










Servicios Personalizados
Revista

Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkRevista argentina de neurocirugía
versión On-line ISSN 1850-1532
Rev. argent. neurocir. vol.24 supl.1 Ciudad Autónoma de Buenos Aires ago. 2010
Resumen del libro de "Teoría de la localización comparativa de la corteza cerebral" expuesta en sus principios en base a la estructura celular1
Horacio Fontana

En el prólogo de su libro, Brodmann expone el objetivo de su trabajo, incluido en el ámbito del programa de investigación del laboratorio neurobiológico de Berlín, dirigido por Oskar Vogt. La tarea que se le encomendó, fue la de realizar un análisis topográfico de la corteza cerebral del ser humano según su estructura celular, para lograr una división topográfico-localizatoria detallada de la misma, que tuviera tanto aplicación clínica, como también ofreciera una imagen de la estructura histológica normal, para su aplicación en la patología.
Pronto observó la necesidad de que su trabajo estableciera las leyes de formación de la corteza cerebral y los fundamentos de la sorprendente variedad de las estructuras encontradas, a través de un estudio más amplio que le proporcionara bases ontogenéticas y de la anatomía comparada. Así su tarea se complicó, pues ya no se trataba más de una investigación sobre el cerebro humano, sino de un estudio de toda la serie de los mamíferos. En ocho años de trabajo, analizó la corteza cerebral de algún representante de los principales grupos de mamíferos, además del humano, en diversas publicaciones que fueron apareciendo antes que el libro. Éste es entonces un extracto de esos estudios y previene al lector acerca de sus numerosas lagunas, por la necesidad de exponer exclusivamente aquello que considera necesario para la deducción y fundamentación de su teoría, y por la insuficiencia general de lo que puede ofrecer en este primer bosquejo. "La teoría de la localización está recién emergiendo".
En la introducción, describe Brodmann sus objetivos específicos:
1) La localización histológica de la corteza cerebral, desde el punto de vista exclusivo de las características anatómicas.
2) La división de la corteza cerebral de acuerdo con sus características anatómicas concordantes, es decir el resumen sistemático de partes y complejos tisulares de similar valor y su separación de otros, todos diferentes entre sí. La localización comparada consiste en la detección de zonas homólogas en el resto de los animales.
3) La obtención de una teoría orgánica de la superficie cortical cerebral basada en características anatómicas, según la propuesta inicial de Meynert.
Descarta así de su estudio, el análisis de los haces de conducción entre partes de la corteza o entre ésta y los centros inferiores y la mielogénesis, que es un estudio ontogenético y no histológico, al que califica de dudoso en sus resultados.
Dentro de los estudios histológicos propiamente dichos, distingue el citoarquitectónico y el fibriloarquitectónico, que en ese momento transitaba sus primeros pasos y el mieloarquitectónico,] que ya se estaba desarrollando en el mismo laboratorio y con el que la citoarquitectura presentaba, aparentemente, resultados similares o complementarios.
Dentro de la citoarquitectura, describe la división cortical de acuerdo a elementos celulares particulares, que es bastante limitada y cuyo paradigma es la zona precentral con las células piramidales gigantes de Betz. Tampoco la división topográfica estratigráfica (por capas), le parece de utilidad, teniendo en cuenta que es difícil atribuir a una capa cortical específica, una determinada función, por lo que prefiere para su investigación, la localización topográfica, o sea: "la división local de la corteza cerebral en campos estructurales o, lo que es lo mismo, la división en regiones espaciales extendidas en superficie, delimitadas regionalmente, unitarias en sí mismas y diferentes entre sí en su estructura" (Areae anatomicae").
Sin solución de continuidad, describe Brodmann su método, que se basa en el análisis del corte transversal de la corteza, especialmente, en su apariencia estratificada. "La cito arquitectónica, está ligada en especial, a la estratificación celular". Estos estratos tienen un comportamiento regional muchas veces extremadamente diferente. La localización comparada se basa en la investigación del comportamiento local de esa estratificación en la escala de los mamíferos y cómo en base a ella, se alcanza una división topográfica de la superficie cerebral en los animales y en el hombre.
Como describe otros aspectos del método en forma desordenada, en otras partes del libro, hemos decidido incluirlos todos aquí.
Brodmann utilizó cortes coronales seriados completos del mismo espesor y también axiales, de cerebros fijados en formol, incluidos en parafina y teñidos con la técnica de Nissl simplificada, que permite ver los cuerpos celulares, y analizó esos cortes bajo aumentos similares, y especialmente, utilizó microfotografías y proyecciones de los cortes para poder compararlos. Para ello, contaba con un microscopio Zeiss de "última generación" que muchos quisieran tener ahora. Estaba equipado con una platina especial para preparados grandes y permitía tomar fotos y realizar proyecciones. En algunos casos, especialmente en animales grandes, realizó "recortes más o menos grandes de hemisferios". Además de los ejemplares de adultos, contó en muchos casos con especímenes fetales y jóvenes. Nos da la lista de especímenes examinados (detallamos los órdenes solamente):
I. Primates: Homo, Simia, Anthropitecus, Semnopithecus, Cercocebus, Cercopithecus, Macacus, Cynocephalus, Ateles, Lagothrix, Cebus, Pithecia, Saimiris, Hapale.
II. Prosimiae: Propithecus, Lindris, Lemur, Pterodicticus, Nycticebus.
III. Chiroptera: Pteropus, Vesperugo. IV. Insectívora: Erinaceus, Talpa, Centetes.
V. Carnívora: Ursus, Cercoleptes, Mustela, Canis, Herpestes, Paradoxurus, Felis.
VI. Planipedia: Phoca.
VII. Rodentia. Sciurus, Mus, Lepus, Spermophilus.
VIII. Ungulata: Hyrax, Elefas, Sus, Dicotyles, Tragulus, Capra.
IX. Edentata: Bradypus.
X. Marsupialia: Phalangista, Macropus, Onychogale, Didelphis.
XI. Monotremata: Echidna.
Pero solamnte describe en detalle los cerebros completos de Homo, Cercopithecus (Chimpancé), Hapale (mono tití o mono ardilla), Lemur, Pteropus (perro volador, orden de los grandes murciélagos), Cercoleptes (Kinkajou o martucha, carnívoro arborícola nocturno pariente del mapache y el coatí), Lepus (conejo), Spermophillus (ardilla de tierra) y Erinaceus. Menciona estudios parciales de Canis, Felis, gorila y orangután y Cebus (mono pequeño del nuevo mundo o platirrino).
Divide su tarea en dos secciones principales:
1) La investigación de la estratificación cortical en el corte y sus modificaciones en la escala de los mamíferos: Tectónica cortical comparativa.
2) La división en campos de la superficie cerebral en los diferentes mamíferos en base a diferencias citoarquitectónicas: Localización topográfica comparativa de la corteza cerebral.
PRIMERA SECCIÓN
Capítulo I
En el primer capítulo se dedica a la tectónica cortical comparativa, y luego de presentar las descripciones de diferentes autores, discute en especial las de Santiago Ramón y Cajal y la de Haller, que piensan que en animales inferiores, desde el orden de los roedores hacia abajo, la corteza tiene menos capas. Para Cajal cinco en los roedores, para Haller, la base filogenética es tres en los marsupiales y va aumentando hasta alcanzar las seis capas en los roedores.
En oposición a estos autores Brodmann sostiene que la corteza cerebral tiene una estructura básica de seis capas en toda la serie y ellas aparecen ya sea en forma definitiva, o al menos transitoria, durante la ontogenia, para después diferenciarse en algunos sectores por pérdida (disminución de capas) o por formación de subcapas (aumento de capas). A esta formación la llama el tipo básico tectogenético de seis capas y a ella se refieren todas las variaciones ulteriores de la corteza. Concede, sin embargo, que las zonas "rudimentarias" pertenecientes al "rinencéfalo", muy desarrolladas en los animales macrosmáticos, y que rodean al cuerpo calloso especialmente en su parte anterior y a nivel del esplenio, constituyen una excepción a la regla y tienen una disposición estratigráfica atípica desde el inicio. En base a estas comprobaciones, divide la corteza cerebral en dos grandes tipos, formaciones homogenéticas que forman la mayor parte del manto cortical en los mamíferos mayores y que presentan gran variabilidad, pero pueden deducirse del tipo básico de seis capas, y las formaciones heterogenéticas, que son aquellas en donde no se puede encontrar embriológicamente un momento transicional de seis capas.
Brodmann fundamenta estas afirmaciones en sus observaciones personales del desarrollo de la corteza en diferentes especímenes, en donde aplicando la ley fundamental de Haeckel que dice que la ontogenia repite la filogenia, encuentra que el tipo básico de seis capas está presente en todos los estados fetales de los mamíferos investigados por él. En el hombre, a partir del quinto mes de gestación, los neuroblastos migran desde la matriz o placa interna de His y comienzan a diferenciarse las capas. En el sexto mes, ya se delimitan la quinta y sexta. Luego la segunda tercera y cuarta se depositan como una sola capa. La diferenciación de las seis capas se da entre el sexto y octavo mes exceptuando los territorios heterogenéticos del achipallium y la "corteza defectiva" de Meynert. Las capas se ven como bandas alternantes, más claras y más oscuras, a partir de la primera, la más externa, que es clara (Fig. 1).
Este estadio no se alcanza simultáneamente en toda la superficie cortical, y puede durar muy poco tiempo, para después sufrir eventuales transformaciones (secundarias) definitivas en más o en menos, como describimos antes (corteza homogenética heterotípica) o mantener el tipo básico de seis capas (corteza homogenética homotípica). De la primera, pone como ejemplo la corteza agranular de la región precentral, donde desaparece la capa IV o capa granular interna y la capa granular externa está débilmente constituida. De la segunda, da como ejemplo la corteza estriada o "tipo calcarina", en donde la capa granular interna, se divide en tres, pasando a poseer ocho capas. De las tres, la sublamina granular interna intermedia, contiene menos células que las otras dos, y corresponde a la estría de Gennari.
Brodmann reconoce que en mamíferos inferiores, como Didelfis, un marsupial, la extensión de la corteza de seis capas es muy pequeña y afirma que también existe en los Monotremas como Echidna, aunque "en una forma esencialmente modificada". Pero en todos los cerebros por él investigados, el tipo básico existe y es fácilmente comprobable. Reconoce aquí, que no tiene ningún espécimen de los cetáceos, y no dispone de fotos de Echidna y Didelphis.
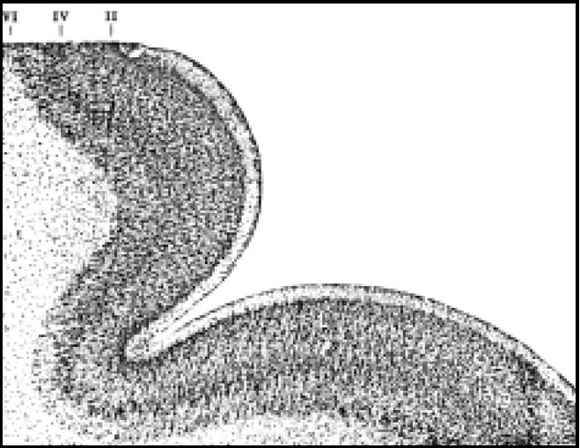
Fig. 1. Reproducción de la figura 2 del libro de Brodmann que muestra un corte de la corteza parietal de un feto humano de 8 meses de gesta en vista panorámica (20:1, 10 micrones. Las capas corticales se distinguen bien como bandas alternas claras y oscuras, comenzando por la primera, más externa, que es clara.
Capítulo II
En el segundo capítulo, Brodmann se dedica a las variaciones regionales de la estructura celular de la corteza. En cuanto a las leyes generales de la variabilidad dice que" los cambios tectónicos afectan constantemente ya sea el número y estructura especial de las capas particulares, o la densidad y tamaño de los elementos celulares en todo el corte y dentro de capas particulares, o la estructura de determinadas formas celulares o finalmente, el espesor de toda la corteza y la relación de espesor de las distintas capas entre sí". En estos criterios se basa la teoría de las diferencias locales de la estructura celular. Los cambios pueden producirse con límites muy netos o mediante transiciones más fluidas.
Da ejemplos en la corteza homotípica de aumento de la densidad celular, como en el tipo calcarina (área 17), que se establecen netamente. O el área preparietal (área 5) que muestra un aumento de la celularidad de la capa IV.
De cambios en el tamaño celular pone como ejemplo el tipo calcarina en que predominan las células granulares pequeñas, o el tipo de las células piramidales gigantes de la zona prerrolándica o el tipo preparietal.
Como ejemplos de cambios en el espesor de una capa, dice que las más frecuentemente afectadas son la II, V y IV y muestra las transiciones de la corteza entorrinal a la perirrinal y a la ectorrinal.
Dice que la corteza puede variar su espesor, sin que cambien los espesores relativos de las capas.
En el caso de las formaciones heterotípicas, reconoce aquellas con aumento por división de una capa y diferenciación de subcapas, o con disminución de capas por pérdida de una de ellas, dando aquí, nuevamente, el ejemplo de la corteza calcarina y la precentral respectivamente. También hay una disminución en el paso del neopalio al arquipalio.
Otra forma de disminución es por fusión de capas originarias y su ejemplo es algún tipo de la corteza retrolímbica. También se produce una forma de fusión entre las capas V y VI de tal forma que parecen ser una sola, en las partes orales del cíngulo24,25,33 y tipos frontales particulares (áreas 6, 8).
Una forma interesante de seudodesaparición de una capa se da frecuentemente en la capa II o granular externa, que en los individuos jóvenes presenta esa característica, y la va perdiendo por transformación de los granos en células piramidales pequeñas, lo que hace que se diferencie poco de la capa III y ha llevado al error a algunos autores. En los marsupiales conserva esta capa durante toda la vida sus caracteres juveniles.
Analiza después Brodmann, el comportamiento regional de las distintas capas y encuentra algunas muy constantes como la I y la VI. Ambas se encuentran en todo tipo de corteza tanto homo como heterogenética, aunque pueden variar de espesor en diferentes regiones.
Las capas más inconstantes son las Granulares tanto externa (II), como interna (IV).
La capa II como dijo antes, presenta una regresión clara entre estadios más tempranos y el cerebro adulto, confundiéndose muchas veces con la capa III. Esto sucede a todos los niveles en la escala de los mamíferos. Por otro lado, se presenta una capa II muy desarrollada en las regiones heterogenéticas vecinas a la cisura rinal (28 y 36).
La capa IV es la más variable de todas, ya que puede faltar en las cortezas agranulares o por el contrario ser gruesa y densa o poco visible, con escasas células, o escindirse dando origen a subcapas como en la corteza calcarina.
Las capas III y V presentan una variabilidad inter-media. La capa III se fusiona con la II formando ambas una capa reducida en la corteza retrolímbica. Frecuentemente, cambia su espesor como capa individuada, como en la corteza occipital. A veces, se divide en una capa más superficial con células pequeñas (piramidal parvocelular) y otra profunda con células más grandes (piramidal magnocelular), como ya describiera Meynert.
La capa V desarrolla a veces células ganglionares de gran tamaño, como las de Betz en el área motora (4), o en el área preparietal (campo 5). En las zonas granulares frontales, suele escindirse en dos subcapas, una superficial más celular y otra más profunda con células más dispersas. A veces se fusiona con la capa VI como en la parte anterior del cíngulo y las porciones oromediales del lóbulo frontal.
De lo expresado, se deduce que son muy variados los procesos que producen los cambios a partir del estado indiferenciado de seis capas y hacen a la rica variedad de tipos adultos. Los cambios de diferenciación se completan a veces en forma bien delimitada localmente, mientras que en otras se producen en forma muy progresiva, de tal manera que no se puede hablar de límites netos, por lo que se necesitan mucho entrenamiento y experiencia para reconocer las diferencias estructurales entre campos vecinos. Este aspecto pre-domina más en el hombre, en los lóbulos parietal y frontal. Las fotografías panorámicas son de gran ayuda en estos casos.
En la mayoría de los casos, no se trata de un solo proceso que da origen a un nuevo campo, sino que se combinan varios de los descriptos. Por ejemplo, se observa casi como regla que un aumento de la celularidad lleva a una disminución del espesor del corte, y viceversa. También los cortes con células más grandes tienen un espesor mayor que aquellos con células pequeñas. También hay que tener en cuenta que cuando en un corte aparecen nuevos tipos celulares, su tectónica cambia, y viceversa, cuando se encuentran cambios tectónicos significativos, hay que tratar de encontrar modificaciones del tipo celular, por lo que toda la tectónica del corte de un lugar de la corteza, es la que representa la unidad localizatoria.
Capítulo III
Comienza el tercer capítulo con un corto resumen de los anteriores diciendo "que la corteza de los mamíferos no sólo se origina a partir de un plan genético general, sino que en su desarrollo ulterior también está construida siguiendo leyes uniformes. Sin embargo a pesar de estas coincidencias en construcción, desarrollo y crecimiento, cada animal presenta como producto de la adaptación y herencia, ciertas propiedades estructurales según la dirección especial del desarrollo durante la vida de su cerebro tanto de la corteza en general, como de tipos particulares de la misma. Éstas son de un carácter bastante definido para cada tipo o cada grupo mayor, lo que le da a la estructura del cerebro una cualidad tal que en ciertas condiciones, es posible discernir, a partir de la forma de la estructura cerebral, la pertenencia a un tipo de animal u Orden.
No se detiene a analizar la multiplicidad de particularidades, sino que se restringirá a:
1) Cambios estructurales de tipo general en el aspecto de todo el espesor de la corteza.
2) Estructuración específica de capas particulares en ciertos animales
3) Caracteres especiales en la tectónica de áreas particulares en una Especie u Orden.
Hace notar que aunque se dedicará a estas formas de variación, ellas aparecen junto a otras y que es debido a su efecto conjunto, que se condiciona el carácter particular de la corteza en una Especie.
Respecto al primer punto, dice que las diferencias se deben a la constitución de las capas en general, al espesor de la corteza, a la densidad celular y al tamaño medio de las células. Comenta resultados contradictorios de la literatura acerca de estos puntos, y dice que su método, más arriba descripto, le permite una mayor exactitud.
1. Respecto al espesor cortical, observa que en el ser humano, varía ampliamente y da valores medios de 1,5- 4,5 mm. Como en otros animales pasa lo mismo, concluye que sólo se deben comparar zonas homólogas de la corteza, por lo que se circunscribe a dar datos de aquellas partes de la corteza de cuya homología no hay dudas (áreas 4, 6, 7, 17, 28, 29), presentándolos en forma de tabla. Aunque establece que para estas áreas la corteza humana es la más ancha al corte, halla diferencias en la serie, que son referibles especialmente al tamaño total del cerebro, más que a la ubicación del animal en la escala de los mamíferos. Sin embargo, este espesor no es proporcional a la diferencia de tamaño de los cerebros, por lo que, relativamente, animales más pequeños tienen proporcionalmente un espesor cortical mayor.
Respecto al tamaño de las células también dice que deben compararse células homólogas y el único ejemplo útil serían las células de Betz. Comenta las teorías de otros autores que sostienen que están en relación con el tamaño corporal o que dependen de la longitud del axón o relacionan su tamaño con el del músculo, grupo muscular o movimiento al que están dirigidas. Él piensa que ninguna de éstas es correcta y aporta sus propias medidas en distintos animales, encontrando que alcanzan su mayor talla en algunos carnívoros como el tigre, el lobo y el kinkajou, que porta las más grandes, a pesar de tratarse de un animal relativamente pequeño (2 kg aproximadamente). Agrega a las otras teorías, que también podría influir, la diferenciación, precisión y asociación del movimiento al que la célula está destinada.
En cuanto a la riqueza celular, se deben comparar también zonas homólogas. En esta tarea observa que las cortezas agranulares tienen menos celularidad que las occipitales, y que las mayores concentraciones celulares se encuentra en el mono, en grado intermedio en el hombre y en menor grado en el kinkajou. Si compara tipos heterólogos, la mayor celularidad se encuentra en el tipo calcarina del mono capuchino y la mas escasa en el tipo piramidal gigante de los lemures. "No es la cantidad de células por unidad de espacio, sino su cualidad, la diferenciación más fina, el desarrollo superficial, es decir, número de dendritas, que fundamenta la riqueza de sus conexiones, como lo ha expresado Bielchowsy en otro sentido, lo que constituye el patrón de la dignidad funcional de la corteza o de un sector de la misma". Como en el sistema periférico, también aquí, jugará el principal rol la diferenciación de la tarea del territorio inervado en su totalidad, junto a su extensión, la amplitud del movimiento, el tamaño corporal y sobre todo el encadenamiento cerebral.
2. Modificaciones de las capas básicas particulares en la escala de los mamíferos.
A pesar de que las seis capas son estructuras constantes, pueden sufrir modificaciones tan intensas en los distintos mamíferos que sólo a alguien entrenado le resultará posible su reconocimiento, como sucede muchas veces con las capas granulares.
I. Capa básica. Lamina zonalis. Es muy variable en espesor entre animales y entre zonas homólogas entre sí. (Brodmann da números y medidas en una tabla) No se puede sacar ninguna conclusión acerca de esta variabilidad.
II. Capa básica. Lamina granularis externa. Refiere al texto previo en cuanto a su regresión en la mayoría de los animales adultos. Permanece preservada en algunos tipos corticales en los monos inferiores y el canguro y falta casi completamente en toda la corteza del conejo y otros roedores, lo que ha llevado al error a algunos autores en cuanto al número de capas en este Orden. En los que se ha preservado, ya no se trata de células pequeñas, sino más grandes y multipolares o estrelladas, que se han diferenciado a partir de aquellas. En el hombre, las células originales de aspecto neuroblástico, se diferencian en pequeñas pirámides, lo que confunde esta capa con la siguiente.
III. Capa básica. Lamina pyramidalis. A pesar de que en forma absoluta y relativa, el ser humano posee la capa III más gruesa, no puede extraerse una ley acerca de la relación entre su espesor y la calidad de la organización de una corteza.
IV. Capa básica. Lamina granularis interna. Reconoce tres modalidades de diferenciación.
A: Las células mantienen durante toda la vida una forma juvenil (aspecto neuroblástico), como en monos inferiores y semimonos.
B: las células se diferencian hacia formas multipolares y estrelladas más grandes, que se diferencian poco de las de otras capas (hombre, carnívoros, ungulados).
C: Cambio regresivo de las células que se hacen más claras y toman forma vesicular uniforme y la capa regresa en su estructura de corte.
V. Capa básica. Lamina ganglionaris. Sufre modificaciones de espesor y de estructura celular. Repite conceptos previos sobre esta capa y se refiere al área de las "grandes pirámides". Estas células pueden estar:
A: agrupadas en nidos como en el hombre ordenamiento cumular.
B: formar una capa cerrada, ordenamiento laminar (monos inferiores, lemures, carnívoros, felinos, caninos, úrsidos, ungulados y pinipedios). El ordenamiento puede ser unilaminar (Cercoleptes) o multilaminar (Cercopithecus).
C: ordenamiento solitario células aisladas dispersas (Hombre, en los sectores inferiores del área 4)
VI. Capa básica. Lamina multiformis. Está muy desarrollada en los roedores y en algunos tipos de corteza del hombre y es muy variable en su espesor. Se refiere aquí, a la división de la corteza en dos capas principales, según Kaes: la zona principal externa (capas I- III) y la zona principal interna (IV-VI). En general, pero no sin excepciones, la capa interna es más gruesa que la externa en animales inferiores.
3. Diferenciaciones específicas de los tipos homólogos particulares.
Dice que en algunas cepas o especies la tectónica de una región específica, puede modificarse considerable-mente, sin dejar de pertenecer al tipo original, pero de forma tan característica, que con sólo estudiar esa región, se puede reconocer la especie del animal en estudio.
Divide los tipos de corteza homologa en dos grandes grupos:
1. Tipos monomorfos. Permanecen estables en toda la clase que se está estudiando con pequeñas desviaciones.
2. Tipos polimorfos. Aquellos presentan cambios tan notables, que dan origen a nuevos tipos.
A los tipos monomorfos los divide a su vez en aquellos pertenecientes a zonas corticales homotípicas (seis capas) cuyo ejemplo más representativo es la región parietal, y los pertenecientes a zonas heterogenéticas, como p. ej., las cortezas límbicas y retrolímbicas.
Da el ejemplo del tipo retrosplenial granular en Pteropus, en donde regresan las capas II y III, y se desarrolla en forma progresiva la capa VI, quedando así una zona externa mucho menos importante que la zona interna a pesar del engrosamiento de la capa I.
Un ejemplo característico de tipos monomorfos, dentro de las cortezas heterotípicas, lo constituye nuevamente, la corteza del área 4. Si bien se da prácticamente en toda la serie, está tan desarrollada y "adornada", que representa una característica de Or-den en los carnívoros, como se viera más arriba.
Para los tipos polimorfos, da el ejemplo nuevamente del tipo calcarina, que se trata de una corteza polimorfa heterotípica. El principio que la diferencia es la hipertrofia de la capa granular interna, y mieloarquitectónicamente, la aparición de la estría de Gennari.
Esta corteza puede diferenciarse de tres formas. En una, se forman tres subcapas en la capa IV (forma triestriada del tipo calcarina) características del hombre, monos y lemúridos (cerebro pitecoideo). Pero en algunos casos como el mono capuchino, la subcapa granular intermedia, se separa todavía en varias subcapas.
En la forma biestriada, la capa IV se divide en dos subcapas de las cuales la superficial contiene menos células y corresponde a la estría de Gennari, como en el gato y conejo. Finalmente, en la forma uniestriada puede haber sólo un engrosamiento de la capa IV como en los roedores pequeños o el perro volador.
Otros ejemplos de una corteza heterotípica, polimorfa, lo constituyen ciertos tipos de la corteza retrolímbica en roedores, insectívoros y quirópteros.
Da también ejemplos de tipos heterogenéticos, ya sea monomorfos o polimorfos, en el erizo, por jemplo, se encuentra un desarrollo polimorfo de la corteza olfatoria.
SEGUNDA SECCIÓN
Capítulo IV
La segunda sección del libro comienza con el capítulo IV, en el que Brodmann se propone realizar un mapa histológico-topográfico de la superficie cerebral de las diferentes especies.
Hace notar que existen en todas las especies, zonas de la corteza cerebral en donde se encuentran transiciones bruscas de un tipo a otro, y que aparecen así campos de estructura celular coincidente, Areae cytoarchitectonicae en los distintos Ordenes, permitiendo poner las bases de una división comparativa de la superficie de la corteza cerebral. También se propone constatar el tipo de división especial en áreas de los animales particulares, para detectar lo común y lo diferente en esta división.
Dice que el problema de la topografía comparativa se puede resumir en las siguientes preguntas:
1. ¿Existe coincidencia o parecido en la división topográfica en los distintos Ordenes de los mamíferos? o en otras palabras, ¿está dividida la corteza en toda la escala de los mamíferos, según una ley común, en regiones estructurales coincidentes, o se debe suponer un principio de división topográfica especial para cada orden, o al menos cada grupo mayor?
2. ¿En qué consisten en general las coincidencias y diferencias en la división cortical en campos de las distintas especies?
3. ¿Hay campos constantes e inconstantes, y cómo varían los primeros en forma, tamaño y localización en especies particulares?
Referimos a la sección de Material y método, para recordar los cerebros de los cuales tiene mapa. Brodmann reconoce que estos mapas no son exactos, sobre todo en cerebros con surcos y en zonas en donde los límites no son netos, pero dan una idea bastante útil de las regiones y áreas, que es lo que se desea. Los mapas tienen categoría de esquema y son un "medio de ayuda para la orientación". Presenta los mapas en orden descendente y después hará comentarios. Nosotros mostraremos solamente los mapas, ya que sus descripciones se limitan a determinar la zona de la corteza que ocupan, las áreas sin referirse a las características que diferencian un campo de otro. Las zonas homólogas o análogas han sido marcadas con los mismos signos diacríticos. En los epígrafes comentaremos los aspectos generales de cada mapa (Figs. 3 a 10).
Si resumimos el resultado de la descripción presentada se llegará, como respuesta a las preguntas originarias, a las siguientes conclusiones: que en principio, en todos los animales investigados, se encontró una amplia concordancia en relación con la localización topográfica cortical, pero a pesar de estas semejanzas en las tendencias básicas, en numerosos puntos, incluso entre especies estrechamente emparentadas, aparecen notables desviaciones de la división superficial. La constancia de caracteres es la expresión de una dirección del desarrollo semejante, ya sea en el sentido de una correspondencia filética, o en el sentido de una convergencia. Las diferencias, en cambio, expresan la especialización morfológica y con ella la funcional, de los cerebros particulares de los diferentes tipos y familias.
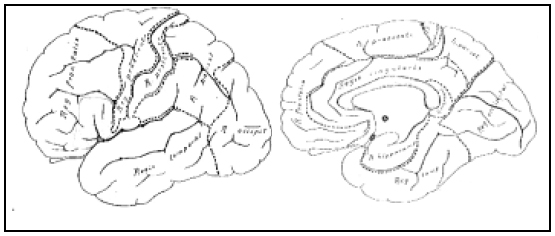
Fig. 2. Regiones principales del cerebro humano. Estas regiones existen con un desarrollo mayor o menor (tamaño y áreas) en toda la serie de los mamíferos, por lo que son consideradas homólogas. Dentro de la región frontal hemos dibujado con lápiz, el territorio de las áreas 44 y 45, corespondientes al "centro de Broca", que Brodmann dice que "merecerían formar una región aparte".

Fig. 3. El conocido mapa cortical humano de Brodmann. Una de las figuras más reproducidas en la literatura neurológica, en la misma proporción en que se desconoce su obra.
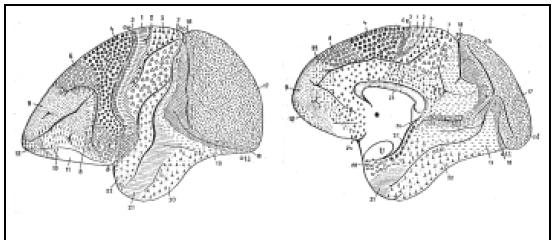
Fig. 4. Mapa de la corteza del Cercopiteco (chimpancé). Obsérvese la reducción de la región frontal, con menos áreas, y el aumento de la región occipital, especialmente el área 17 que se extiende notable-mente hacia la convexidad. Tampoco existen las áreas 39 y 40 del lóbulo parietal inferior del hombre, y aumenta la extensión del área preparietal (5). Así como el cerebro del hombre, se ha desarrollado ha-cia adelante y es un cerebro "frontal", el del mono ha experimentado un gran desarrollo posterior y es un cerebro "visual".
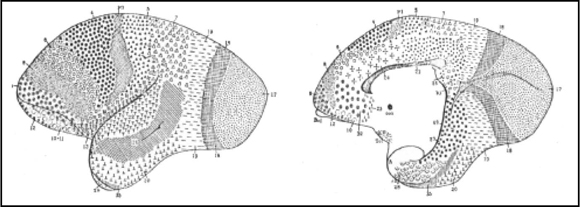
Fig. 5. Mapa cortical del mono tití (Haphalidae). Los cerebros lisencéfalos son bendecidos por Brodmann, porque en ellos se notan mejor las transiciones entre áreas. Como en los otros monos, gran extensión de la región occipital, reducción mayor de la frontal y apertura de la cisura de Silvio, que permite apreciar el área 13 (ínsula). Gran extensión del área preparietal (5).

Fig. 6. Mapa cortical de cerebro de lemur. Persiste el predominio occipital y reducción mayor frontal, aparece más expuesta la ínsula (13-16) y las áreas mediales del lóbulo temporal comienzan a aparecer en la convexidad (28), lo mismo que las olfatorias (51) iniciando la torsión hacia atrás y afuera del lóbulo temporal descripta por Brodmann. En la región poscentral se distingue sólo un área. Comienza el desarrollo del área retrosplenial, que es francamente regresiva en los primates (26, 29, 30).
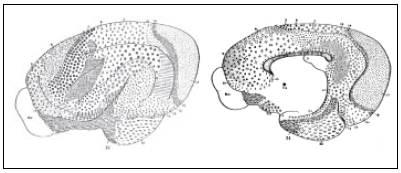
Fig. 7. Mapa cortical de Cercoleptes (kinkajou) carnívoro nocturno pequeño, emparentado con la marta y el coatí. Regresión importante de la región frontal, donde sólo persisten las áreas 8, 12, 25 y 32. El área 4 tiene una forma muy característica en los carnívoros: rodea a la región poscentral por debajo, incurvándose en forma de jota. El área auditiva se encuentra por encima y rodeando la terminación de la cisura de Silvio.La región retrosplenial está bien desarrollada con sus tres campos 26, 29 y 30.

Fig. 8. Mapa cortical del perro volador (Pteropus).Queda solamente el área 12 de la región frontal. Gran desarrollo de la ínsula y del lóbulo olfatorio. En la región parietal no hay diferenciación neta entre campos 5 y 7. En la región hipocámpica aparece el área 48 y 49, como campos nuevos. En el lóbulo olfatorio se diferencia bien el área prepiriforme, el tubérculo olfatorio y el núcleo de la amígdala. La región temporal es pequeña, así como la parietal.
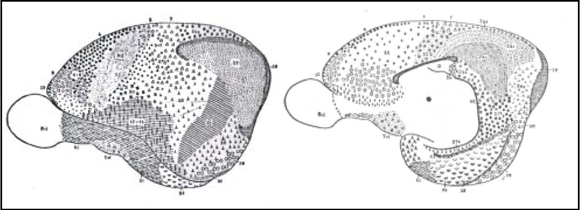
Fig. 9. Mapa cortical de Lepus (conejo). La región frontal es muy pequeña y está representada por el área 12 que es dudosamente granular según Brodmann. Gran desarrollo de la región retrosplenial, que presenta seis áreas distintas, cinco de las cuales son subáreas de la 29. Desarrollo importante de la ínsula y el lóbulo olfatorio. Las áreas neopaliales terminan arriba, en el margen superior del hemisferio, y el área 4 se dispone más horizontalmente. Las áreas 17 y 18 ocupan sólo la parte posterior de la convexidad.
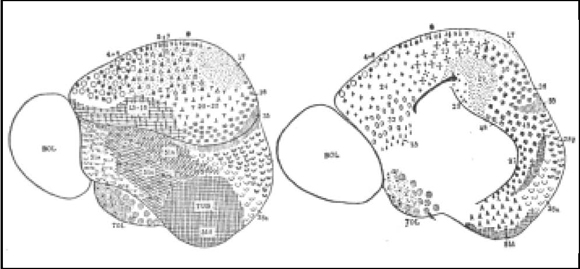
Fig. 10. Mapa cortical de Erinaceus (erizo). La corteza homogenética (neopalio) es pequeña, mientras que la heterogenética (rinencéfalo) es extensa. El área olfatoria presenta además del tubérculo olfatorio y la amígdala, cuatro campos que Brodmann designa como 51 a, b, c y d. En el Tubérculo olfatorio encuentra tres núcleos, anterior, medio y posterior. En la región hipocámpica encuentra los campos 27, 28a, 28p, 48 y 49. En el área precentral faltan las células de Betz, por lo que no se distingue más 4 de 6. Lo mismo sucede con 1, 5 y 7 en el parietal y 20- 22 en el temporal.
Capítulo V
En el capítulo V se ocupa de las coincidencias de la división en campos y dice que se exteriorizan en tres puntos: la semejanza de la construcción total, la coincidencia de regiones estructurales principales y la persistencia de campos particulares en toda la serie.
1. La semejanza de la construcción total
Al respecto dice que mirando toda la serie sin prejuicio, hay que aceptar tal semejanza. Tanto en cerebros con surcos, como en lisencéfalos, se encuentra la misma división en regiones estructurales. En todos los mapas se encuentran segmentos totales, parciales, campos coronales y calotas terminales, en el ordenamiento uno detrás del siguiente, que se aprecia mejor en el corte horizontal.
De los segmentos en cinturón, menciona las áreas 18 y 19 en el lóbulo occipital y las áreas 9 y 10 en el frontal. Como segmentos parciales tenemos las áreas 19 y los campos 21 y 22 temporales y 5 y 7 parietales. También existe gran semejanza en la división de las circunvoluciones del cíngulo y del hipocampo. "La base de la división en campos de la corteza cerebral es la misma en todos los Órdenes de los mamíferos y en ella reina el principio de la segmentación, que se manifiesta más o menos claramente. Su sentido y contenido dice que la corteza se divide en un gran número de zonas estructurales que se ordenan groseramente en sentido oro-caudal, y asumen la forma de segmentos o fragmentos de segmentos". Esta disposición se manifiesta más claramente en los cerebros lisencéfalos.
2. La constancia de las regiones principales
A este respecto reconoce a las mismas como circunscripciones cerradas histológicamente, que engloban varias áreas particulares parecidas entre sí y que aparecen básicamente en todos o casi todos los grupos de animales aunque modificadas en detalles y a menu-do fuertemente disminuidas. Si se comparan estas regiones principales del hombre con las de los otros animales, se comprobará que presentan una sorprendente constancia, aunque varíen en el número de áreas particulares o en su forma y tamaño.
Analiza estas regiones principales, para determinar sus caracteres generales, comenzando por la región precentral, que se caracteriza por la ausencia de la capa granular interna y su gran espesor. Está constituida por dos tipos particulares, más o menos discretos: el área gigantopiramidal y el área frontal agranular, salvo en algunos mamíferos inferiores como insectívoros, microquirópteros y algunos roedores. Se ubica lateral-mente por encima de Silvio y se extiende hasta la callosomarginal, por delante de Rolando. Cuando estas cisuras no existen se encuentra por encima de la ínsula y su límite posterior es incierto, lo mismo que el anterior, éste en todos los casos, salvo en Cercopithecus, en que coincide con el surco arcuato, y en Cercoleptes, con el sulcus praesylvius.
La región principal frontal granular ocupa todo el lóbulo frontal y se extiende hasta el polo, por delante de la región agranular frontal. Su característica principal es la reaparición de la capa granular interna. Presenta también una estratificación débil, escasez celular, mayor espesor y una transición progresiva a la sustancia blanca. Es la región más variable en tamaño y localización, como en número de áreas. En el hombre constituye partes del lóbulo frontal y contiene ocho a nueve áreas: 8-11 y 44- 47. Es más pequeña en los monos y contiene sólo cinco áreas, y va descendiendo en tamaño y número de campos, hasta desaparecer en los mamíferos inferiores nombrados en el párrafo anterior.
La región principal postcentral y parietal aparece en todos los mamíferos por detrás de la zona agranular precentral, en forma brusca, constituyendo así una característica constante en la escala de los mamíferos. En los más pequeños y primitivos, no se pueden diferenciar subregiones, mientras que los más evolucionados presentan ya sea un área postcentral con un solo campo, o varios, como en el hombre y los monos superiores. Por detrás está la región parietal, separada en dos campos en los animales superiores, que se van desdiferenciando a medida que se desciende en la escala, hasta constituir uno solo, que se termina fusionando también con la región postcentral, formando un solo campo y una sola región.
La región principal de la ínsula es la más constante de la escala y notable por su forma y tamaño. Se la reconoce por la presencia del claustro, que para Brodmann sería un desprendimiento de la VI lámina, como una subcapa. De los lemures hacia abajo, aparece en la superficie cerebral y llega hasta el surco rinal posterior y anterior. Característicamente, se divide en una porción agranular anterior y una posterior, granular.
La región principal occipital se caracteriza por la corteza estriada o campo 17, con un crecimiento inhabitual de la capa IV, y un espesor cortical pequeño, con riqueza celular importante. Su lugar corresponde al lóbulo occipital y su vecindad, extendiendose más o menos en la superficie externa. Debido a estos caracteres, su reconocimiento no es difícil.
Como zona general, la región temporal principal, puede ser homologizada, pero no en sus áreas particulares. Si bien no presenta características especiales, es reconocible por su localización. Disminuye su extensión a medida que se desciende en la escala.
La región principal cingular se destaca por su localización constante inmediatamente por fuera del cuerpo calloso, pero también es identificable en los animales sin cuerpo calloso, como monotremas y marsupiales. Se distingue también por su estructura celular característica. Tiene una porción anterior agranular y otra posterior granular, que se diferencian en un número mayor o menor de áreas de acuerdo a la especie.
La región principal retroesplenial se delimita de la sección posterior de la región cingular por la regresión de varias capas básicas, y posee un tamaño y desarrollo tectónico muy variable. Por su localización corresponde al istmo de la circunvolución cingular. Está muy desarrollada en extensión y número de áreas en algunos macrosmáticos como el conejo y es muy rudimentaria en los primates en donde está constituido por uno a tres campos.
La región principal hipocámpica es junto a la precentral e insular, la más constante, y pertenece a las formaciones heterogenéticas. Su estratificación atípica la hace reconocible en todos los mamíferos. Se extiende por dentro de los surcos rinal posterior hasta el surco hipocámpico, es decir la cabeza del hipocampo y una parte del lóbulo piriforme lo mismo que la corteza profunda del surco hipocámpico. Corresponde a las áreas 27, 28, 35, 48 y 49.
La región principal olfatoria está constituida por formaciones corticales rudimentarias. A veces puede distinguirse una especie de capa I y VI, pero casi siempre falta una verdadera estratificación (tubérculo olfatorio y amígdala). Su presencia y tamaño en la serie son muy inconstantes. En macrosmáticos de organización simple es muy prominente, ocupando 1/3 de toda la corteza, como en el erizo, y cubre casi todo el lóbulo piriforme y el lóbulo olfatorio anterior. También está muy desarrollada en pequeños roedores, mientras que en microsmáticos está minimizada, como en los lemures. Dice Brodmann que no ha encontrado su homólogo en los primates.
3. La persistencia de campos individuales
Como las regiones, también se encuentran áreas que persisten a través de toda la escala o de casi toda ella. Característica de estos campos es su estructura muy diferente de la originaria, por lo tanto, su pertenecía a las formaciones heterotípicas y heterogenéticas. La homología de éstas no está dada solamente por la semejanza de la estructura estratigráfica que puede llegar a ser bastante variable, sino por la coincidencia de su localización respecto a campos homólogos vecinos.
Pueden ser designadas como homólogas las siguientes áreas:
1 (1-3 o postcentral), 4 (gigantopiramidal), 6 (frontal agranular), 5 (preparietal), 13-16 (insular), 17 (estriada), 18 (occipital), 26 (retroesplenial), 27 (presubicular), 28 (entorrinal), 35 (perirrinal), 51 (prepiriforme).
Capítulo VI
En el sexto capítulo, Brodmann se dedica a analizar las diferencias de la división cortical en áreas, y dice que son más numerosas que las coincidencias. Se pueden deber a características superficiales (no esenciales) como forma, localización o tamaño, o a diferenciaciones tectónicas específicas (esenciales), que llevan a la adquisición de nuevos campos o a la regresión, minimización o fusión de áreas y regiones en su totalidad. Estos factores casi siempre suceden simultáneamente.
1. Variaciones no esenciales
Son tan frecuentes, como los mapas dejan ver, que sólo da algunos ejemplos, haciendo notar que para ello, se prestan especialmente los campos absolutamente homólogos que describimos arriba. El tipo 4, gigantopiramidal p. ej., ocupa el tercio medio o anterior del hemisferio, de acuerdo al desarrollo del lóbulo frontal, pero las diferencias localizatorias son tan importantes, que para cada grupo mayor se puede describir un tipo localizatorio propio, en los primates con forma de cuña vertical, en los carnívoros con una angulación hacia atrás en su extremo inferior, en los ungulados una posición más horizontal, que se acentúa en insectívoros y roedores, acercándose al margen superior del hemisferio. Hemos visto que su relación con la cisura central no es constante. En cuanto al tamaño absoluto, es más grande en el hombre, pero relativamente al tamaño general del cerebro, es la más pequeña.
En cuanto al área 17, ocupa el polo occipital y sus alrededores, formando una especie de calota que se extiende más hacia la cara externa o interna, en relación con la cisura calcarina. En cuanto a su extensión, es máxima en los grandes monos; entre los antropoides, especialmente el orangután. En el hombre representa el 2% de la superficie total, mientras que en el mono, la relación es del 10 al 15%.
El área entorrinal (28) es el territorio estructural más constante junto con el área presubicular, más desarrollada en los macrosmáticos, y relativamente pequeña en el hombre (1/2 a 1% del total) y en los monos (2%). Su localización varía según la diferente estructura del archipallium, especialmente del lóbulo piriforme. En el hombre y los monos está en la cara interna del lóbulo temporal y representa una pequeña zona en el extremo del surco hipocámpico en la cabeza de la circunvolución del mismo nombre. En los lemures alcanza el polo temporal y la cara externa, y va exteriorizándose cada vez más cuando descendemos en la escala zoológica, respondiendo a una rotación dorsocaudal del borde basal del cerebro.
2. Variaciones esenciales
Se trata de la aparición de nuevas zonas, que faltan o están constituidas en forma rudimentaria en otras especies, o viceversa, de desaparición o reducción de áreas que existen en otras especies. Desde el punto de vista genético, son en parte adquisiciones, en parte atrofias o reducciones de campos anatómicos corticales, las que se resumen aquí. No es fácil determinar cuál de estos procesos se ha puesto en juego, cuando se encuentran cambios en la división en campos. Sin duda ambos compiten entre sí en el desarrollo de las áreas corticales. Cuando existe una presión exterior hacia un determinado desarrollo, p. ej., la visión o el oído o el olfato, los mecanismos del sistema nervioso y entre ellos los centros corticales, experimentan un desarrollo unilateral y deben hipertrofiarse en cierta manera, mientras que los otros, que no mantienen el paso de esta progresión, retrasan su crecimiento, o se atrofian relativamente.
En un sentido inverso, cuando un sentido regresa por condiciones externas contrarias, la disminución de la función se acompaña de la atrofia anatómica o desaparición, en casos extremos, de los centros correspondientes y un predominio relativo de los otros. Las condiciones que influyen en la división localizatoria de la corteza cerebral son entonces realmente complicadas y su juego simultáneo e interacción, determinan las grandes diferencias de división en áreas incluso entre especies relativamente emparentadas. Estas influencias son parcialmente accesibles debido al escaso conocimiento de la funcionalidad de la corteza cerebral, por lo que nos resulta muy difícil determinar las razones de una división localizatoria particular. Sólo podemos registrar los hechos e interpretarlos comparativamente como desarrollos progresivos o regresivos.
Como progresivos señalaríamos junto al aumento de la superficie de una región principal, el aumento de campos particulares dentro de ella. Por ejemplo, el desarrollo extremo de la región olfatoria en los macrosmáticos, junto con la rica diferenciación localizatoria como se observa p.ej., en el erizo.
Lo mismo sucede con la región retroesplenial en el conejo de la que ya hemos hablado, la región hipocámpica en Pteropus con un área 49 que no encontró en otros mamíferos , y el área 27 en el conejo; la región cingular, que presenta siete u ocho campos en Pteropus y la región principal frontal, que no existe en los mamíferos inferiores y recién aparece en los carnívoros, para ir aumentando progresivamente a través de los Ungulados, Pinipedios, Prosimios y Primates y que además en los monos y el hombre ha desarrollado una gran cantidad de nuevas áreas.
En el hombre es donde se manifiestan las apariciones de nuevos campos exclusivamente en el neopalio, y predominantemente en los lóbulos frontal, parietal y temporal. El hecho de que estas transformaciones aparezcan en especies inferiores indicaría que se trata de un desarrollo unilateral, condicionado por la división del trabajo, es decir, de una especialización funcional (sensorial, motora, intelectual) del grupo zoológico correspondiente.
Es más difícil reconocer a las transformaciones regresivas. Se trata aquí de órganos cataplásticos o regresivos (Haeckel), desde el punto de vista fisiológico. En casos extremos, se trata de partes "degeneradas" o de "instrumentos sin utilidad". La prueba morfológica del desarrollo cataplástico se basa en tres tipos de hechos: primero, a la marcha empíricamente comprobada de la ontogenia individual en la que una parte atrófica, presenta en la vida embrionaria un desarrollo mejor, ya sea en cuanto a masa, o cualidad; segundo, en la anatomía comparada de la misma parte en grupos emparentados, y tercero, en la comprobación del grado de diferenciación morfológica de ese sector dentro del órgano en un determinado animal (casi nunca claro y muy poco decisivo). En estos casos, se debe ser sumamente cuidadoso en la interpretación.
Algunos casos, sin embargo, serían pasibles de ser tomados como regresivos: aquellas áreas que están en relación con partes atróficas del cuerpo, p.ej., un órgano de los sentidos, como los ojos en topos y ratoncitos ciegos, no presentan trazas de corteza del tipo calcarina, o los microsmáticos, que presentan las correspondientes partes cerebrales sumamente atróficas. Lo mismo puede decirse del área retrosplenial de los primates y las zonas de "defekte rinde" de Meynert, como las tenia tectae, striae de Lancisi, septum pellucidum y el área preterminal.
Como dice Haeckel, la degeneración de un órgano está asociada a su diferenciación ulterior y a menudo no puede separarse de ella. La lucha por la supervivencia genera cambios no sólo progresivos, sino regresivos, cuando los últimos son más útiles a todo el organismo. El perfeccionamiento de un órgano compuesto está unido a la degeneración de algunas partes constitutivas, si ésta se constituye en una ventaja decisiva para el desarrollo progresivo de las partes restantes y a través de ello, del todo.
Capítulo VIII
En el octavo capítulo, Brodmann se ocupa de las implicancias que sus observaciones puedan tener en el progreso del conocimiento de la patología de distintas enfermedades del sistema nervioso. No las trataremos en profundidad, pues como él mismo dice, estos estudios estaban en ese momento en sus comienzos y los autores no coincidían sobre el tipo de alteraciones que se presentaban. Trata primero la esclerosis lateral amiotrófica y las alteraciones que se detectaron en el área motora y luego, lesiones secundarias de la corteza pos central en el tabes dorsal. Dice que muchas alteraciones descriptas, pueden deberse a una falta de conocimiento de las diferencias de la estructura cortical en las distintas áreas, que llevan al error, considerando patológico, lo que es normal pero con una estructura diferenciada.
Es más interesante el párrafo que dedica a la explicación de la idiocia. Según descripciones recientes (para ese momento), de Alzheimer, las idiocias tendrían tres orígenes: a) a partir de focos lesionales múltiples predominantemente inflamatorios; b) por "procesos difusos corticales"; c) por trastornos primarios del desarrollo (agenesia, aplasia etc.). Se ocupa lógicamente, de éste último grupo.
Según O. Vogt, en las formas de idiocia producidas por trastornos del desarrollo, incluso aquellas que se establecen después del nacimiento, existen estadios de detención de la ontogénesis. Como vimos antes, existe una forma originaria de corteza cerebral en el desarrollo embrionario, que por transformación ulterior produce los diferentes tipos corticales locales. Se podría pues encontrar en algunas idiocias una detención en una fase del desarrollo embrionario. Será pues necesario para la comprensión de las formas congénitas de debilidad mental, conocer aproximadamente el momento en que se instala la inhibición del proceso de desarrollo ontogenético. La estructura tectónica podría dar un punto de referencia acerca del estado más o menos indiferente del estadio de transición ontogenético para decidir sobre esta pregunta. El hecho de que diferentes partes de la corteza se desarrollan en diferentes momentos, no ha sido debidamente considerado, y es importante su conocimiento exacto para evitar errores diagnósticos. Las alteraciones tectónicas pueden darse por inhibición del desarrollo o también por un exceso de producción en algunas capas como describiera Vogt.
Menciona en este sentido, la corea de Huntington, en la que Kölpin habría detectado un aumento en la construcción de la cuarta capa, constituida por elementos de apariencia joven, lo que se interpretaría como la persistencia de un estadio juvenil de la corteza, en el estado adulto.
inalmente menciona las alteraciones de las cortezas sensoriales, cuando el órgano periférico falta.
Capítulo IX
En el último capítulo de su libro, Brodmann teoriza sobre una organología fisiológica de la corteza cerebral.
Discute inicialmente el concepto de equivalencia de la corteza cerebral emitido por Flourens a principios del siglo XIX, cuando nada se conocía de las diferencias locales que existen en ella. Dice que esta tendencia todavía se mantiene a principios del siglo XX, aunque son pocos los autores que adhieren a ella. Meynert sostenía que las células cumplen la misma función y que el resultado depende más bien de las conexiones entre ellas y de las aferencias y eferencias de la zona. Exner y Wundt, el fundador de la psicología moderna, compartieron esta opinión.
En la diferenciación celular que existe en la corteza, generada a partir de una base originaria en que todas eran similares, ya se expresa una especificación histológica y una división funcional del trabajo. Tanto en el hombre como en el animal, la diferenciación se hace de igual manera en iguales sitios, y muchas veces tempranamente. "Donde en un organismo, o dentro de un órgano, se encuentran nuevas categorías celulares, suponemos que existe un rendimiento cualitativo especial". Y da el ejemplo de las pirámides gigantes de Betz.
"El polimorfismo de las células, su diferenciación en distintos momentos, la separación regional estricta de ciertas formas celulares y la regular aparición de los mismos tipos celulares (homólogos) en lugares idénticos de la corteza de todos los mamíferos, permiten la suposición de que en la corteza cerebral ya entre los elementos celulares, ha tenido lugar una amplia especialización de las funciones, una distribución de diferentes tareas sobre células de diferente característica morfológica, diferente localización y diferente valor".
Rechaza de plano cualquier atribución de una función psíquica a cualquier tipo de célula, ya que sólo se puede esperar de una estructura tan simple, realizaciones del tipo más elemental.
Discute también la postura de Kappers que le da valor psíquico a la agrupación celular en capas, p.ej., los elementos granulares tendrían funciones receptoras, las capas infragranulares serían responsables de las asociaciones intrarregionales y las pirámides supragranulares de las asociaciones interregionales de orden superior. Dice Brodmann que no se puede deducir la importancia psicológica por la mayor o menor masa o tamaño del grupo, sino por la organización interna, (química, dinámica, o aún histológica, pero inaccesible a las demostraciones microscópicas del momento).
En cuanto a la localización funcional en general, dice con Von Monakow, que hay dos tipos de investigadores: los "localistas estrictos" y los "semilocalistas".
El hecho de la diferencia de las áreas corticales, es el fundamento del "Principio de la función localizada". Las distintas posturas se basan sólo en la cantidad y tipo de aquello que debe ser localizado.
Golz, por ejemplo, sostiene que el cuadrante anterior y posterior del hemisferio se diferencian funcionalmente, pero no reconoce la existencia de "esferas de los sentidos" especiales. Gudden, sostiene que al menos se deben reconocer dos regiones principales, una para el movimiento y otra para las representaciones de las sensaciones.
Los localistas estrictos como Munk, sostienen que hay campos perfectamente delimitados para los distintos sentidos y para el movimiento y que existe una equivalencia de los centros corticales con las distintas superficies periféricas de los sentidos. Flechsig no sólo proyecta los campos sensoriales en forma estrictamente regional, sino que habla también de "centros de la comprensión", localmente ubicados. Últimamente, dice, ha predominado la localización estricta por la teoría de la afasia y los trabajos de Flechsig, pero piensa con von Monakow, que hay un error en la forma en que se plantea el problema a partir de las conclusiones extraídas de las observaciones.
Habla después, de uno de los temas más apasionantes de su libro, que es su interpretación del principio de la función localizada desde el punto de vista de la morfología.
Recuerda que la corteza cerebral está constituida por una suma de órganos particulares altamente diferenciados (ver capítulo VII), cada uno en un lugar y con una estructura determinados, no sólo por el agrupamiento y asociación de los elementos celulares, sino también por las diferencias en los tipos de células individuales. Estos "centros" histológicos se dan no sólo en el hombre, sino en todos los animales con la misma claridad, aunque en menor número. "Partes fisiológicamente diferentes, tienen una estructura diferente". "Partes que son diferentes, deben servir a tareas diferentes".
El principio de la función localizada dice como ley infranqueable, que cada tarea bajo condiciones dadas, indica una región determinada en el órgano central o, si se trata de una función compleja corresponde a un complejo de regiones. Dice Brodmann que sus comprobaciones certifican con total seguridad la presencia de una localización regional estricta de ciertas funciones. En este caso, los campos están delimitados en forma neta de sus vecinos, p. ej, 4, 17, 27, 28 y la ínsula. El significado de estos campos, su aparición en toda la serie de mamíferos, serían difíciles de explicar, si no se acepta que se trata de sustratos anatómicos para funciones electivas fijas, preformadas en el sitio y delimitadas espacialmente, en realizaciones o tareas que le corresponden exclusivamente a esos territorios y que de alguna manera se diferencian específicamente de las actuaciones de todos los otros territorios. ¿Qué debemos localizar en esas áreas?
Funciones totales o colectivas. La suposición de "centros psíquicos de orden superior "es insostenible". Se trata de "puntos de vista ingenuos". Las capacidades mentales son conceptos que involucran extraordinariamente, complejos enmarañados de funciones elementales. Requieren entonces, el entrelazamiento e interacción de procesos elementales, es decir de la función simultánea de numerosas áreas corticales, si no de toda la corteza, incluyendo también partes subcorticales.
En verdad, existe sólo un centro psíquico y éste es el cerebro, como un todo, con todos sus órganos.
No se debería decir con esto, que para los procesos psíquicos superiores todos los órganos particulares colaboran de igual manera, sino que para cada caso concreto, los "focos funcionales" a disposición, entran en acción en distinto número, grado y forma de conexión. Se podría así, suponer una cierta predilección de lugar para determinados procesos. Un proceso de este tipo será siempre la resultante y no la suma de las funciones de un gran número de órganos parte, distribuidos más o menos ampliamente sobre la superficie. La diversidad, tipo y graduación de los procesos más altos de la conciencia son, según lo dicho, la expresión de una variedad interminable de combinaciones funcionales de órganos corticales particulares.
Funciones localizadas
La localización absoluta. Como la corteza está dividida en áreas distintas que tendrán distintas funciones, en aquellas en que los límites son netos, consideraremos necesariamente, que la función está estrictamente localizada. De aquí no surge que una capacidad elemental esté localizada en estos centros, por ejemplo, los centros de los sentidos, sino que serían áreas que resumen las diferentes funciones periféricas de los órganos correspondientes (p.ej. para la visión, no sólo la percepción de la luz sino del movimiento de los ojos, la integración de reflejos visuales, etc.). Los límites fisiológicos son tan netos e invariables como los morfológicos, y también los correspondientes haces tienen origen sin excepción dentro de estos límites.
La localización relativa. En aquellas áreas con límites poco netos, hay superposición de campos y, se podría suponer, también de funciones. Se entiende con esto, que existen para diferentes zonas del organismo, capacidades corticales subordinadas que se superponen parcialmente en la corteza. Se trata de territorios que pueden también abastecer otras funciones (Principio de la representación múltiple de territorios corticales).
Estas zonas, serían quizá responsables de la restitución de funciones perdidas por procesos patológicos incluso luego de años de instalado el déficit, por el reemplazo de las zonas lesionadas por otras relacionadas (Principio de la suplencia funcional). Esta restitución suele ser debida a la ejercitación.
Esto se explicaría de dos maneras. Una, es que la zona cortical no está diferenciada funcionalmente, está en "indiferencia fisiológica", preparada para el reemplazo. La descarta, porque recuerda que las partes sin función, se atrofian.
La segunda es que la zona que realiza la suplencia toma parte normalmente en el proceso que ha sido dañado y asume entonces toda la función perdida, o que esa zona prestaba una colaboración que regresó con el tiempo, y luego tuvo que fortalecerse por la necesidad. En el primer caso, se trata de una adquisición, en el segundo, de un aumento de la función por ejercitación.
Esta suplencia puede darse por áreas simétricas del hemisferio contralateral, o por sectores vecinos o alejados del mismo hemisferio que están sin embargo, relacionados por su participación normal en la función ahora dañada.
En una sección sobre la localización especial, da datos sobre el área motora, el área visual, las áreas del lenguaje y centros funcionales en los animales para terminar su libro con el famoso adagio de von Gudden:
"Un hecho fisiológico contradictorio, pierde su significado ante un hecho anatómico comprobado, ... Primero pues, anatomía y luego fisiología; cuando sin embargo, primero fisiología, nunca sin anatomía".
COMENTARIO
La traducción
Hemos traducido directamente del alemán el capítulo VII del libro de Brodmann, ya que estimamos que resume bastante bien sus investigaciones histológicas sobre la corteza cerebral y presenta algunos puntos polémicos como la deducción de la organología cortical y la posición del hombre en la escala de los mamíferos desde el punto de vista de la teoría de la localización cerebral citoarquitectónica.
Además presentamos un resumen del resto del libro, lo más claro, breve y ajustado a las ideas de Brodmann, capítulo por capítulo, rescatando textualmente algunas de sus frases, que nos parecieron destacables. Éstas aparecen entre comillas o remarcadas en negrita.
La traducción fue realizada en el mes de enero de 2010 en una apacible ciudad de la costa bonaerense y, aunque hemos tratado de comunicarle algo de nuestro estado anímico en esas circunstancias tan ideales, no sé si habremos alcanzado a hacer amena su lectura. Nos hemos sí, mantenido fieles en la transmisión de las ideas de Brodmann, a través de su texto verborrágico y repetitivo, al que hemos pulido al máximo, tratando a la vez, de no ser doblegados por la gramática alemana, para permitir una lectura al menos, cómoda. Agradecemos a la Dra. Zúccaro la lectura y corrección final del manuscrito, en busca de lo mucho inadvertido por nosotros, para mejorar la calidad del texto.
LOS APORTES DE BRODMANN
1. Seis capas
Aunque fueran Baillarger y Berlin transitoriamente y después ya definitivamente Bevan Lewis2 quienes primero describieran la estructura cortical en seis capas, recién con Brodmann alcanzó su instalación definitiva como conocimiento adquirido para la neurobiología, gracias a sus estudios embriológicos, ya que fue el primero en indagar la generación de la corteza cerebral a partir de la migración de los neuroblastos desde la capa germinativa subventricular descripta por His3. La primera capa se forma primero como zona limitante, antes de la migración de los neuroblastos. Luego, describe Brodmann la migración neuroblástica en dos movimientos, uno primero, que forma la capa VI y luego, a partir de una acumulación de células por encima de esta capa, se diferencian las otras cuatro. Esto sucede entre el sexto y octavo mes, en diferentes tiempos para diferentes zonas de la corteza. En este estadio primitivo, las seis se destacan muy bien, porque se alternan a bajo aumento, una capa clara, con otra oscura, a partir de la primera, que es clara.
Según la ley de Haeckel, que dice que la ontogenia repite la filogenia, Brodmann estableció como principio, que este estadio representaba la corteza primitiva, y que se lo debía encontrar en estadios fetales de otros especímenes de la serie, comprobándolo también en sus observaciones. A las formaciones corticales derivadas de ésta, las llamó corteza homogenética, que puede mantenerse toda la vida con seis capas, recibiendo la calificación de homotípica, o diferenciarse por disminución o pérdida de una capa (áreas 4 y 6), o por subdivisión y aumento de capas (área 17), tipos corticales a los que calificó de heterotípicos.
A las cortezas "rinencefálicas", no pudo encontrarles nunca un estadio intermedio de seis capas, por lo que las llamó cortezas heterogenéticas, y las dividió en tres tipos de acuerdo a la complejidad de su estructura.
El punto de vista de Brodmann es extraño, en este sentido, porque le cuesta reconocer que estas cortezas son las más antiguas filogenéticamente, y que sería imposible pedirles un período embriológico o fetal de seis capas por más transitorio que éste pudiera llegar a ser. Así que cuando discute a Haller, quien considera que los animales más primitivos tienen corteza de tres capas, no posee toda la razón. En este grupo, la corteza de seis capas está territorialmente muy poco desarrollada, y es la de tres capas la que predomina.
Es criticable también, tomar un estadio determinado del desarrollo intrauterino, para evaluar la ontogenia cortical. Cuanto más temprano es el estadio en que aparecen modificaciones, más importantes serán las mismas para el objetivo de la división cortical (ley ontogenética). Así, Filimonoff4 estima que ya en estadios previos al evaluado por Brodmann que es entre los 6-8 meses, la estructura del telencéfalo muestra diferencias locales en el desarrollo de la placa cortical de His3, que permiten dividir la corteza de forma "simple", no en dos, sino en tres territorios fundamentales. Éstos son: allocórtex, corteza intermedia, e isocórtex. El allocórtex o corteza incompleta (más primitiva), se dividiría en dos tipos, el semicórtex, o corteza no separada, a la que pertenecen el tuberculo olfactorio, la región diagonal, el septum pellucidum, la región periamigdalina y, parcialmente, la corteza prepiriforme; y el archicórtex que incluye, el asta de Ammon, el subículum, la fascia dentata y las tenia tectae.
La corteza intermedia o periallocórtex, incluye el perisemicórtex y el periarchicórtex. La zona perisemicortical, incluye la zona insular, y parcialmente, la prepiriforme. La zona periarchicortical incluye el presubículum, que se extiende también sobre el cuerpo calloso, y la zona entorrinal.
El isocórtex es la corteza de seis capas u homogenética de Brodmann, con las diferencias territoriales por él descriptas.
De esta manera, la neocorteza o isocórtex, se encuentra separada en todo su perímetro del allocórtex, por la corteza intermedia. Cada una de estas cortezas tiene una estructura bien definida que permite su reconocimiento.
La nomenclatura de Brodmann ha sido modificada por otros autores, y la corteza homotípica ha recibido las denominaciones de neocórtex, isocórtex, corteza eulaminada, corteza gris, neopalio, corteza completa, entre otros. Para las cortezas heterogenéticas los sinónimos son arquicórtex, paleo y arquipalio, corteza blanca, corteza defectiva, entre otros.
2. La división topográfica de la corteza cerebral y los mapas
Brodmann es muy honesto en su exposición, cuando dice que hay zonas en que las transiciones son muy netas, y otras, en que son graduales "y se necesita mucha práctica para aprender a diferenciarlas". von Ekonomo2 agrega a esto, que a veces los límites son subjetivos.
2a) Las regiones mayores. A pesar de estas dificultades, hay una disposición topográfica muy característica y repetida a lo largo de la escala, que permite diferenciar sin dudas, una serie de regiones mayores. Esta disposición consiste en la inserción de dos regiones heterotípicas entre regiones homotípicas: un área agranular en la región central del hemisferio (región precentral, áreas 4 y 6), y un área con una capa granular interna muy diferenciada, a veces con subcapas (área 17) en la región occipital (Fig. 11).
Ésta forma una especie de calota en el polo occipital, extendida más hacia lateral o hacia medial, de acuerdo a la especie. Entre estas zonas, se expanden más o menos vastas extensiones de corteza homotípica. La zona que queda por delante de la región precentral (agranular), es la región frontal, de extensión máxima en el hombre, y que va disminuyendo su tamaño, hasta prácticamente desaparecer en los roedores. Lo mismo sucede con las regiones insertas entre ambas áreas heterotípicas, la región parietal, y la región temporal aunque de éstas quedan pequeños representantes incluso entre los insectívoros. En la cara medial, todas estas regiones limitan con la circunvolución límbica y el hipocampo, que contienen tres regiones: la región cingular, la retroesplenial y la hipocámpica, reconocidas por su relación con el cuerpo calloso y la cara medial del temporal. Finalmente, por delante y debajo del hemisferio, se extiende la más primitiva región olfatoria. Estas cortezas heterogenéticas también pueden distinguirse muy bien de las homogenéticas.
Estas grandes regiones se subdividen en áreas, cuyos límites son precisos en las regiones heterotípicas y heterogenéticas, y bastante imprecisos en las homotípicas. Todas ellas forman en su conjunto tanto en el hombre como en otras especies superiores, una calota en el polo frontal (Área 10) y luego, regiones en forma de franjas que se disponen verticalmente en sentido anteroposterior. En la región occipital forman dos franjas que rodean al hemisferio en forma de cinturón por delante del área 17 (18 y 19). Así, un corte horizontal del hemisferio muestra una sucesión de campos "bien reconocibles" en el sentido anteroposterior. Muchas veces, las características estructurales de algunas regiones son tan típicas, que permiten reconocer la especie de que se trata. En la región temporal, las áreas se disponen en franjas paralelas a la cisura de Silvio. Las regiones homotípicas disminuyen francamente su extensión y número de campos, a medida que se desciende en la escala, a la vez que las diferencias entre los campos se van atenuando. Esto también pasa incluso en la región heterotípica agranular precentral, en que desaparece la diferencia entre el área 4 y la 6 en los insectívoros, mientras que las regiones parietal y temporal se confunden en una pequeña zona de la corteza dorsolateral. El área 17 también pierde su complejidad característica, sin perder, sin embargo, su identidad. En estas especies, se desarrolla, por otro lado, una poderosa región olfatoria, heterogenética, donde se diferencian varias áreas que desaparecen en especies superiores, especialmente en los "microsmáticos".
Estas similitudes básicas (homología) aún con sus diferencias entre especies y nivel en la escala, son tan típicas, que, a pesar de esas diferencias, y de las lagunas del material disponible para mantener una continuidad más estrecha en el estudio de la escala, le permitirían a Brodmann afirmar un origen monofilético de la serie de los mamíferos.

Fig. 11. Esquemas de cerebros de tití y de lemur, que muestran la inserción de las áreas heterotípicas (punteado). A las que aparecen en blanco hay que restarles las áreas heterogenéticas (lóbulo olfatorio y región hipocámpica).Así, las áreas homotípicas se agrupan en la región frontal, parietal y temporal. Brodmann tiene más dificultad en aceptar las otras "esferas de los sentidos" como heterotípicas (3, 1, 2 y 41,42).
2b) La organología cortical. Este punto de vista, no es original de Brodmann, sino que tiene una larga historia, que comienza con las teorías de Spurtzheim y Gall, de fines del s XVIII y comienzos del XIX5 y que, aunque totalmente teóricas y comprobadamente erróneas, por difundidas, podrían haber influido en mentes preclaras del s XIX como Broca o Meynert, que con sus observaciones promocionaron la idea de que la corteza podría estar dividida en centros, de estructura anatómica distinta, y diferente función.
Broca en 1861, revolucionó el campo de la neurología al describir su paciente Le Borgne5, quien presentaba un trastorno severo de la producción del lenguaje, que hizo que se lo apodara "Tan" que era el único vocablo que emitía, con una comprensión parcialmente conservada. El paciente tenía una lesión en el pie de la tercera circunvolución frontal izquierda. En 1868 Broca presentó otro paciente con un cuadro muy similar, y una lesión coincidente, lo que comprobaba su teoría de un centro del lenguaje hablado.
Karl Wernicke, en 1874, describió una serie de diez pacientes con un tipo de afasia completamente diferente, con un exceso de producción de un lenguaje incomprensible, "jerga", con numerosas parafasias, tanto fonológicas como semánticas y cargado de neologismos, con defecto severo de la comprensión. Presentaban una lesión en la porción posterior de la primera circunvolución temporal izquierda. A partir de sus observaciones y las de Broca, Wernicke ideó el primer modelo de funcionamiento conjunto de diferentes "centros cerebrales" unidos por fascículos de asociación, para la realización de una función "psicológica"6.
En 1867, Meynert, al observar la distinta estructura microscópica de diferentes zonas de la corteza cerebral, supuso para estas "unidades estructurales" diferentes prestaciones y concibió una visión de toda la corteza cerebral como un "complejo orgánico"2. Por la variedad y profundidad de sus observaciones, se dijo de él que "sólo desde Meynert el cerebro ha tenido alma".
Para 1874 también, las investigaciones sobre los vasos sanguíneos cerebrales realizadas por Duret a instancias de Charcot, lo llevaron a observar que diferentes zonas cerebrales, presentaban en forma constante un vaso que las irrigaba, cuyo origen podía ser diverso, mientras que los territorios se mantenían estables. Esbozó de esta forma, en un trabajo de revisión de 1910, una teoría acerca de "órganos" individuales neurovasculares, (Fig. 12), que él relacionaba especialmente con lóbulos y circunvoluciones7. Mostramos las áreas vasculares del territorio silviano según Salamon8, para que sean comparadas con las de Brodmann.
Estas observaciones se potenciaron cuando para la misma época, Fritsch y Hitzig y algo más tarde Ferrier y Bastian9,10, consiguieron estimular eléctricamente la corteza cerebral en perros, y observaron que la estimulación por delante de la cisura central producía movimientos en la mitad contralateral del cuerpo. Algo después, Munk consiguió producir déficits localizados, mediante ablaciones corticales definidas.
Todas estas observaciones, impactantes, llevaron a los investigadores a buscar una estructura diferenciada para esos distintos "centros" y, si bien en algunos casos existía, como el área precentral (4), en otros, las diferencias eran menos evidentes y "algo voluntarias". Brodmann insiste en que las áreas 44 y 45 "son dignas de formar una región aparte", o una subregión dentro de su Regio Frontalis. Algo más compleja, resulta la posición de las áreas 39 y 40, que requerirían que el área de Wernicke invadiera fuertemente el lóbulo parietal inferior.
Posiblemente por eso, en la interpretación de la función a partir de la estructura, se muestra Brodmann muy prudente y genera una hipótesis muy adelantada para su época, de corte netamente sistémico, aseverando que el tipo de procesamiento realizado por células o regiones es muy elemental, y para cada capacidad mayor de la mente humana se requeriría del trabajo conjunto y simultáneo de muchas de estas regiones o bien del cerebro entero. El resultado funcional, no es el de la suma de los trabajos de cada región, sino la resultante nueva de la labor de conjunto. Admite así que cada región de las que él identificó, puede tomar parte en la expresión de distintas capacidades, adelantándose en décadas a las modernas concepciones. Admite también la posibilidad de cierta regionalización de las estructuras involucradas.
Introducimos unos fragmentos de un trabajo de Patricia Goldman-Rakic11 1988, para mostrar la vigencia de las ideas de Brodmann a través del tiempo: ....la pregunta de cómo organiza el cerebro sus subsistemas, es quizá la más desafiante que se pueda hacer... El cuadro que surge de la nueva anatomía es el de un mecanismo altamente integrado, pero distribuido, cuyos recursos son asignados a varios sistemas básicos paralelos, que conectan todas las subdivisiones mayores del cerebro.

Fig. 12. Áreas corticales de las ramas de la cerebral media según Salamon(8). Se trata, como describiera Brodmann, de una disposición ánteroposterior en franjas, de las áreas. Obsérvese una tendencia a la horizontalidad de las áreas temporales- Si bien esta disposición, no sigue exactamente las áreas citoarquitectónicas, es posible que varias áreas sean irrigadas por una misma rama por razónes del tipo de la conveniencia económica. Por otro lado, una misma región citoarquitectónica podría ser irrigada por más de un vaso, ya que puede participar en la compleción de más de una función.
3. Plasticidad cortical
En la última parte de su capítulo sobre fisiología, se refiere a la restitución espontánea por parte del cerebro, de funciones perdidas por efecto de alguna noxa. Aquí también sus opiniones son notables. Concibe la restitución como la asunción de la función por otra región preservada del efecto de la noxa, y que normalmente toma parte en el trabajo de expresión de esa función. La ejercitación (rehabilitación), sería el mecanismo por el cual el área involucrada se hace cargo de la función perdida. Descarta que se produzca la utilización de una zona indiferente, puesto que áreas que no ejercen un trabajo, se atrofian.
Finalmente, la observación que consignamos más abajo, de "maduración" de las capas II y IV durante las primeras décadas de la vida, podría ser vista como una manifestación de plasticidad puesta al servicio del aprendizaje. La experiencia novedosa podría actuar desencadenando la maduración de elementos esenciales para el reconocimiento de un nuevo estímulo.
4. La posición del hombre
El "zeitgeist".
...los negros de Africa carecen por naturaleza de una sensibilidad que se eleve por encima de lo insignificante. El Sr. Hume desafía a que se le presente un ejemplo de que un negro haya mostrado talento, y afirma que entre los cientos de millares de negros transportados a tierras extrañas, y aunque muchos de ellos hayan obtenido la libertad, no se ha encontrado uno solo que haya imaginado algo grande en el arte, en la ciencia o en cualquiera otra actividad honorable... (Emmanuel Kant. Observaciones sobre el sentimiento de lo bello y lo sublime)12.
La frase del mayor representante y aún superador del Iluminismo13, asintiendo a las observaciones de Hume, no deja lugar a dudas acerca del racismo del hombre europeo en épocas de esclavismo. Al principio del siglo veinte, el hombre blanco europeo ocupaba una posición hegemónica en el mundo. Inglaterra, la primera potencia mundial, era dueña de un imperio "donde nunca se ponía el sol" y del que formaba parte nuestro país, "el quinto dominio inglés"14, "la perla más preciada de la corona británica", padeciendo todas las desventajas y sin gozar casi, de ninguna ventaja de esa secreta (a voces) dependencia. Los adelantos tecnológicos y la industria le daban al hombre europeo una preeminencia sobre todos los otros pueblos. Éstos y otros factores, hicieron que se creara el mito de la diferencia racial, que trató de comprobarse científicamente a partir de media-dos del siglo XIX, y se intensificó en el XX.
Broca y sus mediciones de la capacidad craneana y la reinterpretación de sus datos cuando no concordaban con la preeminencia blanca y del varón, constituyen un clásico muy bien relatado por Gould en su obra "La falsa medida del hombre"15. Se encontrarán allí otros relatos de la historia del racismo o la discriminación, basados en razones científicas.
El origen de las especies de Darwin aparecido en 1859, expandió rápidamente la teoría de la evolución por el mundo y, aparte de las encarnizadas discusiones que generó, puso al hombre en la cima del reino animal (Fig. 13) y dio pie a interpretaciones racistas que acentuaron la de por sí natural disposición humana a esas tendencias.
Este prejuicio ha tenido, sin duda, un efecto sobre los estudios de Brodmann, constituye la parte más oscura de su obra, y lo ha llevado al error, aunque él ha sido siempre prudente y escueto en la emisión de sus opiniones, como corresponde a una mentalidad científica.
Fig 13. El árbol genealógico de los mamíferos según Haeckel (1874).Clasificación utilizada aproximadamente por Brodmann.
4a. Los errores de Brodmann. Es posible que por tratarse los monos de una especie inferior, haya estudiado Brodmann sus cerebros con el prejuicio de que su estructura debía ser menos compleja, como efectivamente lo manifiesta, cuando asevera que tienen menos áreas que el del hombre, es decir, una diferencia no sólo cuantitativa (menos volumen cortical total), sino cualitativa.
Vamos a analizar algunos aspectos nada más de estos fallos, básicamente porque no nos encontramos en condiciones de conocimiento como para hacerlo con todos. Constituyen un relato apasionante, en donde entra en juego la neurocirugía.
El lóbulo frontal y la neurocirugía. El interés por el lóbulo frontal se desató un par de décadas después de la publicación de Brodmann, a partir de las clásicas experiencias de Jakobsen en chimpancés (1935)(16), precedidas por las de Jackson (de su mismo equipo) en otras clases de monos. A estos animales se les realizó una lobectomía prefrontal (incluyendo el área 8, ya que llegaba hasta el surco arcuato) (Fig. 14), y esta operación, cuando era bilateral, les hacía perder la capacidad de enfrentarse a pruebas de respuesta retardada, que requirieran mantener un dato importante en la memoria, para obtener la recompensa en comida. Quien realizaba las lobectomías, era John F. Fulton, neurofisiólogo, fundador unos años después, del Journal of Neurosurgery.
Jakobsen16 observó también que a una de las monas chimpancé que estudiaron, antes de la lobectomía, una de las pruebas le producía un tremendo desagrado, tal que, luego de algunos intentos, Becky (que así se llamaba el chimpancé), parecía enloquecer cuando se le administraba la prueba, entraba en un ataque de ira y berrinche, rodaba sobre sí misma y finalmente orinaba y defecaba. De esta forma, nunca se le pudo administrar la prueba antes de ser operada.
Luego de la resección seriada (separada por unos meses) de ambas regiones prefrontales, el animal aceptaba tranquilamente la administración de la prueba, pero, por supuesto no lo aprobaba, salvo que el lapso no superara unos escasos segundos. Esta observación de la "curación" de lo que Jacobsen llamó una "neurosis experimental" semejante a las producidas por Pavlov mediante condicionamiento, llevó a Egas Moniz a realizar un año después (1936) junto con Almeida Lima, la primera lobotomía frontal.
Estas impactantes observaciones que significaron el premio Nobel para Moniz, hicieron que se investigara en mayor profundidad el lóbulo frontal, incluso hasta nuestros días.
Un punto culminante de esos estudios lo alcanzó otro neurocirujano, Earl Walker en 194017, quien revisó la cartografía del lóbulo frontal realizada por Brodmann, encontrando más áreas de las que éste había descripto, incluso para el hombre: 45, 46, 13 y 14, y modificó bastante la disposición del resto, como se puede apreciar en la figura 15. Hay que aclarar que Walker, si bien utilizó los mismos números que Brodmann, en algunos casos no estaba totalmente seguro acerca de la homología, lo mismo que el mismo Brodmann, acerca de ésta frente a la corteza humana.
Otros mapas han sido publicados, como el de Sanides, y más recientemente, el estudio de Petrides18, que encuentra una total homología entre la corteza humana y la del macaco, existiendo sólo una diferencia cuantitativa (Fig. 16).
Las cortezas son difíciles de diferenciar a veces, y Petrides recurre al mismo artilugio de Brodmann, estableciendo la homología no sólo cito y fibriloarquitectónicamente, sino que se apoya muchas veces en la topografía.
Es interesante destacar en estos estudios, la relación del área 46 con el surco principal de la corteza del mono, cuyo homólogo en el hombre, sería el surco frontal medio, descripto por los anatomistas clásicos19, y luego dejado un poco de lado en las descripciones de la topografía del lóbulo frontal.
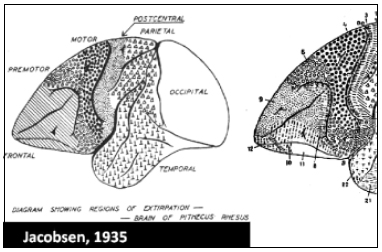
Fig. 14. En rayado, resecciones realizadas por Fulton a dos monas chimpancé, en forma bilateral escalonada para el trabajo de Jakobsen, que fue el primero en demostrar un efecto psíquico y mental de manera experimental. A la derecha, el mapa de Brodmann de la corteza de chimpancé, usado por Fulton, que muestra que se resecó hasta el área 8 inclusive (labio anterior del surco arcuato).

Fig. 15. Mapas de las áreas frontales del mono según Brodmann (derecha) Y según Earl Walker (izquierda). Obsérvese la aparición del área 45, 46, 13 y 14, lo mismo que el desplazamiento del área 10 hacia el polo.

Fig. 16. Áreas del lóbulo frontal en el humano y el mono según Petrides.
4b. El lóbulo parietal inferior. Como para las áreas frontales, Brodmann considera los campos 39 y 40 como exclusivos del ser humano. Sin embargo, ya von Ekonomo había notado la existencia del área PG en el cerebro del mono, mientras que otros consideraron que la corteza de la parte posterior del surco temporal superior era su homólogo20. Aunque la escala de los mamíferos puede ser recorrida en ambas direcciones y de hecho Brodmann lo hace, predomina una franca tendencia en él en hacerlo de arriba hacia abajo, por eso p. ej, puede hablar de "adquisiciones" en la corteza olfatoria del erizo.
4c. Diferencias raciales. Brodmann, siguiendo el ejemplo de Elliot Smith con el cerebro de los egipcios, compara el cerebro del hombre europeo con el de los hotentotes y hereros, que habitaban colonias alemanas de África y con el del javanés, coincidiendo con aquel autor en las diferencias en la estructura de los surcos del lóbulo occipital y su citoarquitectura, que se parecería más al del mono en estas razas, y propone con circunspección y respeto, profundizar estos estudios incluso desde el punto de vista citoarquitectónico.
5. Aspectos grises de la teoría de la localización cerebral: la petición de principio
Brodmann afirma repetidamente que hay veces en que la distinción de un área de otra es difícil, y propone las siguientes soluciones: estar sumamente entrenado, observar imágenes panorámicas a bajo aumento, recordar que las transiciones pueden tener cierta extensión y finalmente si aún no es reconocible, ver de qué parte (topográfica) del cerebro había sido extraída y su relación con zonas vecinas! Es decir p.ej., una zona es parietal, porque se extrajo del lóbulo parietal. Sin esta información, podría ser muy difícil reconocer un campo, sobre todo en regiones homotípicas. Es decir, hay órganos corticales que no son tan definidos como una cierta euforia organológica lo haría creer. Este aspecto del trabajo de Brodmann ha sido duramente criticado por Bailey y von Bonin21. Sin embargo, el hecho de que según estos autores, no se pueda diferenciar una región parietal de una frontal o temporal por su aspecto histológico, no impide que dentro del respectivo lóbulo o región principal, puedan ser detectadas diferentes áreas con caracteres levemente distintos. El hecho de las grandes similitudes debe indicar que el tipo de procesamiento que realizan es parecido, pero su ubicación y conexiones les otorgan una función diferente, que se manifestará necesariamente en diferencias citoarquitectónicas sutiles22. Es quizá por esto, que la labor de Brodmann ha permanecido a través de los años, y otros autores en distintas partes del mundo han comprobado, más tarde, sus observaciones, lo que agiganta sus logros de principios de siglo XX. Presentamos a continuación, algunos de estos mapas, junto al más indiferenciado de Bailey y von Bonin (Figs. 17, 18 y 19).

Fig. 17. Mapa cortical de von Ekonomo y Koskinas2. La obra de estos autores es posiblemente la mayor realizada en este aspecto, por su extensión, minuciosidad y respaldo iconográfico. Es famosa por haber cambiado números por letras, pero su mapa es muy parecido al de Brodmann. Consta de 57 áreas en lugar de 52.

Fig. 18. Mapa de Sarkisov, del instituto neurológico de Moscú. Reproduce aproximadamente las áreas de Brodmann. (Tomado de Petrides17).

Fig.19. Mapa de Bailey y vonBonin()i, (1951). Rojo: koniocortezas. Marrón: áreas yuxtasensoriales. Amarillo, áreas agranulares. Naranja: áreas disgranulares. Rosa: cortezas eulaminadas. Azul: allocortex. Púrpura: yuxtallocortex. Verde: mesocortex . Utilizan los colores para no dar la sensación de separaciones tajantes entre los campos. Los colores compuestos pertenecen a zonas de transición.
6. Otras contribuciones
Brodmann ha realizado el estudio comparado del tamaño de las células, el espesor de las cortezas, la constancia de las capas y otros caracteres generales de la corteza cerebral con minuciosidad.
Así obtuvo algunas conclusiones interesantes: se deben comparar el mismo tipo de células: observó así que las células de Betz más grandes no las posee el hombre, sino los carnívoros, y su tamaño tiene poco o nada que ver con el tamaño del animal, o la longitud del axón.
El espesor de la corteza está en relación con el tamaño del animal, pero no en forma proporcional, por lo que relativamente, el espesor es mayor en los animales más pequeños. También observó que células grandes y en menor número, ocupan cortezas de más espesor y viceversa.
Una observación interesante es la referida a las capas granulares, que según él, son las más variables, tanto que la capa granular interna puede faltar, como en las cortezas agranulares.
La capa granular externa modifica su aspecto con la edad. Mientras que en estadios infantiles y juveniles hay un gran predominio de células granulares "de aspecto neuroblástico", en la edad adulta los gránulos han desaparecido prácticamente, siendo reemplazados por células piramidales pequeñas o medianas, por lo que se diferencia mal de la capa III. ¿Se tratará realmente de células primitivas (gránulos-neuroblastos) que maduran progresivamente hasta pasada la segunda década? Si, como el mismo autor dice, a modificación de la estructura modificación de la función, esta capa podría tener importancia en los fenómenos de memoria y aprendizaje. Otros autores, como von Ekonomo, han corroborado esta observación que a pesar de parecer tan obvia, no genera ningún comentario por parte de Brodmann. También se producen cambios con el tiempo en la capa IV, en donde aparecen células más grandes, frecuentemente estrelladas, entre los gránulos.
CONCLUSIÓN
Estimamos que este modesto homenaje, que ha consistido en hacer conocer la obra de Brodmann en mayor detalle entre los hispanoparlantes, es poco para el gigantesco trabajo que desarrolló este hombre, solo, con técnicas sencillas y que trascendió su época en varias generaciones. Nos ha legado un mapa de la localización de la corteza cerebral que mantuvo su vigencia a través de décadas, por lo que puede considerarse una de las grandes adquisiciones de la humanidad en el siglo XX.
1. Brodmann, K. Vergleichende Lokalizationslehre der grosshirnrinde in ihren Prinzipien dargestellt auf Grund des Zellenbaues. Leipzig, Barth, 1909. [ Links ]
2. von Ekonomo C, Koskinas GN. Die cytoarchitektonik der Hirnrinde des Erwachsenen Menschen. Julius Springer, Wien, 1925. [ Links ]
3. His W. Die Entwickelung des menschlichen Gehirns während der ersten Monate. Leipzig, 1904. [ Links ]
4. Filimonoff, IN. A rational subdivision of the cerebral cortex. Arch Neurol Psychiatry 1947; 58: 296- 311. [ Links ]
5. Greenblatt SH. Phrenology in the Science and Culture of the 19th Century. Neurosurgery, 1995; 37: 790- 805. [ Links ]
6. Wernicke C. Der aphasische Symptomenkomplex. Eine psychologische Studie auf anatomischer Basis. Reprint. Springer, Berlin, 1974. [ Links ]
7. Duret H. Revue critique de quelques recherches récents sur la circulation cérebrale. L'encephale, 1910, 5emeanée, premier semestre: 7- 24. [ Links ]
8. Salamon G, Huang YP. Radiologic anatomy of the brain. Springer, Berlín, 1976. [ Links ]
9. Penfield W, Jasper H. Epilepsy and the functional anatomy of the human brain. Little, Brown & Co., Boston, 1954. [ Links ]
10. Boling W, Olivier A, Fabinyi, G. Historical Contributions to the Modern Understanding of Function in the Central Area. Neurosurgery, 2002; 50: 1296-1310. [ Links ]
11. Kant E. Observaciones sobre el sentimiento de lo bello y lo sublime. Porrúa. México, 1997. [ Links ]
12. Jaspers K. The great philosophers. Volume I. Kant. A Harvest/HBJ Book, Harcourt Brace Jovanovich, San Diego, 1962. [ Links ]
13. Crouzert, M, Director: Historia General de las civilizaciones. Vol X. Destino, Barcelona, 1967. [ Links ]
14. Gould S J. La falsa medida del hombre. Drakontos Bolsillo, Barcelona, 2007. [ Links ]
15. Jakobsen CF, Wolfe JB, Jackson TA. An experimental analysis of the functions of the frontal association areas in primates. J Nerv Ment Dis 1935; 82: 1-14. [ Links ]
16. Walker AE. A cytoarchitectural study of the prefrontal area of the macaque monkey. J Comp Neurol 1940; 73: 59-86. [ Links ]
17. Petrides M, Pandya DN. Comparative architectonic analysis of the human and the macaque frontal cortex. Handbook of Neuropsychology 1994; 9: 17-58. [ Links ]
18. Retzius G. Das Menschenhirn. Studien in der makroskopischen Morphologie. Königliche Buchdruckerei, Stockholm, 1896. [ Links ]
19. Jones EG, Powell TPS. An anatomical study of converging sensory pathways within the cerebral cortex of the monkey. Brain, 1970; 793-820. [ Links ]
20. Bailey P, von Bonin G. The isocortex of man. University of Illinois Press, Urbana, 1951. [ Links ]
21. Felleman, DJ, van Essen, DC. Distributed hierarchical processing in the primate cerebral cortex. Cerebral Cortex 1991; 1: 1-47. [ Links ]

