










Servicios Personalizados
Revista

Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkRIA. Revista de investigaciones agropecuarias
versión On-line ISSN 1669-2314
RIA. Rev. investig. agropecu. vol.44 no.1 Ciudad Autónoma de Buenos Aires abr. 2018
ARTÍCULOS
Tolerancia a las heladas por superenfriamiento en cultivares de nogal (Juglans regia) y almendro (Prunus amygdalus)
Rodríguez, A.1; Benítez Piccini, E.1; Muñoz, A.1
1 Instituto Nacional de Tecnología Agropecuaria (INTA), Estación Experimental Agropecuaria (EEA) Alto Valle, Ruta Nacional 22, km 1190, CC782, CPc8332, General Roca, Río Negro, Argentina. Correo electrónico: rodriguez.andrea@inta.gob.ar
Recibido 15 de diciembre de 2016
Aceptado 02 de septiembre de 2017
Publicado online 15 de mayo de 2018
RESUMEN
El nogal y el almendro son cultivados en los valles irrigados de Patagonia norte. El daño por heladas es el estrés ambiental más importante que afecta la productividad de estos cultivos. El período de riesgo por heladas se extiende desde el otoño hasta finales de primavera (abril a octubre). Las plantas pueden evitar el daño por congelamiento al prevenir la formación de hielo extracelular bajando la temperatura de congelamiento (superenfriamiento). Nuestros objetivos fueron: 1) evaluar los cambios en el grado de resistencia antes y durante el invierno en segmentos de tallos de un año de edad de nogales (cv Chandler y Franquette) y 2) determinar los umbrales de superenfriamiento en flores de almendro (cv Guara, Ferragnes y Farraduel). Para el estudio se comparó el análisis térmico y la evaluación del tejido sano después del enfriamiento controlado, basado en el amarronamiento del tejido dañado. Nuestras observaciones y resultados indican que para nogal el grado de tolerancia a las heladas depende de la historia climática previa. Durante fines de verano y otoño el valor de temperatura de nucleación del hielo (TNH) es de -5 ºC, alcanzando la LT50 a -7,7 ºC y 100% de daño con temperaturas mínimas de -10 ºC por más de una hora. En pleno reposo invernal la TNH es de -6,5 ºC y se llega al 25% de daño con temperaturas mínimas de -15 ºC. La desaclimatización inducida disminuye la tolerancia del tejido (TNH de -5,6 a -4,6 ºC) y aumenta la variabilidad de respuesta a las bajas temperaturas, con heladas de -8 ºC el grado de daño puede ser del 10 al 50% y con -15 ºC de 25 al 100%. En madera joven de nogal, la concentración de azucares solubles es el principal indicador y regulador fisiológico de la tolerancia a las heladas. Las flores de almendro tienen un TNH de -1,7º a -2,5 ºC. La exposición por más de una hora a estas temperaturas provoca la muerte del 45% de los órganos. Si el período prefloración es fresco, el grado de desaclimatación es menor, los valores de TNH disminuyen a -3,8 ºC otorgándole mayor tolerancia a las heladas.
Palabras clave: Desaclimatación; Madera de nogal; Flores de almendro.
ABSTRACT
Walnuts and almond grown in irrigation valley of Patagonia. The freezing damage is one of the most important environmental stresses that affect productivity. The periods of frost risk is from autumn to late spring (April to October). Plants can avoid freezing damage by preventing extracellular ice formation below the equilibrium freezing temperature (supercooling). The objectives were to: 1) evaluate changes in the degree of hardening before and during winter in walnuts stem segments of one-year-old (cv Chandler y Franquette); 2) determine the threshold for supercooling of almond blossom (cv Guara, Ferragnes y Farraduel). To study were compared thermal analysis and the evaluation of tissue health after controlled freezing, based on tissue browning. Our observations and results indicate that the degree of walnut frost tolerance depends on the previous climate history. During late summer and autumn the value TNH is -5ºC, reaching the LT50 to -7.7ºC and 100% damage with minimum temperatures of -10°C for more than an hour. In the winter rest TNH is -6.5ºC and reaches 25% damage with minimum temperatures of -15°C. The desaclimatización induced decreases tissue tolerance (TNH -4.6ºC to -5.6ºC) and increases variability in response to low temperatures, with frost -8ºC the degree of damage can be from 10 to 50% and -15ºC of 25 to 100%. Young walnut wood, the concentration of soluble sugars is the main indicator and physiological regulator of frost tolerance. Almond flowers have TNH of -1.7ºC to -2.5°C. Exposure for more than an hour at these temperatures causes the death of 45% of the organs. If the pre-flowering period is fresh, the degree of desaclimation is lower, TNH values decrease -3.8ºC giving it greater tolerance to frost.
Keywords: Deacclimation; Walnut stem; Almond flowers.
INTRODUCCIÓN
Producción de nogales y almendros
En la Patagonia argentina el cultivo de nogales (Juglans regia) y almendros (Prunus amygdalus) se concentra en los valles irrigados de las provincias de Río Negro y Neuquén. Actualmente la superficie plantada de nogales es de 1.580 hectáreas, principalmente desarrollada en Valle Inferior, Valle Medio y Alto Valle (Iannamico, 2009; Iannamico, 2015a). En lo que respecta al cultivo de almendros, existen 230 ha principalmente en Alto Valle. Los dos cultivos se adaptan a las condiciones climáticas de la región (semidesértica), siendo el factor de mayor riesgo las heladas. Para los nogales tienen mayor relevancia las heladas otoñales (marzo-abril) que afectan a plantas jóvenes y a la madera joven de plantas adultas. Las heladas tardías o primaverales (agosto-septiembre-octubre) son más preocupantes para el cultivo de almendro, dado que afectan la floración (Iannamico, 2015b).
Caracterización de heladas
Los valles patagónicos se caracterizan por estar en un clima semidesértico con un período libre de heladas de 180 a 210 días. El período con heladas se extiende desde la primera quincena de abril hasta primera quincena de octubre. La frecuencia anual media de heladas en el valle irrigado de Río Negro es de 67,5 días. El 12% del total de las heladas anuales corresponde a las tardías. Estas, durante los meses de agosto, septiembre y octubre, son mucho menos frecuentes que las invernales y otoñales, pero de mayor interés agronómico dada la mayor susceptibilidad de los tejidos, afectando flores y frutos en el caso de almendros y el desarrollo de brotes y yemas que portan las flores pistiladas en nogal. Si bien la frecuencia de heladas otoñales es menor al 4% entre marzo y abril, pueden provocar serios daños en tejidos jóvenes que aún no han agostado. La intensidad y duración de las heladas es variable, registrándose temperaturas mínimas absolutas de hasta -13 ºC en el trimestre invernal y de -8 ºC en el período primaveral con una durabilidad variable, pero que puede en algunos casos superar 7 horas (Rodríguez y Muñoz, 2006). La duración de un evento de helada contempla el número de horas en que la temperatura del aire está por debajo de los 0 ºC. Los daños físicos y económicos son muy importantes, disminuyendo notablemente el rendimiento potencial y retardando la entrada en producción de cultivos jóvenes.
Características ecofisiológicas de las especies a las bajas temperaturas
En los vegetales existe más de un mecanismo para tolerar las bajas temperaturas y evitar el daño por congelamiento; pueden sobrevivir a temperaturas de congelamiento evitando o tolerando la formación de hielo extracelular (Levitt, 1978). Los árboles frutales de hoja caduca presentan este tipo de estrategias evitando en un primer momento la formación de hielo extracelular a través del superenfriamiento y posteriormente tolerando ese congelamiento con el inicio del proceso de deshidratación celular (Aslani Aslamarz et al., 2010; Meng et al., 2007; Mathers, 2004). Los umbrales para cada especie y variedad son diferentes y además cada tejido se comporta de manera específica (Proebsting y Mills, 1978; Ashworth y Wisniewski, 1991). El objetivo de nuestro trabajo es evaluar el comportamiento de la madera joven de nogal a las heladas ante diferentes condiciones fisiológicas de aclimatación y en almendros determinar el punto crítico de daño por temperaturas subcero en el estado de plena floración para distintas variedades.
MATERIALES Y MÉTODOS
Para la simulación de heladas se utilizó un prototipo de enfriamiento controlado (Rodriguez et al., 2012) el cual consiste en módulos termoeléctricos (celdas peltier 12V- 63 watts) para enfriar un recipiente de metal (cámara) aislado térmicamente. El conjunto incluye un disipador de calor y se encuentra alojado dentro de un refrigerador que debe mantenerse a temperatura estable. La precisión de las mediciones fue de ±0,1 ºC, a un valor de temperatura de 0 ºC, con un error fijo y por lo tanto compensable, de aproximadamente 0,15 ºC. El registro térmico se realizó de forma individual para cada muestra (tallo o flor) con 24 termocuplas tipo T de cobre-costantan de 1 mm de diámetro, instaladas a un data logger DLZe (Delta T Devices). En los tallos se realizó una pequeña perforación lateral para colocar la termocupla y en la flor se colocó dentro del cáliz. La tasa de enfriamiento fue de 2 a 3 ºC h-1 desde los 0 ºC hasta llegar a la temperatura mínima preestablecida para cada tejido. El almendro (Prunus amygdalus L.) se evaluó durante septiembre del año 2011 y 2013 con temperaturas mínimas de -2,7 ºC y -4 ºC. Las flores de almendro permanecieron por treinta minutos a esa temperatura. En cuanto a la madera de nogal se trabajó con temperaturas mínimas de -5 ºC a -15 ºC; el lapso de exposición fue de una hora; excepto en las heladas simuladas en julio que tuvieron una duración de 4 horas. Las curvas de enfriamiento fueron expresadas como temperatura en función del tiempo en horas. Las muestras retiradas de la cámara de enfriamiento se dejaron 24 horas a temperatura ambiente (20º a 22 ºC) con un 100% de humedad relativa. Después de cada simulación se determinó el daño producido en los tejidos a través de la técnica de observación visual por amarronamiento utilizando una lupa (Nikon 10X/21). Para esto se utilizaron cortes transversales de las muestras de madera. En cuanto a las flores se las observo enteras para identificar muerte total del gineceo y posteriormente se realizó un corte longitudinal de este. La muerte celular puede tener lugar después de una fase de superenfriamiento y posterior formación de cristales de hielo. La temperatura a la cual ocurre el congelamiento del agua es denominada de nucleación del hielo (TNH). Por un lado, esta se observó a través de las curvas de enfriamiento por la ocurrencia de un eventual incremento de la temperatura del tejido debido al exotermo (pérdida de calor por el cambio de estado del agua intercelular). Por otro lado, se determinó en los tejidos el contenido de almidón y azúcares solubles por el método descripto por Southgate (1991); la absorbancia fue medida a 600 nm en un espectrofotómetro (SP 1105, Spectrum, China). El contenido relativo de agua se determinó por diferencia entre peso seco y húmedo de las muestras. La temperatura letal media (LT50) se determinó por interpolación a través de una curva de respuesta a diferentes valores de temperatura mínima.
Se trabajó con almendro (Prunus amygdalus L.) cultivares Guara, Ferragnes y Farraduel en estado de plena floración. En nogal (Juglans regia L.) se evaluó madera joven de un año de edad para las variedades Chandler y Franquette. En este último caso se evaluó material aclimatado (otoño-invierno) y desaclimatado (verano-invierno). En cuanto al material desaclimatado se evaluó el comportamiento de tejido en estado vegetativo (verano, principios de marzo) y tejido aclimatado, por estar en estado de reposo invernal, que fue desaclimatado artificialmente. Para modificar el grado de aclimatación del material en invierno se lo sometió a 120 horas (5 días) a temperaturas de 18 ºC. También se evaluó el comportamiento del nogal según su lugar de cultivo, Alto Valle-Allen (AV) y Valle Medio-L. Beltrán (VM), dos zonas ubicadas a 200 km de distancia. Las dos zonas en estudio difieren en el régimen diario de temperaturas, principalmente en los valores de mínimas, y por ende existen diferencias en la acumulación de frío y la acumulación de calor asociada al inicio del ciclo de crecimiento. Las variables climáticas analizadas para el cálculo del índice de desaclimatación en almendros pertenecen al registro de la estación meteorológica de la Estación Experimental Alto Valle del INTA (INTA EEA Alto Valle) (Allen, -39º01 Lat. Sur; -67º66 Long. Oeste; 242 m s.n.m. El diseño del muestreo fue completamente aleatorizado, con un n=12 y 2 repeticiones por temporada de estudio. Los factores evaluados fueron aclimatación, variedad, origen (AV/VM). El material se extrajo sobre un total de 20 árboles adultos, tanto en nogal como en almendro. Para nogal se extrajeron ramas de 30 centímetros de longitud. Los trozos sometidos al tratamiento fueron de tres centímetros de longitud y se obtuvieron de la parte media de cada rama. En el muestreo de verano las ramas se colocaron en recipientes con agua y los de invierno se mantuvieron en lugar fresco a 10 ºC. Para el caso de las flores, se extrajeron ramas de 15 centímetros de longitud para llevar las flores desde el campo experimental al laboratorio. Luego se sacaron las flores y se sometieron inmediatamente al experimento de frío. Para el análisis estadístico se realizó una comparación de medias (ANAVA) con un subsecuente test de comparaciones múltiples de Tukey con un umbral de significancia del 5% (P=0,05).
RESULTADOS
Grado de resistencia antes y durante el invierno en segmentos de tallos de nogal de un año de edad
En madera joven de nogal variedades Chandler y Franquette se observó amarronamiento, deshidratación del tejido floemático y ruptura de la médula con temperaturas mínimas superiores a los -20 ºC (figura 1). Con una temperatura mínima de -15 ºC, las dos variedades en estado aclimatado presentaron, en un 37,5% de las muestras, amarronamiento del tejido. El daño por heladas afectó de un 10 a un 20% de la superficie radial observada.

Figura 1. A-daño por helada en madera de nogal Chandler (1 año de edad), B-sección de madera sana.
La desaclimatización en invierno incrementó el daño, afectando del 10 al 50% de la superficie radial con temperaturas mínimas de -8 ºC. Con temperaturas de -15 ºC, en Chandler la zona radial analizada fue afectada del 25 al 100% y en Franquette entre 60 y 80%. En estado vegetativo (marzo) Chandler, con temperaturas mínimas de -7,7 ºC presentó el 50% del tejido necrosado y llegó al 100% de daño con -10 ºC por más de una hora. Para las dos variedades los valores de TNH obtenidos fueron iguales para marzo y mayo, pero disminuyeron en aproximadamente 1,5 ºC en julio (tabla 1). Estos resultados indican mayor sensibilidad del tejido en la etapa de inicio de reposo invernal. Como se observa en la tabla 1, no hubo diferencias significativas en el contenido relativo de agua entre marzo y julio. Por lo cual el incremento de mayo se debe a las intensas lluvias ocurridas en el mes de abril (115 a 150 mm). Este mayor contenido hídrico no mostró efecto significativo sobre el valor de TNH. El contenido de azúcares solubles fue variable en los tres momentos medidos, pero entre marzo y julio se observó un incremento del 40% que se corresponde con valores de TNH más bajos (mayor tolerancia). El contenido de almidón no presentó diferencias significativas entre los meses analizados, sin embargo existe una aparente tendencia a la disminución de marzo a julio (tejido desaclimatado y aclimatado respectivamente). Los valores de LT50 (temperatura letal media) variaron en función del grado de desaclimatación en el que se encuentre el tejido (tabla 1 y 2).
Tabla 1. Respuesta de la madera de nogal joven (Juglans regia L.) a las heladas simuladas en estado vegetativo (marzo), inicio de reposo invernal (mayo) y pleno reposo invernal (julio).

*TNH: temperatura de nucleación de hielo. LT50: temperatura a la cual muere el 50% de las muestras. Valores medios. Medias con una letra común no son significativamente diferentes (se lee horizontal), Test de Tukey (α > 0,05).
Tabla 2. Valores de TNH (ºC), azúcares solubles (mg/g), almidón (mg/g), LT50 (ºC), para tejido aclimatado y desaclimatado (mes de julio) para madera de 1 año de edad de nogales (Juglans regia L) de las zonas de Alto Valle (AV) y Valle Medio (VM).

* *valores medios y error estándar. Medias con una letra común no son significativamente diferentes (se lee horizontal), Test de Tukey (α > 0,05).
La desaclimatación con tan solo 4 días de temperaturas de 18º C produjo un efecto significativo en la madera joven de nogal tanto en la TNH, como en LT50 y el contenido de azúcares solubles (tabla 2). En el material desaclimatado la TNH fue más alta por lo cual su tolerancia al daño fue menor que en el tejido aclimatado. Los valores de TNH en tejido con desaclimatización forzada en julio, no difieren significativamente de los obtenidos en las muestras con desaclimatización natural y plena de marzo; los valores medios fueron de -4,55+-0,43 y -5,22+-0,38 respectivamente (test de Tukey α= 0,05). Del análisis del mes de julio se observó que el porcentaje de daño fue de hasta 30% para la madera en estado aclimatado y de 25 a 100% en el tejido desaclimatado (figura 2). Para las heladas de mayor intensidad el tiempo entre el exotermo y la temperatura mínima fue de 1 a 2 horas mayor que en las de menor intensidad. A modo de ejemplo se incorpora una gráfica de las curvas de descenso térmico y exotermo en madera de nogal (figura 3). La velocidad de descenso térmico fue de 2,5 ºC/h (-8 ºC) y 3,5 ºC/h (-15 ºC).

Figura 2. Porcentajes de daño para madera de nogal de 1 año de edad, mes de julio en estado desaclimatado y aclimatado. Rango térmico evaluado -8 ºC a -15 ºC.

Figura 3. Curvas de enfriamiento en madera de nogal de 1 año de edad, variedad Chandler. Temperatura mínima -15 ºC. T4, 3, 18, 11 y 25 son termocuplas dentro de trozos madera.
El contenido de azúcares solubles disminuyó al igual que la resistencia al congelamiento (TNH mayor) cuando el tejido está desaclimatado (tabla 2). En cuanto al contenido hídrico del tejido no se observaron diferencias significativas entre el estado aclimatado y desaclimatado. En estado aclimatado (mayo y julio) las variedades Chandler y Franquette difieren significativamente en TNH. Franquette presentó un menor contenido de agua y valores de TNH más bajos (tabla 3). Esto debería otorgarle mayor capacidad de superenfriamiento y en consecuencia mayor grado de tolerancia a las bajas temperaturas, pero en nuestros ensayos no hemos encontrado diferencias significativas en el grado de daño. De acuerdo a la zona de la cual provenía el material de estudio, no se observaron diferencias significativas en cuanto al grado de tolerancia a las heladas, pero sí en el contenido de azúcares y almidón, VM presentó mayores concentraciones de almidón y menor contenido de azúcares solubles que AV.
Tabla 3. Comportamiento de las variedades Chandler y Franquette ante heladas simuladas. TNH (ºC) y contenido relativo de agua (%) en el tejido aclimatado y desaclimatado, mz: marzo; m: mayo y j: julio.

*valores medios. Medias con una letra común no son significativamente diferentes (se lee horizontal), Test de Tukey (α > 0,05).
Umbrales de superenfriamiento en flores de almendro
En almendro (Prunus amygdalus L.) variedad Guara se observó un comportamiento diferencial entre temporadas. Para las heladas de mayor intensidad, -4 ºC, en el 2011 se registró un 25% de daño por congelamiento del ovario (figura 6 A) y se alcanzó un TNH de -3,8 ºC. En la segunda temporada de estudio, las flores manifestaron exotermos con valores de TNH de -2 ºC a -2,5 ºC, pero con temperaturas mínimas de -2,7 ºC; los daños por congelamiento alcanzaron el 45% de la población muestra. El valor de TNH fue de 1 a 2 ºC más alto un año respecto al otro y el daño se incrementó un 20%. Las temperaturas ambientales previas a las heladas simuladas influyen fuertemente en este comportamiento. Para la comparación se utilizó un índice de desaclimatación para frutales de hoja caduca sujeto a la frecuencia de temperaturas horarias mayores a 16 ºC, 4 días previos a la helada (Rodríguez et al., 2014). El grado de desaclimatación al momento de plena floración fue diferente para cada temporada, siendo mayor en el 2013, lo que podría explicar los valores de TNH más altos y el mayor porcentaje de daño en flores de almendro. Ferraduel y Ferragnes se evaluaron en septiembre del 2013, el día 13 y 20 respectivamente, con heladas de -2,7 y -2,5 ºC (figura 7). Los valores de TNH fueron de -1,7 ºC con un 45% de daño.
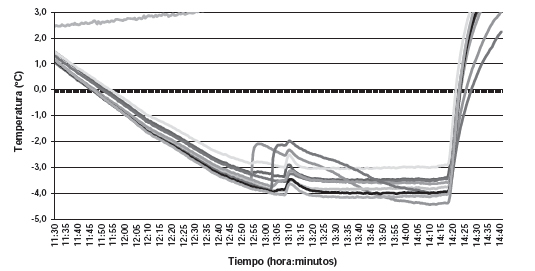
Figura 4. Curva de enfriamiento en flores de almendro (Prunus amygdalus L.) cv Guara. Estado de plena floración (12/9/2011).

Figura 5. Curva de enfriamiento para almendro (Prunus amygdalus L.) cv Guara. Estado de plena floración (13/9/2013), mayor desaclimatación.

Figura 6. Flor de almendro cv Guara. A. daño total por congelamiento del ovario y estigma; B. ovario sano, sin daño.

Figura 7. Curva de enfriamiento para almendro (Prunus amygdalus L.) cv Ferraduel. Estado de plena floración (13/9/2013).
Analizando las temperaturas mínimas de 1990 a 2015 se observa que durante el 26 de agosto hasta el 18 de septiembre (período medio de floración de almendros), la probabilidad de ocurrencia de heladas es del 28% y se ha mantenido constante con 7 eventos por año. Si bien, en los últimos 25 años, ha disminuido la frecuencia de heladas fuertes y muy fuertes, la frecuencia de temperaturas de entre -2º y -3,9 ºC no ha variado. La peligrosidad de daño es alta para cualquiera de los subperíodos analizados (tabla 4).
Tabla 4. Peligrosidad de heladas primaverales para Alto Valle durante el período de floración de almendros*, serie 1990-2015.

*fechas medias de inicio y fin de floración de Guara (serie 1978- 2014) según Iannamico (2015b).
DISCUSIÓN Y CONCLUSIONES
Grado de resistencia antes y durante el invierno en segmentos de tallos de nogal de un año de edad
Definir la temperatura crítica de daño en madera es complejo, la temperatura mínima alcanzada no es un indicador directo de la muerte de madera joven, varios factores influyen en este proceso. Entre ellos está la capacidad de reparar el tejido ante daños parciales, el tiempo transcurrido entre la TNH y el fin de la helada, la diferencia de sensibilidad entre las distintas estructuras de un mismo árbol y el grado de desaclimatización del tejido dependiendo de las temperaturas de los días previos a la helada. Diferentes trabajos sobre congelamiento de tejido leñoso señalan que la concentración de carbohidratos totales se relaciona con la resistencia a las heladas porque regula el ajuste osmótico que previene el congelamiento intracelular (Morin et al., 2007; Poirier et al., 2010; Charrier et al., 2013). En términos generales, las ramas de un año de edad en nogales tienen de 79-85 mg g-1 de azucares solubles en invierno y esto se corresponde con LT50 de -21 ºC a -24 °C. En nuestros estudios el contenido de azucares solubles no superó los 54 mg g-1, y con -15 ºC se alcanzó 25% de daño en pleno reposo invernal. Hemos observado que si el tejido permanece menos de 1 hora al valor de TNH, no sufre daño por helada. En mayo si bien la TNH no difiere estadísticamente con respecto a marzo, el tejido soportó temperaturas más bajas antes de alcanzar un 50% de daño. Durante el otoño hubo menor tolerancia al congelamiento y más variabilidad en la respuesta que en el invierno. Durante el invierno los árboles de hojas caducas utilizan una alta proporción de hidratos de carbono para resistir a las bajas temperaturas, mientras que en otoño y primavera esta relación disminuye porque se destinan también a procesos asociados al crecimiento. Nuestros resultados son similares a los presentados por Aslani Aslamarz et al. (2010), en donde las temperaturas exotérmicas en tallos de 1 año de edad para las variedades Pedro y Lara de nogal son de -4 ºC en mayo, -5 ºC en julio y en primavera de -2,5 ºC. Con respecto a los LT50 hemos estimado en Franquette y Chandler valores de -7,7 ºC para marzo. Estos resultados coinciden con los calculados por Charrier et al. (2011) cercanos a los -10 ºC en junio, hemisferio norte (diciembre, equivalente hemisferio sur) para las mismas variedades.
A mitad del invierno el proceso de aclimatación es completo. Hemos observado en este estudio que con tres días de temperaturas a 18 ºC, la madera 100% aclimatada perdió tolerancia alcanzando el mismo valor de TNH que el tejido en estado vegetativo. En algunas especies semileñosas se ha comprobado que con 4 días a temperaturas de 17º/22 ºC, la desaclimatización provoca una pérdida de resistencia de hasta 5 ºC (Pagter et al., 2011). En nogal la movilización de almidón en invierno genera una acumulación de sucrosa relacionada con la tolerancia a las heladas porque interviene en la reparación de vasos conductores afectados por embolismo provocados por congelamiento (Améglio et al., 2002). Esta podría ser la razón principal por la cual los daños en madera pueden ser parciales o nulos en tejidos en proceso de aclimatación (otoño). Probablemente la madera nueva al estar sometida a una mayor variabilidad metabólica, por el ritmo de crecimiento, responda rápidamente a los cambios en el contenido de azucares solubles afectando la TNH y grado de tolerancia. Las prácticas de manejo (fertilización, riego, poda, etc.) y climáticas ligadas al proceso fotosintético condicionan el nivel de reservas nutricionales y por ende los procesos asociados al balance total de carbohidratos.
Umbrales de superenfriamiento en flores de almendro
Los tejidos florales pierden capacidad de superenfriado al avanzar el desarrollo fenológico y mueren luego de que se inicia la formación de hielo (Ballard et al., 1971; Proebsting yMills, 1978). Los efectos de las heladas primaverales sobre los órganos reproductivos del almendro son muy variables, se han observado respuestas diferentes entre genotipos, tejidos dentro de una misma planta y entre ciclos productivos (Imani y Khani, 2011). En nuestros estudios observamos que si las flores permanecen por más de 30 minutos a temperaturas iguales a la TNH se produce muerte celular y consecuentemente daño total del ovario. El valor de LT50 depende de cada temporada evaluada al igual que el TNH, sin embargo podemos decir que ante condiciones térmicas propicias para la desaclimatación de los tejidos, las flores de almendro mueren por congelamiento al exponerse por más de treinta minutos a temperaturas subcero de 2,5 ºC o menores. En nuestra investigación los datos obtenidos pueden considerarse como los primeros valores referenciales de tolerancia a las bajas temperaturas para el cultivar Guara, y están dentro de los rangos térmicos obtenidos por otros autores en otros cultivares; por ejemplo Imani y Khani, 2011 y Kodad et al., 2010 obtienen en floración 100% de daño con temperaturas mínimas de -4 ºC en la variedad Ferragnes y para Felisia más del 50% de daño con temperaturas de -2,5ºC, ambas expuestas por el lapso de 1 hora.
AGRADECIMIENTOS
A Celeste Kloster y Fernanda Toscana por acompañarnos durante su pasantía en este trabajo.
BIBLIOGRAFÍA
1. AMÉGLIO, T.; BODET, C.; LACOINTE, A.; COCHARD, H. 2002. Winter embolism, mechanisms of xylem hydraulic conductivity recovery and springtime growth patterns in walnut and peach trees. Tree Physiology, 22:1211–1220.
2. ASLANI ASLAMARZ, A.; VAHDATI, K.; RAHEMI, M. 2010. Supercooling and Cold hardiness of acclimated and deacclimated buds and stems of Persian walnut cultivars and selections. Hort- Science 45(11): 1662–1667.
3. BALLARD, J.; PROEBSTING, E.; TUKEY, R.; MIUS, H. 1971. Critical temperatures for blossom buds. Was. Agr. Ext. Circ. Nos., 369–374.
4. CHARRIER, G.; BONHOMME, M.; LACOINTE, A.; AMEGLIO, T. 2011. Are budburst dates, dormancy and cold acclimation in walnut trees (Junglans regia L.) under mainly genotypic or environmental control? Int. J. Biometeorol, 55: 763–774.
5. CHARRIER, G.; POIRIER, M.; BONHOMME, M.; LACOINTE, A.; AMEGLIO, T. 2013. Frost hardiness in walnut trees (Junglans regia L.): How to link physiology and modeling? Tree Physiology, 33: 1229–1241.
6. IANNAMICO, L. 2009. El cultivo de nogal en clima templado fríos. Primera edición, Instituto Nacional de Tecnología Agropecuaría (INTA). Buenos Aires. 116 p. [ Links ]
7. IANNAMICO, L. 2015a. Cultivo de Nogal. Ediciones INTA. [ Links ]
8. IANNAMICO, L. 2015b. Cultivo de Almendro. Ediciones INTA. [ Links ]
9. IMANI, A.; KHANI, Y.M. 2011. Characteristics of almond selections in relation to late frost spring. International Journal of Nuts and Related Sciences 2(2): 77–80.
10. KODAD, O.; MORALES, F.; SOCIAS, R. 2010. Evaluación de la tolerancia de las flores de almendro a las heladas por fluorescencia de clorofila. ITEA, 106 (2): 142–150.
11. LEVITT, J. 1978. Plant Cold Hardiness and Freezing Stress: An overview of freezing injury and survival, and its interrelationships to other stresses. Ed. Li P.H and Sakaia. Nueva York, San Francisco, 1: 3–15.
12. MATHERS, H.M. 2004. Supercooling and cold hardiness in sour cherry germplasm: Flower buds. J. Amer. Soc. Hort. Sci. 129(5):675–681.
13. MENG, Q.; LLIANG, Y.; WANG, W.; DU, S.; LI, Y.; YANG, J. 2007. Study on supercooling point and freezing point in floral organs of apricot. Agricultural Sciences in China, 6 (11): 1330–1335.
14. MILLS, L.J.; FERGUSON, J.C.; KELLER, M. 2006. Cold-Hardiness Evaluation of Grapevine buds and Cane Tissues.Am.J.Enol. Vitic. 57(2): 194–200.
15. MORIN, X.; AMÉGLIO, T.; AHAS, R.; KURZ-BESSON, C.; LANTA, V.; LEBOURGEOIS, F.; MIGLIETTA, F.; CHUINE, I. 2007. Variation in cold hardiness and carbohydrate concentration from dormancy induction to bud burst among provenances of three European oak species. Tree Physiology, 27: 817–825.
16. PAGTER, M.; HAUSMAN, J.F.; ARORA, R. 2011. Deacclimation kinetics and carbohydrate changes in stem tissues of Hydrangea in response to an experimental warm spell. Plant Science, 180: 140–148.
17. POIRIER, M.; LACOINTE, A.; AMÉGLIO, T. 2010. A semi-physiological model of cold hardening and dehardening in walnut stem. Tree Physiology 30, 1555–1569.
18. PROEBSTING, E.L.; MILLS, H.H. 1978. Low temperature resistance of developing flower buds of six deciduous fruit species. J.Amer.Soc.Hort.Sci. 103(2): 192–198.
19. RODRÍGUEZ, A.; MUÑOZ, A. 2006. Síntesis Agro-meteorológica para el período 1990-2004 EEA Alto Valle. Ed. INTA. Boletín Divulgación Técnica n.º 53, 38 p. [ Links ]
20. RODRIGUEZ, A.; BENITEZ PICCINI, E.; MUÑOZ, A. 2012. Desarrollo de un prototipo para determinar temperaturas críticas de daño por heladas en órganos de cultivos frutícolas. xiv Reunión Argentina de Agrometeorología, Malargüe, Mendoza, Argentina. [ Links ]
21. RODRÍGUEZ, A.; KLOSTER, C.; BENÍTEZ PICCINI, E.; MUÑOZ, A. 2014. Estimación de un índice de desaclimatación para prever sensibilidad al daño por heladas en frutales del Alto Valle de Río Negro y Neuquén. Reunión Internacional De Agrometeorologia Uruguay. [ Links ]
22. SOUTHGATE, D.A.T. 1991. Determination of food carbohydrate. Second Edition, Elsevier Applied Science, Nueva York. [ Links ]

