










Servicios Personalizados
Revista

Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkRevista argentina de neurocirugía
versión On-line ISSN 1850-1532
Rev. argent. neurocir. v.22 n.2 Ciudad Autónoma de Buenos Aires abr./jun. 2008
Esclerosis temporal mesial: Paradigma de la epilepsia de resolución quirúrgica
Premio júnior NeuroRaquis 2007
Claudio Vázquez, Mariano Cuello, Pablo Rubino, Alberto Prosen, Necmettin Tanriover, Mónica Perasolo, Manuel Martínez, José C. Morales, Hugo Pomata
Hospital General de Agudos "Dr. Cosme Argerich", Hospital de Pediatría "Prof. Dr. Juan P. Garrahan", FLENI, Hospital General Interzonal de Agudos "Prof. Dr. Rodolfo Rossi", Fundación Científica del Sur.
Correspondencia: clauvazquezl4@hotmail.com
Recibido: mayo 2008;
aceptado: mayo 2008
RESUMEN
Objetivo. Describir la Esclerosis Temporal Mesial (ETM) en sus aspectos anatómico, fisiopatológico, clínico, imagenológico, neurofisiológico, neuropsicológico y quirúrgico; y su relación con la epilepsia de resolución quirúrgica. Luego analizar estadísticamente nuestra casuística y resultados.
Material y método. De los 469 (115 adultos y 354 niños) pacientes intervenidos quirúrgicamente por presentar epilepsia entre los años 1989-2007 en el Hospital Dr. Juan P. Garrahan, FLENI, Hospital Dr. Cosme Argerich y Hospital Prqf. Dr. Rodolfo Rossi, se analizan 91 pacientes(19,4%) con el diagnóstico de ETM; 38 de los cuales (41.75%) son niños y 53 (58.25%) adultos
Resultados. Los resultados son evaluados mediante el score de Engel. Aplicando esta clasificación, nuestra población tiene la siguiente distribución: 69 pacientes (75,8 %) se encuentran en clase IA de Engel, de ellos 36 son niños (52,2 %) y 33 adultos (47,8 %); 4 pacientes adultos se encuentran en Engel IB (4,4%), 3 en Engel IC (3,3%), 1 adulto en Engel ID (1,1 %), 4 adultos en Engel IIA (4,4%), 5 adultos en Engel IIB (5,5%) y 3 pacientes en Engel IVA (3,3 %) de ellos 1 niño a quien posteriormente se le implantó un estimulador del nervio vago y los 2 restantes adultos. Un paciente adulto se suicidó luego de un año de la cirugía (en clase IA de Engel). Un paciente se encuentra durante su primer año postoperatorio por lo cual no puede ser incluido aún en la estadística de resultados.
Comclusión. La ETM es el paradigma de la epilepsia refractaria en la población adulta y la causa de un porcentaje significativo de pacientes pediátricos sometidos a cirugía. La evaluación prequirúrgica debe ser exhaustiva para lograr la selección del paciente. La resección quirúrgica debe ser realizada con la mayor selectividad posible. El diagnóstico y tratamiento precoz permiten lograr un alto índice de curación.
Palabras Clave: Esclerosis temporal mesial; Epilepsia; Hicocampo; Engel.
Abstract
Objectives: to describe the Mesial Temporal Lobe Sclerosis (MTS), in relation to its anatomical, clinical, iconographic, neurophysiologic, neuropsychologic, and surgical aspects, in reference to the epilepsy cases that needed a surgical resolution. Additionally, It's realized a statistical analysis of our series and its results.
Material and Methods: From the series of 469 patients (115 adults and 354 children) operated on between 1989 and 2007, at National Pediátrica Prof Dr. Juan Garrahan, FLENI, Dr. Cosme Argerich, and Prof Dr. R. Rossi Hospitals; who harbored Refractory Epilepsy; were analyzed 91 cases (19,4%) with the diagnosis of MTS; 38 patients belong to the pediatric group and 53 were adults.
Results: The results were evaluated by the Engel score. Applying this classification, our population of patients showed the next pattern of distribution: 69 (75,8%) are in Engel's class IA, from this group, 36 (52,2%) are children, and 33 (47,8%), adults; 4 adults patients: ( 4,4%) are in Engel's class IB, 3 patients (3,3%) in Engel's class IC; 1 (in Engel's class ID (1,1%); 4 adults in Engel's class ILA (4,4%); 5 in Engel's class IIB (5:5%) and 3 (3,3%) in Engel's class TVA; from then, one patient was a child, and underwent the implantation of VNS, the rest were adults. One adult patient committed suicide, one year after surgery (was in Engel's class LA). One patient is on his first post operative year, and then couldn't been included in statistical analysis, because follow up wasn't enough.
Conclusión: The MTS is the paradigm of Refractory Epilepsy in the adult population, and in the pediatric subgroup involved a significant percentage: the presurgical evaluation must be exhaustive for adequate selection of cases. The extent of resection should be done with high degree of selectivity. The early diagnosis and treatment can obtain a high índex of good results without íctal phenomena and the absence of necessity of antiepileptic drugs.
Key words: Mesial temporal sclerosis; Epilepsy hyppocampus; Engel.
INTRODUCCIÓN
La epilepsia es una afección de la cual existen registros antiguos; ya en la Mesopotamia, en 5000 aC, se alude a personas que presentaban crisis epilépticas. En algunas lápidas de piedra halladas en Babilonia, se han encontrado inscripciones con observaciones detalladas sobre epilepsia, distintos tipos de crisis y sus factores desencadenantes. Para los griegos era considerada un fenómeno sobrenatural, una enfermedad sagrada; entendiendo que solo los dioses eran capaces de producir ese comportamiento en una persona y luego devolverlo a la vida. En el período grecorromano se creía que era controlada por los cuerpos celestes y particularmente por la luna. Los romanos suspendían los comicios si alguno de ellos era afectado por epilepsia durante los mismos, de ahí el nombre de mal comicial. Es descripta con precisión por los autores clásicos como Celso, Galeno e Hipócrates. Este último en 450 a.C. discute las ideas místicas al respecto, sosteniendo que la enfermedad reconocía causas naturales. En las culturas precolombinas también era reconocida esta entidad. Los incas y aztecasl consideraban que la causa de las enfermedades, incluyendo a la epilepsia, se debía a un disturbio en la relación establecida con las fuerzas sobrenaturales. En la cultura China, las primeras descripciones aparecen en el Clásico de medicina interna del Emperador Amarillo, escrito entre 770 a 221 a.C. La primera teoría considerada por ellos fue la causa congénita, siendo contraída en el útero materno, cuando la madre sufría algún compromiso emocional2.
La palabra epilepsia coincide con el homónimo griego epilepsia que significa ser atacado, agarrado o capturado. Varios sinónimos han sido utilizados para señalar a esta enfermedad, por ejemplo enfermedad sagrada (Morbus sacer), enfermedad de la luna, enfermedad del que se cae, morbocomicial, mal de San Valentín, mal de corazón, perlesía y gota coral2.
Las epilepsias constituyen una enfermedad neurológica heterogénea, caracterizada por la recurrencia de crisis epilépticas, en ausencia de lesión cerebral aguda. El síndrome epiléptico es definido como la asociación de uno o varios tipos de crisis con alteraciones electroencefalográficas interictales o letales, con o sin compromiso del sistema nervioso central (SNC) y otros componentes, tales como edad de comienzo, gravedad y curso evolutivo3. El término crisis se refiere a un comportamiento anormal (con síntomas o signos) que resulta de una descarga anormal de neuronas corticales4. El origen topográfico de esta afección puede ser variado, siendo la manifestación clínica un elemento útil para reconocerlo.
Las epilesias originadas en el lóbulo temporal en particular, constituyen en sí mismas un síndrome epiléptico. Incluyen crisis originadas en las estructuras mesial/basal o la neocorteza temporal lateral. Son las más frecuentes de las epilepsias de origen focal, alcanzando el 80% de todas las crisis; el 90% son originadas a nivel temporal mesial3. La esclerosis temporal mesial (ETM) es la entidad epileptógena más frecuente en esta localización en los adultos. Presenta pobre respuesta al tratamiento médico, por lo que su resolución está estrechamente ligada a la cirugía. Debido a la documentada coexistencia de la ETM con otras lesiones a nivel temporal (patología dual) y al compromiso de la región neocortical en los abordajes al sectormesial, para tratar esta entidad se requiere un conocimiento anatómico, neuroñsiológico y funcional del lóbulo temporal en su totalidad.
ANATOMÍA MICROQUIRÚRGICA DEL LÓBULO TEMPORAL5 6 7 8 9 10
Los hemisferios cerebrales, se encuentran divididos en lóbulos, cuya delimitación se basa en relación con el hueso que lo cubre, pero como veremos más adelante, tomando corno ejemplo el lóbulo temporal, no existe una real separación entre los diferentes lóbulos, que componen un hemisferio cerebral. El lóbulo temporal presenta límites un tanto arbitrarios. En la superficie lateral del cerebro, se encuentra separado de los lóbulos frontal y parietal, por la raíz posterior de la fisura silviana, hacia atrás se continua con el lóbulo occipital sin una demarcación clara, pero existe una línea imaginaria que se llama línea parietotemporal, que va desde la incisura preoccipital hasta la impresión en la superficie lateral del surco parietooccipital, que lo separa del lóbulo occipital. Por otro lado si trazamos una línea desde la raíz posterior de la fisura silviana hasta la parte media de la línea parietotemporal, completamos la separación superior con el lóbulo parietal, esta última línea se llama línea temporoccipital. En la cara basal también el lóbulo temporal se continúa con el occipital, por lo tanto, la proyección basal de la línea parietotemporal, lo separa del lóbulo occipital.
Todas las caras del lóbulo temporal se encuentran en continuidad con el resto de los hemisferios. Como ya observamos, su cara lateral y basal se continúan con el occipital, su cara superior se conecta con el lóbulo frontal a través del limen de la ínsula y por otro lado la amígdala lo conecta con el putamen. La cara medial se continúa hacia atrás y arriba con el cíngulo y hacia atrás y abajo con la circunvolución lingual del lóbulo occipital.
Si logramos separar el lóbulo temporal del resto del hemisferio, observamos que el mismo presenta cuatro superficies, lateral, superior, basal y medial o mesial. La superficie lateral presenta tres circunvoluciones temporales; superior, media e inferior, separadas por dos surcos temporales; superior e inferior, la circunvolución inferior se continua en la cara basal, donde encontramos tres circunvoluciones, ellas son de lateral a medial: temporal inferior, fusiforme y parahipocampal. Éstas están separadas por lo surcos occipitotemporal y colateral. El surco colateral se continúa anteriormente con el surco rinal, que lo separa del polo temporal. La cara superior tiene tres estructuras, de adelante hacia atrás ellas son, el plano polar, las circunvoluciones transversas de Heschl y el plano temporal. La cara mesial es la más compleja de todas y de importancia crucial en la cirugía de la epilepsia.
La cara mesial del lóbulo temporal tiene como componentes, varias estructuras, que podemos dividir en extraventriculares e intraventriculares. Las primeras son: la circunvolución parahipocampal, el uncus, y la circunvolución dentada, en tanto que las estructuras intraventriculares son; el hipocampo, la fimbria del fornix y el complejo amigdalino.
La circunvolución parahipocampal, marca la transición entre la cara mesial y basal, de la cual está separada por el surco colateral posteriormente y por el surco rinal anteriormente. La circunvolución parahipocampal, se continúa hacia atrás y arriba con el cingulum y hacia atrás y abajo con la circunvolución lingual del occipital. Hacia adelante se curva medialmente para dar lugar al uncus, que rodea medialmente al mesencéfalo. Hacia arriba se relaciona con la circunvolución dentada, de la cual la separa el surco hipocampal.
El uncus es la proyección medial del parahipocampo. Es de forma triangular con una superficie anterior y otra posterior, las cuales se unen en un ápex. La cara anterior presenta dos circunvoluciones, la semilunar e inferior a ésta, la ambiente. La cara posterior tiene a su vez dos superficies, una posteromedial y otra inferior, que para exponerla es necesario observarla desde abajo removiendo el parahipocampo. La cara posterior esta formada por tres circunvoluciones, que desde adelante hacia atrás son, uncinada, banda de Giacomini e intralimbica.
La circunvolución dentada, se llama así por sus típicas identaciones, se continua anteriormente con la banda de Giacomini y posteriormente con la circunvolución fasciolar, la cual a su vez sigue como el indiseum griseum, que corre por la superficie del cuerpo calloso para alcanzar la circunvolución para terminal, por debajo del pico del cuerpo calloso.
El hipocampo, se encuentra dentro del asta temporal del ventrículo lateral y forma la parte medial del mismo, tiene tres segmentos, que son desde adelante hacia atrás; cabeza, cuerpo y cola, la cabeza se caracteriza por tener tres a cuatro digitaciones, que simulan las patas de un león, también vemos que es el único segmento libre de plexo ceroideo. Se relaciona hacia adelante y arriba con la amígdala, que incluso a veces hace una prominencia que contacta con la cabeza hipocampal y lateralmente con la eminencia colateral y la proyección anterior y medial de ésta, espacio que se llama receso uncal. El cuerpo del hipocampo comienza a partir del punto coroideo inferior, que es por donde entra la arteria coroidea anterior al asta temporal, también a este nivel comienza la fimbria del fornix y es el limite inferior de la fisura coroidea, hendidura natural entre el fornix y el tálamo, de extrema utilidad e importancia quirúrgica. Lateralmente se relaciona con la eminencia colateral y medialmente con la fisura coroidea y a su través con la cisterna ambiente. La cola comienza cuando el hipocampo se adelgaza y hace una nueva curvatura medial alrededor del pulvmar del tálamo para terminar macroscópicamente a nivel del calcar avis en la pared medial del atrio ventricular, se relaciona lateralmente con el trígono colateral.
La fimbria es la parte inicial del fornix, que se forma a partir de la sustancia blanca que cubre el hipocampo, se dirige hacia atrás donde se engrosa y conforma el pilar, que rodea al tálamo, y pasando por debajo del espíenlo del cuerpo calloso, se une al del otro lado a través de la comisura hipocampal posterior, para formar el cuerpo del fornix, el cual a su vez a nivel del foramen de Monro se vuelve a separar para formar las columnas del fornix que terminan en los cuerpos mamilares.
La amígdala, sobrepasa los límites del lóbulo temporal. Dentro del mismo está contenida en lo que es considerado como uncus. Está compuesta de tres núcleos de sustancia gris, que son el basolateral, el mediocortical y el central. La amígdala se continúa superiormente con el globus basolateral, el mediocortical y el central. La amígdala se continúa superiormente con el putamen, se relaciona medialmente con las superficies anterior y posterior del uncus y hacia atrás con la cabeza del hipocampo, a la cual a veces contacta y forma a su vez la pared anterior del asta temporal del ventrículo lateral.
La fisura coroidea, es una hendidura natural entre el tálamo y fornix, tiene forma de letra C, y se extiende desde el foramen de Monro hasta el punto coroideo inferior, que se encuentra por detrás de la cabeza del hipocampo. El plexo coroideo se encuentra a lo largo de toda su extensión y se une al tálamo y al fornix a través de la tenia coroidea y la tenia fornicis respectivamente. La importancia en cirugía de la epilepsia de esta estructura es que marca un límite muy importante. Las estructuras mediales a la misma corresponden al tálamo en tanto que las laterales corresponden al lóbulo temporal.
El asta temporal del ventrículo lateral, tiene cinco paredes, una anterior que la representa la amígdala y el receso uncal, una pared lateral dada por el tapetum del cuerpo calloso, un piso formado lateralmente por la eminencia colateral (proyección ventricular del surco colateral) y medialmente por el hipocampo, recubierto en su superficie superior por la fimbria del fornix y el plexo coroideo. La pared medial la conforma la fisura coroidea. Finalmente el techo tiene dos componentes mayores el tapetum y más medialmente las radiaciones ópticas, las cuales pueden ser lesionadas durante la cirugía de la epilepsia, ya que se encuentran en intima relación con la amígdala e hipocampo (Figs. 1, 2 y 3).

Fig. 1. Lóbulo temporal en las tres caros del hemisferio. A. Vista inferior, la línea de trazos es la línea paríetotemporal basal que divide el temporal del occipital. B. Vista lateral, la línea paríetotemporal es intersectada por la línea temporooccipital, así se delimita el lóbulo temporal en la cara lateral. C. Cara mesial. I: línea paríetotemporal; 2: línea temporooccipital; 3: circunvolución temporal superior; 4: circunuolución temporal media; 5; fisura silviana; 6: circunvolución temporal inferior; 7: circunvolución fusiforme; 8: circunvolución parahipocampal; 9: uncus; 10: polo temporal.

Fig. 2. Relaciones entre el hipocampo amígdalas y el techo ventrícular. A. Las radiaciones ópticas son inmediatamente laterales a la amígdala u se ubican 3 cm por detrás del polo temporal. B. Vista superior del asta ventrícular con su techo removido. C. Vista anteroíateral que muestra la relación en el techo entre las radiaciones ópticas y las amígdalas. D. Mismo preparado vista inferior. E. Se removió el hipocampo, obsérvese la relación del asa de Meyer con las amígdalas. F. En otro preparado solóse preservaron las radiaciones ópticas. 1: radiaciones ópticas; 2: hipocampo; 3: amígdala; 4: polo temporal; 5: tapetum; 6: pulvinar; 7: putamen; 8; tracto óptico.
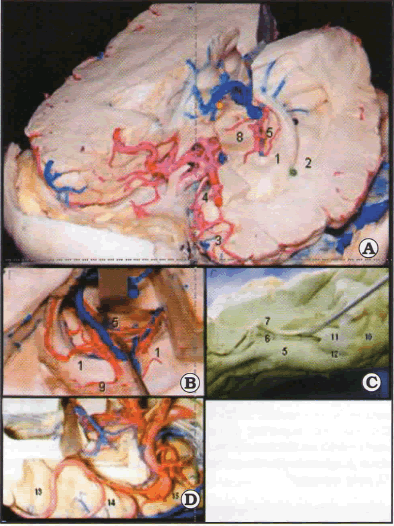
Fig. 3. Lóbulo temporal cara mesial, superior y configuración interna. A. Vista anterolateral. Se observa el piso del asta ventricular. B. Vista superior del piso, cara mesial y fisura coroidea. C. Cara mesial. D. Cara superior u opercular. 1: hipocampo; 2: eminencia colateral; 3: polo temporal; 4: uncus; 5: parahipocampo; 6: dentado; 7: fimbria; 8: mesecéfalo; 9: plexo coroideo; 10: uncus cara anteromedinl; 11: uncus cara pos terosuperior; 12: uncus caraposteromferior; 13: plano temporal; 14: circunvolución de Heschl; 15: piano polar.
Anatomía vascular del lóbulo temporal
La superficie lateral del lóbulo temporal es irrigada principalmente por ramas de la arteria cerebral media, las mismas son la arterias uncal, temporopolar, temporal anterior, las cuales emergen del segmento M1 prebifurcación. Las temporales media y posterior emergen de la rama inferior postbifurcación. La superficie inferior esta abastecida por ramas que provienen de la arteria cerebral posterior, las cuales salen a nivel de los segmentos P2 y P3 e incluyen a las arterias temporales anterior, media y posterior. La región mesial es irrigada por ramas que provienen de la arteria coroidea anterior y cerebral posterior. El hipocampo es irrigado por las arterias hipocampales que en número de 2 a 6, emergen del segmento P2 proximal de la cerebral posterior cursando a través de la cisterna ambiens. La amígdala es irrigada por ramas de la arteria coroidea anterior y la temporal anterior rama de la silviana. El parahipocampo es irrigado por las temporales anterior, media y posterior de la cerebral posterior. Finalmente el uncus es suplido por pequeñas arterias que pueden provenir de la carótida interna, cerebral media y coroidea anterior.
El drenaje venoso de la superficie lateral esta dado por venas corticales que generalmente drenan a través de la vena silviana superficial, la que a su vez drena en el seno esfenoparietal. En menor medida el drenaje puede ser hacia la vena de Labbe y a su vez al seno transverso. Una tercera posibilidad es el drenaje al seno sagital superior a través de la vena de Trolard. La vena silviana profunda corre en la profundidad de la fisura silviana y drena en el segmento estriado de la vena basal de Rosenthal, la cara mesial drena en la vena basal de Rosenthal, a través de las venas hipocampal anterior, uncal anterior y posterior, hipocampales longitudinales, ventricular inferior y coroidea inferior. La vena basal tiene tres segmentos, estriado, peduncular y mesencefálico posterior. El último segmento a nivel de la cisterna cuadrigeminal va a drenar en la vena de Galeno (Fig. 4).
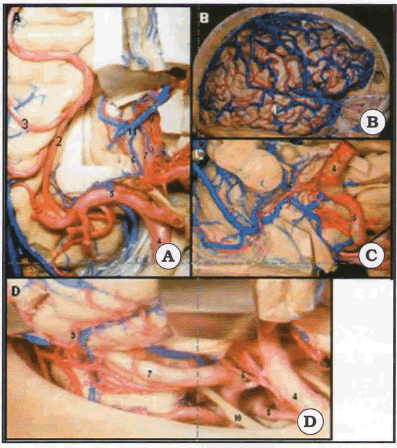
Fig. 4. Vosos en relación al lóbulo temporal. A. Cara superior principalmente irrigada por la cerebral media, B. Lo mismo en su cara lateral. C. La cara mesial anterior recibe de I segmento M1 y/ o arteria coroidea, que a su vez irriga el ventrículo entrando por el punto coroideo inferior. D. La cara mesial posterior e inferior recibe irrigación de ramas de la cerebral posterior. 1: ramas M4 de la. arteria silviana; 2: porción silviana M3; 3: ramas operculares silvianas; 4:. carótida supraclinoidea; 5: segmento silviano M1; 6: arteria coroidea anterior; 7: arteria cerebral posterior; 8: arteria comunicante posterior; 9: arteria temporal; 10: nervio oculomotor.
FISIOPATOLOGÍA DE LAS CRISIS EPILÉPTICAS ASOCIADAS A LA ESCLEROSIS TEMPORAL MESIAL
La fisiopatología de los fenómenos letales que subyacen a la esclerosis temporal mesial (ETM) son motivo de un activo trabajo de investigación, el cual involucra muchas líneas. Más allá del gran cuerpo de conocimientos en torno de esta patología, muchos son los puntos oscuros que imposibilitan una comprensión holística del fenómeno.
Las características inherentes a las epilepsias cuyo origen reside en las estructuras de la cara mesial del lóbulo temporal, depende estrechamente de las conexiones, así corno de las propiedades intrínsecas del sistema límbico. Para la adecuada comprensión de los aspectos fisiopatólogicos de la ETM es preciso tener presente las aferencias, eferencias, así como los circuitos intracorticales del hipocampo, su parcelación, y su interacción con numerosas estructuras límbicas; de ellas, con mucho, la más importante es el complejo amigdalino, verdadero punto nodal.
El concepto de lóbulo límbico se atribuye a P. Broca11 quien definió así a un grupo de estructuras filogénicamente primitivas ordenadas en torno del límite cortical (limbo), circunscribiendo al hilio del cerebro; este autor fue el que hizo hincapié en asociarlo a las funciones olfatorias, de ahí el termino que se le dio entonces de rinencéfalo. En 1937 James Papez12 propuso la idea de que las funciones emocionales residían en este disperso conjunto de estructuras12. Paul Maclean, más tarde incluyó dentro del sistema límbico al hipotálamo, al núcleo accumbens septi, al área septal, así como a diferentes sectores corticales como el área orbitofrontal y, más importante, al complejo amigdaloide13,14. Actualmente se sabe que diversas regiones antes asociadas a las funciones motoras, poseen subcompartimentos límbicos, como es el caso del neoestriado15-16. Aunque Papez y Maclean consideraron al hipocampo como una estructura central en los procesos emocionales, hoy se sabe que posee un valor indirecto y que las reacciones emocionales tienen enlace en el complejo amigdalino fundamentalmente, el cual establece un nexo entre la corteza cerebral y los centros de control visceral14,17.
Arquitectura y parcelación de hipocampo
El hipocampo, se lo puede dividir en dos regiones diferentes, el hipocampo dorsal y el ventral. El primero está representado, en el hombre, por una circunvolución rudimentaria conocida como indiseum griseum, en tanto que el ventral tiene un desarrollo mucho más importante. Su relieve en el piso ventricular está representado por el arrollamiento del arquicórtex, que asume ante la gran expansión que alcanza en el ser humano el neocortex18,19 (Fig. 5)

Fíg. 5. Visión esquemática de un corte coronal que involucra a las estructuras temporales mesiales. 1: parahipocampo; 2: subículum; 3: fisura hipocampal; 4: giro dentado; 5: surco fimbria-dentado; 6: fisura coroidea; 7: cuerpo geniculado lateral; 8: stria terminalis; 9: cola del caudado; 10: fimbria; 11: plexo coroideo; 12: asta temporal; 13: hipocampo propio; 14: eminencia colateral; 15: fisura colateral; 16: giro fusiforme.
El hipocampo del mamífero es el paradigma de la simplicidad cortical; un tipo básico de células, más sus interneuronas. Las neuronas de proyección se hallan empaquetadas en un solo estrato, configurando así un modelo de corteza trilaminar. En un corte coronal del hipocampo se puede apreciar la gradual transición entre la compleja corteza entorrinal hexalaminar y el arquicórtex. Lorente de No (1934)20; en función de las características citoarquitectónicas clasifica a la interpuesta región subicular en cuatro sectores llamados: parasubiculum, presubiculum, subiculumyprosubuculum; representados por una corteza transicional de 4 o 5 capas.
Al hipocampo mismo en un corte coronal (Figs. 5 y 6), se lo puede ver formado por dos láminas en "U" entrelazadas; la fascia dentada (área dentata) y el hipocampo propio (cornu ammonis). El hipocampo propio, basándose en diferencias citoarquitectónicas se lo subdivide en regio superior, adyacente al subículo y un regio inferior, el cual se caracteriza por poseer células piramidales gigantes, éste se arquea en torno de la fascia dentada. Lorente de No, divide al hipocampo propio en cuatro sectores; los cuales son CA1, CA2, CA3 y CA4 (Fig. 6). CA1 corresponde al regio superior; mientras que CA2 y CA3 corresponden el regio inferior y llamó CA4 a un sector conformado por escasas células rodeadas por el hilus del giro dentado. Estas células fueron incluidas en el hipocampo propio porque poseen morfología piramidal, reciben fibras musgosas y envían axones a la fimbria, pero se diferencian de las células piramidales en que no reciben "inputs" de las células en cesto.

Fig. 6. Sectores de laformación del hipocampo, vistos en un corte coronal que pasa por el cuerpo del mismo. 1: stratum granulosum del giro dentado; 2: hilus; 3: CA4; 4: CA3; 5: CA2; 6: CA1; 7: región subicular.
Con excepción de CA4, el patrón general del hipocampo es el mismo. Una lámina ordenada de grandes células de proyección, cuyas dendritas se extienden paralelamente varios micrómetros. Las fibras aferentes las cruzan en ángulos rectos haciendo múltiples sinapsis "en passage". Estas neuronas son las pirámides gigantes de CA3, las pirámides pequeñas de CA1, y las células granulosas del giro dentado. Las pirámides tienen dendritas básales y apicales, en tanto que las células granulosas tienen solo dendritas apicales20.
Laminación interna
Fascia dentada. El giro dentado posee tres estratos; la capa de células granulares, formada por neuronas esféricas, pequeñas, densamente agrupadas; la capa molecular, compuesta principalmente por axones aferentes al giro dentado y las dendritas de los granos, así como fibras de interneuronas; y por último, la capa polimorfa, en el hilus de la fascia dentada, la cual se funde con CA4, formando la folia terminal. Esta contiene los segmento axonales iniciales de las células granulares, que componen la vía de las fibras musgosas, así como diversas células no granulares, de las cuales las más importantes son las células en cesto.
Hipocampo propio. También se halla compuesto por tres láminas, la capa de células piramidales, rodeada por los estratos molecular y polimorfo, respectivamente. La sustancia blanca del hipocampo, está representada por una delgada lámina adyacente a la capa polimorfa conocida como alveus. No obstante esto, la particular distribución de fibras, así como diferentes aspectos referidos a la configuración de sus células determinan la aparición de una laminación más compleja. De esta manera, desde la superficie ventricular se pueden enumerar múltiples capas, de las cuales la primera es el alveus, sigue a este el stratum oriens, stratum pyramidale, stratum radiatum, stratum lacunosum y stratum moleculare. En el campo CA3 se puede reconocer una séptima capa, alojada entre el stratum pyramidale y el radiatum, es el stratum lucidura20.
Conectividad interna del hipocampo
Un hecho tempranamente identificado respecto a la red interna del hipocampo es que cada sector se proyecta al siguiente, configurando una vía estimulatoria trisináptica (Figs. 7 y 8). Es así que las células granulosas del giro dentado proyectan sus axones al sector CA3, formando una vía conocida como de las fibras musgosas; la zona CA2 no recibe proyecciones musgosas. Este vía glutamatérgica, y sus terminales se articulan con dendritas de las células piramidales, así como de interneuronas locales. Las grandes pirámides de CA3 originan axones que se extienden por el fórnix y envían colaterales retrógrados a CA1, los cuales hacen slnapsis con células piramidales, esta constituye la vía colateral de Schaffer. Las pirámides de CA1 se proyectan alsubículoyéste envía axones fuera del hipocampo. Esta concepción, a la luz de investigaciones recientes no parece real, por ejemplo se sabe que las fibras musgosas hacen contactos en el hilus, y que colocalizan Zn y péptidos opioides, junto al glutamato, que estos axones estimulan interneuronas del giro dentado, así como a las mismas células granulares por medio de fibras recurrentes17. Por otro lado, se han registrado influencias de las células de CA1 sobre las de CA3, probablemente por mecanismos de naturaleza antidrómica16. Bajo determinadas circunstancias, las fibras musgosas expresan glutamato descarboxilasa y proteína transportadora de GABA, manteniendo la capacidad de neurotransmisión glutamatérgica, por un mecanismo de tipo compartimentalizado, estimulando algunas células e inhibiendo otras21. Además, no debe dejarse de tener presente que todo esto se encuentra en medio de una extensa red de interneuronas, muchas de ellas gabaérgicas, configurando múltiples vías de "feed-back" y "feed-forward".

Fig. 7. Vista esquemática de los circuitos intrahipocampales. 1: célula piramidal de CA1; 2:Jíbra musgosa; 3: célula granulosa del giro dentado; 4: vía perforante; 5: intemeurona inhibitoria del hilus; 6: célula musgosa; 7: fibras de proyección fimbria/ fomix; 8: célula piramidal de CA3; 9: intemeurona inhibitoria; 10: vía colateral de Schqffer, GD: giro dentado; H: hilus; Fm: fimbria; Sub: subiculum

Fig. 8 Vista semiesquemática de los circuitos intrahipocampales. 1: célula piramidal del subiculum; 2: célula granulosa del giro dentado; 3: Fibras comisurales de las células musgosas; 4: Fibras de proyección de al sistema fimbría/fórnix; 5: célula piramidal de CA3; 6: vía de lasfigras musgosas; 7: vía colateral de Scaffer; 8: célula piramidal de CA; 9: fibras de proyección de CAÍ al subiculum; 10: fibras de proyección subiculares al área entorrinál; 11: vía perforante; 12: vía tempo-amónica.
Aferencias corticales del hipocampo
La principal vía de aferencia cortical del hipocampo está representada por las fibras que nacen en la corteza entorrinal. Estas nacen en las láminas II y III, y se extienden hasta la formación del hipocampo siguiendo la vía perforante, la cual atraviesa y obtura la fisura hipocampal, terminando dentro del estrato molecular del giro dentado y haciendo sinapsis con dendritas de las células granulares (Figs. 7 y 8). Existen diferencias entre las proyecciones de la corteza entorrinal lateral respecto de la medial, siendo esta última glutamatérgica y su actividad genera EPSP por acción de receptores NMDA y no-NMDA. La subdivisión lateral produce potenciación de larga duración (LTP) dependiente de péptidos opioides, que se presume que actúan sobre interneuronas y tienen una gran influencia en los procesos de epileptogénesis, al mediar la desinhibición de los granos. Una segunda vía de arribo cortical es la temporoamónica (Fig. 4); estas fibras nacen el la lámina III de la corteza entorrinal y se extienden al estrato molecular/lacunosum de CA1 y CA3; esta vía es glutamatérgica.
Aferencias subcorticales del hipocampo
Vía septohipocampal: estas fibras (Fig. 9) nacen del núcleo septal medio y del de la banda diagonal de Broca y está compuesto por axones positivos para la acetilcolinesterasa, además, contiene fibras gabaérgicas22. La proyección septohipocampal es responsable de la casi totalidad de la inervación colinérgica del hipocampo; hace arribo por medio del sistema fimbria-fornix, y es responsable de los fenómenos de actividad theta del hipocampo23.

Fig. 9. Esquema de las conexiones intralímbicas. GD: giro dentado; SUB: subiculum; LC: locus coeruleus; ATV: área tegmentaría ventral; SN: sustantia nigra.
La vía monoaminérgica talloencafálica (Fig. 9) corresponde a las proyecciones de los núcleos del rafe (5HT), locus coeruleus (norepinefrina), del locus Níger y otros sectores (dopamina), las cuales ejercen numerosos efectos sobre el umbral convulsivante del hipocampo, la norepinefrina por ejemplo, lo disminuye, las restantes, a la inversa, lo incrementan17.
Eferencias corticales del hipocampo
La principal eferencia es hacia la corteza entorrinal (Fig. 9); la cual funciona como un verdadero corredor entre el hipocampo y el resto de la corteza, ésta se interpone tanto en la ruta aferente como la eferente14,24, de aquí que se puede inferir la importancia de la corteza entorrinal en los procesos de almacenamiento mnésico, su lesión desvincula prácticamente a la formación del hipocampo del resto de la corteza. Es sabido, no obstante, que existen conexiones entre el hipocampo y la corteza perirrinal, así como cingular, orbitaria e infralímbica, aunque la importancia de estas conexiones no se encuentra bien dilucidada.
Eferencias subcorticales del hipocampo
La proyección mejor conocida del hipocampo es aquella que lo vincula con el área septal, de hecho CA1 y el subiculum envían fibras a través del fornix precomisural al núcleo septal lateral (Fig. 9). Otra proyección relacionada con ésta es la que se establece con el núcleo accumbens, por otro lado, el hipocampo se proyecta hacia los núcleos talámicos anteriores, amígdala e hipotálamo (núcleos mamilares, área preóptica). Otro aspecto de importancia es la participación del hipocampo en el eje hipotálamo-pituitario-adrenal, ya que posee grandes cantidades de receptores para gluco y mineralocorticoides8.
Características regionales
Fascia dentada. Las fibras que ingresan al hipocampo, tienen, en su mayoría destino en el giro dentado. Es así, que esta rudimentaria circunvolución constituye la compuerta de entrada a la mayor parte de las Incitaciones nerviosas que tienen por arribo al hipocampo (Fig. 7). El stratum granulosum, esta cubierto por el stratum moleculare, el que se subdMde en tres regiones, el tercio profundo recibe fibras comisurales y de asociación, el tercio medio la proyecciones entorrinales mediales y el tercio superficial, las entorrinales laterales. El estrato polimorfo, inmerso en el hilus, contiene el origen de las fibras musgosas (Fig. 7), así como varios tipos de células no granulares, incluidas las células musgosas excitatorias (glutamatérgicas), que dan origen a fibras de asociación/ comisurales, así como numerosas interneuronas inhibitorias. Las células granulares presentan algunas características. Son relativamente resistentes al insulto que genera la crisis epiléptica, tienen un alto umbral de descarga, poseen gran capacidad de reorganización de la red que sustentan, son principalmente excitatorias, y tienen la capacidad de regenerarse continuamente, con la subsecuente capacidad incrementada de crear circuitos aberrantes. Este continuo "turnover" dura toda la vida, y es dependiente de la concentración de esteroides suprarrenales25.
Las células granulares poseen potenciales de membrana en extremo altos, de manera que son difícilmente despolarizables. Una vez excitadas, generan importantes potenciales de hiperpolarización, esto es debido a la intensa actividad inhibitoria de las interneuronas circundantes. Los granos, envían axones a la región CA3, los cuales hacen sinapsis con células piramidales e interneuronas, a las que excitan. Las fibras musgosas contienen altas concentraciones de Zn, cuyo significado se desconoce.
Es interesante destacar que la célula granulosa no es la única neurona de proyección del giro dentado (Fig. 7). El otro tipo es la neurona musgosa hilar, la cual asienta en el estrato polimorfo. Siendo de carácter excitatorio, envía sus axones a la región profunda del stratun granulosum, activando células granulosas e interneuronas inhibitorias contra e ipsilaterales. Estas células tienen bajo umbral de excitabilidad, y resultan muy vulnerables a los procesos excitotóxicos. Es muy factible que esto último se halle ligado a las bajas concentraciones intracelulares de proteínas ligadoras de calcio. La vulnerabilidad exquisita que presentan estas neuronas puede que sea de extrema importancia para la epileptogénesis, ya que la depleción de ellas inactivaría una fuente de estimulación importante para las interneuronas inhibitorias, bajando así el umbral de descarga de las células granulosas17.
CA3: La región CA3 (Fig. 6) se caracteriza por la presencia de grandes células piramidales. Clásicamente se la suele dividir en tres subregiones; CA3a; en contacto con CA2; CA3b, formado la parte media de la banda en cuestión, localizada en las inmediaciones de la unión fimbria-fórnix; y CA3c, cerca del hilus, en continuidad con la región CA4. Por sus características, CA3a se la puede asimilar a CA2, de hecho, no recibe fibras musgosas17. Cada subsector posee propiedades independientes, pero analizada en conjunto, CA3 puede ser considerado como el marcapaso del hipocampo. De hecho, las neuronas de esta región tienen la tendencia a descargar en forma rítmica, involucrando a grandes poblaciones celulares. La frecuencia y amplitud de estos eventos bioeléctricos, parece justificar los hallazgos en EEG y particularmente los registros profundos interictales de los pacientes portadores de esclerosis mesiales temporales. La aparición de respuestas altamente sincronizadas parece deberse a las propiedades intrínsecas de las neuronas piramidales, así como la distribución de la red que configuran los circuitos locales26. Los axones de las grandes células piramidales emiten colaterales proximales de trayecto recurrente que se arborizan rápida y extensamente en las inmediaciones de los cuerpos neuronales, haciendo sinapsis con células piramidales y no piramidales. Estos circuitos piramidopiramidales generan por un lado, fenómenos de retroestimulación positiva, y por otro, como la pirámide que generó el axón no es la única que recibe sus colaterales, sino que estos se contactan con infinidad de otras células, la característica de esta red, es su tendencia a la sincronización, aspecto crítico de la epileptogénesis. No obstante, la mayoría de los potenciales excitatorios son contrarrestados por una fuerte actividad inhibitoria derivada de la acción de las interneuronas locales. Una mínima disrupción de esta última lleva a la generación de descargas rítmicas altamente sincronizadas17. La región CA3 ejerce su efecto en CA1 ipsilateral por medio de la vía colateral de Schaffer (Fig. 6 y 7), y en CA1 y CA3 contralaterales por medio de fibras comisurales17. Otro aspecto interesante de este sector es que los colaterales axónicos se extienden por grandes distancias interconectando células piramidales a lo largo de todo el eje hipocampal27, al punto de que una actividad epileptiforme que involucre gran parte del hipocampo rara vez se puede objetivar en la región de origen debido a la gran influencia inhibitoria de los circuitos locales, y es porque las interneuronas inhibitorias próximas a la célula de origen reciben un fuerte contingente de terminales. Las células piramidales gigantes tienen una gran densidad de canales de calcio en su árbol dendrítico, lo cual las hace altamente excitables. Esto último, ligado a las características de la red neural, hace que una vez producida la descarga, se propague hacia CA1 y hacia el resto de CA3, así como al hipocampo contralateral. Presumiblemente, el mecanismo gatillo de estas descargas, sea el "input" masivo proveniente del giro dentado, puntualmente, las influencias de las células granulares a través de las fibras musgosas. Las fibras musgosas, establecen complejas sinapsis con las grandes células piramidales a nivel del stratum lucidura, donde son capaces de generar EPSP mediados por receptores no NMDA28.
Dos aspectos resultan de interés, uno de ellos es que la frecuencia de descarga de las fibras musgosas es baja, el otro es que estos "Inputs" también actúan sobre las redes inhibitorias locales, de esta manera, la actividad ictal se torna poco probable en condiciones locales, ya que las pirámides se encuentran tónicamente inhibidas por su entorno neuronal. Existen diversos tipos de interneuronas inhibitorias, algunas llamadas "fast spiking cells"; que producen cortos IPSP, y las conocidas como "slowspiking cells", capaces de general largos IPSP en las células piramidales.
El árbol dendrítico se encuentra sectorizado; el sector proximal, formado por los troncos de menor orden de las dendritas apicales, a nivel del stratum lucidum, recibe sinapsis de las fibras musgosas, el sector medio, formado por los segmentos intermedios de las dendritas apicales (stratum radiatum) y por las dendritas básales (stratum oriens), recibe fibras comisurales y de asociación (fibras A/C), mientras que el sector distal, conformado por las últimas ramificaciones de las dendritas apicales (stratum lacunosum/moleculare) recibe las fibras del haz temporoamónico (Figs. 6 y 7), provenientes de la corteza entorrinal17. De todas formas, todo este sistema está modulado por aferencias subcorticales17. Un hecho relevante es que las sinapsis musgosas, al actuar por un mecanismo dependiente de receptores no NMDA, presentan un patrón de plasticidad diferente al LTP ("long term potentiation")29, que se puede apreciar a nivel del stratum radiatum. Este mecanismo de potenciación parece dependiente del incremento de la concentración intracelular de cAMP derivado de la actividad del sistema monoaminérgico16.
CA1._Las células piramidales de CA1, junto con las del subiculum, constituyen los eferentes primarios del hipocampo. Los aferentes excitatorios principales de este sector son los colaterales de Scaffer, ipsi y contralaterales (Figs. 6 y 7), los cuales vierten contactos sinápticos con dendritas básales del stratum oriens y apicales del stratum radiatum17, otro contingente aferente excitatorio de importancia son las fibras que conforman la vía temporoamónica, que proviene del córtex entorrinal, éstas hacen sinapsis con las dendritas apicales en el stratum lacunosum/moleculare30. Si bien las pirámides de CA1 están sometidas a un entorno inhibitorio, este no es tan fuerte como el del giro dentado o de CA3, por lo cual son más fácilmente reclutables que las neuronas piramidales de otros sectores, además presentan mayor labilidad al estímulo excitotóxico que otras. Un aspecto para recalcar, es que las interacciones del las células piramidales de CA1 entre sí, son escasas, por lo cual se puede admitir que los patrones de sincronización no dependen de una actividad intrínseca, sino de las propiedades de los circuitos de CA3, si bien este sector no tiene tendencia a generar descargas masivas en sus poblaciones neuronales, estas pueden ser rápidamente involucradas por la acción de sus aferencias excitatorias. El entorno inhibitorio está compuesto por una población heterogénea de neuronas gabaérgicas con sensibilidad variable a la acetilcolina31 y a la norepinefrina32, estas células han recibido gran atención, por su posible papel en la generación de ritmo theta. Muchos estudios se han centrado en los mecanismos de plasticidad sináptica inherente a este sector. Los EPSP de las células piramidales dependen de mecanismos mediados por receptores NMDA y no NMDA. Los receptores NMDA resultan cruciales para la iniciación de los fenómenos de LTP, pero ante intensos niveles de estimulación excitatoria, las corrientes de Ca++ pueden generar formas de LTP33. El concepto de neurona desconectada o dormida originado para explicar los fenómenos de epileptogénesis en el giro dentado, parece apropiado para CA134, ya que se han registrado disminución de los IPSP GABA dependientes, encontrándose preservadas las neuronas gabaérgicas, incrementándose así la excitabilidad de las células piramidales.
Subiculum. El subiculum es la mayor fuente de eferencias de la formación del hipocampo (Fig. 6 y 7) está interpuesto entre el hipocampo mismo y la corteza14,17,24 por un lado, así como los centros subcorticales y el hipocampo por otro17, de esta manera, constituye un punto clave para la difusión de las crisis de origen hípocampal. Dado que en la ETM, el subiculum muestra escasa depleción celular respecto de otras regiones 17,35,36, cosa por completo opuesta a lo que ocurre en CA1, este sector cobra aun mayor trascendencia. Las neuronas piramidales del subiculum presentan EPSP dependientes de receptores glutamatérgicos NMDA y no NMDA, así como IPSP GABA-dependientes. La proyección cortical fundamental es hacia el área entorrinal14,17,24
Corteza entorrinal. La corteza entorrional está interpuesta funcionalmente entre el hipocampo y el resto de la corteza14,24. Grandes porciones de la neocorteza se proyectan el área entorrinal y la adyacente corteza perirrinal17 (Fig. 9), es así que esta región recibe un importante contingente de fibras provenientes de la corteza orbitofrontal, insular, infralímbica, singular y temporobasal. Por otro lado, el subiculum se proyecta extensamente hacia la corteza entorrinal. Las neuronas del córtex entorrinal poseen la capacidad de descargar en forma masiva37, de esta manera, el área podría actuar como un verdadero amplificador en la difusión de las crisis epilépticas hipocampales17.
Los "inputs" subcorticales son muy numerosos, los bulbos olfatorios inervan los estratos superficiales, la corteza prepiriforme hace lo propio, al punto que al área entorrinal se la considera la corteza olfatoria secundaria15, 16, el núcleo septal medial envía gran cantidad de fibras colinérgicas17, la amígdala envía proyecciones tanto hacia la EC medial como lateral38, del tallo llegan fibras provenientes de los núcleos del rafe, del locus coeruleus, etc. El tálamo envía fibras desde el nucleus reuniens y los núcleos anteriores. Las eferencias son recíprocas, pero mientras que las aferencias arriban a las capas I y III, las eferencias se originan en los estratos IV y VI17. Extensas redes existen entre las diferentes capas de la corteza entorrinal, lo cual tiene importantes implicancias en la generación y difusión de las crisis. La capa II da origen a la vía perforante, que se dirige al giro dentado, y la capa III destina fibras al sector CA1. Estos dos estratos son los que aportan la casi totalidad de los inputs corticales del hipocampo.
La mayor parte de las neuronas del área entorrinal son de configuración triangular. La vía perforante se origina en las células estrelladas de la capa II y las piramidales de la capa III, se impregnan intensamente con las tinciones para metales pesados de Timm, y perece utilizar glutamato corno neurotransmisor38. Los preparados de corteza entorrinal e hipocampo, sometidos a bajas concentraciones de Mg+ o altas de K+, generan un patrón de descarga muy similar al que se puede objetivar durante los fenómenos ictales17. Es aquí donde podría jugar un papel importante las modificaciones del medio intercelular que general los procesos de gliosis.
Corteza perirrinal._Los índices más rápidos de epileptogénesis por "kindling" se han obtenido por estimulación de la corteza perrinal17; esto ha generado gran interés en el estudio de esta región. Extendida dorsalmente desde el margen posterior del área entorrinal a la corteza insular, este sector se encuentra ricamente inervado por los lóbulos frontal, parietal, occipital, insular y temporal, incluida la corteza piriforme. Las aferencias subcorticales involucran al tálamo, claustrum, amígdala, núcleos del rafe y al núcleo endopiriforme. Las conexiones, son en su totalidad, recíprocas, de extrema importancia resulta el importante contingente que se extiende al área motora primaria. Desde un punto de vista histoarquitectónico, la corteza perirrinal es del tipo agramilar, contiene una población heterogénea de neuronas, cuya excitabilidad parece corresponder a la de la corteza entorrinal18. Interesantemente, las células de la lámina V que se proyectan a la corteza motora frontal, presentan tendencia a las descargas masivas cuando son estimuladas por corrientes despolarizantes40. La exéresis de la corteza perirrinal previene el desarrollo de crisis clónicas en los experimentos de "kindling" del hipocampo dorsal41.
Corteza piriforme. La piriforme es el área olfatoria primaria, de diminuto volumen en el ser humano, en las especies macrosomáticas adquiere gran desarrollo. Desde el punto de vista citoarquitectónico, es una corteza trilaminar; configurada por una lámina I llamada plexiforme, una lámina II o de células piramidales y, por fin, una lámina III o de células polimorfas. La lámina I esta compuesta por el árbol dendrítico de las células profundas y las ramificaciones terminales de las múltiples aferentes, se subdivide en un sector la y otro Ib. La lámina II contiene células piramidales y semilunares y la III células piramidales en su región superficial y no piramidales en su estrato profundo.
Las fibras olfatorias primarias terminan en el estrato la, las del núcleo olfatorio anterior en el estrato Ib, el núcleo de la estría olfatoria lateral en el estrato II, el cortical de la amígdala en el III y la corteza insular en las tres capas. Profundamente alojado a la corteza piriforme se encuentra el núcleo endopiriforme, esta estructura provee una vasta inervación a la capa Ib, cabe acotar que el núcleo endopiriforme es una formación altamente excitable17.
El locus coeruleous, el área tegmentaria ventral, la sustantia nigra, la sustantia innominata, el hipotálamo y el tálamo, constituyen importantes aferentes subcorticales
La importancia del córtex piriforme en epileptología se ha puesto en evidencia por la rápida respuesta en los modelos de "kindling", su sensibilidad exquisita a sustancias convulsivantes y su extrema labilidad a la injuria ictal. Sus conexiones entre vastas áreas neocorticales y las cortezas ento y perirrinales, su organización convergente y divergente dan cuenta de su importante papel en la generación y difusión de fenómenos críticos.
Amígdala. El complejo nuclear amigdalino, constituye un aglomerado de aproximadamente 10 núcleos alojados en el extremo dorsorrostral del cuerno temporal, tiene una configuración ovoidea y aproximadamente dos centímetros de longitud. Esta estructura existe en todos los mamíferos, y ha sido homologado al arquiestriado de las especies no mamíferas. Clásicamente, sus diferentes núcleos son agrupados en una división corticomedial y una basolateral, el núcleo central se lo suele considerar aparte, pero en ocasiones se lo incluye dentro de la división corticomedial. En el ser humano, el grupo corticomedial, se aloja en posición dorsal o dorsomedial del complejo, por la rotación medial del lóbulo temporal. Las subdivisiones de este grupo son: el área amigdalina anterior, el núcleo de la cintilla olfatoria lateral, el núcleo medial, y el cortical. El núcleo de la cintilla olfatoria lateral es el menos desarrollado en el hombre. El área amigdalina anterior, la parte más rostral del complejo, esta poco diferenciada Al conjunto formado por el área amigdalina anterior, el núcleo de la estría lateral y el núcleo cortical se lo se los denominan grupo olfatorio. El grupo basolateral es la parte más desarrollada en el hombre; sus subdivisiones son; el núcleo amigdalino lateral, el basal y el basal accesorio.
Los aferentes del grupo olfatorio son los bulbos olfatorios y la corteza piriforme. El hipocampo se proyecta también, pero lo hace mayormente a través del subículum y CA1. Otras aferencias corticales están formadas por la corteza entorrinal lateral y la insular agranular. Las subcorticales provienen del hipotálamo, tálamo, núcleos del rafe, locus coeruleus, núcleo de la banda diagonal y núcleo endopiriforme17 (Fig. 9). Los eferentes son mayormente recíprocos, aunque un componente importante está representado por las fibras que se extienden el estriado ventral y el núcleo accumbens. A su vez, este grupo se interconecta con los demás núcleos del complejo, a excepción del núcleo lateral.
El núcleo medial recibe fibras de la corteza piriforme, entorrinal, retroesplenial, insular agranular, del subiculum y del hipocampo. Los aferentes subcorticales corresponden a los núcleos talámicos de línea media, hipotálamo, así como de los otros componentes nombrados para el grupo olfatorio. Los eferentes del núcleo medial que tienen importancia para el conocimiento de la fisiopatología de la ETM son las proyecciones a la corteza entorrinal y el estriado ventral. El hipotálamo y el tallo reciben fibras de este núcleo. El núcleo medial expresa CCK, somatostatina, hormona liberadora de tirotrofina, VIP, encefalina, y otros, pero el GABA es el mejor caracterizado de todos los neurotransmisores, esto puede dar cuenta de la relativa resistencia al kindling de este sector del complejo. El núcleo central, por el contrario, es el más vulnerable al proceso de kindling, recibe aferencias de las cortezas, piriforme, insular agranular, entorrinal, perirrinal, etc. Tiene varios inputs del hipotálamo y tálamo, núcleos del rafe y locus coeruleus. Las eferencias involucran a la corteza perirrinal, el estriado, el hipotálamo, la sustancia gris periacueductal, los núcleos pedúnculopontinos, parabraquiales, el NTS, el motor dorsal de vago, etc. Todas estas proyecciones subcorticales proveen importantes vías de expresión visceral de la actividad del núcleo, de aquí que se puede entender su importancia en los aspectos fenomenológicos de los procesos ictales de etiología temporal mesial.
El grupo basolateral se distingue por sus vastas conexiones con el neocórtex, además de aquellas que presenta con la corteza entorrinal, perirrinal, piriforme, insular agranular, etc. Las conexiones subcorticales de importancia corresponden al núcleo de la banda diagonal, el pálido ventral y la sustancia innominada. Otros aferentes son el tálamo, hipotálamo, así como las proyecciones noradrenérgicas, seritoninérgicas y dopaminérgicas del tallo cerebral17 (Fig. 9).
Según diversos estudios experimentales17 la aparición de descargas epileptiformes en la amígdala basolateral involucra a fenómenos mediados por receptores NMDA y no NMDA; así como la disminución de la actividad GABA.
De esta manera, se puede concluir que los fenómenos de epileptogénesis en el ETM obedecen a alteraciones en los mecanismos de inhibición mayormente gabaérgicos en el giro dentado, verdadera compuerta de entrada a la formación del hipocampo, a la exquisita tendencia a la sincronización que tiene la región CA3, en función de la extensa red de colaterales axónicos recurrentes, al importante poder de amplificación que posee la región CA1 y a las características de las conexiones entre el subiculum y la corteza entorrinal.
Los aspectos fenomenológicos de las crisis temporales son marcadamente atinentes a las características de las conexiones de la corteza entorrinal, la amígdala, el mismo hipocampo con el hipotálamo, el estriado, el neocórtex, etc (Fig. 9).
No resultan claras aún las particularidades inherentes a la difusión de las crisis fuera del hipocampo ante la presencia de una región CA1 despoblada, que constituye la principal estación de relevo entre el resto del hipocampo y el subiculum.
Un aspecto interesante es que cuando una actividad eléctrica anormal queda limitada a menos de 1000 neuronas, esta no se manifiesta clínicamente14. Durante el período interictal la actividad del foco se mantiene confinada a una región pequeña de la corteza, esto se debe a que este grupo de neuronas se halla inmerso en un entorno inhibitorio determinado por IPSP generados por neuronas mayormente gabaérgicas. Durante el desarrollo de la crisis, los fenómenos de posthiperpolarización, dependientes de la acción de circuitos locales desaparecen, de esta manera es que grupos más extensos de neuronas son reclutados; las causas de este fenómeno de ruptura del entorno inhibitorio no se saben. Al parecer, la transmisión gabaérgica es algo lábil, aunque de ser así, se desconoce si los mecanismos íntimos del proceso son de extirpe pre o postsináptica. Muchos aspectos inherentes a la naturaleza de los procesos subyacentes a la generación y difusión de las crisis epilépticas resultan oscuros a los actuales conocimientos y requieren de futuras investigaciones para ser resueltos.
CUADRO CLÍNICO DE LAS CRISIS EPILÉPTICAS ASOCIADAS A LA ESCLEROSIS TEMPORAL MESIAL
Al analizar clínicamente las crisis ocasionadas por la EMT debemos tener en cuenta las estructuras involucradas en la génesis de las mismas, siendo la formación hipocampal y las estructuras adyacentes: amígdala, ínsula, ganglios básales y cíngulo.
Al interrogatorio pueden encontrarse antecedentes de presentación de crisis febriles complejas (crisis tónico-clónico generalizadas con predominio en un hemicuerpo), crisis de larga duración hasta 10 minutos y/o presencia de estado de mal epiléptico. Suelen presentarse antes de los 4 años y luego de intervalo libre de ataques hasta 10 años, comienzan con crisis. Aún se discute si estos son causa o consecuencia de la EMT. Se describen padecimiento de traumas, agresión perinatal o malformaciones del hipocampo2,3.
Las crisis típicamente se presentan como parciales complejas. Esta semiología es característica, aunque no patognomónica, de las crisis temporales. Aproximadamente un 80% de las crisis parciales complejas provienen de estructuras temporales y de estas hasta un 90% son de semiología mesial temporal. La EMT es un síndrome epiléptico, en ello se incluyen crisis que provienen de estructuras mesiales o básales o de la neocorteza del lóbulo temporal2,3.
Estas crisis pueden estar precedidas en ocasiones por un aura típica, referida como "sensación que asciende" retroesternal hasta el cuello, en los niños puede dar sensación de cólico abdominal o necesidad de defecar (epilepsia abdominal). Este tipo de aura pertenece a la irritación insular. En caso de comprometerse la amígdala y giro parahipocámpico, sentimientos de despersonalización, miedo y pánico pueden evidenciarse. El aura puede presentarse en forma aislada o tornarse en crisis parcial compleja luego de 5 a 30 segundos. En caso de progresar hacia la crisis hay ausencia de respuesta al medio, mirada fija, conducta automática (simple o compleja) como masticación, movimientos de lengua, y otros más complejos como refregarse las manos, tocarse la ropa, desnudarse, caminar, frotarse la nariz con la mano ipsilateral al foco epiléptico, etc. Los eventos clínicos que siguen a este cuadro inicial dependen de las estructuras involucradas en la propagación de las descargas. En caso de involucrar al cíngulo se manifestará como movimiento de pedaleo. La postura distónica de la mano contralateral al foco que descarga indica participación de ganglios de la base ipsilaterales. Puede presentarse un lenguaje inteligible (discurso ictal) donde se sospecha participación del lóbulo temporal no dominante. Un cuadro de disfasla postictal sugiere descarga del lóbulo dominante. Si en su propagación se involucran estructuras suprasilvianas alcanzando al lóbulo frontal, el paciente presentará desviación cefálica y ocular forzada contralateral al foco que descarga. En resumen, una crisis que comienza con aura gástrica, automatismos, postura distónica y desviación cefálica forzada presuponen origen en el lóbulo temporal, contrariamente si de inicio muestra desviación cefálica presupone crisis de inicio frontal.
Posteriormente puede concluir con crisis tónica o tónico clónica generalizada. Sobreviene confusión post ictal (postcrítica) que dura minutos, puede estar afásico por lo menos unos 30 minutos si la afectación primaria es el temporal dominante izquierdo. A veces predominan los síntomas psiquiátricos postictales como ansiedad irritabilidad, depresión inmediatamente posterior o en el período que abarca hasta 72 horas postcrisis2,3.
Comorbilidad psiquiátrica en la epilepsia del lóbulo temporal. Hay alteraciones neuropsiquiátricas y déficit neurocognitivo cuando hay afectación del lóbulo temporal: memoria y dificultad para la aplicación de pruebas ofrecidas por medios verbales (lóbulo dominante) y los test ofrecidos por vía visual (lóbulo no dominante).
Otros síntomas como ansiedad y depresión son inherentes a afectación de estructuras mesiales temporales y otras no límbicas.
Todavía se halla en discusión la relación del riesgo de presentación en estos pacientes de la denominada "personalidad del lóbulo temporal": que no es específico de las crisis del lóbulo temporal y se encuentra en otras patologías psiquiátricas y neurológicas.
Estos pacientes se presentan además con alteración en la esfera sexual como lo son la disminución de la libido, impotencia, anorgasmia, eyaculación retardada y dispareunia en las mujeres que al parecer se halla vinculada al consume de las DAE3.
EEG EN LAS EPILEPSIAS DEL LÓBULO TEMPORAL
El hallazgo más consistente en las epilepsia del Lóbulo Temporal (ELT) es la espiga temporal anterior. No obstante esto, las epilepsias del lóbulo temporal pueden generar alteraciones electroencefalográficas en otras localizaciones, además las espigas temporales anteriores no son patognomónicas de ELT, ya que se pueden ver en las epilepsias del lóbulo frontal, especialmente en las epilepsias fronto-orbitarias. La unilateralidad de los hallazgos en registros repetidos confiere alta probabilidad de que la crisis se origine en lóbulo involucrado electroencefalográficamente y confiere un buen pronóstico3.
El patrón ictal de EEG de "scalp" (Fig. 10) varía según la localización del generador dentro del lóbulo. En el 50 al 80% de los casos de las epilepsias de origen temporal mesial, la actividad es rítmica, habitualmente en el rango theta o alfa, que aumenta en amplitud y disminuye en frecuencia42. Esta actividad que se obtiene en superficie no corresponde al registro del hipocampo, sino a la propagación de la crisis hacia el cortex temporo-basal, ya que los generadores profundos no suelen tener expresión directa en el EEG de superficie. Los ritmos alfa, delta o theta rítmicos son los predominantes51 . La actividad ictal generalmente se propaga al resto del lóbulo, y después al contralateral.

Fig. 10. EEG de SCALP que muestra el inicio de una crisis temporal.
La lentificación post-ictal es un hecho de valor lateralizador en la epilepsia de origen temporal mesial3.
El registro con electrodos profundos (Fig. 11) se indica cuando las dudas de lateralidad persisten. En caso de sospechar un componente neocortical, una opci ón apropiada puede ser la colocación de electrodos ortogonales, ya que registran estructuras mesiales y laterales. Las crisis hipocampales normalmente comienzan con salvas de espigas seguidos por actividad rítmica alfa o beta de baja o gran amplitud43. La epilepsia del lóbulo temporal mesial comienza con una actividad de 10 a 16 Hz. mientras que las epilepsias neocorticales combinan actividad de baja con alta frecuencia44. Spencer sugiere que la actividad de baja amplitud y alta frecuencia (más de 13 Hz.) suele corresponder a ETM, mientras que las de baja frecuencia y alta amplitud al comienzo de la crisis suelen tener mayor correlación con hallazgos histopatológicos normales. Las crisis temporales mesiales pueden tener comienzo regional dentro del hipocampo o amígdala o extenderse por todo el eje de estas estructuras4. Es mas frecuente que las crisis comiencen en la porción anterior del hipocampo que el la posterior; esto se suele asociar a mejor pronóstico43, los registros intracraneales pueden ser utilizados para evaluar el patrón de propagación de las crisis, éstas se pueden difundir a estructuras ipsilaterales como la corteza temporal ipsilateral u órbito-frontal, o al hipocampo contralateral por media de fibras A/C a través de la comisura hipocampal, antes de hacerlo a estructuras ipsilaterales, esto tiene valor pronostico, a mayores tiempos de propagación contralateral, mejores resultados postquirúrgicos42.

Fig 11: Registro del inicio de una crisis temporal mesial con electrodos profundos implantados en hipocampo.
EVALUACIÓN NEUROPSICOLÓGICA
La evaluación neuropsicológica constituye una herramienta fundamental en la evaluación del paciente epiléptico candidato a cirugía. Debido al deterioro de las funciones cognitivas presente en estos pacientes, es necesario evaluaciones pre y postquirúrgicas.
El deterioro cognitivo particular de cada paciente está relacionado con distintas variables: etiología, uso de fármacos antiepilépticos, edad de inicio de la enfermedad, tipo de crisis, duración y frecuencia de las crisis epilépticas3,4,45. En cuanto a la etiología, se propuso en un principio que las epilepsias sintomáticas cursaban con mayor deterioro que las idiopáticas. Esto podría deberse al compromiso de la lesión en sí misma aunque no hay consenso al respecto; algunas epilepsias consideradas idiopáticas con leve compromiso cognitivo resultaron ser sintomáticas con el advenimiento de métodos de diagnóstico más sensibles4,46. Los estudios que relacionan este deterioro al uso de fármacos antiepüépticos revisten dificultades metodológicas propias debido a la gran cantidad de variables que entran en juego al evaluar esta asociación4,46,47. No obstante, el grado de compromiso documentado no llega a presentar significación estadística. De todas las funciones cognitivas analizadas, la que presentaría mayor compromiso es la atención, siendo la alteración de la memoria una consecuencia indirecta. De los fármacos utilizados, el que impresiona presentar mayor compromiso a altas dosis es el fenobarbital, afectando a las capacidades visoespaciales4. Respecto a la edad de inicio de la enfermedad, existe relación inversamente proporcional entre ésta y el deterioro cognitivo; cuanto más temprano sea el inicio de la enfermedad, mayor es el potencial de deterioro4,4,50. Otra variable analizada es el tipo de crisis: las crisis generalizadas presentan mayor deterioro que las parciales. Una excepción son las crisis generalizadas de ausencias no atípicas, en las cuales el efecto deletéreo es comparable con el producido por las parciales. Aquellos que presentan más de un tipo de crisis tienen mayor riesgo de deterioro respecto a los que experimentan un solo tipo. El status epilepticus es el tipo de crisis que mas deterioro produce4,45,50. Por último, al analizar la duración y frecuencia de las crisis epilépticas se considera que la relación del deterioro es directamente proporcional a estas variables4,50.
Los objetivos de la evaluación neuropsicológica son: obtener una línea de base de cada paciente en particular, mediante el estudio y la valoración de las funciones superiores, permitiendo una evaluación evolutiva de los resultados de la cirugía, y estudiar la locáiización del complejo lesivo epileptógeno y la laterálizacián hemisférica, dos ítems cruciales y en ocasiones de complejo diagnóstico. Respecto a la lateralización, es fundamental la identificación prequirúrgica del hemisferio dominante para el lenguaje y del compromiso amnésico asociado. Resulta ser clave en ocasiones la realización del test de Wada con esta finalidad4,45.
Si bien no existe consenso, algunos autores afirman que puede ser utilizada como elemento de predicción prequirúrgico sobre la respuesta de las crisis y cambios en la esfera cognitiva luego de la cirugía45,51,53.
Desde el punto de vista quirúrgico, esta evaluación nos aporta información que va a influir en parte a la hora de planear la táctica y técnica quirúrgica, en función del grado de compromiso previo de las estructuras involucradas.
Por último, permite brindarle a cada paciente en particular, información con mayor certeza de los potenciales riesgos y beneficios que brinda el procedimiento a realizarse45,55.
1. Pomata HB, Campos M. Epilepsy in pre-colombian times, (en prensa- Child's Nerv Syst). [ Links ]
2. Micheli F, Nogués MA, Asconapé JJ, Fernández Pardal MM, Biller J. Tratado de Neurología Clínica. 1ra ed. Buenos Aires (Arg.): Ed. Médica Panamericana; 2002. [ Links ]
3. Campos MG, Kanner AM. Epilepsias: diagnóstico y tratamiento. 1ra ed. Santiago (Chile): Ed. Mediterráneo; 2004. [ Links ]
4. Villarejo Ortega F. El Tratamiento de
5. Rhoton A. The cerebrum. Neurosurgery 51 [ suppl 1] 1-51, 2002. [ Links ]
6. Rhoton A. The supratentorial arteries. Neurosurgery 2002; 51 [ suppl 1] 53-120. [ Links ]
7. Roper S, Rhoton A. Surgical Anatomy of the temporal Lobe. Neurosurg Clin N Am 1993; 4: 223-31. [ Links ]
8. Rubino P, Rhoton A, Tong X, et al. Three-dimensional relationships of The optic radiation. Neurosurgery 2005; 57 [ONS Suppl 3]: ONS- 219-ONS-27. [ Links ]
9. Wen H, Rhoton A, de Oliveira E, et al. Microsurgical anatomy of the temporal lobe: Part 1. Mesial temporal lobe anatomy and its vascular relationships as applied to amigadlohippocampectomy. Neurosurgery 1999; 45: 549-92. [ Links ]
10. Yasargil M, Wieser H, Valavanis A et al. Surgery and results of selective Hippocampectomy I one hundred patients with nonlesional limbic epilepsy. Neurosurg Clin N Am 1993; 4:243-61. [ Links ]
11. Broca P. Anatomie comparée des circonvolutions cerebrales. Le grand lobe limbique et la scissure limbique dans le serie des mammiferes. Rev Anthropol Ser 1878; 2: 385-498. [ Links ]
12. Papez JW. A proposed mechanism of emotion. Arch Neurol Psychiat 1937; 38: 725-43. [ Links ]
13. Maclean PD. Some psychiatric implications of physiological studies on frontotemporal portion of limbic system (visceral brain). EEG Clin Neurophysiol 1952; 4: 407-18. [ Links ]
14. Kandel E. R. Schwartz J. H. Jessell T. M. Principies of Neural Science. Mc Graw Hill. Fourth Edition. U. S. A. 2000. [ Links ]
15. Carpenter M. Sutin J. Neuroanatomía Humana. VI Ed. El Ateneo. Argentina. 1985 [ Links ]
16. Carpenter M. Neuroanatomía-Fundamentos. Ed. Médica Panamericana. Barcelona. 1994. [ Links ]
17. Engel J Jr, Pedley TA. Epilepsy: The Comprehensive CD-ROM. © 1999. Lippincott Williams & Wilkins. [ Links ]
18. Henry Rouviére; Andre Delmas. Anatomía Humana. 11° Ed. Masson. Barcelona. 2005. [ Links ]
19. Ranson S. W. Clark S. L. Anatomía Del Sistema Nervioso. X Ed. Editorial Interamericana. México. 1964 [ Links ]
20. O'KeefeJ, NadelL. The Hippocampus as a Cognitive Map Oxford University Press. 1978 [ Links ]
21. Romo-Parra H, Vivar C, Maqueda J, Morales MA, Gutiérrez. Activity- Dependent Induction of Multitransmitter Signaling Onto Pyramidal Cells and Interneurons of Hippocampal Área CA3. J Neurophysiol 2003; 89: 3155-67. First published February 12, 2003; 10.1152/jn.00985.2002 [ Links ]
22. Amaral DG, Kurz J. An analysis of the origins of the cholinergic and noncholinergic septal projections to the hippocampal formation of the rat. J Comp Neurol 1985; 240: 37-59. [ Links ]
23. Bradley P., Wyble, Linster C, Hasselmo ME. Size of CA1-Evoked Synaptic Potentials Is Related to Theta Rhythm Phase in Rat Hippocampus. The American Physiological Society. 2000 [ Links ]
24. Milner B, Squire LR, Kandel ER. Cognitive Neuroscience and the Study of Memory. Neuron, 1998; 20: 445-68. [ Links ]
25. Gould E, Cameron HA, Daniels DC, Woolley CS, McEwen BS. Adrenal hormones suppress cell división in the adult rat dentate gyrus. J Neurosci 1992; 12: 3642-50. [ Links ]
26. Traub RD, Miles R. Neurona! Networks of the Hippocampus. Cambridge, UK: Cambridge University Press, 1991 [ Links ]
27. Li X-G, Somogyi P, Ylinen A, Buzsáki G. The hippocampal CA3 network: an in vivo intracellular labeling study. J Comp Neurol 1994; 339: 181-208. [ Links ]
28. Brown TH, Johnston D. Voltage-clamp analysis of mossy fiber synaptic input to hippocampal neurons. J Neurophysiol 1983:50: 487-507. [ Links ]
29. Weisskopf MG, Zalutsky RA, Nicoll RA. The opioid peptide dynorphin mediates heterosynaptic depression of hippocampal mossy fibre synapses and modulates long-term potentiation. Nature 1993; 362: 423-27. [ Links ]
30. Soltesz L, Jones RSG. The direct perforant path input to CA1: excitatory or inhibitory? Hippocampus 1995; 5:101-3. [ Links ]
31. Reece LJ, Schwartzkroin PA. Effects of cholinergic agonists on two non-pyramidal cell types in rat hippocampal slices. Brain Res 1991; 566: 115-26. [ Links ]
32. Doze VA, Cohén GA, Madison DV. Synaptic localization of adrenergic disinhibition in the rat hippocampus. Neuron 1991; 6: 889-900. [ Links ]
33. Grover LM, TeylerTJ. N-methyl-d-aspartate receptor-independent long-term potentiation in área CA1 of rat hippocampus: input specific induction and preclusion in a non-tetanized pathway. Neuroscience 1992; 49: 7-11. [ Links ]
34. Bekenstein JW, Lothman EW. Dormancy of inhibitory interneurons in a model of temporal lobe epilepsy. Science 1993:259:97-100. [ Links ]
35. Campos M; Kaner A. Epilepsias. Diagnóstico y Tratamiento. Ed. Mediterráneo. Chile. 2004. [ Links ]
36. Lüders H; Comair Y. Epilepsy Surgery. 2a Edición. Lippincott Williams & Wilkins. U.S.A. 2000 [ Links ]
37. Stewart M, Wong RKS. Intrinsic properties and evoked responses of guinea pig subicular neurons in vitro.J Neurophysiol 1993; 70: 232-45. [ Links ]
38. Beckstead RM. Afferent connections of the entorhinal cortical área in the rat as demonstrated by retrograde labeling with horseradish peroxidase. Brain Res 1978; 152: 249-64. [ Links ]
39. Storm-Mathisen J. Glutamate in hippocampal pathways. In: Di-Chara G, Gessa GL, eds. Glutamate as a Neurotransmitter. New York: Raven Press, 1981; 43-55. [ Links ]
40. Mclntyre DC, Kelly ME, Staines WA. Efferent projections of the anterior perirhinal cortex in the rat. J Comp Neurol 1996; 369: 302-18. [ Links ]
41. Kelly ME. An investigation into temporal lobe seizure networks: role of the piriform and perirhinal cortices in hippocampal kindled motor seizures. [ PhD dissertation] Ottawa: Carleton University, 1995. [ Links ]
43. Lüders H; Comair Y. Epilepsy Surgery. 2a Edición. Lippincott Williams & Wilkins. U.S.A. 2000 [ Links ]
44. Spencer S; Guimaraes P; Katz. Morphological pattems of seizures recorded intracranially. Epylepsia 1992; 33: 537-45 [ Links ]
45. Wyler AR, Hermann BP. The Surgical Management of Epilepsy. 1ra ed. USA: Ed. Butterworth-Heinemann; 1994 [ Links ]
46. Dodrill CB. Interictal cognitiva aspects of epilepsy. Epilepsia 1992a; 33 (Suppl. 6): S7-S10. [ Links ]
47. Dodrill CB. Neuropsychological aspects of epilepsy. Psych Clin North Am 1992b: 15(2): 383-94. [ Links ]
48. Tañer RE. Intelectual and adaptlve functioning in epilepsy: a review of fifty years of research. Diseseases of the nervous system 1972: 33: 763-9. [ Links ]
49. Perrine K. Gershengorn J, Brown ER. Interictal neuropsychological function in epilepsy. En: Devinsky O, Theodore WH (eds.). Epilepsy and behavior. New York: Wiley-Liss, 1991: 181-193. [ Links ]
50. Dodrill CB. Corrwelates of generalized tonic-clonic seizures with intellectual, neuropsychologocal, emotional and social function in patients with epilepsy. Epilepsia 1986; 27: 399-411. [ Links ]
51. Smitj DB. Kraff BR, Collins J et al. Behavioral characteristics of epilepsy patients compared with nirmal controls. Epilepsia 1986: 27: 760-8. [ Links ]
52. Wannamaker BB, Matthews CG. Prognostic implications of neuropsychological test performance for surgical treatment of epilepsy. J Nerv Ment Dis 1976; 163: 29-34. [ Links ]
53. Lieb JP, Rausch R, Engel J Jr, Brown WJ, Crandall PH. Changes in intelligence following temporal lobectomy: relationship to EEG activity, seizure relief, and pathology. Epilepsia 1982; 23: 1-13. [ Links ]
54. Rausch R. Crandall PH. Psychological status related to surgical control of temporal lobe seizures. Epilepsia 1982; 23: 191-202. [ Links ]
55. Ivnik RJ, Sharbrough FE, Laws ER. Anterior temporal lobectomy for control of partial complex seizures: information of counseling patients. Mayo Clin Proc 1988; 63: 783-91. [ Links ]

