










Servicios Personalizados
Revista

Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkCiencia del suelo
versión On-line ISSN 1850-2067
Cienc. suelo vol.35 no.2 Ciudad Autónoma de Buenos Aires dic. 2017
BIOLOGÍA DEL SUELO
Fósforo del suelo: calidad del rastrojo y descomposición microbiana bajo diferentes contenidos hídricos
Dannae Lilia Serri1; Valeria Soledad Faggioli2 & Claudio Antonio Lorenzon2
1. Instituto de Patología Vegetal, Centro de Investigaciones Agropecuarias
2. EEA INTA Marcos Juárez
* Autor de contacto: serri.dannae@inta.gob.ar
Recibido: 07-04-17
Recibido con revisiones: 08-06-17
Aceptado: 08-06-17
RESUMEN
Los residuos de los cultivos y rastrojos son un importante recurso para los agroecosistemas. Sin embargo, aún hay escasa información sobre procesos de liberación e inmovilización microbiana de nutrientes en relación a la calidad del material vegetal y las condiciones del medio. El objetivo de este trabajo fue evaluar el efecto de la calidad de residuos vegetales y contenidos de humedad edáfica sobre procesos bioquímicos relacionados a la dinámica del fósforo (P) en el suelo. Se realizó un experimento en condiciones controladas con los siguientes tratamientos: a) Rastrojo: Testigo, sin residuo vegetal (Te); residuo de Vicia villosa (V); residuo de Triticum aestivum L. (T) y V. T, correspondiente a la mezcla de los dos últimos; y b) Humedad del suelo: capacidad de campo (CC), 75% CC y punto de marchitez permanente (PMP). A los 10, 20, 40 y 60 días de iniciado el experimento se midió en el rastrojo la materia seca (MS) remanente, P (PR), C:N (C:NR) y C:P (C:PR). En el suelo se cuantificó: C:N(C:NS), respiración microbiana (RM), hidrólisis de diacetato de fluoresceína (FDA), fosfatasa ácida (FA), P (PS), P de biomasa microbiana (PBM), C de biomasa microbiana (CBM). Los resultados demostraron que, aunque el contenido hídrico del suelo afectó la descomposición del material vegetal, la calidad del rastrojo tuvo mayor influencia sobre la actividad de los microorganismos. En general, la mayor actividad biológica se observó en CC. Independientemente del contenido hídrico, la presencia de rastrojo en el suelo incrementó RM y FDA; FA y PS fueron mayores cuando se adicionó V. El CBM asimiló el P liberado, generando una inmovilización a los 20 días, seguido de una mineralización. Los resultados demuestran la importancia de la calidad del material vegetal en mayor medida que la humedad del suelo sobre las funciones biológicas relacionadas a la descomposición del residuo y dinámica del fósforo edáfico.
Palabras clave: Calidad del rastrojo; Actividad microbiana; Mineralización e inmovilización de fósforo.
Soil phosphorus: Stubble quality and microbial decomposition under contrasting water contents
ABSTRACT
Crop residues and stubble are important agroecosystem resources, but there is scarce information on the processes of microbial nutrient release and immobilization related to plant material quality and environmental conditions. The objective of this work was to evaluate the effect of different soil moisture contents and plant residue quality on biochemical processes related to soil phosphorus (P) dynamics. A bioassay was performed and the treatments were: a) Crop residues: Control, without plant residue (Te); Vicia villosa residue (V); Triticum aestivum L. residue (T) and V. T, corresponding to the combination of V and T; and b) Soil moisture: field capacity (CC), 75% CC and wilting point (PMP). After 10, 20, 40 and 60 days of the experiment we measured MS (dry matter), PR (P of residue), C:NR (C:N of residue) and C:PR (C:P of residue) in plant residue. In the soil we quantified: C:N (C:NS), microbial respiration (RM), fluorescein diacetate hydrolysis (FDA), phosphatase acid (FA), P (PS), microbial biomass P (PBM) and microbial biomass C (CBM). The results showed that although in the course of the experiment the soil water content affected plant material decomposition, the stubble quality had a greater influence on the microorganism activity. In general, the greatest decomposition and biological activity was observed in CC. Regardless of the water content, the incorporation of plant tissues in soil increased FDA and RM; and FA and PS were higher when V was added. The released P was assimilated by CBM at 20 days, but was followed by a mineralization phase. These results allowed to assert the importance of the plant material quality on the biochemical functions related to the stubble decomposition and crop residues, and soil phosphorus dynamics.
Keywords: Stubble quality; Microbial activity; Phosphorus mineralization and phosphorus immobilization.
INTRODUCCIÓN
La optimización de la eficiencia del uso del fósforo (P) es una de las metas perseguidas en los agroecosistemas. Para ello es necesario contar con un conocimiento adecuado de la dinámica de las reservas edáficas de P y las necesidades de aportes externos para un crecimiento óptimo de los cultivos. Si bien nuestra comprensión sobre P inorgánico del suelo es relativamente amplia, el P devuelto al suelo en los residuos del cultivo ha sido menos estudiado. Los residuos de los cultivos de cobertura y luego de la cosecha de granos pueden presentar cantidades significativas de P además de contribuir a mejorar la retención de agua y condiciones físicas del suelo, y proporcionar energía para los procesos biológicos (Lal, 2005; Damon et al., 2014).
El aprovechamiento de P de los rastrojos depende de procesos bioquímicos que generan un continuum entre el rastrojo y la solución de suelo cuyo punto de interacción es la microbiota edáfica. La biomasa microbiana juega un papel clave en el proceso de transformación bioquímica del P orgánico a su forma inorgánica, disponible para la absorción por las raíces de las plantas (Stewart & Tiessen, 1987). Enzimas microbianas,tales como la fosfatasa cobran gran importancia en la nutrición fosforada de las plantas ya que la mayor parte del P existente en el suelo se encuentra en formas orgánicas y por ello representa el mecanismo bioquímico más importante en la mineralización de P del suelo (Wick et al., 1998).
Entre los factores que controlan las tasas de descomposición y mineralización se encuentra la calidad o composición química de los materiales vegetales (Johnson et al., 2005).Se sabe que la composición química dirige el sentido de los procesos biológicos (Kwabiah et al., 2003), y varía de acuerdo a la especie vegetal (Palm & Sánchez, 1991) y su estado fenológico (Kachaka et al., 1993). Aportes de P agronómicamente significativos podrían estar siendo subestimados en los actuales sistemas de producción, principalmente a través del uso de cultivos de cobertura de fabáceas (Varela et al., 2014). En contraste, los rastrojos de cereales al contar con una baja concentración de P-debido a la re-translocación de P al grano- pueden reducir la disponibilidad de P del suelo debido a la asimilación en la biomasa microbiana (Damon et al., 2014).
En suelos sin limitaciones físicas o químicas, su contenido hídrico es vital puesto que el agua interviene en la movilidad y disponibilidad de los diferentes elementos del medio (Coyne, 1999). Reducciones del nivel de humedad afectan la disponibilidad de P ya que su tasa de disolución es proporcional a la cantidad de agua (Thompson & Troeh, 2002). A su vez, el agua constituye un factor determinante en los procesos biológicos puesto que la mayoría de ellos son oxidativos, pero necesitan del agua para mantener el equilibrio osmótico y funcionamiento de las células microbianas (Halverson et al., 2000). Según Coyne (1999) el nivel óptimo es de -0,1atm, es decir, capacidad de campo. En consecuencia, la dinámica de P dependerá directamente de la interacción de tres componentes clave: la biomasa y actividad microbiana, las propiedades del material vegetal y las condiciones edáficas.
En un contexto productivo donde tanto el P como el agua pueden ser limitantes, el conocimiento de un potencial aporte o inmovilización vinculado al material vegetal remanente y condiciones hídricas deben ser considerados al momento de diagramar una estrategia de fertilización y/o rotación. El objetivo de este trabajo fue evaluar el efecto de la calidad de residuos vegetales y contenidos de humedad edáfica sobre procesos bioquímicos relacionados a la dinámica del fósforo (P) en el suelo.
MATERIALES Y MÉTODOS
El experimento se realizó con suelo Argiudol típico franco limoso, proveniente de un lote con historial agrícola de INTA EEA Marcos Juárez, Córdoba (INTA, 1978). Se evaluaron los siguientes tratamientos de calidad de rastrojos: Testigo, sin aplicación de residuos (Te); Vicia, Vicia villosa (V); Trigo, Triticumaestivum L. (T) y Vicia. Trigo (V.T). Dichos tratamientos se sometieron a diferentes escenarios de contenido hídrico del suelo: capacidad de campo (CC), 75% CC y punto de marchitez permanente (PMP). Se utilizó un diseño completamente aleatorizado con arreglo factorial de 3 repeticiones.
Preparación del experimento
Se extrajo suelo de 0-20 cm (MO: 3,08%; P-Bray1: 39,5 ppm; NO3: 113 ppm; pH (1:2,5) 5,6; CE (1:2,5): 0,14 mS/cm 25 ºC). El suelo se secó a temperatura ambiente, se tamizó (2 mm) y se colocaron 150 g en recipientes plásticos de 375 cc (10*15*2,5 cm). El tratamiento de humedad se aplicó ajustando por peso cada 24 h la cantidad de agua destilada necesaria para llegar a CC (0,3 g H2O g-1 suelo), 75% CC (0,225 g H3O g-1 suelo) y PMP (0,126 g H2O g-1 suelo). Según el respectivo tratamiento, a cada recipiente se le adicionó 4,5 g de residuos de T, V o V.T cortado en fragmentos de 1,5 cm de longitud y se mezcló homogéneamente con el suelo. El residuo de V provenía de plantas en estado de floración (C:N 12:1), mientras que T consistió del material remanente luego de la cosecha del cultivo (C:N 43:1). El experimento se mantuvo a 25 ºC durante 60 días.
Muestreo
La extracción de las muestras se realizó a los 10, 20, 40 y 60 días. El procesamiento consistió en el tamizado de la totalidad del suelo contenido en cada bandeja por un tamiz de 2 mm de malla para separar el suelo del material vegetal. Se secaron aproximadamente 10 g de suelo en estufa a 105 ºC para las determinaciones químicas y estimación del contenido de humedad para corrección de las variables biológicas (expresadas en base seca). El suelo restante se conservó en bolsas plásticas cerradas a 4 ºC hasta la realización de las determinaciones biológicas durante el lapso de una semana. El material vegetal contenido en la malla del tamiz se separó del suelo mediante enjuagues con agua destilada y se secó en estufa a 60 °C para estimar su tasa de descomposición por variaciones en el peso seco.
Determinaciones
Sobre las muestras de rastrojo se midió: materia seca (MS) remanente, donde la tasa de descomposición se cuantificó por peso seco del material vegetal contenido en tamiz; C y N total del rastrojo (C:NR) por combustión seca con analizador automático LECO Truspec (no se contó con repeticiones debido a cuestiones operativas),concentración de P(PR) por calcinación a 500 °C (Fernández al., 2009) y colorimetría de Murphy & Riley (1962), y relación C:P del rastrojo mediante las técnicas antes mencionadas. Sobre las muestras de suelo secas se analizó C y N total del suelo (C:NS) por combustión seca con analizador automático LECO Truspec y concentración de P (PS) extraído con 0,5 M NaHCO3 pH 8,5 y colorimetría de Murphy & Riley (1962).Sobre las muestras de suelo fresco se determinó: la actividad enzimática de la hidrólisis del diacetato de fluoresceína (FDA) y fosfatasa ácida (FA) utilizando las técnicas de Alef (1995); respiración microbiana (RM) por incubación durante 7 días y C de biomasa microbiana (CBM) por el método de fumigación-incubación, ambas técnicas de Jenkinson & Polwlson (1976) y P de biomasa microbiana (PBM) según Brookes et al. (1982).
Análisis estadísticos
Los resultados se analizaron mediante análisis de la varianza por modelos lineales generales y mixtos (considerando heterogeneidad de variancias para los muestreos) y la comparación de medias se estimó a través del test LSD (p<0,05) con el programa Statistical Analysis System (SAS Institute, 1990). La relación entre variables se estimó a través del análisis de correlación de Pearson (p<0,05) para el cual se utilizó el programa estadístico InfoStat (Di Rienzo et al., 2015).
RESULTADOS
Descomposición del material vegetal
El contenido de humedad del suelo y la calidad del rastrojo afectaron la descomposición del material vegetal. Se observó que la situación hídrica óptima para la actividad microbiana fue CC (Fig 1a). En CC el rastrojo remanente al finalizar el experimento llegó a valores cercanos a 3% del material inicial en V y V.T, mientras que T registró un remanente cuatro veces mayor.
La relación C:NR y C:PR se ordenó de la siguiente manera: T > V.T > V, siendo sus valores iniciales de C:NR de 43:1, 21:1 y 12:1, y C:PR de 479:1, 206:1 y 113:1 respectivamente. Se observó para T una relación C:NR superior a 25:1 (umbral teórico de inmovilización - mineralización) durante el tiempo del ensayo, en tanto que V y V.T mantuvieron una relación menor (Fig 1b). La evolución de la relación C:NS para los distintos tratamientos expresó para T, V.T y Te una relación C:NS superior a 10:1, mientras que V se mantuvo por debajo de dicho valor a lo largo del tiempo de ensayo (Fig 1c).

Figura 1. a) Evolución de la materia seca (MS) remanente del rastrojo (%); b) Evolución de la relación C:NR y c) Evolución de la relación C:NS, para un suelo incubado con rastrojos de trigo (T), vicia (V), vicia×trigo (V.T) y testigo sin material vegetal (Te), en diferentes intervalos de tiempo. De izquierda a derecha resultados en suelo con humedad gravimétrica de capacidad de campo (CC), 75% CC y punto de marchitez permanente (PMP).
Figure 1. a) Dry matter (DM) remaining stubble (%) evolution;b) C:NR ratio evolution and c) C:NS ratio evolution. Soil incubated with wheat (T), hairy vetch (V), hairy vetch plus wheat (V. T) and control without plant material (Te), at different time intervals. From left to right results in soil with gravimetric moisture of field capacity (CC), 75% CC and permanent wilting point (PMP).
Se registró una correlación altamente significativa y positiva para las variables C:NR y C:NS (Fig 2). Se registró para V C:NR <25 y C:NS <10, para T C:NR >25 y C:NS >10 y V.T obtuvo valores intermedios. En la Figura 2, la línea de puntos perpendicular al eje ‘‘x’’ corresponde a la relación C:NS 10:1 necesaria para el funcionamiento de hongos y bacterias (Kaye & Hart, 1997), en el eje ‘‘y’’ la línea corresponde a la relación C:NR 25:1 umbral teórico de inmovilización-mineralización (Palma & Segat, 2000).
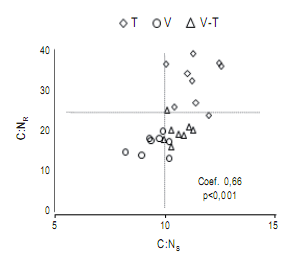
Figura 2. Correlación significativa entre las variables C:NR y C:NS para un suelo incubado con rastrojos de trigo (T), vicia (V) y vicia×trigo (V.T). Se presentan el p-valor y el índice de correlación de Pearson.
Figure 2. Significant correlation between C:NR and C:NS in a soil incubated with wheat (T), hairy vetch (V), hairy vetch plus wheat (V.T). p-value and Pearson coefficient are shown.
Actividad microbiana del suelo
La adición de tejidos vegetales al suelo afectó significativamente la RM en los tres contenidos de humedad estudiados (Fig 3a). Los residuos de T y V.T registraron las medias más altas de CO2, seguidos de V y Te. El análisis estadístico registró los valores máximos de RM de cada rastrojo a los 20 días.
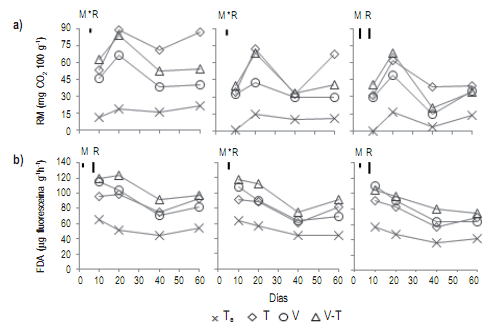
Figura 3.a) Evolución de la respiración microbiana (RM: mg CO2 100 g-1) y b) Evolución de la hidrólisis de diacetato de fluoresceína (FDA: μg fluoresceína g-1h-1), de un suelo incubado con rastrojos de trigo (T), vicia (V), vicia×trigo (V.T) y testigo sin material vegetal (Te), en diferentes intervalos de tiempo. De izquierda a derecha resultados en suelo con humedad gravimétrica de capacidad de campo (CC), 75% CC y punto de marchitez permanente (PMP). Las barras representan la diferencia mínima significativa (LSD p<0,05) de los factores: momento de muestreo (M), tipo de rastrojo (R) o su interacción (M*R), según corresponda.
Figure 3. a) Microbial respiration (RM: mg CO2 100 g-1) evolution, and b) FDA activity (FDA: μg fluoresceína g-1h-1) evolution. Soil incubated with wheat (T), hairy vetch (V), hairy vetch plus wheat (V.T) and control without plant material (Te), at different time intervals. From left to right results in soil with gravimetric moisture of field capacity (CC), 75% CC and permanent wilting point (PMP). Bars represent the least significant difference (LSD p<0.05) of the factors: sampling time (M), stubble type (R) or their interaction (M*R), as appropriate.
En general el contenido hídrico del suelo afectó la RM (Fig 3a), siendo el CO2 liberado superior en CC para todos los residuos. El promedio de RM durante el tiempo de estudio para T a CC fue 31,4% y 42,9% superior a la observada para el mismo residuo en 75% CC y PMP, respectivamente.
La actividad microbiana estimada por FDA registró los valores de medias más altos durante los primeros 10 y 20 días de iniciado el ensayo (Fig 3b), y fue más alta en el suelo con residuos vegetales, en el orden V.T > V > T > Te. Además, el contenido de humedad del suelo afectó la FDA. Para CC, mostró diferencias significativas entre los tratamientos, siendo V.T superior y registrando en promedio para los 60 días del ensayo un 50% más que Te.La actividad enzimática FA fue superior para los tratamientos con residuos vegetales. Estos mantuvieron la misma actividad a lo largo del tiempo como así también dentro de cada uno de los muestreos (V > V.T > T > Te) (Fig 4a). El contenido hídrico del suelo no afectó significativamente la actividad FA, aunque a 75% CC se observaron los valores de medias más altos. Independientemente de esto, los rangos medios de cada rastrojo durante todo el ensayo indicaron que V registró un 10,3%, 21,7% y 32% más actividad FA que V.T, T y Te respectivamente. Los muestreos registraron diferencias significativas, observándose a los 20 días de iniciado el ensayo los mayores valores para los tratamientos en todas las humedades.

Figura 4. a) Evolución de la actividad fosfatasa ácida (FA: μg pnf g-1h-1); b) Evolución del P remanente del rastrojo (PR remanente: mg P) y c) Evolución del P del suelo (PS: μg P g-1), de un suelo incubado con rastrojos de trigo (T), vicia (V), vicia×trigo (V.T) y testigo sin material vegetal (Te), en diferentes intervalos de tiempo. De izquierda a derecha resultados en suelo con humedad gravimétrica de capacidad de campo (CC), 75% CC y punto de marchitez permanente (PMP). Las barras representan la diferencia mínima significativa (LSD p<0,05) de los factores: momento de muestreo (M), tipo de rastrojo (R) o su interacción (M*R), según corresponda.
Figure 4. a) Phosphatase activity (FA: μg pnf g-1h-1) evolution; b) Stubble P (PR remaining: mg P) evolution, and c) Soil P (PS: μg P g-1) evolution. Soil incubated with wheat (T), hairy vetch (V), hairy vetch plus wheat (V.T) and control without plant material (Te), at different time intervals. From left to right results in soil with gravimetric moisture of field capacity (CC), 75% CC and permanent wilting point (PMP). Bars represent the least significant difference (LSD p <0.05) of the factors: sampling time (M), stubble type (R) or their interaction (M*R), as appropriate.
Liberación neta de fósforo
El PR remanente, estimado a partir del contenido inicial de P del residuo (V: 9,72; T: 6,62 y V.T: 3,51 mg P), registró una fase inicial de rápida liberación de P en los primeros 20 días de ensayo, seguido de una fase estable hasta la finalización del mismo (Fig 4b). En CC se registró una liberación de PR en V del 89%, en V.T del 80% y para T del 52%. En la medida que disminuyó el contenido hídrico del suelo, el PR liberado fue menor.
El PS fue diferente entre tratamientos a lo largo de todo el período de incubación. Se observó que PS fue superior significativamente para V respecto de V.T,T y Te, lo que se registró en todos los contenidos hídricos(Fig 4c). A los 20 días se obtuvieron los valores más bajos de PS para los residuos en todas las humedades; mientras que los más altos fueron a los 60 días.
La correlación hallada para las variables FA y PS fue significativa y positiva (Fig 5). Se observó que los tratamientos con residuos se ubicaron de forma descendente en el orden V > V.T > T > Te, mostrando una clara separación entre los mismos.

Figura 5. Correlación significativa entre las variables FA (μg pnf g-1 h-1) y PS (μg P g-1) para un suelo incubado con rastrojos de trigo (T), vicia (V), vicia×trigo (V.T) y testigo sin material vegetal (Te). Se presentan el p-valor y el índice de correlación de Pearson.
Figure 5. Significant correlation between FA (μg pnf g-1 h-1) and PS (μg P g-1) in a soil incubated with wheat (T), hairy vetch (V), hairy vetch plus wheat (V.T) and control without plant material (Te). p-value and Pearson coefficient are shown.
Inmovilización neta de fósforo
El CBM registró diferencias significativas en los tratamientos de rastrojos y muestreos para la humedad CC (Fig 6). Paraésta, los residuos de V y V.T presentaron medias superiores con diferencias significativas respecto de T y Te. A los 20 días del ensayo se registró el CBM más bajo para todos los residuos. Además, evaluando el CBM en conjunto con la MS remanente se pudo apreciar una correlación altamente significativa y negativa (Coef. -0,8; p-valor 0,001).
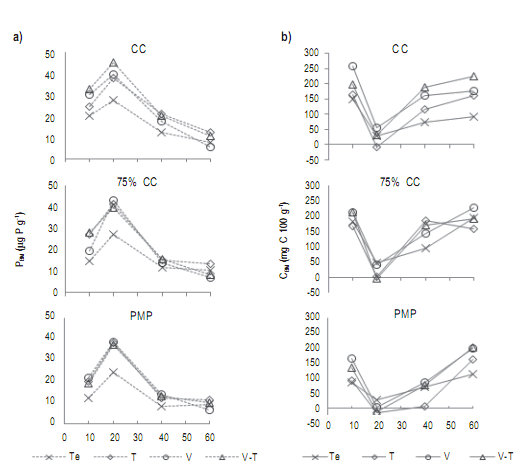
Figura 6. a) Evolución del P de biomasa microbiana (PBM: μg P g-1); b) Contenido de C de biomasa microbiana (CBM: mg C 100 g-1), de un suelo incubado con rastrojos de trigo (T), vicia (V), vicia×trigo (V.T) y testigo sin material vegetal (Te), en diferentes intervalos de tiempo. Suelo con humedad gravimétrica de capacidad de campo (CC), 75% CC y punto de marchitez permanente (PMP). Las barras representan la diferencia mínima significativa (LSD p<0,05) de los factores: momento de muestreo (M), tipo de rastrojo (R) o su interacción (M*R), según corresponda.
Figure 6. a) Microbial biomass P (PBM: μg P g-1) evolution; b) C content in microbial biomass (CBM: mg C 100 g-1). Soil incubated with wheat (T), hairy vetch (V), hairy vetch plus wheat (V.T) and control without plant material (Te), at different time intervals. Soil with gravimetric moisture of field capacity (CC), 75% CC and permanent wilting point (PMP). Bars represent the least significant difference (LSD p <0.05) of the factors: sampling time (M), stubble type (R) or their interaction (M*R), as appropriate.
El PBM registró los mayores valores para los tratamientos con residuos, mientras que los niveles más bajos se observaron en Te (Fig 6). La evolución en el tiempo de los niveles de PBM de los tratamientos fue similar para todas las humedades, alcanzando a los 20 días los mayores valores, seguidos de una tendencia decreciente hasta finalizar el ensayo. A los 60 días V registró el menor PBM con 6,2 µg P g-1, en promedio para todos los tratamientos de humedad del suelo.
DISCUSIÓN
La biomasa microbiana ha sido definida por algunos autores como el ojo de la aguja que todo material orgánico del suelo debe atravesar para que los nutrientes aportados estén disponibles para las plantas (He, 1997). En nuestro estudio se observó que independientemente del tratamiento de rastrojos, hubo mayor CBM en suelos mejores provistos de agua (CC y 75%CC; Fig 6), es decir, que hubo una respuesta de la biota del suelo al ambiente edáfico acorde a lo reportado por Butterly et al. (2009). Del mismo modo, Banu et al. (2004) hallaron una fuerte correlación positiva entre el CBM y el contenido de humedad del suelo. El tratamiento sin incorporación de material vegetal (Te) mostró los menores valores de CBM (Fig 6). Resultados hallados por Sanaullah et al. (2011) estudiando el efecto de la humedad en comunidades vegetales de diferente composición sobre la biomasa microbiana, observaron que ésta siempre se incrementó cuando el suelo estuvo con vegetación cualquiera haya sido el tratamiento de humedad. La correlación negativa hallada entre las variables MS del rastrojo y CBM, indicaría que en la medida que se descompuso el rastrojo incorporado al suelo se produjo el aumento del CBM, es decir, el crecimiento de las comunidades microbianas. Este hecho fue explicado por Goldman et al. (1987) como el resultado de un crecimiento microbiano balanceado debido al suministro apropiado de nutrientes. Stark et al. (2007) estudiando la influencia de las enmiendas orgánicas sobre las propiedades microbianas del suelo, encontraron que la adición de una fabácea (Lupinus sp) al suelo estimuló el crecimiento y la actividad de la comunidad microbiana simultáneamente.
La actividad biológica es fundamental para la solubilización, movilización y disponibilidad de los nutrientes (Borie et al., 1999). La mineralización comprende dos procesos: uno dirigido por enzimas según la demanda del medio; y el otro, es la oxidación de hidratos de C por los microorganismos del suelo para proveerse de energía con la subsecuente liberación de nutrientes contenidos en los compuestos oxidados (McGill & Cole, 1981). El pico de actividad microbiana observado en V, V.T y T (Figs 3a, 3b y 4a), podría atribuirse a la estimulación originada por el agregado de material vegetal (Martens et al., 1992; Bandick & Dick, 1999; Sánchez Monedero et al., 2008); en cambio en Te correspondería a la mineralización de compuestos orgánicos del suelo (Tabatabai, 1994). Se ha comprobado que la eficiencia microbiana (i.e. la fracción de C que asignan para crecer) disminuye cerca de 100 veces con incrementos de las relaciones C:N y C:P de los sustratos (Goldman et al., 1987). Esto explica la baja descomposición observada en T y lo opuesto en V (Fig 1a). Además, elevadas relaciones C:N:P pueden generar oxidaciones de los compuestos orgánicos del suelo para satisfacer la demanda microbiana y mineralización de nutrientes (Goldman et al., 1987). En el ensayo, se observó que los residuos con mayor relación C:N aumentaron el C:NS (Fig 1b y 1c). Esto fue confirmado por la correlación altamente significativa y positiva registrada entre las variables C:NR y C:NS (Fig 2), demostrando un importante efecto de la calidad del rastrojo sobre la relación C:N del suelo. La relación C:N tiene efectos directos sobre los procesos bioquímicos del suelo puesto que a mayor C:N predominan procesos de inmovilización mientras que si aumenta ocurre lo opuesto (Damon et al., 2014).
Dado que la fosfatasa ácida es una exoenzima, puede permanecer retenida en los sitios de intercambio y, en condiciones de baja concentración de P en la solución del suelo, libera fosfatos de los complejos orgánicos para equilibrar los niveles de P edáfico (Tabatabai, 1982). Este hecho quedó demostrado en la correlación positiva entre FA y PS (Fig 5) y pone de manifiesto la importancia de la enzima en el ciclo del P y en el mantenimiento de niveles de P biodisponible del suelo (Wanner, 1996; Quiquampoix & Mousain, 2005). Podría inferirse, entonces, que la cuantificación de las fosfatasas sería un complemento a los diagnósticos de fertilidad fosforada convencionales que sólo contemplan el P extractable. Más aún en sistemas con cultivos de cobertura leguminosos por el potencial aporte de P del residuo vegetal (Kwabiah et al., 2003).
Una vez en la solución del suelo los iones fosfato pueden ser inmovilizados por la biomasa microbiana según la oferta de nutrientes del medio (Kwabiah et al., 2003). En este estudio, los iones fosfato liberados del material vegetal (disminución de P en los tejidos) no permanecieron en la solución de suelo como para ser detectados a través de PS sino que fueron asimilados por la biomasa microbiana (PBM). Esto pudo observarse a los 20 días de iniciado el ensayo, período en el cual se registraron los valores más bajos de PS (Fig 4c) y más altos de PBM (Fig 6). Como se mencionó anteriormente para ese momento, se registró la mayor pérdida de MS y PR coincidente con el aumento de la actividad microbiana (RM, FDA y FA). Resultados semejantes han sido descriptos por Bünemann et al. (2012), quienes al agregar diferentes tipos de sustratos también observaron elevada respiración microbiana del suelo que no se correlacionó con pulsos en la liberación de fosfatos a la solución suelo.
Nuestros resultados sugieren que luego de un corto período luego del aporte de material vegetal (20 días), los procesos microbiológicos se exacerban en detrimento de la disponibilidad de P del suelo, con un predominio de la inmovilización de P. Al transcurrir el tiempo, los tratamientos con residuos de mayor calidad pueden propiciar la liberación de P a la solución de suelo, especialmente en situación de buena oferta de humedad edáfica.
CONCLUSIONES
La calidad del material vegetal y la humedad gravimétrica del suelo afectaron en diferente medida a las funciones biológicas relacionadas a la descomposición del rastrojo y dinámica del fósforo edáfico. En este sentido, la composición química del material vegetal fue más importante que la humedad del suelo en los procesos metabólicos de descomposición. El residuo de vicia (fabácea) fue el que generó mayor actividad fosfatasa, mayor liberación de P al suelo y perdió mayor proporción de P de sus tejidos en todas las condiciones de humedad estudiadas.
CONSIDERACIONES FINALES
El aporte del fósforo removilizado por el residuo del cultivo de vicia y el consecuente incremento de los niveles de fósforo extractable debido a las funciones biológicas del suelo podría representar una mejora en el reabastecimiento de fósforo biodisponible del suelo y la reducción de las necesidades de fertilización del cultivo siguiente gracias a la rápida mineralización de este residuo vegetal. Para ello se sugiere profundizar los estudios a campo sobre la relación entre la actividad de las fosfatasas ácidas y el P disponible del suelo bajo diferentes condiciones de aporte de rastrojos (rotaciones) y regímenes de disponibilidad hídrica.
BIBLIOGRAFÍA
1. Alef, K. 1995. Estimation of microbial activities. In: Alef, K & P Nannipieri (eds). Methods in Applied Soil Microbiology and Biochemistry. Ac. Press. London. Pp. 193-270. [ Links ]
2. Bandick, AK & RP Dick. 1999. Field management effects on soil enzyme activities. Soil Biol. Biochem. 31: 1471-1479. [ Links ]
3. Banu, NA; B Singh & L Copeland. 2004. Soil microbial biomass and microbial biodiversity in some soils from New South Wales, Australia. Aust J. of Soil Res. 42: 777-782. [ Links ]
4. Borie, G; S Aguilera & P Peirano.1999. Actividad biológica en suelos. Frontera Agrícola (Chile) 5: 29-32. [ Links ]
5. Brookes, PC; PS Powlson & DS Jenkinson. 1982. Measurement of microbial biomass phosphorus in soil. Soil Biol. Biochem. 14: 319-329. [ Links ]
6. Bünemann, EK; O Oberson; F Liebisch; F Keller; KE Annaheim; O Huguenin-Elie& E Frossard. 2012. Rapid microbial phosphorus immobilization dominates gross phosphorus fluxes in a grassland soil with low inorganic phosphorus availability. Soil Biol. Biochem. 51: 84-95. [ Links ]
7. Butterly, CR; EK Bünemann; AM McNeill; JA Baldock& P Marschner. 2009. Carbon pulses are related to decreases in microbial biomass during repeated drying and rewetting of soils. Soil Biol. Biochem. 41(7): 1406-1416. [ Links ]
8. Coyne, MS. 1999. Soil Microbiology: an exploratory approach. 1ra edn.Delmar Publishers. 462 pp. [ Links ]
9. Damon, PM; B Bowden; T Rose & Z Rengel.2014. Crop residue contributions to phosphorus pools in agricultural soils: A review. SoilBiol.Biochem. 74: 127-137. [ Links ]
10. Di Rienzo, JA; F Casanoves; MG Balzarini; L Gonzalez; M Tablada & CW Robledo. InfoStat Versión 2015. Grupo InfoStat, FCA, UNC, Argentina. URL http://www.infostat.com.ar. [ Links ]
11. Fernández, MC; H Belinque; FH Gutierrez Boem & G Rubio. 2009. Compared phosphorus efficiency in soybean, sunflower and maize. J. Plant Nutr. 32(12): 2027-2043. [ Links ]
12. Goldman, JC; DA Caron & MR Dennett.1987. Regulation of gross growth efficiency and ammonium regeneration in bacteria by substrate C:N ratio. Limnol.Oceanogr. 32(6): 1239-1252. [ Links ]
13. Halverson, LJ; TM Jones & MK Firestone. 2000. Release of intracellular solutes by four soil bacteria exposed to dilution stress. Soil Sci. Soc. Am. J. 64: 1630-1637. [ Links ]
14. He, Z. 1997. Turnover of soil microbial biomass and its relation to nutrient cycling in agricultural system: A review. Tura 29: 61-69. [ Links ]
15. INTA - Secretaría de Agricultura y Ganadería de la Nación. 1978. Carta de suelos de la República Argentina. Hoja 3363-17, Marcos Juárez. [ Links ]
16. Jenkinson, DS & DS Powlson. 1976. The effect of biocidal treatments on metabolism in soil. I. Fumigation whit chloroform. Soil Biol. Biochem. 8: 167-177. [ Links ]
17. Johnson, JM; NW Barbour & S LachnichtWeyers. 2005. Chemical composition of crop biomass impacts its decomposition. Soil Sci. Soc. Am. J. 71:155-162. [ Links ]
18. Kachaka, S; B Vanlauwe& R Merckx. 1993. Decomposition and nitrogen mineralization of pruning of different quality. In: Mulongoy, K. & R Merckx. (eds). Soil organic matter dynamics and sustainability of tropical agriculture.Wiler, Chichester. Pp. 199-208. [ Links ]
19. Kaye, JP & SC Hart. 1997. Competition for nitrogen between plants and soil microorganisms. Trends Ecol. Evol. 12(4): 139-143. [ Links ]
20. Kwabiah, AB; CA Palm; NC Stoskopf& RP Voroney. 2003. Response of soil microbial biomass dynamics to quality of plant materials with emphasis on P availability. Soil Biol. Biochem. 35(2): 207-216. [ Links ]
21. Lal, R. 2005. World crop residues production and implications of its use as a biofuel.Environ. Int. 31(4): 575-584. [ Links ]
22. Martens, DA; JB Johason& WT Frankenbergenr, Jr. 1992.Production and persistence of soil enzymes with repeated addition of organic residues.Soil Science 153(1): 53-61. [ Links ]
23. McGill, WB & CV Cole. 1981. Comparative aspects of cycling of organic C, N, S and P through soil organic matter. Geoderma 26: 267-286. [ Links ]
24. Murphy, J & JP Riley. 1962. A modified single solution method for the determination of phosphate in natural waters. Anal.Chim.Acta 27: 31-36. [ Links ]
25. Palm, CA & PA Sánchez. 1991. Nitrogen release from leaves of some tropical legumes as affected by their lignin and polyphenolic contents. Soil Biol. Biochem. 23: 83-88. [ Links ]
26. Palma, RM & AL Segat. 2000. Nitrógeno del suelo. En: Conti, M. (ed). Principios de Edafología. Facultad de Agronomía. 2da edn. 430 pp. [ Links ]
27. Quiquampoix H & D Mousain. 2005. Enzymatic hydrolysis of organic phosphorus. In: Turner BL; Frossard E & DS Baldwin (eds). Organic phosphorous in the environment. CABI, Wallingford, pp 89-112. [ Links ]
28. Sanaullah, M; E Blagodatskayac; AChabbia; C Rumpela& Y Kuzyakovc. 2011. Drought effects on microbial biomass and enzyme activities in the rhizosphere of grasses depend on plant community composition. Appl. Soil Ecol. 48: 38-44. [ Links ]
29. Sánchez Monedero, MA; C Mondini; ML Cayuela; A Roig; M Contin & M De Nobili. 2008. Fluorescein diacetate hydrolysis, respiration and microbial biomass in ferhly amended soils. Biol. Fertil. Soils 44: 885-890. [ Links ]
30. SAS Institute, 1990. SAS/STAT user’s guide, Version 6. SAS Institute, Inc., Cary, NC, 1848 pp.
31. Stark, C; LM Condron; A Stewart; HJ Di & M O’Callaghan. 2007. Influence of organic and mineral amendments on microbial soil properties and processes. Appl. Soil Ecol. 35: 79-93.
32. Stewart, JWB & H Tiessen. 1987. Dynamics of soil organic phosphorus. Biogeochemistry 4: 41-60. [ Links ]
33. Tabatabai, MA. 1982. Soil Enzymes. In: Page, AL; RH Miller & DR Keeney (eds). Methods of analysis, Part 2: Microbiological and Biochemical Properties. American Society of Agronomy, Madison. Pp. 903-948. [ Links ]
34. Tabatabai, MA. 1994. Soil enzymes. In: Weaver RW; S Angle; P Bottomley; D Bezdicek; S Smith; ATabatabai& A Wollum (eds). Methods of soil analysis.Part 2.Microbiological and biochemical properties.Soil Science Society of America, Madison.Pp. 775-833. [ Links ]
35. Thompson, LM & FR Troeh. 2002. Los suelos y su fertilidad. 4ta edn. España. Ed. Reverté. 657 pp. [ Links ]
36. Varela, MF; CM Scianca; MA Taboada & G Rubio.2014. Cover crop effects on soybean residuedecomposition and P release in no-tillage systems of Argentina. Soil Till. Res. 143(0): 59-66. [ Links ]
37. Wanner, BL. 1996.Phosphorus assimilation and control of the phosphate regulon. In: Neidhardt, FC. (ed). Escherichia coli and Salmonella: Cellular and Molecular Biology, ASM Press, Washington, DC, USA. Pp. 1357-1381. [ Links ]
38. Wick, B; RF Kühne& PLG Vlek. 1998. Soil microbiological parameters as indicators of soil quality under improved fallow management systems in south-western Nigeria. Plant Soil 202(1): 97-107. [ Links ]

