










Servicios Personalizados
Revista

Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkRevista veterinaria
versión On-line ISSN 1669-6840
Rev. vet. vol.28 no.1 Corrientes ene. 2017
TRABAJOS DE INVESTIGACIÓN
Cambios morfométricos de lagunas aluviales del río Paraná y su incidencia en la diversidad íctica
Contreras, F.I.1; Meza, J.C.1; Scipioni, N.2; Hernández, D.R.3; Ruiz-Díaz, F.J.3
1Centro de Ecología Aplicada del Litoral (CONICET-UNNE) Ruta Provincial N° 5 km 2,5; Corrientes (3400), Argentina. Tel-Fax: +54 (379) 4454417; 2Instituto de Investigaciones Geohistóricas (CONICET-UNNE); 2Instituto de Ictiología del Nordeste, Facultad de Ciencias Veterinarias (UNNE), E-mail: figcontreras@hotmail.com
Resumen
Para comprender cuáles son las características ambientales que inciden en las comunidades acuáticas de una laguna, es necesario poseer conocimientos referidos a sus formas y dimensiones, ya que su morfometría interviene sobre variables relacionadas con la erosión, sedimentación, carga de nutrientes y características físico-químicas, entre otras. Sin embargo, la renovación del agua de las lagunas del valle aluvial se encuentra íntimamente ligada a los pulsos de inundación de este último, entendiéndose que es tan importante la altura del nivel de agua, como su duración en el tiempo. Diferentes niveles topográficos de la llanura aluvial, pueden generar distintas respuestas en las lagunas, al margen de encontrarse próximas entre ellas. En tal sentido, el objetivo de este trabajo fue aplicar modelos de anegamiento en la llanura aluvial del río Paraná, a fin de establecer las diferencias topográficas entre las lagunas Galloso y Melilla, para explicar sus respuestas frente a los pulsos de inundación y por consiguiente sus diferencias morfométricas y biológicas. Para ello se utilizó la herramienta Increase level water del software Global Mapper 15.1, teniendo como base un modelo de elevación digital generado a partir de una imagen SRTM de 30 m. Los resultados demostraron que la laguna Galloso se encuentra 2 m por debajo de la laguna Melilla, quedando más expuesta a los pulsos de inundación del río Paraná. Dicha situación se ve reflejada en sus variaciones morfométricas entre los años estudiados, en donde la laguna Galloso ha aumentado 19 veces su superficie mientras que la laguna Melilla se mantuvo estable. En conclusión, esta diferencia topográfica y la mayor conectividad con el río Paraná, sería el principal motivo por el cual la primera posee 80 especies de peces, mientras que la segunda solo 49 especies.
Palabras clave: peces, lagunas, río Paraná, repercusión en comunidades acuáticas.
Abstract
To understand which are the environmental characteristics that affect aquatic communities of floodplain lakes it is necessary to possess beforehand information regarding shapes and dimensions as morphometry intervenes in the variables related to erosion, sedimentation, nutrient`s charge, physicochemical characteristics, among others. Renovation of the waters of floodplain lakes is closely linked to the flood pulse of the latter, considering the importance of the height of water level and the duration of the flood period. Different topographic levels of the floodplain can generate different responses in the lagoons, regardless of being close to each other. The aim of this assay is to apply flooding models in the study of the floodplain of the Paraná river in order to establish the topographical differences between two lakes and their implications for the diversity of fish species to determine their responses to flood pulses and consequently their morphometric and biological differences. For this, the Increase level water tool of the Global Mapper 15.1 software was used considering a digital elevation model generated from an SRTM image. The results showed that the Galloso lagoon is 2 m below the Melilla lagoon leaving the former more exposed to flood pulses of the Paraná River. This situation is reflected in their morphometric variations among the years under consideration, having Galloso lagoon increased 19 times its surface while Melilla lagoon remained stable. In conclusion, this topographic difference and proximity to the Paraná River may be the main reason why Galloso lagoon has 80 fish species while Melilla lagoon only 49.
Key words: fish, floodplain lakes, Paraná River, repercussion in aquatic communities.
Recibido: 3 octubre 2016
Aceptado: 7 febrero 2017
INTRODUCCIÓN
El río Paraná posee la segunda mayor cuenca de América del Sur y es el principal curso de agua de la Cuenca del Plata, extendiéndose 3.780 km desde la confluencia de los ríos Paranaíba y Grande hasta el estuario del río de La Plata.Comprende la sección denominada Paraná superior, desde la confluencia del río Grande hasta las cataratas de Sete Quedas en el estado de Guairá, Brasil, la sección llamada Paraná medio que se extiende desde Sete Quedas hasta los saltos de Yacyretá-Apipé y por último la sección del Paraná Inferior extendiéndose desde los saltos de Yacyretá-Apipé hasta su desembocadura 2, 3. Sobre el territorio de la Provincia de Corrientes es un río de valle aluvial que se abre en varios brazos que encierran grandes islas como las de Yacyretá, Talavera, Apipé Grande, Apipé Chico y otras menores. Tras la represa de Yacyretá el río presenta una clara dirección hacia el oeste, ensanchándose progresivamente (Figura 1) y ramificándose en varios canales que forman gran cantidad de islas fluviales. Luego de recibir la confluencia del río Paraguay se dirige hacia el sur para desembocar en el río de La Plata.
La comunidad de peces de la cuenca del río Paraná y sus ambientes asociados está integrada por alrededor de 400 especies 12. Sin embargo, el conocimiento sobre la diversidad de peces y sus patrones espacio-temporales aún es escaso. La identificación de los patrones de ocurrencia de especies y su relación con el hábitat permiten plantear hipótesis respecto a la estructura y la dinámica de la comunidad 17.
En los ríos neotropicales, el régimen hidrológico es considerado un factor regulador de los patrones de biodiversidad. El valle aluvial de estos ríos contieneuna amplia variedad de biotopos con gradientes espaciales asociados a la conectividad hídrica 18. El pulso de inundación produce un intercambio lateral de agua, nutrientes y organismos entre el cauce principal del río y los ambientes asociados al valle aluvial, con efectos significativos en la organización de los ensambles de peces 11, 13.
Un ejemplo de ello ocurre en Pantanal (Mato Grosso, Brasil), donde las inundaciones anuales cubren de agua alrededor de un 80% de la superficie, incidiendo en el metabolismo de éste bioma a causa de la hidrología y el enriquecimiento de nutrientes 1, 4. El agua que ingresa de un río hacía las planicies laterales genera como resultado una disminución de oxígeno disuelto y dióxido de carbono, incrementando la cantidad de sólidos suspendidos, principalmente de detritos y algas 9.
En el valle aluvial del río Paraná inferior se localizan cuerpos lénticos, los cuales se ven sometidos a dichos pulsos de inundación, representando un escenario útil para la identificación de los principales factores o procesos que inciden en la diversidad íctica.
En este sentido, son fundamentales los estudios geomorfológicos, ya que condicionan los parámetros fisicoquímicos y biológicos de estos ambientes. Sin embargo, si bien existen antecedentes sobre el estudio de variables morfométricas del valle aluvial del río Paraná, en la actualidad no se han encontrado trabajos que hagan referencia a la relación existente entre dichas variables y su incidencia en la diversidad de peces, como así también en la digitalización, la geo-referenciación de información de campo y la generación de cartografía temática. El objetivo de este trabajo fue determinar las características ambientales que inciden en la diferencia de diversidad de comunidades de peces entre dos lagunas de la zona, empleando herramientas propias de los sistemas de información geográfica.
MATERIAL Y MÉTODOS
El área de estudio correspondió al tramo del río Paraná entre la represa Yacyretá y la confluencia con el río Paraguay, seleccionándose las lagunas Galloso y Melilla, las cuales están separadas por una distancia de 564 m debido a la interposición de un brazo del río Paraná (Figura 2). Ambos cuerpos de agua fueron digitalizados y se calcularon su valores morfométricos.
Las lagunas Galloso y Melilla poseen una significativa diferencia en la diversidad de especies de peces (80 y 49 respectivamente), pese a encontrarse muy próximas entre sí, poseer una misma génesis y presentar valores morfométricos relativamente similares.
Tomando el día 27/05/2008 como de aguas altas y el 26/08/2013 como de aguas bajas (según disponibilidad de imágenes de alta definición), se procedió a calcular las siguientes variables morfométricas:
- Longitud Máxima (LM): es la distancia, en línea recta, entre los dos puntos más distantes de las orillas de las lagunas. Esta distancia representa la longitud máxima de agua continua de la superficie de la laguna con la cual puede interactuar el viento.
- Ancho Máximo (AM): corresponde a la distancia máxima entre las orillas de la laguna, en un ángulo recto con la línea de longitud máxima. La anchura media es igual al área dividida por la longitud máxima.
- Perímetro (PE): es la línea de intersección de la tierra con el agua.
- Superficie (SU): área correspondiente a la cubeta del cuerpo de agua medida en m2.
- Desarrollo de la Línea de costa (DL): es el cociente entre el perímetro del lago y la superficie que se obtiene a través de la siguiente fórmula. DL=P/2.√(A.π). Posteriormente se clasificarán a los cuerpos de agua según sus formas.
- Cociente de elasticidad (CE): es el resultado del cociente entre la superficie del cuerpo de agua en un período húmedo y un período seco.
- Índice de cambio (IC): es el promedio del cociente entre LM, AM, PE y SU del año húmedo y los valores del año seco: (PH/PS).(SH/SS).(LMH/LS).(AMH/AS)/4. Los resultados esperados son: sin cambios = 1, cambios moderados = 1,1 a 1,9 y cambios significativos = +2.
Con los valores de DL se efectuó una clasificación de formas de la cubeta teniendo en cuenta las categorías establecidas por Hutchinson 10 y Timms 16. Mediante el software Global Mapper 15.1 se obtuvo una imagen SRTM de 90 m con la cual se generó un modelo de anegamiento que permitió determinar las diferencias topográficas entre ambas lagunas y sus posibles respuestas frente a las crecientes del río Paraná.
RESULTADOS Y DISCUSIÓN
Considerando la sumatoria de los valores de superficie de ambas lagunas, solo representarían el 0,004% en aguas bajas y 0,05% en aguas altas de la superficie total del área de estudio, estimada en 2.064 km2.
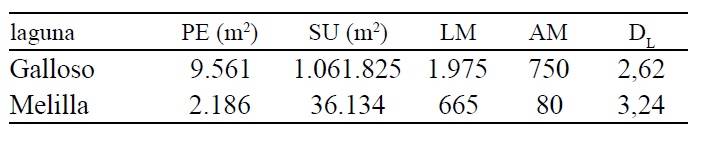
Sin embargo, las diferencias significativas de las variables morfométricas entre las altas y bajas aguas, suceden en la laguna Galloso, mientras que en la laguna Melilla, no se observan cambios considerables (Tablas 1 y 2).
Teniendo en cuenta el DL, si bien ambos cuerpos de agua poseen forma compleja, irregular en el caso de la laguna Galloso y dendrítica para la laguna Melilla, con la mera observación parecerían adoptar la misma morfología. Sin embargo esta última posee un mayor desarrollo del perímetro por poseer un canal conector mucho más angosto. En este sentido, en trabajos anteriores se estableció que existe una mayor correlación del DL con el perímetro y no con la superficie, lo cual evidencia que independientemente de su tamaño, las lagunas pueden adoptar diferentes formas.
Las formas complejas, que superan el valor de 2 DL, son típicas del valle de inundación 14. Atendiendo a una clasificación antemencionada 16 tanto las de forma triangular, como dendríticas e irregulares tienen su origen en la inundación del terreno. El valor máximo calculado para este tipo de lagunas es 7,61 14 y ellas se caracterizan por la presencia de altos valores que posteriormente van adquiriendo formas circulares a medida que evolucionan hacia una senescencia, pasando por un ambiente de pantano y finalmente a terrestre (proceso de terrestralización o colmatación). Esta situación se aprecia en las Tablas 1 y 2, ya que durante aguas bajas los valores de DL descienden en ambas lagunas.
Tabla 1. Valores morfométricos de las lagunas, año 2013.

Tabla 2. Valores morfométricos de las lagunas, año 2008.
Por otra parte, este proceso puede verse interrumpido cuando ocurren los pulsos de inundación 11, e ir variando la importancia espacial que ocupan las aguas de acuerdo a procesos climáticos como ENSO (El- Niño-Southern-Oscillation); factores que generan un "reseteo" 15, en el cual el ambiente de pantano, geomorfológicamente hablando, vuelve a su estado anterior como laguna.
En la Figura 3 quedan en evidencia las marcadas diferencias morfométricas entre ambas lagunas. Los valores de CE e IC de la laguna Galloso fueron 19,4 y 8,8; mientras que la laguna Melilla poseía 1 y 1 respectivamente. Considerando que en ambas variables el valor de 1 indica que el cuerpo de agua mantuvo su misma forma y tamaño y que por el contrario, a partir de 1,9 de IC ya se consideran como cambios significativos, un valor de 8,8; sumado a que el CE está indicando que en momentos de aguas altas la laguna Galloso puede albergar 19,4 lagunas con su misma superficie que en aguas bajas, evidencia que esta última posee un dinamismo mucho más marcado. Por su parte, esta situación incidirá en las características físico-químicas de sus aguas, generando así un ambiente mucho más heterogéneo. En este sentido, el balance de las comunidades, los patrones de biodiversidad y la organización funcional dependerían del efecto anual de la crecida que transporta una gran cantidad de carbono orgánico disuelto alóctono que sufre un proceso de metabolización 5, 8.
Esta variación del carbono disuelto en la planicie, se correlaciona con los cambios en el nivel hídrico 7, con el contenido de materia orgánica acumulada en los suelos inundados y con la tasa de descomposición de la materia orgánica particulada. Moviliza y resuspende los nutrientes contenidos en los sedimentos y aporta a la productividad primaria de algas y macrófitas 6. A su vez el detritus formado y acumulado, soporta una rica invertebrofauna y representa la base trófica de varias especies de peces. Al iniciarse la estación seca, importantes cantidades de este carbono disuelto y particulado son exportados al cauce principal.
La generación de una simulación de anegamiento teniendo como punto de partida un modelo de elevación digital (Figura 4), demostró que la laguna Galloso se encuentra 2 m por debajo de la laguna Melilla. Este dato es muy significativo, ya que establece que la laguna Galloso se encuentra más expuesta frente a los pulsos de creciente del río. En otras palabras, el recambio de las aguas se produce mucho antes y culmina mucho después.
En síntesis, si el nivel de la creciente no supera los 2 m, no existirá una renovación de las aguas, con lo cual la laguna Melilla puede mantenerse aislada del río Paraná, al mismo tiempo que la laguna Galloso se encuentra total o parcialmente conectada. Por otra parte, se puede apreciar el parecido de la morfología que posee la laguna Galloso en el año 2008 (Figura 3) con la simulada a cota de 59 m (Figura 4).
Esta situación no resulta menor, ya que estaría explicando la mayor diversidad de peces en esta última laguna mencionada. Cabe destacar que las especies más abundantes de la laguna Galloso (Figura 5) fueron Odontostilbe pequira, Astyanax lacustris, Serrapinus calliurus, Moenkhausia intermedia y Psellogramus kennedyi, siendo Odontostilbe pequira, cinco veces más abundantes que las demás; mientras que en la laguna Melilla (Figura 6) las especies más abundantes fueron Moenkhausia intermedia, Poptella paraguayensis, Odontostilbe pequira, Moenkhausia dichroura y Otocinclus vestitus.
En definitiva, resulta de interés hacer un seguimiento de las formas de estas lagunas, ya que sus cambios morfológicos pueden suceder de un pulso de inundación a otro y dicha situación incide en la diversidad de peces.
Agradecimientos. Este trabajo fue financiado por la Secretaría General de Ciencia y Técnica (UNNE, PIB009- 2014) y por el convenio Entidad Binacional Yacyretá – Universidad Nacional del Nordeste.
REFERENCIAS
1. Abdo MS, Silva CJ. 2004. Limnological characteristics of the water bodies of the Corutuba Nesting Site in Brazil’s Pantanal. Acta Limnol Brasil 16: 359-368.
2. Agostinho AA, Gomes LC, Zalewski M. 2001. The importance of floodplains for the dynamics of fish communities of the upper river Paraná. Intern J Ecohydrol Hydrobiol 1: 209-217. [ Links ]
3. Albert JS, Carvalho TP. 2011. Neogene assembly of modern faunas. In: Historical Biogeography of Neotropical Freshwater Fishes (Albert JS, Reis RE, ed.), University of California Press, p. 119-136. [ Links ]
4. Alho CJ. 2008. Biodiversity of the pantanal: response to seasonal flooding regime and environmental degradation. Braz J Biol 68: 957-966. [ Links ]
5. Baigún C, Oldani N, Nestler J. 2005. Integridad ecológica en los ríos Paraná y Mississippi: ¿trayectorias paralelas o divergentes? Miscel Insugeo 14: 91-104. [ Links ]
6. Bayley PB. 1995. Understanding large river-floodplain ecosystems. BioScience 45: 153-158. [ Links ]
7. Depetris PJ, Cascante E. 1985. Carbon transport in the Parana River. In: Transport of carbon and minerals in major world rivers (Degens ET, Kempe S, Herrera R, eds.), Mitteilungen des Geologish-Paläeontologischen Instituts der Universität Hamburg, SCOPE/UNEP Sonderband 52: 385-395. [ Links ]
8. Depetris PJ, Paolini JE. 1991. Biogeochemical aspects of South American Rivers: The Paraná and the Orinoco. Mitteilungen des Geologish-Paläeontologischen Instituts der Universität 42: 105-125. [ Links ]
9. Granado DC, Raoul H. 2012. Changes in abiotic characteristics of water in the Paranapanema River and three lateral lagoons at mouth zone of the Jurumirim Reservoir during the flood period, São Paulo, Brazil. Latin Am J Aquat Res 40: 79-89. [ Links ]
10. Hutchinson G. 1957. A treatise on limnology: geography, physics and chemistry, Vol. 1, Chapman and Hall, London. [ Links ]
11. Junk W, Bayley P, Sparks R. 1989. The flood pulse concept in river-floodplain systems. In: Proceedings of the International Large River Symposium LARS (Dodge D.P. ed.), 110-127. [ Links ]
12. Mirande JM, Koerber S. 2015. Checklist of the freshwater fishes of Argentina (CLOFFAR). Ichthyol Contribut Peces Crioll 36: 1-68. [ Links ]
13. Neiff JJ. 1990. Ideas para la interpretación ecológica del Paraná. Interciencia 15: 424-441. [ Links ]
14. Paira AR, Drago E. 2007. Origin, evolution, and types of floodplain water bodies. In: The Middle Parana River Limnology of a Subtropical Wetland (Iriondo M, Paggi J, Parma M, eds.), Springer, p. 53-81. [ Links ]
15. Sparks R, Bayley P, Kohler S, Osborne L. 1990. Disturbance and recovery of large floodplain rivers. Environ Manag 14: 699-709. [ Links ]
16. Timms B. 1992. Lake geomorphology, Gleneagles Publishing, Adelaide. [ Links ]
17. Townsend CR. 1989. The patch dynamics concept of stream community ecology. J North Am Benthol Soc 8: 36-50. [ Links ]
18. Welcomme RL. 1985. River Fisheries, In: FAO Fisheries Technical Paper, Food and Agriculture Organization of the United Nations, Nº 262. [ Links ]

