidad de tener un hijo no es la misma en todas las edades (Livi-Bacci, 1993). De la sumatoria de las tasas específicas de fecundidad se obtiene una estimación del número de descendientes del grupo de mujeres de la muestra, si todas ellas sobrevivieran hasta el final del período reproductor y estuvieran expuestas a las tasas de fecundidad específicas para cada edad (Campbell y Wood, 1988). El valor obtenido es de 5,5 hijos nacidos vivos.
DISCUSIÓN
El ritmo de la maduración y de la edad al primer nacimiento afecta el potencial individual del éxito reproductivo del curso de la vida, que tiene importantes implicancias como fuerza de la selección. El comienzo relativamente tardío en relación a otras sociedades de pequeña escala, con una edad media a la primera maternidad de 20,2 años, muestra que prolongan hasta la segunda década su etapa de crecimiento mejorando sus condiciones fenotípicas para enfrentar posteriores embarazos (Allal et al., 2004; Hill y Hurtado, 1996).
Los Chorote registran una de las longitudes del intervalo entre nacimientos más altas descritas para diversas sociedades humanas, con valores similares a los Aché y a los Wichí otros grupos de cazadores-recolectores del Gran Chaco Sudamericano (Tabla 9). Los datos mostrados dan cuenta de la amplia variabilidad del parámetro, con los valores más bajos para los Amish y poblaciones históricas europeas y con los valores más altos para grupos del África sub-sahariana con prácticas extensivas de lactancia materna y abstinencia sexual, como los ¡Kung. El valor del intervalo en los Chorote se expresa en meses para facilitar las comparaciones.

Tabla 9. Longitud media del intervalo entre nacimientos en diversas poblaciones (en meses)
La sucesión de partos a lo largo de todo el intervalo fecundo hace que la edad media de la maternidad, considerando todos los órdenes de paridad, sea de 32,3 años evidenciando que no existen prácticas contraconceptivas en esta población. También lo confirman los valores de la edad media de la última maternidad (39,1 años) y de la longitud media del intervalo fecundo (18 años). Para las mujeres Chorote con fertilidad completada el intervalo fecundo es largo, aún si se lo confronta con otras mediciones en poblaciones con fertilidad natural, sobre todo teniendo en cuenta que el valor medio de inicio de la maternidad supera en dos y hasta tres años a los registrados en otras poblaciones cazadoras-recolectoras. El inicio tiene su influencia sobre la longitud del intervalo fecundo, así las mujeres que comienzan antes de la edad media al primer nacimiento ganan hasta cinco años de fecundidad.
La regulación temporal de los nacimientos es otra medida para evaluar el rendimiento reproductivo del período fecundo y del gasto de tiempo y energía que los progenitores están dispuestos a hacer por su descendencia. Se ha observado repetidamente una asociación entre la abundancia de cortos intervalos intergenésicos y una posterior elevada mortalidad infantil en diversidad de poblaciones, incluidos países industrializados (Miller, 1989), países contemporáneos en desarrollo (Hobcraft et al, 1985) y poblaciones históricas (Pebley et al., 1991). El riesgo de mortalidad infantil se reduce drásticamente a medida que la longitud del intervalo entre nacimientos aumenta hasta alcanzar los mínimos estables entre los 24 y los 28 meses (Pebley et al. 1991). En este sentido los intervalos entre nacimientos con un valor promedio alto de 3,2 años (38,2 meses) demuestran un esfuerzo parental para lograr la supervivencia del hijo recién nacido que se vigoriza a través de una prolongada lactancia.
Ahora bien, en contraste con los resultados obtenidos en estudios realizados sobre longitudes medias de los intervalos entre nacimientos en varias poblaciones, los Chorote disminuyen gradualmente la regulación temporal poniendo en evidencia que buscan completar un tamaño de progenie elevado acortando los intervalos a medida que avanzan en su período fecundo. Tanto es así que las mujeres con fertilidad completada que registran intervalos intergenésicos más cortos tienen intervalos fecundos más cortos.
Los intervalos entre nacimientos muestran una correlación negativa, aunque débil, con la edad al primer nacimiento de manera que cuanto más elevada la edad de la primera maternidad, la tendencia es tener hijos a intervalos más cortos. Los Wichí presentan un intervalo medio similar, no obstante, la estrategia difiere. Su primera maternidad asume un valor medio de 18,2 años y las madres que comienzan su maternidad más temprano tienen intervalos más cortos, mostrando una correlación positiva. Considerando estos dos aspectos de las estrategias reproductivas, la diferencia principal reside en comenzar antes aunque tengan los hijos con el mismo ritmo promedio (Mendez y Ferrarini, 2011; Ferrarini y Mendez, 2011).
El efecto del tamaño de la progenie sobre la longitud de los intervalos intergenésicos se puso en evidencia: las mujeres con mayor descendencia tienen sus hijos más próximos, aunque no muestren diferencias significativas con las que tienen menos.
La inversión de energía costosa en la producción de descendientes está en las bases ecológicas para el éxito evolutivo o reproductivo. Las variaciones en recursos y su asignación a la reproducción son básicas para comprender las diferencias en las medidas de fertilidad. Muchos estudios informan que los intervalos entre nacimientos varían según el sexo del hijo (Greenberg y White, 1967; Westoff y col.; 1961; Wyshak, 1969; Blanchart y Bogaert, 1997). Unos pocos estudios empíricos demuestran diferencias dentro de la sociedad en la atención, cuidado y asignación de recursos a hijos e hijas (Hrdy, 1987, Sieff, 1990). Se ha formulado la hipótesis que los padres proveen mejor cuidado a los varones que a las mujeres cuando los varones poseen una ventaja reproductiva sobre las mujeres con respecto a la asignación de recursos (Trivers y Willard, 1973). Una abuela u otra mujer mayor, genéticamente relacionada a la mujer que está criando niños, pueden incrementar la fertilidad de la madre a través de sus contribuciones energéticas de trabajo y producción de alimento, y de esta forma mejorar su propio éxito evolutivo (Hawkes et al., 1998). Dado que el intervalo intergenésico más largo es el que separa el nacimiento de dos hijas mujeres y el más corto aquel del varón seguido por mujer, entre los Chorote la longitud del intervalo mostró estar relacionada con el sexo. El efecto del sexo que abre el intervalo se puso de manifiesto, toda vez que todos los intervalos abiertos por mujer son más largos que los abiertos por varón no importando el sexo que sigue. Se interpreta como una toma de decisión reproductiva sesgada por la matrilocalidad y con fundamento evolutivo. Esto es importante ya que el nacimiento de un nuevo hermano puede tener un inmediato efecto perjudicial sobre la morbilidad y mortalidad de su hermano mayor (Bohler y Bergström, 1995). Entre los Chorote no se hallaron muestras del efecto del sexo que cierra el intervalo, de esto surge que la inversión parental manifiesta un marcado sesgo hacia las mujeres.
El tamaño medio de la progenie entre las mujeres que completaron su intervalo fecundo es de 6,6 hijos nacidos vivos. En el conjunto de las madres la tasa F de fecundidad a edad específica es de 5,5. Para una comparación con otra etnia chaqueña, al final de sus carreras reproductivas las madres Wichí tienen una tasa de fecundidad de 7,5 hijos (Mendez y Ferrarini, 2011). Es decir, que del análisis comparado de las estrategias surge que aún con el mismo espaciamiento intergenésico, por comenzar dos años antes su período fecundo, las Wichí lo concluyen con dos hijos más.
CONCLUSIONES
Las etnias chaqueñas siguen esquemas reproductivos propios. En algunos casos el resultado de la aplicación de estos esquemas conduce a poblaciones de mayor tamaño, con un efectivo numérico importante. En otras se registra una estabilidad a través del tiempo que abre interrogantes acerca de su futuro. Los Chorote constituyen un ejemplo de esta última premisa y merecen ser analizados en todas las variables involucradas en su historia de vida.
La iniciación de la reproducción muestra una tendencia a prolongar la etapa de crecimiento y mejorar las condiciones de salud y nutrición para enfrentar sucesivos embarazos. La longitud media de los intervalos intergenésicos es 3,2 años y se acorta a medida que las mujeres avanzan en su intervalo fecundo. Se manifiesta una tendencia a espaciamientos más cortos en mujeres con inicio rezagado de la maternidad. La tasa de fecundidad fx es de 5,5 de comienzo temprano y finalización tardía. La edad media a las distintas paridades concuerda con el de poblaciones de fertilidad natural con un tamaño final de la progenie moderado en relación a otros grupos cazadores-recolectores. La longitud del intervalo intergenésico es significativa en relación al sexo del hijo que lo abre, favoreciendo a las hijas. Esto se interpreta como una toma de decisión reproductiva sesgada por la matrilocalidad y con fundamento evolutivo. Estos números y las pautas seguidas para lograrlos ponen en evidencia que las estrategias asumidas por los Chorote para su reproducción son expresivas, y también explicativas, de la situación demográfica en que las poblaciones se encuentran hoy. Las mediciones oficiales dan cuenta de una población estable entre 1968 y 1984 con un discreto avance numérico reflejado en los años sucesivos (Mendez y Ferrarini, 2013). Con estas regulaciones de los distintos eventos relacionados con la reproducción se espera que a futuro el efectivo poblacional aumente en forma moderada y con los rasgos más tradicionales que muestran las etnias chaqueñas.
1 ALCARAZ LOPEZ, GM (2000) A fecundidade entre osGuaraní: um legado de kunhankaraí. Tesis Doctoral. http://bases.bireme.br/cgi-bin. [ Links ]
2 ALLAL, N; SEAR, R; PRENTICE, AM; MACE, R (2004) An evolutionary model of stature, age at first birth and reproductive success in Gambian women. Proceedings of the Royal Society of London, Vol. 271: 465-470. [ Links ]
3 BERNIS, C (1990) Canvis en els models de reproducciónhumana: fets i problemasteórics. En: Poblacions, societats i entorns: 260-281. Barcanova. Barcelona [ Links ]
4 BLANCHARD, R; BOGAERT, AF (1997) The relation of closed birth intervals to the sex of the preceding child and the sexual orientation of the succeeding child. Journal of Biosocial Science, 29:111-118. [ Links ]
5 BOHLER, E; BERGSTRÖM, S (1995) Subsequent pregnancy affects morbidity of previous child. Journal of Biosocial Science Vol. 27: 431-442. [ Links ]
6 BONGAARTS, J; POTTER, R (1983) Fertility,biology and behavior. An analysis of the proximates determinants.Academic Press. New York. [ Links ]
7 BUMPASS, L, RIFUSS, R; JONOSIK, R (1978) Age and Marital Status at First Birth and Pace of Subsequent Fertility. Demography, Vol 15 Nº1: 75-80. [ Links ]
8 CAMPBELL, BC; WOOD, JW (1988) Fertility in traditional societies. En: Diggory P. et al., (eds) Natural human fertility. Social and biological determinants: 39-69. MacMillan Press and The Eugenics Society. London. [ Links ]
9 CROGNIER, E (2003) Reproductive Success: Which Meaning? American Journal of Human Biology, Vol. 15: 352-360. [ Links ]
10 CROGNIER, E; ELIZONDO, S; CARATINI, A; ZUBIETA, N; NIBORSKY, R; CARNESE, F (1996) Los comportamientos reproductivos de la población Mapuche de la provincia de Río Negro. Revista Argentina de AntropologíaBiológica, Vol. 1, Nº 1: 86-97. [ Links ]
11 CROGNIER, E; BAALI, A; HILALI, MK (2001) Do “helpers at the nest” increase their parents’ reproductive success? American Journal of Human Biology, 13 (3): 365-373.
12 FELT, JC; RIDLEY, JC; ALLEN, G; REDEKOP, C (1990) High fertility of old colony Mennonites in Mexico. Human Biology 62 (5): 689-700. [ Links ]
13 FERRARINI, SO; MENDEZ, MG (2009) Inversión reproductiva: número, sexo y supervivencia en Chorote y Ayoreo. En: Méndez, MG (ed) Terra incognita: estudios antropológicos en el Chaco Meridional: 56-74. Editorial Universitaria de La Plata. [ Links ]
14 FERRARINI, SO; MENDEZ, MG (2011) Niña o niño: intervalos entre nacimientos y el sexo de los hijos entre los Wichí. Cuadernos Facultad de Humanidades y Ciencias Sociales 39. Suplemento X Jornadas Regionales de Investigación en Humanidades y Ciencias Sociales: 128. [ Links ]
15 GAISIE, SK (1984) The Proximate Determinants of Fertility in Ghana. WFS Scientific Reports, Vol. 53. [ Links ]
16 GRAY, SJ (1994) Comparison of effect of breast-feeding practices on birth-spacing in three societies: Nomadic Turkana, Gainj and Quechua. Journal of Biosocial Science, 26 (1): 69-90. [ Links ]
17 GREENBERG, RA; WHITE, C (1967) The sexes of consecutive sibs in human sibships. Human Biology, Vol. 39:374-404. [ Links ]
18 GREKSA, LP (2002) Population growth and fertility patterns in an Old Order Amish settlement. Annals of Human Biology Vol. 29 Nº 2: 192-201. [ Links ]
19 HAWKES, K; O’CONNEL, JF; BLURTON-JONES, NG; ALVAREZ, H; CHARNOV, EL (1998) Grandmothering, menopause, and the evolution of human life histories.Proceedings of Natural Academy of Science, USA. Anthropology, 95:1336-1339.
20 HENRIPIN, MJ (1954) La fécondité des ménages canadiens au début du XVIIIe siècle. Population, 9: 61-84. [ Links ]
21 HERN, WM (1994) Cultural change, polygyny, and fertility among the Shipibo of Peruvian Amazon. South American Indian Studies, 4: 77-86. [ Links ]
22 HENRY, L (1961) Some data on natural fertility. Eugenics Quaterly, 8: 81-91. [ Links ]
23 HILL, K; HURTADO, AM (1989) Hunther-gatherers of the New World. American Scientist, Nº 77: 437-443. [ Links ]
24 HILL, K; HURTADO, AM (1996) Ache Life History. NY: Aldine de Gruyter. [ Links ]
25 HOBCRAFT, J; MACDONALD, JW; RUTSTEIN, S (1985) Demographic determinants of infant and early child mortality: a comparative analysis. Population Studies, 39: 363. [ Links ]
26 HRDY, SB (1987) Sex-biased investment in primates and mammals. En: Gelles, R; Lancaster, J (eds) Child Abuse and Neglect. Hawthorne, NY: Aldine de Gruyter. 97-147. [ Links ]
27 KAPLAN, HS; LANCASTER, JB (2003) An Evolutionary and Ecological Analysis of Human Fertility, Mating Patterns, and Parental Investment. En: Watcher, K.; Bulatao, R. (eds) Offspring: Human fertility behaviour and biodemographic perspective: 170-223. National Academies Press. Washington D.C. [ Links ]
28 KNODEL, J (1968) Infant mortality and fertility in three Bavarian villages: an analysis of familiy histories from the 19th century. Population Studies, 22: 297-318. [ Links ]
29 KONOGOLO, L (1985): Variation in Entry into Motherhood and Length of Effective Reproductive Life among Women in Kenya. Studies in African and AsianDemography. CDC AnnualSeminar. [ Links ]
30 LIVI-BACCI, M (1993) Introducción a la demografía. Editorial Ariel. Barcelona. [ Links ]
31 MCALPINE, PJ; SIMPSON, NE (1976) Fertility and other demographic aspects of the Canadian Eskimo communities of Igloolik and Hall Beach.Human Biology, Vol. 48 Nº 1: 113-138. [ Links ]
32 MENDEZ, MG; FERRARINI, SO (2009) Rasgos fenotípicos y desempeño reproductivo en hombres y mujeres Chorote. En: Méndez, MG (ed) Terra incognita: estudios antropológicos en el Chaco Meridional: 35-55. Editorial Universitaria de La Plata. [ Links ]
33 MENDEZ, MG; FERRARINI, SO (2011) Madres Wichí: cuándo ser y cuantos niños tener. Cuadernos Facultad de Humanidades y Ciencias Sociales 39. Suplemento X Jornadas Regionales de Investigación en Humanidades y Ciencias Sociales: 228-229. [ Links ]
34 MENDEZ, MG; FERRARINI, SO (2013) Los Chorote en retrospectiva. En: Niños y madres: estudios antropológicos sobre reproducción y fertilidad. Mendez, M.G. (Comp.): 39-84. [ Links ]
35 MILLER, JE (1989) Is the relationship between Barth intervals and perinatal mortality spurious? Evidence from Hungary and Sweden. Population Studies, 43: 479. [ Links ]
36 NATH, DC; LEONETTI, DL; STEELE, MS (2000) Analysis of birth intervals in a non-contracepting Indian population: an evolutionary ecology approach. Journal of BiosocialScience, Vol. 32: 343-354. [ Links ]
37 PASCUAL, J (2004) Caracterización de la historia reproductora y análisis de la fecundidad de las mujeres de Tierra del Fuego. Tesis. Universidad de Barcelona. [ Links ]
38 PEBLEY, AR; HERMALIN, AI; KNODEL, J (1991) Birth spacing and infant mortality: evidence for eighteenth and nineteenth century German villages. Journal of BiosocialScience, 23 (3): 445-459. [ Links ]
39 RODRIGUEZ OTERO, H; BLANCO VILLEGAS; MJ, CARO; L, SÁNCHEZ COMPADRE, E; LÓPEZ MARTÍNEZ, B (2000) Factores asociados a la fertilidad en la comarca leonesa de La Cabrera. En: CARO DOBÓN, L et al., (Eds.) Tendencias actuales de investigación en la Antropología Física Española. Universidad de León, pp. 273-280. España. [ Links ]
40 SÁNCHEZ OCASIO, K (2003) A Fork in Fertility: The Demographic Transition and Factors Mediating a Fertility Decline among the Indigenous Toba of Argentina.Tesis.Harvard University, Cambridge, Massachusetts. [ Links ]
41 SCOTT, S; DUNCAN, CJ (2000) Interacting effects of nutrition and social class differentials on fertility and infant mortality in a pre-industrial population. Population Studies, 54: 71-87. [ Links ]
42 SIEFF, D (1990) Explaining biased sex ratios in human populations – A critique of recent studies. CurrentAnthropology, 31:25-48.
43 SIFFREDI, A (1977) La vida social de los Chorote. Informe preliminar. Cuadernos Franciscanos, Nº 41: 75-99. [ Links ]
44 SMITH, TE (1960) The Cocos-Keeling Islands: a demographic laboratory. Population Studies, 14: 94-130. [ Links ]
45 St.GEORGE, D; EVERSON, P; STEVENSON, JC; TEDROW, L (2000) Birth interval and early childhood mortality in a migrating Mennonite community. American Journal of Human Biology, 12: 50-63. [ Links ]
46 TRIVERS, RL; WILLARD, DE (1973) Natural selection of parental ability to vary the sex ratio offspring. Science, 179:90-92. [ Links ]
47 VALEGGIA, C, ELLISON, P (2004) Lactational amenorrhea in well-nourished Toba women of Formosa, Argentina. Journal of Biosocial Science, 36 (5): 573-595. [ Links ]
48 WESTOFF, CF; POTTER, RG; SAGI, PC; MISHLER, EG (1961) Family Growth in Metropolitan America.Princeton University Press, Princeton, New Jersey. [ Links ]
49 WESTOFF, CF; BLANC, AK; NYBLADE, L (1994) Marriage and entry into parenthood.DHS Comparative Studies Nº 10. Calverton, Maryland: Macro International Inc. [ Links ]
50 WYSHAK, G (1969) Intervals between births in families containing one set of twins. Journal of Biosocial Science, Vol. 1:337-351. [ Links ]

