











 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
PermalinkINTRODUCCIÓN
El cambio de la cobertura vegetal del suelo genera alteraciones en los flujos de agua y solutos, cuya magnitud varía según diversos factores (Nosetto et al., 2008; Jobbágy et al., 2008). Así se han verificado procesos de salinización secundaria como fruto de alguna actividad antrópica que rompe el equilibrio hídrico produciendo la removilización de las sales desde fuentes cercanas (subsuelos salinos, aguas subterráneas, etc.) (Zhang et al., 2003; Bin et al., 2014). Este proceso de salinización secundaria puede surgir como consecuencia de la forestación de pastizales y posee tres componentes impulsores: aspectos de índole biológico, hidrogeológicos y climáticos (Nosetto et al., 2008; Jobbágy et al., 2008).
El componente biológico incide en el proceso de salinización secundaria a través del consumo diferencial de agua dado por las características ecofisiológicas de las especies (Gyenge et al., 2011), su tolerancia a la salinidad (Nosetto et al., 2008) y la redistribución de las precipitaciones a través de la cobertura vegetal (Besteiro y Rodríguez Vagaría, 2012). En general, el consumo diferencial de agua de las forestaciones se encuentra ligado con su productividad, aunque no siempre de manera proporcional (Gyenge et al., 2011). Así los árboles de rápido crecimiento consumen más recursos que los de menor productividad (por ejemplo; Gyenge et al., 2011). En este sentido, ante el caso de que dos especies se cultiven en el mismo sitio, se esperaría la ocurrencia de este proceso de salinización secundaria en la forestación con la especie más productiva. Asimismo, la magnitud de la salinización también dependerá del nivel de tolerancia de las especies a la concentración de solutos en el suelo, tal como lo demostraron Nosetto et al. (2008). Estos autores observaron que las especies más tolerantes pueden llevar a una mayor acumulación de sales antes de sufrir los efectos negativos. Además del uso del agua, es importante analizar la interceptación de lluvias por parte del dosel forestal que influye directamente sobre los flujos verticales de agua. Este proceso también difiere entre especies. Por ejemplo, Besteiro et al. (2012) estimaron que la intercepción de las precipitaciones de forestaciones de Eucaliptus spp. fueron inferiores a forestaciones de Pinus spp. Por un lado, estas diferencias generarían cambios en la cantidad de precipitación directa que llegue al suelo, pudiendo provocar mayores flujos de drenaje y, por lo tanto, un mayor lavado de sales del perfil del suelo (Nosetto et al., 2008; Jobbágy et al., 2008). Por otro lado, tal como menciona Besteiro et al. (2013), el cambio de cobertura vegetal modifica los procesos de lavado / lixiviado de sales del suelo. En este sentido, como la cobertura de una forestación puede ser modificada por el manejo (densidad de plantación, podas, raleos), es posible disminuir la evapotranspiración del ecosistema lo que podría generar mayores excesos hídricos y, por lo tanto, un mayor lixiviado de sales del suelo (Zhang, 2001).
El objetivo principal de este trabajo fue el de cuantificar la conductividad eléctrica del suelo (CE, µS cm-1) sometido a distintos usos y manejos silvícolas: forestaciones con Eucalyptus spp. y Pinus radiata, agricultura y pastizal natural. Se utilizó la CE del suelo como un indicador de los cambios en la concentración de sales, permitiéndonos identificar situaciones que comprometan la calidad / productividad de los cultivos agrícolas y forestales más conspicuos de la región.
MATERIALES Y MÉTODOS
Las sierras de Tandilia, con una altura máxima de 524 m s. n. m., se alzan sobre la llanura bonaerense con orientación NO-SE y tiene una longitud de 350 km desde la ciudad de Mar del Plata hasta la ciudad de Olavarría y un ancho máximo de 60 km (De la Sota, 1967; Dalla Salda et al., 2006). El área de estudio se encuentra en la zona central de este sistema, en el grupo serrano de Tandil-Barker, provincia de Buenos Aires (figura 1). La vegetación natural predominante son los pajonales y matorrales (Frangi, 1975). El clima es templado húmedo con precipitaciones promedio que rondan los 893 mm año-1 y distribuidas de manera más o menos homogénea a lo largo del año (Aeródromo Tandil, 2016), lo que genera un balance hídrico de 0 a 200 mm año-1 (Nosetto et al., 2008, figura 1). Los suelos son de textura franca a franca fina, profundos en el piedemonte y en las sierras son poco desarrollados y están emplazados sobre un manto rocoso (INTA, 1989).
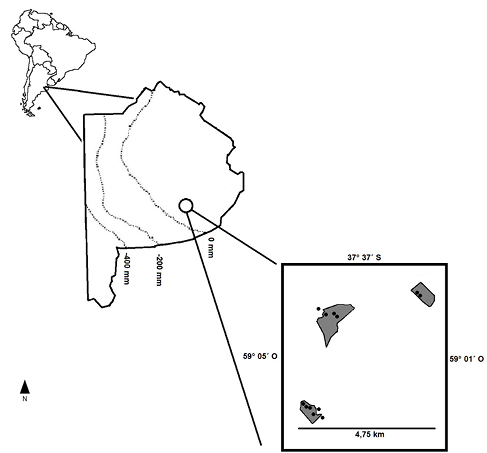
Figura 1Ubicación de la zona de muestreo (círculo hueco) con sus parcelas apareadas (puntos negros), en gris se denota las forestaciones. Con líneas punteadas se denotan los balances hídricos calculados como la diferencia entre la precipitación media anual y la evapotranspiración potencial de Penman-Montheith (modificado a partir de Nosetto et al., 2008).
Durante el mes de febrero del 2016, se establecieron parcelas apareadas en sitios con una cobertura vegetal herbácea (pastura o pastizal) y otras en forestaciones de Pinus radiata y Eucalyptus spp. Todas las forestaciones fueron implantadas en 1990 (28 años de edad al momento del muestreo) y poseían diferentes manejos silvícolas. Los manejos silvícolas abarcaron macizos de Eucalyptus spp. (EU) y Pinus radiata (PD) (códigos EU1, EU2, EU3, PD1, PD2 y PD3, tabla 1), un sistema silvopastoril en fajas de 10 metros de ancho abierta en el año 2011 (código F11, tabla 1), otro silvopastoril con fajas de 14 metros de ancho abierta en el año 2013 (código F21, tabla 1) y una tala rasa realizada dos años antes del muestreo (código Tr2, tabla 1) todos provenientes de macizos de Pinus radiata.
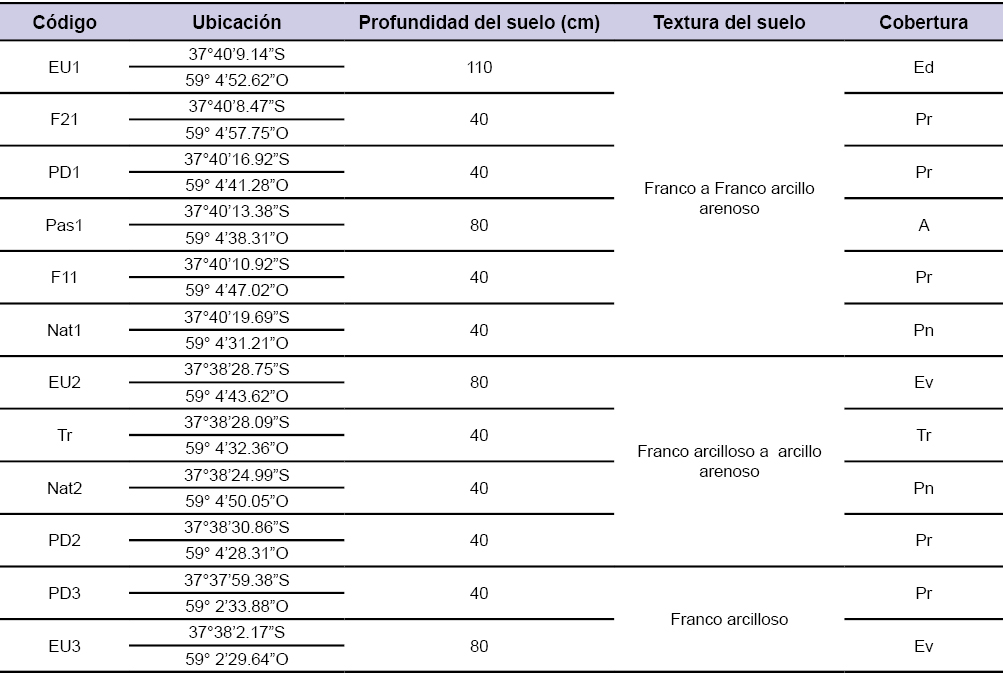
Tabla 1Código y ubicación geodésica de las parcelas, profundidad de suelo (estimada a partir de los mapas de suelo 1:50000, INTA (1989) y corroborado a campo), textura del suelo, cobertura (Ed: Eucalyptus dalrympeana, Ev: Eucalyptus viminalis, Pr: Pinus radiata, Pn: Pastizal natural, A: Avena sativa, Tr: Tala rasa).
Con el objetivo de analizar las características estructurales del rodal y tomar las muestras de suelo, en las parcelas situadas en las forestaciones se demarcó una superficie que tuvo distinto tamaño dependiendo de la densidad y la distribución de los árboles. Así, en unos sitios con distribución irregular de los árboles (sitios EU1, PD2 y EU2) se delimitaron superficies circulares de 10 y 30 metros de diámetro dependiendo del distanciamiento entre los árboles, mientras que en los sitios con una distribución regular (sitios EU3, PD1, PD3) se establecieron parcelas de 4 filas por 10 árboles, midiendo a posteriori la superficie ocupada (figura 2).
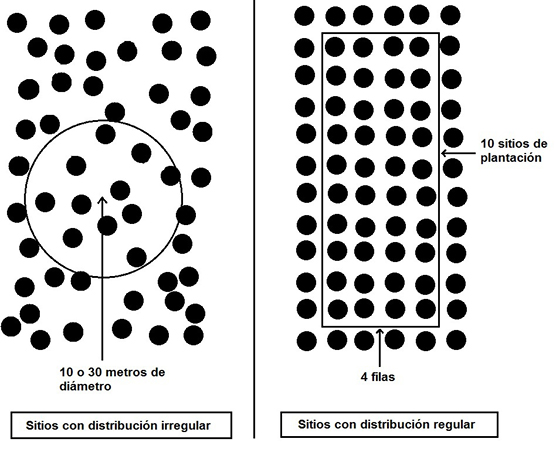
Figura 2Ejemplo de cómo fueron establecidas dentro de la forestación las parcelas muestreadas (los puntos negros representan los árboles). Estas parcelas tuvieron distinto tamaño dependiendo de la densidad y la distribución espacial de los árboles.
Tanto en las parcelas con forestaciones como en las establecidas en los cultivos herbáceos, pastizal natural y tala rasa, se extrajeron muestras de suelo en 5 puntos al azar hasta máxima profundidad posible (las limitaciones de profundidad están dadas en todos los casos por la presencia de un manto rocoso o carbonato de calcio cementado). La distancia entre ambas parcelas apareadas fue de, aproximadamente, unos 50 metros buscando la mayor homogeneidad posible en cuanto a pendiente y exposición. Se analizó la conductividad eléctrica del suelo (CE, µS cm-1) en diluciones 1:2,5 por medio de un lector OAKTON PC700 con sonda de CE-temperatura 35608-74 y se determinó la textura del suelo de cada muestra por el método de Bouyoucos (p.ej., Bouyoucos, 1962; Elliot et al., 1999).
Con el fin de caracterizar las forestaciones y dentro de cada superficie delimitada, se midió el diámetro a la altura del pecho (DAP, cm), con cinta diamétrica, de todos los árboles vivos y la altura de los árboles dominantes (H, m) mediante clinómetro Suunto PM-5/360PC (Suunto, Finlandia). Se consideró como pie individual a todos aquellos fustes mayores de 3 cm de diámetro. A partir de estos estadísticos se estimó la densidad (árboles ha-1) y área basal del rodal (AB, m2 ha-1; tabla 2) estimada a partir de la sumatoria de todas las áreas individuales obtenidas a partir del DAP. La cobertura forestal se estimó a partir del análisis de fotografías planas mediante el programa ImageJ 1.48v (Wayne Rasband, National Institutes of Health, EE. UU.).
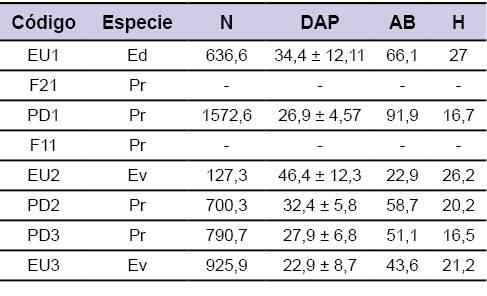
Tabla 2Variables descriptoras de los rodales. Especie= Ed: Eucalyptus dalrympeana, Ev: Eucalyptus viminalis, Pr: Pinus radiata; N= Densidad (árboles ha-1); DAP= diámetro del fuste a 1,30 m (cm); AB= área basal (m2 ha-1); H= altura dominante (m).
Se analizaron diferencias entre tratamientos (eucaliptos macizo, pinos macizo, pinos silvopastoril, pastizal natural, avena y tala rasa; tabla 3) mediante ANOVA con un α = 0,05 aplicándose test de LSD de Fisher (InfoStat versión 2011). En el caso de las forestaciones con pinos, la mayor profundidad encontrada fue de 40 cm. Por este motivo, se compararon los valores promedio de CE por cada tratamiento de las profundidades 0-20 y 20-40 cm.
RESULTADOS
Caracterización de las forestaciones
Las forestaciones que denominamos macizos (códigos EU1, EU2, EU3, PD1, PD2 y PD3, tabla 1) carecían de manejo silvícola. La densidad de plantación varió desde 127,3 hasta 1572,6 árboles ha-1 (para macizos de Eucalyptus viminalis y Pinus radiata respectivamente), mientras que el área basal de los rodales estuvo entre los 22,9 y 91,9 m2 ha-1 (tabla 2). La cobertura del dosel arbóreo varió entre 11,2% y 76,3%, estimándose para las fajas del sistema silvopastoril y macizo de Pinus radiata respectivamente (figura 3), mientras que los macizos de Eucalyptus spp. mostraron valores intermedios (de 50 al 60%) (figura 3).
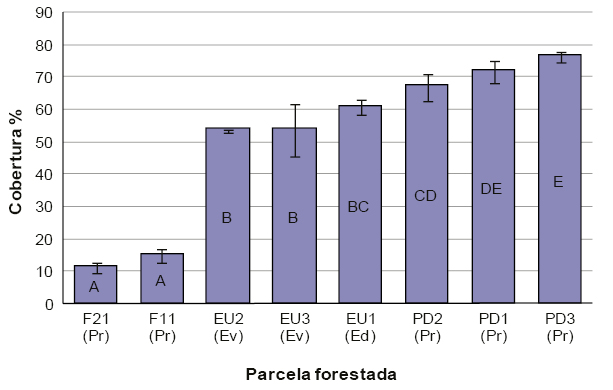
Figura 3Cobertura del dosel arbóreo (%) y sus desvíos para cada parcela de la localidad de Tandil. Entre paréntesis la especie correspondiente a cada parcela (Pr: Pinus radiata, Ev: Eucalyptus viminalis, Ed: Eucalyptus dalrympeana). Letras distintas indican diferencias significativas entre parcelas con un α = 0,05.
Variables de suelo por especie y manejo
No se observó una gran variación en la textura de los suelos de los distintos tratamientos, estimándose una textura general franca en la superficie (0-20 cm) a franca arcillosa o arcillosa arenosa en profundidad (20-40 cm, tabla 1).
Por un lado, en general, se observó una tendencia a obtener menores valores de CE en los perfiles de suelos más profundos (tabla 3). No se encontraron diferencias significativas entre tratamientos en la CE del suelo a una profundidad de 0-20 cm, a pesar de los manejos y especies contrastantes (tabla 3). Por el contrario, la CE varió en la profundidad de 20-40 cm en respuesta al uso y al manejo silvícola. Así en el aprovechamiento a tala rasa, con tan solo 2 años de antigüedad, se midieron los valores más bajos de CE (65,08 µS cm-1, tabla 3), mostrando diferencias significativas con todos los tratamientos (herbáceas, silvopastoriles y macizos de Pinus radiata) excepto con el suelo donde se situaron los macizos de Eucalyptus spp. En contraste, a esta profundidad del suelo, los macizos de Pinus radiata mostraron los valores más altos de CE (110,44 µS cm-1, tabla 3) diferenciándose de la tala rasa y los macizos de Eucalyptus spp. Por último, la CE del suelo en los sistemas silvopastoriles mostraron valores intermedios (101,31 µS cm-1, tabla 3), que no se diferenciaron del pastizal ni del cultivo.
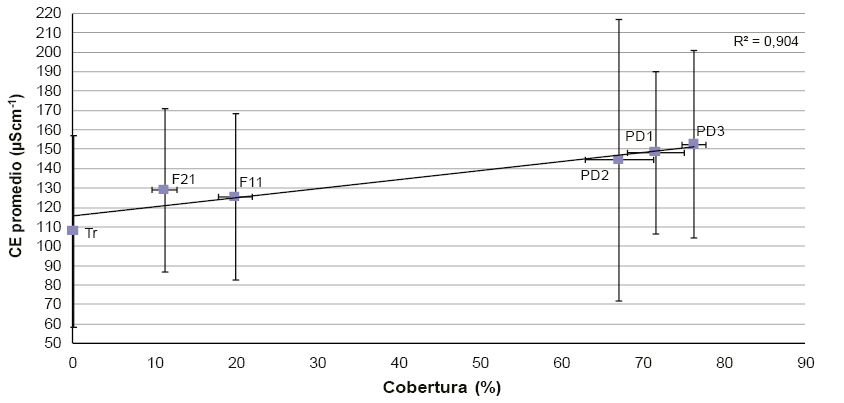
Por otro lado, al analizar los distintos tipos de manejo forestal para la especie Pinus radiata, se encontró una relación positiva entre la CE promedio del suelo y la cobertura (R2 = 0,94, figura 4).
DISCUSIÓN
Características de las forestaciones
La variación de la cobertura arbórea se relacionó de manera lógica con los manejos forestales (tabla 2). Así, las mayores coberturas se estimaron en las parcelas de los macizos de Pinus radiata, los que, además, mostraron una mayor densidad y área basal. De manera contraria, las fajas silvopastoriles mostraron valores bajos de cobertura (figura 3), propios de estos sistemas de manejo (Fassola et al., 2005).
Efectos en la CE del suelo producto del cambio de uso
No se observaron diferencias en la CE del suelo entre los tratamientos en los primeros 20 cm de profundidad, más allá del tipo de cobertura vegetal y manejo. Esto posiblemente se deba a un fácil lavado de los cationes debido a la buena infiltración de las texturas francas en la superficie (Weil y Brady, 2017). No obstante, se encontraron diferencias significativas entre los tratamientos en la profundidad de 20-40 cm, observándose una mayor CE en los macizos de Pinus radiata (tabla 3). Este efecto puede ser producto de la mayor cobertura arbórea (figura 3) en conjunto con una acumulación de solutos lixiviados sobre el manto rocoso. Sin embargo, estos valores están muy por debajo de los 1000 µS cm-1 (1 dS m-1), que podrían afectar la producción de los cultivos agrícolas o forestales más sensibles de la región (Hillel, 2000; Marcar et al., 2002; Stirzaker et al., 2002). De manera contraria, se observó la menor CE en el suelo de los macizos de Eucalyptus spp. a 20-40 cm, los que poseían una menor cobertura (figura 3) y se encontraban sobre suelos más profundos (tabla 1), permitiendo probablemente un mayor lixiviado de sales en profundidad. Así también, existen antecedentes que plantean la baja capacidad de retención de agua en el suelo forestal en rodales de Eucalyptus spp. frente a pastizales naturales. Esto debido a un mayor porcentaje de macro/mesoporos y a una mayor hidrofobicidad de la materia orgánica (Silveira et al., 2006). Esta combinación de factores favorecería el lavado de sales hacia perfiles más profundos durante los periodos de exceso hídrico.
Como mencionamos, a una profundidad de 20-40 cm, el aprovechamiento a tala rasa obtuvo los valores más bajos de CE (tabla 3). Como es de esperar, el efecto más notorio ocurre al disminuir bruscamente la cobertura forestal. Tal como menciona Roberts (2001), la tala rasa genera un aumento instantáneo del rendimiento hídrico debido a la consecuente caída del consumo de agua. Así, es interesante recalcar lo rápido que se observa una reversibilidad del proceso, ya que se midió una disminución significativa de la CE con solo dos años desde que la forestación fue cosechada. Como complemento, sería interesante analizar los cambios en la calidad de las salidas de agua en las forestaciones sometidas a este tipo de manejo.
La relación positiva entre la CE del suelo y la cobertura forestal (figura 4) pone de manifiesto el valor del manejo silvícola como herramienta para disminuir la acumulación de sales en la rizosfera. En este sentido, las fajas silvopastoriles a una profundidad de 20-40 cm obtuvieron valores intermedios de CE, solo diferenciándose de la tala rasa (mínimo valor absoluto de CE para todas las situaciones consideradas) y muy similares a los del pastizal natural (tabla 3). Así, es importante resaltar la importancia del manejo forestal en cuanto a la acumulación de sales en el suelo, como también a la disponibilidad relativa de los recursos que limitan la productividad (por ejemplo, el agua; Martínez Meier et al., 2015). Entonces, como mencionan Jobbágy et al. (2006), la instalación de plantaciones con bajas densidades podría ser considerada como una alternativa de menor impacto en estas regiones que pueda reducir la intensidad de los procesos de salinización secundaría.
Efecto de las especies en la CE
Tal como se esperaba, los mayores valores de CE fueron estimados en los pinares (tabla 3). En este sentido, Eucalyptus viminalis y Eucalyptus dalrympleana fueron descriptas como especies de tolerancia baja a moderada (2000 a 4000 µS cm-1; FAO 1981, Marcar et al., 2002), mientras que P. radiata se describe como de una mayor tolerancia a la salinidad que estas especies (4000 a 8000 µS cm-1; Marcar et al., 2002). Esto estaría en concordancia con lo encontrado por Nosetto et al. (2008), quienes indican que las especies más tolerantes podrían llevar a una acumulación mayor de sales. Finalmente, si bien los crecimientos en el centro de la provincia de Buenos Aires de Eucalyptus viminalis son mayores a los de Pinus radiata (SINAVIMO, 2014; Ferrere et al., 2015) la CE del suelo mostró una relación inversa, indicando que la mayor productividad no fue un factor determinante como impulsor de un proceso de salinización secundaria. En este sentido, sería importante determinar el consumo y la eficiencia en el uso del agua de ambas especies, no solo teniendo en cuenta el efecto sobre la dinámica de sales en el suelo, sino también sobre el posible impacto sobre los recursos hídricos locales.
CONCLUSIONES
El cambio de uso del suelo, las distintas especies forestales y el manejo silvícola generan cambios en la magnitud de las sales acumuladas en el suelo. Sin embargo, estas diferencias son reversibles y no de una magnitud tal como para generar consecuencias negativas en los cultivos agrícolas o forestales. Estos resultados contribuyen a comprender que la especie vegetal dominante y el manejo son factores clave para tener en cuenta, ya que impactan rápidamente en los flujos de sales del sistema. Así, una menor expresión de los procesos de acumulación de sales puede ir de la mano de una correcta selección de la especie y de un manejo adecuado de los turnos de corta, densidades de plantación, raleos y escamondos.
