











 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
Permalink
Introducción
Echinococcus vogeli (Rausch y Bernstein 1972) y E. oligarthra (Diesing 1863) ocasionan la Equinococosis Neotropical (EN), enfermedad zoonótica parasitaria emergente en países tropicales de Centro y Sur América. E. vogeli origina la equinococosis neotropical poliquística (ENP) que por su frecuencia y por la patología que produce en el hombre, es la más importante de las dos especies neotropicales (OPS/OMS 2015, D’Alessandro y Vizcaychipi 2019, Vuitton et al. 2020). El ciclo natural de E. vogeli está adaptado a una relación predador - presa entre el perro de monte o zorro pitoco (Speothos venaticus, Lund 1842), único hospedador definitivo natural conocido, y el majaz o paca (Cuniculus paca, Linnaeus, 1766) principal hospedador intermediario y presa predilecta del primero (D’Alessandro y Rausch 2008). El ciclo de vida silvestre se inicia cuando S. venaticus elimina los huevos de E. vogeli por materia fecal hacia el medio ambiente. Posteriormente estos son ingeridos por su hospedador intermediario, donde se desarrolla el metacestode, principalmente en hígado y generalmente visible en la superficie del órgano. Este ciclo se completa cuando la presa es ingerida por el predador, el S. venaticus, desarrollando las formas adultas del parásito en el intestino delgado (D’Alessandro et al. 1981, D’Alessandro y Rausch, 2008).
La enfermedad en los humanos se da por la ingesta accidental de huevos del parásito presentes en suelos, vegetales, frutos silvestres y aguas contaminadas o por contacto directo con perros domésticos parasitados con E. vogeli (D’Alessandro et al. 1981, Noya-Alarcón et al. 2011, Kern et al. 2017).
El majaz es un roedor grande (peso promedio 7,0 kg), sus hábitos alimenticios son vegetarianos y se lo encuentra desde el sur de México hasta la zona tropical de Perú, Bolivia, Paraguay, Brasil y Norte de Argentina principalmente en la provincia de Misiones (Vizcaychipi et al. 2013). Debido a la presión de caza y a la destrucción de su hábitat la densidad poblacional de este roedor, al igual que la del armadillo de nueve bandas (Dasypus novemcinctus) y el añuje (Dasyprocta spp.), está en descenso (Moya 2011, Cobos et al. 2013, El Bizri et al. 2020). En la Amazonía, el majaz es una de las especies más frecuentemente cazadas con fines de subsistencia y comercialización, siendo la caza de subsistencia considerada una actividad legal (El Bizri et al. 2020, Mayor et al. 2022). En la ciudad de Iquitos se comercializan aproximadamente 82.200 kg/año de carne de majaz (equivalente aprox. 10.200 individuos) (Moya 2011, Cobos et al. 2013, El Bizri et al. 2020). Estos datos evidencian la existencia de un riesgo de transmisión del ciclo silvestre de E. vogeli a poblaciones humanas y animales domésticos (D’Alessandro 2010, Noya-Alarcón et al. 2011, D’Alessandro y Vizcaychipi 2019). El objetivo del presente trabajo consistió en estudiar la presencia de E. vogeli en majaces cazados para consumo familiar por pobladores de comunidades de la cuenca alta del Río Itaya.
Materiales y métodos
Área de estudio. Elestudio fue llevadoacaboencuatro comunidades situadas enlacuencaalta del río Itaya, al sureste de la ciudad de Iquitos, entre los ríos Amazonas, Marañón y Nanay: Melitón Carbajal (4°16´1.12´´S-73°35´50.80´´O), Luz del Oriente (4° 16´56.96´´S-73°38´3.87´´O), 28 de Enero (4°16´17.67´´S-73°35´31.67´´O) y Nueva Villa Belén (4°16´49.44´´S-73°42´46.33´´O) (Figura 1).
La fisiografía de la cuenca alta del Río Itaya está conformada por bosques de colinas bajas con pendientes que varían entre 15 y 70% según presencia de cauces de quebradas, disectando la superficie por acción erosiva. Estas quebradas le proporcionan una gran riqueza de hábitats de flora y fauna, así como cuerpos de agua, aguajales, con una gran diversidad en especies acuáticas, de aves y reptiles. En el área de estudio, las precipitaciones y temperaturas anuales oscilan entre los 2500 a los 3000 mm y entre los 27°C a los 40°C respectivamente (Cobos et al. 2013, El Bizri et al. 2020).
Colecta de muestras biológicas. Se realizó un muestreo por conveniencia supeditado a las circunstancias de la captura de majaces en su medio natural. Previo a la colecta del material biológico, en cada comunidad de estudio se identificaron cazadores voluntarios, que fueron capacitados en el conocimiento, uso de medidas preventivas de la EN, reconocimiento y almacenamiento post-mortem de masas quísticas halladas en órganos torácico-abdominal de majaces.
La colecta de hígado y pulmones de individuosabatidos por los cazadores se realizó entre diciembre del 2016 a diciembre del 2017 (Figura 2A). Cada cazador registró el sexo y estadio etario aproximado (juvenil, adulto) del animal, fecha, lugar de caza y nombre del colector. Durante la colecta, se obtuvo de forma ocasional el hígado de un armadillo de nueve bandas. Los órganos colectados por los cazadores fueron conservados en solución de formaldehído 4% (v/v) y solo un órgano de majaz en alcohol al 70%. Las muestras fueron trasladadas al Laboratorio de Biotecnología y Bioenergética de la Universidad Científica del Perú y al Laboratorio de Histología, Embriología y Patología Veterinaria de la Facultad de Medicina Veterinaria de la Universidad Nacional Mayor de San Marcos en Lima, Perú, para sus análisis posteriores.
La colecta biológica se fundamentó en la caza de subsistencia que realizan de forma habitual las comunidades locales. Este estudio en ningún aspecto estimuló actividades ilegales e irregulares, no supuso el aumento de la presión de caza, y en ningún caso existió incentivo al ejercicio de la caza, utilizando únicamente el material biológico que los cazadores de subsistencia suelen desechar. El estudio contó con la autorización de la Dirección Ejecutiva Forestal y Fauna Silvestre (007-2017-GRL-GGR-ARA-DEFFS).
Análisis parasitológico. Macroscópicamente se estudió la localización, número, tamaño y aspecto de los quistes presentes en cada hígado o pulmón. La observación microscópica incluyó el análisis descriptivo y morfométrico de los ganchos rostelares de la larva utilizando el método de Raush et al. (1978). Se utilizó microscopio óptico con objetivo de inmersión (100x) y ocular micrométrico calibrado. Por cada muestra de quiste se tomaron 50 µl de líquido hidatídico y se realizaron preparados en fresco entre porta y cubreobjeto aplicando sobre este último suficiente presión para provocar la separación de los ganchos rostelares de protoescólices. Se seleccionaron al azar y se tomaron longitudes de los ganchos grandes y pequeños, evaluándose un total de 50 ganchos. En la medición de los ganchos se consideró la medida de longitud total de los ganchos grandes (LGG) y la de los ganchos pequeños (LGP), longitud del mango (LM), longitud de la hoja (LH) y la relación porcentual del tamaño hoja/mango (Raush et al. 1978, D’Alessandro et al. 1981).
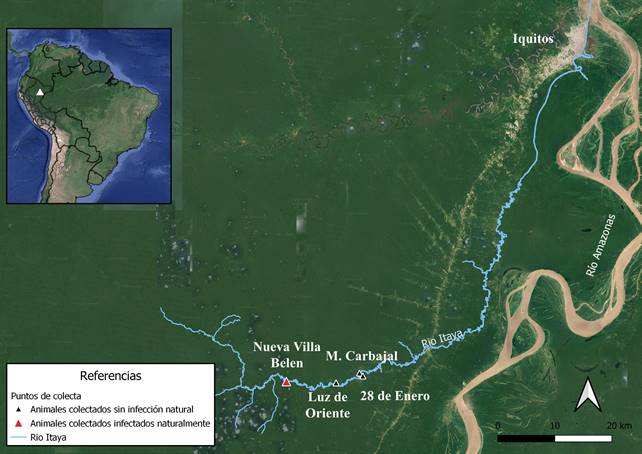
Figura 1 Mapa indicando las zonas geográficas de las comunidades en la cuenca alta del Río Itaya, Perú donde se realizaron las colectas de majaces y armadillo de nueve banda durante el periodo 2016 - 2017. Como referencia, se muestran las comunidades que presentaron individuos infectados naturalmente por E. vogeli (triángulo rojo) y no infectados (triangulo negro).
Análisis estadístico. Se evaluó la asociación de la presencia de quistes en mamíferos entre diferentes comunidades, edad y sexo mediante la prueba de Chi- cuadrado con un nivel de significancia de 0,05. Para la comparación de la longitud de los ganchos rostelares entre los hospedadores intermediarios se utilizó el test t de Student para muestras independientes y se verificaron los supuestos correspondientes. Los resultados se presentan como media e intervalo de confianza 95% (IC 95%). Se utilizó Microsoft Excel 2010 para el análisis estadístico. El mapa que representa las comunidades donde se efectuaron las colectas fue generado empleando el software QGIS 3.8, Google Maps como mapa de referencia y GEO GPS Perú para los ríos.
Resultados
De las 4 comunidades estudiadas, se identificaron y capacitaron a 8 cazadores, 2 por cada comunidad. Se colectaron vísceras de 36 majaces (36 pulmones, 32 hígados) y el hígado de un armadillo de nueve bandas. La comunidad de Nueva Villa Belén presento el mayor número de colecta de majaces (n=24) incluido el armadillo de nueve bandas (n=1), seguido por la comunidad 28 de Enero (n=8); Luz de Oriente (n=2) y Melitón Carbajal (n=1) (Figura 1).
Del total de los animales obtenidos por los cazadores, sólo 25 majaces contaron con ficha de registro, 20/25 correspondieron a individuos adultos (14 machos; 6 hembras) y 5/25 a juveniles (1 macho; 4 hembras), el armadillo de nueve bandas correspondió a un individuo macho adulto de aproximadamente 8 kg de peso. La época de mayor caza se registró durante los meses de temporada húmeda (noviembre a abril).
El 13,5% (5/37) de los mamíferos silvestres colectados (majaz (n=4); armadillo de nueve bandas (n=1)) presentaron quistes hepáticos blanquecinos de 0,5-3,0 cm de diámetro, semisólidos al tacto, con el aspecto típico de hidátides poliquísticas de E. vogeli (Figura 2 D-E). Los majaces parasitados correspondieron a individuos machos (3 adultos y 1 juvenil) provenientes de la comunidad Nueva Villa Belén, al igual que el armadillo de nueve bandas (Figura 1). De los individuos evaluados ninguno presento masas quísticas en los pulmones.
En relación a los individuos colectados por comunidad, no se observó asociación estadística en función de la comunidad y animales parasitados (p= 0,82), ni entre infección natural y edad aproximada (p= 0,54) o el sexo (p= 0,40) de los majaces evaluados.
En el análisis microscópico en fresco del líquido hidatídico se observaron protoescólices, corpúsculos calcáreos y ganchos rostelares (Figura 2F-G). En los majaces infectados (n=4) la medición promedio de los ganchos rostelares fue de LGG 40,9 ± 0,3 µm; LGP 32, µm ± 0,5 µm, LM 14,1 ± 0,5 µm y LH 26,7 ± 0,5 µm. Para el armadillo de nueve bandas las mediciones promedio de los ganchos rostelares fueron de LGG 40,6 ± 1,6 µm, LGP 33,8 ± 1,6 µm, LM 15,6 ± 1,6 µm y LH 22,9 ± 1,6 µm. La comparación estadística de las longitudes promedio de los ganchos grandes y chicos entre ambas especies de mamíferos no presentó diferencias estadísticamente significativas. En base a la clasificación morfométrica, el tamaño, forma y proporciones de los ganchos rostelares de protoescólices obtenidos de los majaces y del armadillo de 9 bandas son compatibles con E. vogeli (Tabla 1, Figura 2G).
Tabla 1 Longitud total (en micrones) de los ganchos rostelares de protoescólices de E. vogeli prevenientes de hospedadores intermediarios de la Cuenca alta del Río Itaya, Perú en comparación con las medidas patrón.
| Especie y hospedador | 68.LGG 69. 70. | 71.LGP 72. 73. | 74. 75. 76.||||
|---|---|---|---|---|---|---|
| Mín. | 77.Máx. | 78.X̅ | 79.Mín. | 80.Máx. | 81.X | 82. 83.|
| E. vogeli | 84. 85. 86. 87. 88. 89. 90. 91. 92.||||||
| Majaz 1 | 93. 94.40,1 | 95.42,1 | 96.41,2 | 97.24,2 | 98.35,0 | 99.32,8 | 100. 101.
| Majaz 2 | 102. 103.40,2 | 104.42,4 | 105.40,6 | 106.23,2 | 107.34,2 | 108.31,8 | 109. 110.
| Majaz 3 | 111. 112.39,1 | 113.41,2 | 114.40,9 | 115.30,7 | 116.35,3 | 117.32,8 | 118. 119.
| Majaz 4 | 120. 121.41,2 | 122.43,0 | 123.42,1 | 124.24,2 | 125.34,2 | 126.31,1 | 127. 128.
| Armadillo de nueve bandas | 129. 130.38,3 | 131.42,6 | 132.40,8 | 133.31,5 | 134.35,3 | 135.33,8 | 136. 137.
| Medidas patron* | 138. 139.38,2 | 140.45,6 | 141.41,8 | 142.30,4 | 143.36,9 | 144.33,1 |
LGG: Longitud Gancho Grande; LGP: Longitud Gancho Pequeño. *Rausch, Rausch y D’Alessandro 1978.
Discusión
Este estudio, llevado a cabo en la cuenca alta del Río Itaya, confirma morfológicamente nuevos registros de E. vogeli en poblaciones de majaz en su rango geográfico natural de distribución y confirma el primer reporte de E. vogeli en armadillo de nueve bandas en la Amazonia peruana, aportando al mayor conocimiento epidemiológico de la EN.
La EN, con más de 260 casos humanos distribuidos en 15 países tropicales de América Central y del Sur (Nicaragua, Costa Rica, Panamá, Colombia, Venezuela, Suriname, Guayana Francesa, Ecuador, Brasil, Perú, Bolivia, Chile, Paraguay, Uruguay, Argentina) presenta una mortalidad para la ENP del 15,5% al 29,0% (D’Alessandro et al. 1979, D’Alessandro 2010, OPS/OMS 2015, Kern et al. 2017, D’Alessandro y Vizcaychipi 2019, Vuitton et al. 2020).
En Perú existe poca información sobre esta zoonosis. Se describe un caso humano en Contamana (Departamento de Loreto) ubicado en la selva central del Perú (Somocurcio et al. 2004). No obstante la prevalencia de esta enfermedad principalmente en áreas de selva como la Amazonia puede estar siendo subestimada (D’Alessandro 2010, Siqueira et al. 2013, Kern et al. 2017). Respecto a los hospedadores silvestres, los estudios de E. vogeli se limitan a dos reportes. El primero, realizado en el mercado de Belén, Iquitos donde se informa metacestode de E. vogeli en cuatros hígados de majaz Tantalian et al. (2012) y el segundo al realizado por Mayor et al. (2015) en dos comunidades de la Amazonía peruana (Nueva Esperanza y Diamante/7 de Julio), donde encontraron un 11,7% (15/128) de majaces infectados con E. vogeli. Estando este trabajo en coherencia con la epidemiologia de la enfermedad (Thatcher 1972, Gardner et al. 1988, D’Alessandro et al. 1979, Soares et al. 2004, Somocurcio et al. 2004, D’Alessandro 2010, Noya-Alarcón et al. 2011, Vizcaychipi et al. 2013, Mayor et al. 2015, das Neves et al. 2017, Kern et al. 2017, Vuitton et al. 2020).
En la Amazonia en general y en la peruana en particular ambos hospedadores intermediarios (majaz y armadillo de nueve bandas) se encuentran en la zona de distribución del S. venaticus, único hospedador definitivo natural conocido de E. vogeli (das Neves et al. 2017, D’Alessandro y Vizcaychipi 2019). Teniendo en cuenta la relación especie especifica parásito-predador y sus hábitos alimenticios predador-presa, el hallazgo en este estudio de un armadillo de nueve bandas naturalmente infectado con E. vogeli como el de cinco ejemplares de armadillo de nueve bandas descritos por otros autores en la cuenca del Amazonas oriental del Brasil (Soares et al. 2004, Santos et al. 2012, Souza et al. 2022), confirma que este mamífero placentario de la familia Dasypodidae actuaría como hospedador intermediario natural, requiriéndose mayores estudios para entender su rol en el ciclo de E. vogeli.
En el ámbito geográfico de E. vogeli, el perro doméstico es un hospedador definitivo adecuado y puede cumplir el rol principal de transmisor de la enfermedad a humanos cuando son alimentados con vísceras crudas de majaz naturalmente infectados (D’Alessandro 2010, Souza et al. 2022). Las observaciones de campo muestran que los pobladores de la cuenca alta del río Itaya ofrecen a sus perros domésticos vísceras crudas de los animales cazados, principalmente las vísceras del majaz, representando un riesgo epidemiológico potencial en salud pública para la infección por E. vogeli. No obstante, la presencia de E. vogeli en el perro domestico aún es un interrogante a ser investigado en comunidades de la Amazonía.
Si bien la EN guarda un gran componente geográfico cultural de riesgo: el hábito de caza de mamíferos silvestres principalmente majaz para la ENP y vivir o haber vivido en zonas de forestas tropicales, bosques de galería, con presencia de sus hospedadores silvestres definitivo e intermediario (D’Alessandro et al. 1979, Kern et al. 2017), también se debe tener en cuenta otros factores de riesgo asociadoscomo ser, elcontexto social enelcual viven; la falta de disponibilidad de agua potable que lleva a condiciones higiénico personales y alimentarias deficientes; dificultad de acceso a centros de salud, atención médica inadecuada y conocimiento insuficiente sobre enfermedades tropicales locales (D’Alessandro 2010, Noya-Alarcón et al. 2011, Vizcaychipi et al. 2013, Mayor et al. 2015, das Neves et al. 2017, Kern et al. 2017, D’Alessandro y Vizcaychipi 2019, Vuitton et al. 2020, Souza et al. 2022). Ante esta situación, donde la salud en estas comunidades es una cuestión social compleja, puesto que se relaciona específicamente con las condiciones de vida en los órdenes económico, ambiental, cultural y político, este estudio refuerza la necesidad de concientizar, prevenir, monitorear, controlar y ampliar la búsqueda de la EN y otras zoonosis en fauna silvestre y en poblaciones humanas en coexistencia, desde el enfoque integral y multisectorial de “Una Salud”

Figura 2 A. Majaces obtenidos por caza de subsistencia en la cuenca alta del río Itaya, Perú. B. Majaz. C. Armadillo de nueve bandas. D. y E. Masas poliquísticas de E. vogeli (metacestodes) en hígado de majaz (D) y en armadillo de nueve bandas (flechas) (E). F y G. Análisis microscópico y morfométrico de metacestode de E. vogeli en armadillo de nueve bandas, protoescólice invaginado (F) y ganchos rostelares grandes y pequeños de protoescólice de E. vogeli (G) mostrando sus características distintivas (observación en fresco, 100 X).

