










Servicios Personalizados
Revista

Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkCiencia del suelo
versión On-line ISSN 1850-2067
Cienc. suelo vol.27 no.1 Ciudad Autónoma de Buenos Aires ene./jun. 2009
TRABAJOS
Caracterización de la meso y macrofauna edáfica en sistemas de cultivo del Sudeste Bonaerense
Antonio H. Gizzi; Héctor A. Álvarez Castillo; Pablo l. Manetti; Alicia N. López*; Natalia L. Clemente & Guillermo A. Studdert
*Unidad Integrada Facultad de Ciencias Agrarias (UNMdP)- EEA INTA Balcarce C.C. 276, (7620) Balcarce, Argentina. Tel. 02266-439100 int. 222 * Correo electrónico: acastillo@balcarce.inta.gov.ar
Recibido: 27-05-08
Aceptado: 30-12-08
RESUMEN
Los objetivos de este estudio fueron: i. determinar la composición taxonómica y la abundancia relativa de la meso y macrofauna y de sus grupos tróficos en suelos del sudeste de la provincia de Buenos Aires, ii. evaluar la densidad de los organismos que integran dichas comunidades y iii. estudiar el efecto de los sistemas de cultivo (combinación entre sistema de labranza y rotación) y la fertilización sobre la meso y macrofauna. Se utilizaron cuatro sistemas de cultivo: LC50 [ (50% pastura - 50% agricultura bajo labranza convencional (LC)] , SD50 [ (50% pastura - 50% agricultura bajo siembra directa (SD)] , LC75 (25% pastura - 75% agricultura bajo LC), SD75 (25% pastura - 75% agricultura bajo SD) y cuatro dosis de fertilización nitrogenada (0, 60, 120 y 180 kg de N ha-1). En el año 2000, en LC50 y en SD50 se sembró una pastura (P), la que en 2001 tenía un año de implantada. En el 2000, en LC75 y en SD75 se sembró maíz y en el 2001, soja. Dentro de la mesofauna, la mayoría de los individuos fueron enquitreidos, que en el primer año no presentaron diferencias entre LC y SD, mientras que en el segundo, los mayores porcentajes se observaron en P cuando ésta tenía más de un año de implantada. En la macrofauna predominaron oligoquetos megadrilos, isópodos y miriápodos y en P en ambos años los isópodos y miriápodos fueron dominantes. En el primer año, las densidades de la meso y macrofauna no mostraron diferencias significativas (p>0,003) entre sistemas de cultivo, sin embargo, fueron mayores en los cultivos bajo SD. En el segundo año, las densidades fueron significativamente mayores (p<0,003) en P. En los dos años, las densidades de detritívoros, herbívoros, predadores y omnívoros no mostraron interacción significativa (p>0,003) entre los sistemas de cultivo y las dosis de fertilización. A pesar de no haberse observado diferencias significativas (p>0,003) entre los sistemas en el primer año, los cuatro grupos fueron más abundantes en P y SD que en LC. En el segundo año, los detritívoros, los predadores y los omnívoros fueron significativamente más abundantes en P que en el cultivo de soja. Los resultados confirmaron que los sistemas de cultivo afectaron la composición taxonómica y la abundancia relativa de la meso y macrofauna y de sus grupos tróficos. Por otro lado, ambas comunidades aumentaron numéricamente durante los períodos bajo P desde su implantación hasta los 15 meses.
Palabras clave. Fauna edáfica; Sistemas de labranza; Rotaciones de cultivos; Composición taxonómica; Grupos tróficos; Densidad.
Characterization of the meso and macrofauna of a Southeastern Buenos Aires province soil under different cropping systems
ABSTRACT
The aim of this work was to determine the taxonomic composition and relative abundance of the soil meso and macrofauna and their different trophic groups as affected by soil management practices. Four cropping systems were evaluated: LC50 [ (50% pasture - 50% cropping under conventional tillage (LC)] , SD50 [ (50% pasture - 50% cropping under no-tillage (SD)] , LC75 (25% pasture - 75% cropping under LC), SD75 (25% pasture - 75% cropping under SD). In the year 2000, a pasture (P) was sown on LC50 and SD50 and in 2001 it was one year old. On LC75 and SD75, corn was planted in 2000 and soybean was planted in 2001. Most of the individuals observed in 2000 within the mesofauna were enchytraeids and their abundance was not different between LC and SD, whereas in 2001 the highest abundance of enchytraeids was observed under the 15-year-old P. Within the macrofauna, earthworms, isopods and miriapods predominated, but isopods and miriapods were dominant under P in both years. In the first year, the meso and macrofauna densities were not significantly different (p>0.003) among cropping systems, although they tended to be greater under SD. In the second year, such densities were significantly higher (p<0.003) under P. In both years, density interactions between cropping systems and fertilization rates were not significant (p>0.003) for detritivours, herbivours, predators and omnivous. Despite the lack of significant differences (p>0.003) among systems during the first year, all four trophic groups were more abundant under P and SD than under LC. In the second year, detritivours, predators, and omnivours were significantly more abundant under P than under soybean. Our results confirm that the cropping systems affected the taxonomic composition of the soil meso and macrofauna and their trophic groups. On the other hand, the abundance of both communities increased under P throughout the experiment.
Key words. Soil fauna; Tillage systems; Crop rotations; Taxonomic composition; Trophic groups; Density.
INTRODUCCIÓN
Durante los últimos años y a nivel mundial, la degradación del recurso suelo ha sido cada vez mayor. De acuerdo a Solbrig (1997), la capacidad productiva de un suelo depende del manejo que el agricultor haga del mismo. Los disturbios producidos por la actividad humana tradi cionalmente producen erosión, pérdida de la materia orgánica y alteran tanto la biodiversidad como las condiciones del ambiente edáfico (Potter & Meyer, 1990).
Consecuentemente, como una medida para controlar la erosión y preservar la calidad del sistema suelo, a comienzos de los '90 los productores de la Región Pampeana comenzaron a adoptar con éxito labranzas más conservacionistas, particularmente la siembra directa (García et al., 2000).
Los sistemas conservacionistas tienden a conservar la estructura del suelo, mejorar la disponibilidad de nutrientes y, debido a la presencia de los rastrojos en superficie, a reducir el efecto de la erosión y la pérdida de agua del perfil (Studdert & Echeverría, 2002). Stinner & House (1990) consideran que en estos sistemas la actividad de los invertebrados del suelo y su interacción con la microflora se beneficia en relación con los sistemas de labranza convencional. La presencia de una cubierta vegetal proporciona un ambiente especial continuo tanto en el espacio como en el tiempo para los organismos del suelo. Estos afectan la descomposición de los residuos vegetales directamente a través de la ingestión, trituración y redistribución de la materia orgánica e indirectamente actúan en la dinámica de los hongos y de las bacterias.
La mesofauna edáfica comprende a los organismos cuyo diámetro corporal varía de de 0,1 a 2,0 mm (Swift et al., 1979) e incluye a los enquitreidos (Annelida: Oligochaeta, Enchytraeidae) y a los microartrópodos (Symphyla, Pauropoda, Acarina, Collembola, Protura y Diplura). Actúan en el ciclo de los nutrientes de manera indirecta a través de la ingestión de hongos y bacterias y en forma directa, fragmentando el material vegetal en descomposición (Conti, 2000). Además algunos integrantes de la mesofauna, como los enquitreidos, debido a su actividad cavadora, son importantes en el mantenimiento de la estructura del suelo (Primavesi, 1982).
La macrofauna comprende a los organismos cuyo diámetro corporal varía entre 2 y 20 mm e incluye principalmente a los Annelida: Oligochaeta: Megadrilli (lombrices) y a los Arthropoda. Mediante un sistema de digestión simbiótica con la microflora del suelo, las lombrices promueven la fertilidad produciendo la mineralización del material orgánico y la movilización de nutrientes necesarios para el crecimiento y producción de las plantas (Lavelle, 1997). Junto a las hormigas (Insecta: Hymenoptera: Formicidae) y a las larvas de los insectos de suelo (Insecta: Coleoptera: Scarabaeidae, Curculionidae, Elateridae, Chrysomelidae), modifican la estructura del suelo mediante la formación de macroporos y agregados (Linden et al., 1994).
En el sudeste bonaerense, son numerosos los estudios sobre cómo los sistemas de cultivo influyen sobre los organismos que viven en la superficie del suelo y que se han realizado utilizando trampas de caída como método de muestreo (Carmona et al., 2002; Carmona et al., 2004; Cicchino, 2003; Marasas et al., 1997; Marasas et al., 2001; Turienzo et al., 2001). Sin embargo, es escasa la información disponible sobre los organismos que viven o cumplen parte de su ciclo de vida dentro del suelo. En consecuencia se hipotetiza que: a) los sistemas de cultivo y la fertilización inciden en la composición taxonómica y en la abundancia relativa de los organismos de las comunidades meso y macrofaunal, b) la densidad de los organismos de la mesofauna y de la macrofauna se incrementa en las pasturas y con la implementación de SD y c) los detritívoros y los predadores son más abundantes en los sistemas con pastura, mientras que los herbívoros lo son bajo cultivo. Se plantean como objetivos: a) determinar la composición taxonómica y la abundancia relativa de la mesofauna y de la macrofauna y de sus grupos tróficos y b) estudiar la densidad de los organismos que integran dichas comunidades y el efecto de los sistemas de cultivo (combinación entre sistema de labranza y rotación) y la fertilización sobre ambas.
MATERIALES Y MÉTODOS
El estudio se realizó durante los ciclos agrícolas 2000/2001 y 2001/2002 en el campo experimental de la Unidad Integrada Balcarce (37°45' S, 58°18' W, 130 msnm) en un ensayo de rotaciones mixtas (agricultura-pastura) iniciado en 1976 sobre un complejo de suelos de Argiudol Típico y Paleudol Petrocálcico sin riesgo de erosión (2% de pendiente) y con textura superficial franca y elevado contenido de Materia Orgánica (53,7±1,7 g MO kg-1 suelo en otoño de 2000 de 0-20 cm promedio de las unidades experimentales empleadas para este trabajo, Eiza et al., 2006). Hasta 1993 se implementaron rotaciones mixtas bajo labranza convencional (LC), alternando ciclos agrícolas y de pasturas con diferente duración. En 1994 se incorporó la SD como variable y quedaron definidos distintos sistemas de cultivo surgidos de la combinación de LC y SD y varias rotaciones de cultivos de cosecha y pasturas. Para esta experiencia se utilizaron sólo cuatro (Tabla 1) de los sistemas de cultivo definidos en el ensayo: LC50 (50% del tiempo con agricultura-50% del tiempo con pastura todo bajo LC), SD50 (50% del tiempo con agricultura-50% del tiempo con pastura todo bajo SD); LC75 (75% del tiempo con agricultura- 25% del tiempo con pastura todo bajo LC) y SD75 (75% del tiempo con agricultura-25% del tiempo con pastura todo bajo SD). La mezcla de especies utilizada para las pasturas estuvo constituida por pasto ovillo (Dactilis glomerata L.), trébol rojo (Trifolium pratense L.), raigrás perenne (Lolium perenne L.) y trébol blanco (Trifolium repens L.). Durante los ciclos agrícolas se empleó la secuencia de cultivos maíz (Zea mays L.) - soja (Glycine max L. Merr.) - trigo (Triticum aestivum L.). El diseño experimental del ensayo es en bloques completos aleatorizados con arreglo de tratamientos en parcelas divididas con tres repeticiones. A las parcelas principales se asignaron los sistemas de cultivo (Tabla 1) y a las subparcelas se asignaron distintas dosis de fertilización con N: 0, 60, 120 y 180 kg N ha-1.
Tabla 1. Sistemas de cultivo en las parcelas principales del ensayo. Las columnas en negrita indican los años analizados.
Table 1. Cropping systems in the main experimental plots. The columns with bold characters indicate the analyzed years.
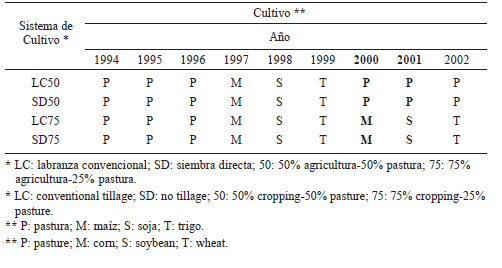
Debido a que la re-definición de la estructura del experimento de largo plazo fue realizada en 1994, hasta el ciclo agrícola 1999/ 2000, los cuatro tratamientos empleados tuvieron la misma historia reciente de manejo (Tabla 1). En el otoño de 2000 correspondió la siembra de pastura bajo LC y SD en las parcelas principales correspondientes a los sistemas de cultivo LC50 y SD50 (Tabla 1), respectivamente. En primavera de 2001 dicha pastura tenía algo más de un año de implantada (Tabla 1). Por otro lado, en la primavera de 2000 correspondió la siembra de maíz bajo LC y SD en las parcelas principales correspondientes a los sistemas de cultivo LC75 y SD75 (Tabla 1), respectivamente, mientras que en la primavera de 2001 correspondió la siembra de soja en las mismas parcelas principales. En el primer año de este estudio (2000/01) se comparó la implantación de las pasturas bajo LC (PLC) y bajo SD (PSD) en los tratamientos LC50 y SD50, respectivamente, con el cultivo de maíz bajo LC (MLC) y bajo SD (MSD) en los tratamientos LC75 y SD75, respectivamente. En el segundo año (2001/02), se compararon las parcelas bajo PLC y PSD (pasturas de algo más de un año de implantadas) en los tratamientos LC50 y SD50, respectivamente, con el cultivo de soja bajo LC (SLC) y bajo SD (SSD) en los tratamientos LC75 y SD75, respectivamente.
En el primer año la extracción de las muestras se realizó durante la última semana de diciembre y la primera de enero y en el segundo año, durante la segunda y tercera semana de enero.
En cada subparcela (17,5 x 7,5 m) se tomó una muestra de suelo al azar, de 25 cm de diámetro por 30 cm de profundidad. Las muestras fueron tamizadas a través de dos mallas de 10 y 2,5 mm, respectivamente. Los elementos groseros y restos vegetales se extrajeron con el tamiz de malla mayor. Luego de tamizado, el suelo quedó lo suficientemente desagregado, extendiéndose sobre bandejas blancas (0,38 m x 0,28 m x 0,06 m) una capa fina de suelo y de ella se extrajeron manualmente los ejemplares («handsorting») (Blair et al., 1996). Los ejemplares extraídos fueron conservados en formol 5% o alcohol 70% para su determinación.
Los organismos de diferentes taxa se agruparon en grupos tróficos teniendo en cuenta la clasificación de Moore et al. (1988): detritívoros, herbívoros, predadores y omnívoros. Los detritívoros se alimentan de restos biológicos en descomposición. El grupo de los herbívoros que se alimentan de vegetales, está compuesto por organismos que, de acuerdo al grado de daño que ocasionan a los cultivos, son considerados plagas. Algunos, como los miriápodos diplópodos (milpiés) y los crustáceos isópodos (bichos bolita) se alimentan de las raíces de las plantas, pero también de restos biológicos en descomposición. Por esta razón, Moore et al. (1988) los considera omnívoros para diferenciarlos de los herbívoros fitófagos que ocasionan daños de significancia económica. Finalmente, los predadores actúan como reguladores naturales de las poblaciones de los herbívoros predando o parasitando sus distintos estados de desarrollo.
Dado que en el segundo año algunos coleópteros no pudieron ser identificados, no fueron incluídos en los grupos tróficos. Por este motivo, la fauna total (meso + macrofauna) del segundo año es numéricamente diferente a la suma de las densidades de los grupos tróficos.
Se calculó la proporción de los organismos de la mesofauna y macrofauna como la relación entre el total de cada taxón y el total de los organismos de la meso o macrofauna, según corresponda. La densidad de los individuos por metro cuadrado se analizó mediante modelos lineales generalizados asumiendo una distribución binomial negativa (α=0,05) con el software SAS V8 (SAS Institute, 2001). Este tipo de distribución se utiliza cuando la varianza de los datos es mayor a dos veces la media (Díaz & Demetrio, 1998). En el caso de detectarse diferencias significativas, los tratamientos se compararon mediante contrastes tipo Bonferroni (McCullagh & Nelder, 1989).
RESULTADOS
En el primer año, los enquitreidos representaron entre el 83 y el 96%. En el segundo año, en cambio, los enquitreidos constituyeron entre el 45 y el 79%, observándose los mayores porcentajes en las pasturas con más de un año de implantadas (Fig. 1).

Figura 1. Composición taxonómica de la mesofauna bajo diferentes sistemas de cultivo en dos ciclos agrícolas. PLC= pastura en labranza convencional, PSD= pastura en siembra directa, MLC= maíz en labranza convencional, MSD= maíz en siembra directa, SLC= soja en labranza convencional y SSD= soja en siembra directa.
Figure 1. Taxonomic compositionof soil mesofauna under different cropping systems in two years. PLC= pasture under conventional tillage, PSD= pasture under no tillage, MLC= corn under conventional tillage, MSD= corn under no tillage, SLC= soybean under conventional tillage and SSD= soybean under no tillage.
La proporción de arañas fue mayor en el segundo año que en el primero (11-23% y 2-4%, respectivamente), hallándose la mayor abundancia relativa en SSD. Los ácaros tuvieron un aporte relativo bajo en la mesofauna en ambos años, y el aporte mayor se observó en PLC. En el primer año los colémbolos se presentaron en mayor porcentaje en las pasturas que en el cultivo de maíz (10% y 2%, respectivamente), mientras que en el segundo se comportaron a la inversa (8-16% y 26-33% para las pasturas y los cultivos de soja, respectivamente) (Fig. 1).
Dentro de la macrofauna se hallaron lombrices (Annelida: Oligochaeta: Megadrilli), estados inmaduros de Coleoptera: carábidos (Carabidae), insectos de suelo (Scarabaeidae, Curculionidae, Elateridae y Chrysomelidae), y adultos de estafilínidos (Staphylinidae). También se encontraron dos especies de Isopoda: el bicho bolita, Armadillidium vulgare Latreille, 1804 y la cochinilla de la humedad, Porcellio scaber Latreille, 1804 y miriápodos quilópodos (ciempiés) y diplópodos (milpiés) (Fig. 2).

Figura 2. Composición taxonómica de la macrofauna bajo diferentes sistemas de cultivo en dos ciclos agrícolas. PLC= pastura en labranza convencional, PSD= pastura en siembra directa, MLC= maíz en labranza convencional, MSD= maíz en siembra directa, SLC= soja en labranza convencional y SSD= soja en siembra directa.
Figure 2. Taxonomic composition soil macrofauna under different cropping systems in two years. PLC= pasture under conventional tillage, PSD= pasture under no tillage, MLC= corn under conventional tillage, MSD= corn under no tillage, SLC= soybean under conventional tillage and SSD= soybean under no tillage.
La composición taxonómica de la macrofauna fue similar en las pasturas en ambos años y los grupos dominantes fueron isópodos y miriápodos. Sin embargo, en maíz fue similar entre sistemas de labranza y distinta en soja (Fig. 2). Los oligoquetos megadrilos, las larvas de carábidos y el complejo de insectos de suelo, contribuyeron con porcentajes menores a la composición macrofaunal en la mayoría de los casos.
En MSD y MLC los grupos dominantes fueron los miriápodos (51 y 55%, respectivamente) y los insectos de suelo (15 y 16%, respectivamente). En el segundo año, en SLC predominaron las lombrices (39%) y en SSD los miriápodos (36%). Las larvas de los carábidos en los sistemas con cultivo tuvieron porcentajes similares entre años, variando entre 4 y 15%. En cambio, los isópodos aparecieron con distinta proporción entre labranzas en el primer año e igual en el segundo con 8%. La composición porcentual de los estafilínidos, fue variable en ambos años, tanto bajo SD como bajo LC (Fig. 2).
Las densidades de la mesofauna y la macrofauna y del total de la fauna edáfica (meso + macrofauna) no mostraron interacción significativa entre los sistemas de cultivo y las dosis de fertilización nitrogenada en los dos años de estudio (P >0,003). No hubo diferencias significativas en las densidades de la mesofauna y la macrofauna y del total de la fauna edáfica entre las dosis de fertilización nitrogenada (P >0,003).
En el primer año, las densidades de la mesofauna y de la macrofauna no presentaron diferencias significativas entre sistemas de cultivo (P >0,003) (Tabla 2). En el segundo año, las densidades de la mesofauna, de la macrofauna y de la fauna total fueron significativamente mayores (P <0,003) en los sistemas con pasturas. Por otro lado, en SSD la densidad de la macrofauna fue significativamente mayor que en SLC. Por otro lado, las labranzas no afectaron a la mesofauna que estuvo constituida mayormente por enquitreidos.
Tabla 2. Densidad (individuos m-2) de la mesofauna, de la macrofauna y del total de la fauna (meso + macrofauna) edáfica bajo distintos sistemas de cultivos. PLC= pastura bajo labranza convencional, PSD= pastura bajo siembra directa, MLC= maíz bajo labranza convencional, MSD= maíz bajo siembra directa, SLC= soja bajo labranza convencional y SSD= soja bajo siembra directa.
Table 2. Density (organism m-2) of soil meso, macro and total fauna (meso + macro) under different cropping systems in two years. PLC= pasture under conventional tillage, PSD= pasture under no tillage, MLC= corn under conventional tillage, MSD= corn under no tillage, SLC= soybean under conventional tillage and SSD= soybean under no tillage.

Se observó que la densidad de la mesofauna y el total de la fauna edáfica de un año a otro fue diferente en las unidades experimentales con cultivos. La densidad de la macrofauna disminuyó en los lotes agrícolas bajo LC, pero se modificó levemente en los lotes bajo SD. En las pasturas, las densidades de la mesofauna y de la macrofauna y del total de la fauna edáfica aumentaron en el segundo año (Tabla 2).
Los detritívoros hallados en este trabajo fueron lombrices megadrilos, enquitreidos, ácaros (Acarina: Cryptostigmata) y colémbolos (Arthropleona: Onychiuridae); los herbívoros (fitófagos sensu, Moore et al., 1988) larvas de insecto de suelo (Coleoptera: Scarabaeidae, Elateridae, Chrysomelidae y Curculionidae): Diloboderus abderus (Sturm, 1826), Cyclocephala signaticollis (Burmeister, 1847), Phillochloenia bonaeriensis (Bruch, 1909), Conoderus scalaris (Germar, 1824, Colaspis bridarollii (Bechyné, 1949) y curculiónidos; los predadores: arañas, quilópodos (Geophilomorpha, Lithobiomorpha y Scolopendromorpha), larvas de carábidos y adultos de estafilínidos y los omnívoros A. vulgare, P. scaber y diplópodos (Juliformia).
La densidad de los grupos tróficos, en los dos años no mostró interacción significativa entre los sistemas y las dosis de fertilización nitrogenada (P >0,003). Tampoco se encontraron diferencias significativas entre las dosis de fertilización (P >0,003). A pesar de que en el primer año no se encontraron diferencias significativas (P>0,003) entre los sistemas en las densidades de los grupos tróficos, en las pasturas y en SD, en general, los cuatro grupos fueron más abundantes que en LC (Tabla 3).
Tabla 3. Densidad (individuos m-2) de los grupos tróficos bajo distintos sistemas de cultivo en dos ciclos agrícolas. PLC= pastura en labranza convencional, PSD= pastura en siembra directa, MLC= maíz en labranza convencional, MSD= maíz en siembra directa, SLC= soja en labranza convencional y SSD= soja en siembra directa.
Table 3. Density (organism m-2) of trophic groups under different cropping systems in two years. PLC= pasture under conventional tillage, PSD= pasture under no tillage, MLC= corn under conventional tillage, MSD= corn under no tillage, SLC= soybean under conventional tillage and SSD= soybean under no tillage.

En el segundo año se observó el efecto de la rotación y en consecuencia, las densidades de los detritívoros, los predadores y los omnívoros fueron significativamente mayores en las pasturas que en los cultivos independientemente de la labranza. En las pasturas, la densidad de los detritívoros fue aproximadamente 10 veces mayor que en los cultivos agrícolas. En los dos años, los herbívoros no presentaron diferencias significativas (P >0,003) entre los tratamientos y, en general, sus densidades fueron bajas. Los predadores tuvieron densidades mayores en las pasturas con respecto a SLC, mientras que SSD no presentó diferencias significativas con las pasturas (Tabla 3).
DISCUSIÓN
La composición taxonómica de la mesofauna y de la macrofauna fue similar a la informada para el sudeste bonaerense por Vincini et al. (2004) y por López et al. (2005). Dentro de los organismos de la mesofauna, los enquitreidos, fueron los más abundantes a pesar de los disturbios producidos por las labranzas (López et al., 2005). En los sistemas bajo LC las labores incorporan los residuos vegetales e incrementan la probabilidad de supervivencia de estos organismos (Lägerlof et al., 1989). Además, como son organismos muy pequeños, permanecen en los agregados del suelo, a pesar de los disturbios producidos por las labranzas. En cambio, en el maíz y la pastura bajo SD, los residuos vegetales que se localizan en la superficie del suelo, estimulan el crecimiento de los hongos, que sirven de alimento a los enquitreidos micrófagos (Hendrix et al., 1986).
Dentro de los microartrópodos, los ácaros se encontraron en los sistemas con y sin disturbios, coincidiendo con los resultados informados por López et al. (2005).
En general, en ambos años, los microartrópodos tuvieron un aporte relativamente bajo en la mesofauna. En contraste con estos resultados, Gómez et al. (1996) hallaron que, en suelos franco-arenosos de la zona semiárida de la provincia de Buenos Aires, la mesofauna estuvo constituída principalmente por microartrópodos, de los cuales aproximadamente el 90% fueron ácaros y colémbolos. Aparentemente los ciclos agrícolas prolongados tuvieron un efecto negativo mayor sobre los enquitreidos que sobre los ácaros, los colémbolos y las arañas, ya que éstos aumentaron su abundancia relativa en el segundo año.
Dentro de la macrofauna, en las pasturas tanto en el primero como en el segundo año los grupos dominantes fueron los isópodos y los miriápodos. Por el contrario, López et al. (2000) reportaron que en las pasturas, los insectos de suelo y los megadrilos (lombrices), fueron los organismos dominantes y que en conjunto representaron el 67% del total de la macrofauna. Posiblemente, en este trabajo influyó la edad de las pasturas, que fue de sólo tres meses en el primer año y de 15 meses en el segundo, pudiendo considerarse que en el primer año estaban en implantación y no se manifestó el efecto del sistema pastura sobre la fauna edáfica. La densidad de algunas especies del complejo insectos de suelo aumenta cuando las pasturas tienen varios años (Aragón, 1998; Ribeiro, 2000). Por otra parte, las diferencias observadas en la composición taxonómica de la macrofauna pueden asociarse a la fecha de los muestreos, fin de diciembre y principio de enero, que es el período en el cual las larvas de los insectos de suelo son escasas (Álvarez Castillo et al., 1993; Lanteri, 1994; Ribeiro, 2000).
Las densidades mayores de la mesofauna y de la macrofauna estuvieron asociadas a los sistemas con menor grado de disturbio, como las pasturas y la SD. Ello se debe a que las pasturas producen un ambiente con temperatura y humedad adecuada y una biomasa radicular abundante que proporciona mayor disponibilidad de alimento y refugios (Primavesi, 1982). Sin embargo, en este estudio, este efecto sólo se observó en el segundo año. En el primer año las pasturas recientemente implantadas, provenían de una rotación agrícola (maíz, soja, trigo) y la fauna del suelo se encontraba en un período de recuperación de los disturbios producidos durante el ciclo agrícola.
La densidad de los detritívoros en las pasturas fue mucho mayor que en los cultivos agrícolas. Algunos estudios realizados comparando lotes con rotación agrícola y pasturas de varios años muestran que en las pasturas la densidad de detritívoros es mayor que en los cultivos (Stinner & House, 1990; Bedano et al., 2000; Clemente et al., 2003). Aparentemente en las pasturas, la biomasa que retorna al suelo, principal recurso alimenticio de los detritívoros facilita el aumento numérico de estos organismos (Wardle et al., 1999). Por el contrario, Marasas et al. (2001) encontraron, un porcentaje mayor de detritívoros en cultivos agrícolas con respecto a pastizales naturales y argumentaron que estos sistemas disponen de mejor calidad y cantidad de materia orgánica.
En general, en los dos años de estudio, los herbívoros no fueron abundantes y en concordancia con Robertson et al. (1994), las prácticas agrícolas no afectaron a los herbívoros fitófagos. Esto fue debido a que los ejemplares recolectados en las muestras fueron insectos de suelo que encuentran sus recursos alimenticios y refugios tanto en los cultivos como en los sistemas bajo pasturas. Por otra parte, los insectos de suelo que predominaron fueron los coleópteros curculiónidos en coincidencia con los resultados reportados por Lanteri (1994), Aragón (1998) y Ríos de Saluso et al. (2001).
Los predadores fueron más abundantes en SD y pasturas, si bien en el segundo año en PLC hubo un número mayor de predadores, a pesar de las labores agrícolas efectuadas. Esto se pudo deber a que los carábidos hallados pertenecieron a especies de los géneros Scarites anthracinus y Notiobia cupripennis que poseen hábitos cavadores y realizan galerías hacia el interior del suelo protegiéndolos de la acción de las labores.
CONCLUSIONES
Bajo las condiciones en que se desarrolló esta experiencia, los sistemas de cultivo (combinación entre sistemas de labranzas y rotaciones de cultivos de cosecha y pasturas) afectaron la composición taxonómica y la abundancia relativa tanto de la mesofauna como de la macrofauna. Cuando se continuó con la agricultura, la mesofauna y la macrofauna disminuyeron con distinta intensidad según el sistema de labranza. La fertilización nitrogenada no tuvo efecto sobre las densidades de la meso, macrofauna y de los grupos tróficos. Los detritívoros, los omnívoros y los predadores fueron más abundantes en las pasturas de 15 meses que en los cultivos, mientras que la densidad de los herbívoros fue similar en todos los sistemas. Las pasturas fueron los sistemas que permitieron el desarrollo de los detritívoros y omnívoros, los cuales intervienen en la mineralización de la materia orgánica y en el reciclaje de los nutrientes.
REFERENCIAS
1. Álvarez Castillo, HA; AN López; AM Vincini; DM Carmona& PL Manetti. 1993. Relevamiento de insectos del suelo en cultivos de papa del sudeste bonaerense. CERBAS INTA. Boletín Técnico 118, 18 pp. [ Links ]
2. Aragón, JR. 1998. Plagas animales del trigo y su manejo integrado. Pp 6-9. En: Actas IV Congreso nacional de trigo. II Simposio nacional de cereales de siembra otoño - invernal. Mar del Plata, 11-13 de noviembre de 1998. [ Links ]
3. Bedano, JC; M. Cantu & ME Doucet. 2000. La influencia de distintos sistemas productivos en las comunidades de fauna edáfica en agroecosistemas del sur de Córdoba. En: CD, 4 pp en Actas XVII Congreso Argentino de la Ciencia del Suelo. Mar del Plata, 11 al 14 de abril de 2000. [ Links ]
4. Blair, JM; PJ Bohlen & DW Freckman. 1996. Soil invertebrates as indicators of soil quality. Pp. 273-291. In: JW Doran & AJ Jones (eds). Methods for assessing soil quality. SSSA Spec. Public. 49. Soil Sci. Soc. Am., Inc. Madison, Wisconsin, USA. [ Links ]
5. Carmona, D; AM Vincini; PL Manetti; AN López & HA Álvarez Castillo. 2002. Densidad activa estacional de insectos plaga y predadores edáficos en agroecosistemas con siembra directa y agricultura convencional. En: V Congreso Argentino de Entomología. Buenos Aires. Museo Argentino de Ciencias Naturales «Bernardino Rivadavia». 18-22, p 245. [ Links ]
6. Carmona, D; P Manetti; A Vincini; A López; HA Álvarez Castillo & M Sorensem. 2004. Abundancia, riqueza y dominancia específica de Carábidos (Insecta: Coleoptera) en agroecosistemas bajo siembra directa y manejo convencional. En: II Reunión Binacional de Ecología. XXI Reunión Argentina de Ecología. Mendoza, p 98. [ Links ]
7. Cicchino, AC; M Marasas & MF Paleologos. 2003. Características e importancia de la carabidofauna edáfica de un cultivo experimental de trigo y sus bordes con vegetación espontánea en el partido de La Plata, provincia de Buenos Aires. Revista Ciencia y Tecnología. Serie Científica: 41-53. [ Links ]
8. Clemente, NL; AN López; AM Vincini; HA Álvarez Castillo; DM Carmona; PL Manetti & S San Martino. 2003. Abundancia de megadrilos (Annelida: Oligochaeta) en diferentes sistemas de producción. Ci. Suelo 21: 35-43. [ Links ]
9. Conti, M. 2000. Principios de edafología, con énfasis en suelos argentinos. Ed. Facultad de Agronomía 2º. ed. 430 p. [ Links ]
10. Díaz, MP & CGB. Demétrio. 1998. Introducción a los Modelos Lineales Generalizados. Su aplicación en las Ciencias Biológicas. Ed. Screen. 105 pp. [ Links ]
11. Eiza, M.; GA Studdert & GF Domínguez. 2006. Dinámica de la materia orgánica del suelo bajo rotaciones mixtas: I. Materia orgánica total. En: CD, 5 pp. En: Actas XX Congreso Argentino de la Ciencia del Suelo. Salta-Jujuy, 19 al 22 de septiembre de 2006. [ Links ]
12. García, FO; M Ambrioggio & V Trucco. 2000. No-tillage in the Pampas of Argentina: A Success store. Better Crops Int. 14: 24-27. [ Links ]
13. Gómez, M; H Krüger & M Sagardoy. 1996. Actividad biológica en un suelo de la zona semiárida bonaerense cultivado con la secuencia soja - trigo bajo tres prácticas culturales. Ci. Suelo 14(1): 37-41. [ Links ]
14. Hendrix, PF; RW Parmelee; DA Crossley; DC Coleman; EP Odum & PM Groffman. 1986. Detritus food webs in conventional and no-tillage agroecosystems. Bio-Science 36: 374-380. [ Links ]
15. Lagerlöf, J; O Andren & K Paustian. 1989. Dinamics and contribution to carbon flows of enchytreidae (Oligochaeta) under four cropping systems. J. Appl. Ecol. 26: 183-199. [ Links ]
16. Lanteri, AA. 1994. Bases para el control integrado de los gorgojos de la alfalfa. De La Campana (ed.). La Plata, Bs. As. 119 pp. [ Links ]
17. Lavelle, P. 1997. Faunal Activities and Soil Processes: Adaptative Strategies That Determine Ecosystem Function. Adv. Ecologycal Res. 27: 93-132. [ Links ]
18. Linden, DR; PF Hendrix; DC Coleman & PCJ van Vliet. 1994. Faunal indicators of soil quality. Pp 91-106. In: JW Doran; DC Coleman; DF Bezdicek & BA Stewart (eds.) Defining soil quality for a sustainable environment. SSSA Special Publication Nº 35. Soil Sci. Soc. Am., Inc.; Am. Soc. Agron., Inc. Madison, Wisconsin, USA [ Links ]
19. López, AN; AM Vincini; DM Carmona; HA Álvarez Castillo& PL Manetti. 2000. Impacto de las prácticas de manejo sobre la fauna edáfica en dos sistemas de producción. En: CD, 4 pp en Actas XVII Congreso Argentino de la Ciencia del Suelo. Mar del Plata, 11 al 14 de abril de 2000. [ Links ]
20. López, AN; AM Vincini; NL Clemente; PL Manetti; DM Carmona & HA Álvarez Castillo. 2005. Densidad estacional y distribución vertical de los Enchytraeidae (Annelida: Oligochaeta) en diferentes sistemas de producción. Ci. Suelo 23: 115-121. [ Links ]
21. Marasas, M.; SJ Sarandon & AC Cicchino. 1997. Efecto de la labranza sobre la colepterofauna edáfica en un cultivo de trigo en la provincia de Buenos Aires (Argentina). Ci. Suelo 15: 59-63. [ Links ]
22. Marasas, ME; SJ Sarandon & AC Cicchino. 2001. Changes in soil arthropod functional group in a wheat crop under conventional and no tillage systems in Argentina. Applied Soil Ecology 18: 61-68. [ Links ]
23. McCullagh, P & JA Nelder. 1989. Generalizaed linear models. Chapman & Hall, Cambridge, Great Britain. 541 pp. [ Links ]
24. Moore, JC & DE Walter.1988. Arthropod regulatio of micro-and mesobiota in below-ground detrital food webs. Ann. Rev. Entomol. 33: 419-439. [ Links ]
25. Primavesi, A. 1982. Manejo ecológico del suelo. Ed. El Ateneo, Bs. As. 5ta edic. [ Links ]
26. Potter, CS & RE Meyer. 1990. The role of biodiversity in sustainable dryland farming systems. Adv. Soil Sci.13: 241-251. [ Links ]
27. Ribeiro, A. 2000. Gorgojos del suelo. Rev. de la EEMAC. Cangüé. 19: 22-25. [ Links ]
28. Ríos de Saluso, MLA; A Saluso; O Paparotti & C Debona. 2001. Asociación entre la macrofauna del suelo y la siembra directa en el oeste de Entre Ríos. Pp 83-95. En: JL Panigatti; D Buschiazzo & H Marelli (eds). Siembra Directa II. Ed. INTA. [ Links ]
29. Robertson, LN; BA Kettle & GB Simpson. 1994. The influence of tillage practices on soil macrofauna in a semi-arid agroecosystem in northeastern Australia. Agric. Ecosyst. Environ. 48: 149-156. [ Links ]
30. SAS INSTITUTE. 2001. SAS/STAT software release 8.02. Cary, USA, SAS Institute Inc. [ Links ]
31. Solbrig, OT. 1997. Sembrando el futuro. Como desarrollar una agricultura para conservar la tierra y la comunidad. En: 5º Congreso Nacional de AAPPRESID, Mar del Plata. [ Links ]
32. Stinner, BR & GJ House. 1990. Arthropods and other invertebrates in conservation - tillage agriculture. Ann. Rev. Entomol. 35: 299-318. [ Links ]
33. Studdert, GA & HE Echeverría. 2002. Soja, girasol y maíz en los sistemas de cultivo del sudeste bonaerense. Pp. 413-443 en Andrade, FH & V Sadras (eds) Bases para el manejo del maíz, el girasol y la soja (2ª ed.). INTA - Fac. Cs. Agrarias (UNMP). Balcarce. [ Links ]
34. Swift, M; O Heal; & J Anderson. 1979. The composition in Terrestrial Ecosystems. Blackwell Scientific, Oxford, UK. 373 p. [ Links ]
35. Turienzo, P; D Carmona; G Studdert; AM Vincini; AN López; PL Manetti & HA Álvarez Castillo. 2001. La siembra directa en el manejo sustentable de los suelos: conservando los carábidos en agroecosistemas. En: I Reunión Binacional de Ecología. XX Reunión Argentina de Ecología: Argentina- Chile. San Carlos de Bariloche. p 232. [ Links ]
36. Vincini, A; P Manetti; D Carmona; A López; HA Álvarez Castillo & M Sorensen. 2004. Abundancia poblacional de crustáceos (Isopoda: Oniscoidea) en cultivos de cereales bajo siembra directa y convencional en el sudeste bonaerense. P. 121. En: II Reunión Binacional de Ecología. XXI Reunión Argentina de Ecología. Mendoza. 31 octubre al 5 noviembre de 2004. [ Links ]
37. Wardle, DA; KS Nicholson; KI Bonner & GW Yeates. 1999. Effects of agricultural intensification on soil associated arthropod population dynamics, community structure, diversity and temporal variability over seven-year period. Soil Biol. Biochem. 31(12): 1691-1706. [ Links ]

