










Services on Demand
Journal

Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO
Related links
-
 Similars in
SciELO
Similars in
SciELO
Share

 Permalink
PermalinkRIA. Revista de investigaciones agropecuarias
On-line version ISSN 1669-2314
RIA. Rev. investig. agropecu. vol.39 no.3 Ciudad Autónoma de Buenos Aires Dec. 2013
ARTÍCULOS
Calidad nutritiva de láminas de festuca alta (Festuca arundinacea Schreb) en rebrotes de verano y otoño
Insua, J.R.1; Di Marco, O.N.1; Agnusdei, M.G.2
1Facultad de Ciencias Agrarias, Universidad Nacional de Mar del Plata.
2Instituto Nacional de Tecnología Agropecuaria, EEA Balcarce. Correo electrónico: insua_juan@hotmail.com
Recibido el 27 de noviembre de 2012
Aceptado el 9 de octubre de 2013
Publicado online el 23 de octubre de 2013
RESUMEN
Se evaluó el efecto de la morfogénesis y estructura foliar en la calidad de las láminas de rebrotes de verano y otoño de Festuca arundinacea Schreb. Se utilizó un diseño completamente aleatorizado (n=3) en 250 macetas al aire libre sin limitantes hídricas de N y P. En los rebrotes de cada estación se realizaron tres cosechas de 300 macollos/repetición para obtener láminas en tres estados de desarrollo (recientemente expandida, adulta y pre-senescencia), correspondientes a tres generaciones consecutivas de hojas. En cinco macollos marcados por repetición se midió: la tasa de elongación foliar (TEF), el intervalo de aparición de hoja (IAH), la vida media foliar (VMF), el número de hojas vivas (NHV) y el largo de lámina y vaina. En las láminas se determinó el contenido de fibra detergente neutro (FDN), la digestibilidad de la FDN (DFDN) y la digestibilidad in vitro aparente de la materia seca (DAMS) (24 h de incubación, digestor DaisyII, ANKOM). Las variables de morfogénesis se evaluaron como medidas repetidas en el tiempo, y las de calidad ajustando funciones lineales con la edad y largo foliar. Las comparaciones se realizaron mediante análisis de variables Dummy y ANVA (Tukey). La temperatura media diaria del rebrote de otoño fue menor que en verano (13±4 vs. 21±3ºC). Los parámetros de morfogénesis TEF, VMF, IAH fueron menores (p<0,001) en otoño que en verano, pero el NHV no fue afectado (p>0,23). La longitud foliar en otoño no varió durante el rebrote y fue menor que en verano (106±10 vs 239±100 mm); que aumentó entre generaciones sucesivas (~110 mm/generación) en estrecha asociación (R²=93%) con el aumento progresivo de la longitud de vaina. Las láminas de otoño presentaron menor contenido de FDN (39,8±0,8 vs. 55,3±0,6%), mayor DFDN (52,7±1,6 vs. 46,8±1,1%) y DAMS (69,3±0,9 vs. 58,7±0,6%%) que las de verano. Se concluye que los rebrotes de verano y otoño difirieron en variables de morfogénesis, estructura y calidad. Asimismo, las diferencias en digestibilidad de la MS estuvieron asociadas al efecto del largo foliar en la dinámica de la FDN y DFDN.
Palabras clave: Morfogénesis; FDN; DFDN; Edad foliar; Largo foliar; Aina.
ABSTRACT
The effect leaf morphoenesis and structure onleaf bades quality was evaluated in summer and autumn regrowths of Festuca arundinacea Schreb. The experiment was conducted in 250 pots arranged in a completely randomised design (n=3) under natural climate conditions, without limitation of water, N and P. In each season swards were sampled 3 times (~300 tillers/replicate) to obtain leaves in 3 ontogenics stages of similar age (just expanded, adult and pre-senescence) corresponding to the 3 first sequentially appearing leaf generations. The leaf elongation rate (LER), leaf app earance interval (LAI), leaf life span (LLS), numb er of living leaves (NLL) and length of the leaf blade and sheath were measured in 5 marked tillers/replicate. In leaf blades, the NDF and the in vitro digestibility (NDFD and DMD) were determined (incubation at 24 h DaisyII digestor). The mo rphog enic variables were evaluated using repeated measures data analysis. Quality traits were analyzed by lineal function with age and leaf length. Comp arisons were made by dummy variable regressions and ANOVA (Tukey). The mean daily temp erature was low er in autumn than in summ er (13±4 vs. 21±3ºC). LER, LLS and LAI were low er (p<0.001) in autumn than in summ er but NLL was similar (p>0.23) in bo th season. Leaf length did not vary during autumn regrow th, but it was shorter than summ er (106±9 vs 239±98 mm ). Leaf blade length increased during summ er amo ng successive leaves of regrow th (~110 mm /generation) in relation (R²=93%) to the prog ressive in crease of the sheath tube length. Autumn leaf blades show ed low er NDF (39,8±0,8 vs 55,3±0,6%), higher NDFD (52,7±1,6 vs 46,8±1,1%) and DMD (69,3±0,9 vs 58,7±0,6%%) than that of summ er regrow th. It is concluded that summ er and autumn regrow ths differ in mo rphog enic, structure and quality variables, and that differences in DM digestibility were associated with the effect of leaf length on NDF and NDFD dynamics.
Keywords: Morphogenesis; NDF; DNDF; Leaf age; Leaf length; Sheath.
INTRODUCCIÓN
Festuca alta (Festuca arundinacea Schreb.) es una gramínea perenne que se adapta muy bien a suelos de baja aptitud agrícola, cuyo rol en los sistemas ganaderos del país puede ser importante en la medida en que se demuestre su capacidad para producir forraje de alta calidad. No obstante, esta especie rústica de excelentes características productivas, disminuye drásticamente su calidad nutritiva en primavera avanzada y verano (Burns, 2009).
La caída continua de calidad entre hojas sucesivas del rebrote se debe a la pérdida de digestibilidad de la fibra detergente neutro (DFDN) con el incremento del largo foliar (Groot y Neuteboom, 1997; Duru y Ducroq, 2002; Agnusdei et al., 2011; Insua et al., 2012; Di Marco et al., 2013). Se demostró que el aumento de la frecuencia de defoliación (Donaghy et al., 2008) y la disminución de la altura de la pastura (Di Marco et al., 2013) permiten mejorar la calidad del forraje en pasturas en estado vegetativo.
Los antecedentes citados llevan a plantear como hipótesis que las condiciones térmicas contrastantes de otoño y verano afectan, diferencialmente, el largo foliar entre hojas sucesivas del rebrote y, en consecuencia, la calidad del forraje, independientemente de la estación del año. El objetivo de este trabajo fue estudiar las características morfogenéticas y estructurales durante rebrotes de verano y otoño de Festuca arundinacea y su relación con la calidad de las láminas en ambas estaciones.
MATERIALES Y MÉTODOS
El ensayo se llevó a cabo en la Estación Experimental Agropecuaria (EEA) INTA de Balcarce (sudeste Bonaerense, 37º 45' S; 58º 18' W) durante el verano de 2009/10 y el otoño de 2011. Se utilizaron plantas de Festuca arundinacea Schreb. (cv El Palenque Plus INTA) sembradas en macetas de plástico (250 macetas de 20 cm de diámetro y 40 cm de altura), con sustrato extraído del horizonte A de un suelo Argiudol típico, en un diseño completamente aleatorizado (DCA, n=3). El ensayo fue al aire libre sin limitantes hídricas ni nutricionales. Se aplicó riego por aspersión con frecuencia variable para mantener las condiciones de capacidad de campo. Además, se realizó una fertilización inicial con Nitrato de Amonio Calcáreo (150 kg N ha-1) y Fosfato Diamónico (50 kg P ha-1), más fertilizaciones semanales de Nitrato de Amonio Calcáreo (35 kg N ha-1) durante todo el ensayo. Antes del inicio del período experimental de cada estación, se realizaron tres cortes a 5 cm para homogenizar la edad de macollos y formar micropasturas densas y foliosas.
Previo al muestreo se realizó un último corte de homogenización y sincronización del crecimiento a 5 cm de altura que dio comienzo al período experimental. Se registró la temperatura media diaria, la cual se mantuvo relativamente constante durante el verano (21±3ºC), y disminuyó durante el otoño en un rango de 21 - 4ºC (13±4ºC). El tiempo se expresó como grados días de crecimiento (GDC), que se calculó como la acumulación de la temperatura media diaria por encima de una temperatura base de 4°C. Se registró el intervalo de aparición de hojas (IAH), la tasa de elongación foliar (TEF), la vida media foliar (VMF), el número de hojas vivas por macollo (NHV), y el largo de lámina y vaina con una frecuencia de una a tres veces por semana en 15 macollos. La VMF se midió como los GDC acumulados transcurridos entre la aparición de la lámina visible y el comienzo de la senescencia.
Paralelamente, se realizaron tres cosechas (300 macollos/ repetición) para obtener láminas en tres categorías de edad (recientemente expandida, adulta y pre-senescente) correspondientes a las tres primeras generaciones de hojas (G1, G2, G3) acumuladas durante los rebrotes. Los macollos se cortaron a nivel de la base y se congelaron, inmediatamente, en N líquido. Luego, se almacenaron en freezer (-20°C) hasta su posterior separación y procesado. A continuación, se descongelaron y se mantuvieron en bandejas sobre hielo durante la separación de las láminas por categorías de edad y generaciones.
Las láminas se liofilizaron y molieron con un molino tipo Ciclotec con malla de 1 mm. Luego se determinó el contenido de fibra detergente neutro (%, FDN, Van Soest et al., 1991) y la digestibilidad de la FDN (%, DFDN) y de la verdadera de la materia seca (%, DVMS) por incubación in vitro de 250 mg de muestra durante 24 horas de incubación en el equipo DaisyII. La DVMS se convirtió a digestibilidad aparente (DAMS), restando el factor metabólico 11,9 propuesto por Van Soest (1994).
El conjunto de datos se analizó con un modelo estadístico lineal para un diseño DCA con tres repeticiones. Las variables de morfogénesis se analizaron como mediciones repetidas en el tiempo tomando en cuenta a los macollos marcados como sujetos de estudio y a la estación del año como factor de tratamiento. Para las variables de calidad se evaluaron ambas estaciones del año realizando ajustes de funciones lineales con los GDC y comparaciones mediante análisis de variables dummy. Los datos se analizaron con los procedimientos PROG REG y PROC GLM de la versión 8 del paquete estadístico SAS (SAS, 2001). Se efectuó un análisis de la varianza para un DCA de las variables de interés y se comparó las medias con la prueba de Tukey (α=0,05).
RESULTADOS
Morfogénesis y largo foliar
Durante el crecimiento y desarrollo de las tres primeras hojas sucesivas acumuladas durante el rebrote (G1, G2 y G3), los parámetros de morfogénesis TEF, VMF e IAH fueron mayores en verano que en otoño. Sin embargo, el máximo NHV acumuladas no se diferenció entre estaciones (tabla 1).
Tabla 1. Variables de morfogénesis de Festuca arundinacea durante el otoño y verano. Letras diferentes indican diferencias significativas entre épocas del año (p<0,05). ±: Error estándar de la media (n=3).

El largo de lámina del rebrote de otoño fue menor (p<0,0026) que en verano y se mantuvo constante (106±10 mm) entre generaciones sucesivas de hojas. En cambio, en verano aumentó (p<0,0001) desde 136±11, 222±24 a 360±53 mm para G1, G2 y G3, respectivamente. El aumento del largo de lámina de las hojas sucesivas del rebrote estival mostró una fuerte relación positiva (R²=93%) con el incremento progresivo del largo del tubo de vainas (figura 1), el cual se mantuvo relativamente constante en otoño (47±10 mm).
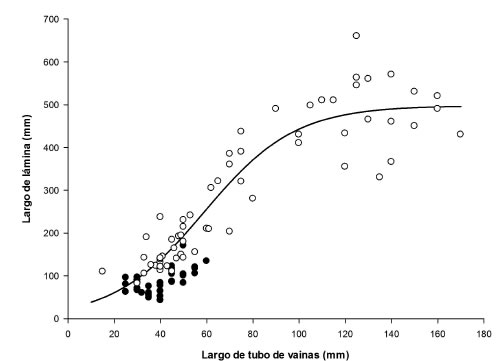
Figura 1. Relación entre el largo de lámina y del tubo de vainas de hojas recientemente expandidas de Festuca arundinacea durante los rebrotes de otoño (?) y verano (.). Otoño no significativo (p>0,05).
Calidad de las láminas al mismo estado ontogénico
Las láminas de otoño tuvieron menor FDN y mayor DAMS y DFDN que las de verano, comparadas a un mismo estado ontogénico foliar (tabla 2). La DAMS y la DFDN disminuyeron (p<0,0001) con el progreso de la ontogenia foliar, pero la FDN se mantuvo (p>0,86) en un promedio de 39,8±0,79 en otoño y de 55,3±0,59 % en verano.
Tabla 2. Contenido de FDN y digestibilidad de la FDN (DFDN) y de la MS (DAMS) de láminas de rebrotes de Festuca arundinacea en diferentes estados ontogénicos (promedio de tres generaciones sucesivas de hojas). Letras diferentes indican diferencias significativas entre épocas del año (p<0,0001). ±: Error estándar de la media (n=3).

Efecto de la edad y largo foliar sobre la calidad de las láminas
El efecto de la edad foliar sobre la calidad de las láminas se analizó durante el desarrollo (desde recientemente expandida hasta pre senescencia) de hojas con longitud similar (G1). El contenido de FDN se mantuvo sin cambios con la edad foliar (GDC), pero fue menor (p<0,0001) en otoño que en verano (36,1±3 vs 52,9±3%). Las hojas de la G1 mostraron similar DFDN en ambas épocas (promedio 54,3%), pero ésta disminuyó con la edad foliar (figura 2a) a una mayor tasa en otoño que en verano (paralelismo p<0,006).
Figura 2. Evolución de la DFDN con la edad foliar (a) y largo de lámina (b) de hojas de Festuca arundinacea durante los rebrotes de otoño (negro) y verano (blanco). En a: G1 recientemente expandida, ?.; adulta, ■□; y pre senescente, ▲Δ. En b: hojas recientemente expandidas de G1, ?.; G2, ■□; G3, ▲Δ. DFDN = -0,06±0,009x + 68±1,6, R² = 0,73, p<0,0001 (n=27).
El largo de lámina afectó diferencialmente la FDN, DFDN y DAMS de hojas recientemente expandidas (coincidencia p<0,0001). Cabe aclarar que para la evaluación del efecto del largo foliar se utilizó hojas de similar estado ontogénico (recientemente expandidas) para eliminar efectos confundidos debidos a la edad foliar. Durante el otoño, la FDN se mantuvo constante (40,2±0,8%), mientras que en verano aumentó de 52 a 59% (0,2%/cm; p<0,02), resultando un valor promedio de 55,3±0,6%. A su vez, la DFDN fue más afectada que la FDN por el aumento del largo de lámina debido a que mostró una pérdida de 0,6%/cm durante el verano (figura 2b). La DFDN de las láminas de ambas estaciones se ajustaron a una misma función con el largo foliar (coincidencia p=0,9). Sin embargo, en otoño los datos se agruparon en los máximos niveles de DFDN debido a que no se observaron cambios en la longitud foliar.
DISCUSIÓN
Crecimiento y recambio foliar
Los rebrotes de otoño y verano crecieron en diferentes condiciones de temperatura, lo cual afectó diferencialmente los parámetros de morfogénesis y estructurales en estudio. En otoño, el crecimiento (TEF) resultó ~5 veces menor que en verano y el recambio foliar fue más rápido (menor VMF e IAH). A pesar de estas diferencias, en ambas épocas se acumuló un máximo de tres hojas vivas por macollo (tabla 1).
La menor VMF de otoño (444 vs 630 GDC) coincide con resultados de Lemaire y Agnusdei (2000) que registraron menores valores durante el invierno-otoño. Al igual que en el presente trabajo, estos autores no encontraron explicaciones claras a las diferencias estacionales en la VMF debido a que se considera una característica de carácter genético y, por ende, inherente a la especie (Lemaire y Chapman 1996).
Aunque las hojas de otoño tuvieron un ciclo de vida un 30% menor (<VMF) que las de verano, el NHV fue similar (~3 hojas) en ambas estaciones. Esto es debido a que en otoño las hojas aparecieron con mayor frecuencia, es decir, un IAH 29% más corto que en verano. Por lo tanto, el cociente VMF/IAH que determina el NHV (Lemaire y Agnusdei 2000) permaneció constante.
La dinámica del crecimiento de las hojas fue diferente entre estaciones, lo cual afectó su estructura. En otoño, las hojas fueron más cortas que en verano y su longitud permaneció constante a través de las generaciones de hojas sucesivas del rebrote. En cambio, en verano la longitud aumentó entre generaciones de hojas (~110 mm), en estrecha asociación (R²=93%) con el aumento progresivo del largo del tubo de vainas (figura 1). Ambas variables, longitud de hoja y tubo de vainas, se mantuvieron relativamente constantes en otoño. Skinner y Nelson (1995) y Kavanová et al. (2006), señalan que el tamaño del tubo de vainas controla el largo de hoja a través de un efecto positivo sobre la tasa y la duración de la elongación foliar.
Calidad láminas
La calidad promedio de las láminas de otoño fue superior a las de verano (tabla 2). En otoño, la DAMS y la DFDN fueron 18 y 12% mayores que en verano, respectivamente; y el contenido de FDN 28% menor (tabla 2).
El menor contenido de FDN en otoño (39 vs 55%) estaría relacionado al menor crecimiento otoñal (ocasionado por las menores temperaturas) que produciría una menor demanda de fotoasimilatos, y la consecuente acumulación de contenido soluble y disminución relativa de la pared celular. Al respecto, Lemaire y Agnusdei (2000) y Burns et al., (2002), también observaron una disminución de la FDN y mayor acumulación de hidratos de carbono solubles en otoño que en épocas más favorables para el crecimiento. Por otro lado, la constancia de la FDN con la edad (VMF) y la escasa variación con el largo foliar concuerda con experimentos previos en otras especies (Avila et al., 2010; Agnusdei et al., 2011; Insua et al., 2012).
Debido a la constancia, o escasa variación, de la FDN con la edad y largo foliar, los cambios de DAMS durante el desarrollo foliar dependieron en mayor medida de la dinámica de la DFDN (tabla 2). La edad foliar mostró un efecto negativo sobre la DFDN que fue similar en ambas estaciones. Éste fue de ~19 unidades porcentuales (de ~62 a ~43%) durante la VMF, es decir, desde que la hoja se expandió hasta pre-senecencia (figura 2a). Esto indicaría que, en hojas de similar largo y estado de desarrollo, no habría diferencias en DFDN entre estaciones del año. Similar comportamiento fue observado por Insua et al. (2012) al comparar láminas de cultivares de festuca alta. Sin embargo, debido a la menor VMF de otoño, la tasa de pérdida de DFDN con la edad foliar expresada en tiempo térmico (GDC), fue mayor que en verano (figura 2a). Los resultados sugieren que los procesos físicos (Wilson y Mertens, 1995) y químicos (Akin 1989; Jung y Allen, 1995), que hacen a la pared celular menos digestible, se aceleran con el aumento del recambio foliar, es decir, cuando se acorta el IAH y/o la VMF.
El largo foliar mostró un efecto negativo sobre la DFDN de las láminas de ambos rebrotes (figura 2b), con una pérdida de 0,6 unidades porcentuales cada 10 mm de incremento en la longitud en ambas épocas del año. Este valor es similar al encontrado en especies C4 por Agnusdei et al. (2011) y Avila et al. (2010). Debido a este efecto sobre la DFDN, el aumento del largo foliar de verano fue el determinante de las mayores pérdidas de calidad durante el rebrote, en comparación con el otoño donde la longitud de las hojas se mantuvo relativamente constante.
CONCLUSIONES
Los rebrotes de otoño y verano se diferenciaron en parámetros de morfogénesis y estructurales, como así también en la calidad de las láminas, la cual fue mayor en otoño. En esta estación el porcentaje de FDN fue menor que en verano (39 vs 54%) y se mantuvo constante con la edad foliar en ambas estaciones. Sin embargo, la FDN aumentó levemente con el largo foliar en verano (0,2%/cm). La DAMS fue mayor en otoño que en verano (18%) y disminuyó diferencialmente durante el rebrote en ambas estaciones. La disminución de calidad durante el rebrote de otoño fue debido a la pérdida de DFDN con la edad foliar, dado que el largo de las hojas sucesivas del rebrote no aumentó. En cambio, en verano, la DFDN disminuyó tanto con la edad, como así también con el aumento del largo foliar entre hojas sucesivas del rebrote. Este aumento estuvo asociado al incremento progresivo del largo del tubo de vainas de verano. Los resultados indican que las variaciones en DFDN son más importantes que las de FDN para explicar los cambios de DAMS durante el rebrote. Además, las variaciones del largo foliar son las principales determinantes de los cambios estacionales de calidad.
BIBLIOGRAFÍA
1. AGNUSDEI, M.G.; DI MARCO, O.N.; NENNING, F.R.; AELLO, M.S. 2011. Leaf blade nutritional quality of rhodes grass (Chloris gayana) as affected by leaf age and length. Crop and Pasture Sci. 62, 1098-1105. [ Links ]
2. AKIN, D.E. 1989. Histological and physical factors affecting digestibility at forages. Agron. J. 1:17-25. [ Links ]
3. AVILA, R.E.; DI MARCO, O.N.; AGNUSDEI, M.G.; MAYORAL, C. 2010. Digestibilidad de la fibra y materia seca de dos gramíneas megatermicas (Chloris gayana y Cenchrus ciliaris) de diferente porte. Relación con la edad y largo foliar. Rev. Arg. Prod. Anim. 30: 1-13. [ Links ]
4. BURNS, J.C. 2009. Nutritive value. In: Fribourg, H.A.; Hannaway, D.B.; West, C.P. (eds.). Tall fescue for the Twenty-first Century. Agron. Monogr. 53. ASA, CSSA, and SSSA, Madison, WI. pp.159-202. [ Links ]
5. BURNS, J.C.; CHAMBLEE, D.S.; GIESBRECHT, F.G. 2002.Defoliation intensity effects on season long dry matter distribution and nutritive value of tall fescue. Crop Science, 42, 1274-1284. [ Links ]
6. CALLOW, M.N.; LOWE, K.F.; BOWDLER, T.M.; LOWE, S.A.; GOBIUS, N.R. 2003. Dry matter yield, forage quality and persistence of tall fescue (Festuca arundinacea) cultivars compared with perennial ryegrass (Lolium perenne) in a subtropical environment. Aust. J. Exp. Agric. 43:1093-1099. [ Links ]
7. DI MARCO, O.N.; HARKES, H.; AGNUSDEI, M.G. 2013. Calidad de agropiro alargado (Thinopyrum ponticum) en estado vegetativo en relación a la edad y longitud de las hojas. Rev. Investigación Agropecuaria (RIA). 39:105-110. [ Links ]
8. DONAGHY, D.J.; TURNER, L.R.; ADAMCZEWSKI, K.A. 2008. Effect of defoliation management on water-soluble carbohydrate energy reserves, dry matter yields, and herbage quality of Tall Fescue. Agron. J. 100:122-127. [ Links ]
9. DURU, M.; DUCROCQ, H. 2002. A model of lamina digestibility of orchardgrass as influenced by nitrogen and defoliation. Crop Sci. 42: 214 - 223. [ Links ]
10. GROOT, J.C.; NEUTEBOOM, J.H. 1997. Composition and digestibility during ageing of Italian ryegrass leaves of consecutive insertion the same levels. J. Sci. Food Agric. 75: 227-236. [ Links ]
11. INSUA, J.R.; AGNUSDEI, M.G.; DI MARCO, O.N. 2012. Calidad nutritiva de láminas de dos cultivares de "festuca alta" (Fes tuca arundinacea Schreb). Rev. Investigación Agropecuaria (RIA). 38(2): 190-195. [ Links ]
12. JUNG, H.J.; ALLEN, M.S. 1995. Characteristics of plant cell walls affecting intake and digestibility of forages by ruminants. J. Anim. Sci. 73: 2774-2790. [ Links ]
13. KAVANOVÁ, M.; GRIMOLDI, A.A.; LATTANZI, F.A.; SCHNYDER, H. 2006. Phosphorus nutrition and mycorrhiza effects on grass leaf growth. P status- and size-mediated effects on growth zone kinematics. Plant, Cell Environm. 29: 511-520. [ Links ]
14. LEMAIRE, G.; CHAPMAN, D. 1996. Tissue flows in grazed plant communities. En: HODGSON, J.; ILLIUS, A.W. (Eds.) The ecology and management of grazing system. CAB International. pp. 3-36. [ Links ]
15. LEMAIRE, G.; AGNUSDEI, M. 2000. Leaf tissue turnover and efficiency of herbage utilization. En: LEMAIRE, G.; HODGSON, J.; de MORAES, A.; CARVALHO, P.C. de F.; NABINGER, C. (Eds.) Grassland ecophysiology and grazing ecology. CAB International. pp. 265-287. [ Links ]
16. SAS. 2001. SAS/STAT® User's guide (Release 8.0). SAS Inst. Inc., Cary, NC. [ Links ]
17. SKINNER, R.H.; NELSON, C.J. 1995. Elongation of the grass leaf and its relationshipto the phyllochron. Crop Sci. 35: 4-10. [ Links ]
18. VAN SOEST, P.J. 1994. Nutritional ecology of the ruminant. Cornell University Press, Ithaca, NY. 476 p. [ Links ]
19. VAN SOEST, P.; ROBERTSON, J.; LEWIS, B. 1991. Methods for dietary fiber, neutral detergent fiber and nonstarch polysaccharides in relation to animal nutrition. J. Dietary Sci. 74: 3583 -3597. [ Links ]
20. WILSON, J.R.; MERTENS, D.R. 1995. Cell wall accessibility and cell structure limitations to microbial digestion of forage. Crop Sci. 35:251-259. [ Links ]

