










Servicios Personalizados
Revista

Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkCiencia del suelo
versión On-line ISSN 1850-2067
Cienc. suelo vol.36 no.2 Ciudad Autónoma de Buenos Aires ago. 2018
BIOLOGÍA DEL SUELO
Caracterización fenotípica y genotípica de rizobios simbiontes de macroptilium erythroloma recuperados de suelos de las provincias de corrientes y santa fe
Phenotypic and genotypic characterization of the rhizobial symbionts of macroptilium erythroloma recovered from soils in the provinces of corrientes and santa fe
Laura Viviana Fornasero1*, María Antonieta Toniutti1, Nicolás Emilio Zuber1
1. Facultad de Ciencias Agrarias. Universidad Nacional del Litoral. R.P. Kreder 2805. (3080) Esperanza, Santa Fe.
*Autor de contacto: lfornase@fca.unl.edu.ar
Recibido: 26/01/2018
Recibido con revisiones: 10/05/2018
Aceptado: 11/06/2018
RESUMEN
Macroptilium erythroloma es una leguminosa perenne de amplia distribución en el centro y norte de Argentina, que se destaca entre las forrajeras nativas por su potencial productivo. Hacia el uso sustentable de esta leguminosa, se presenta el aislamiento y evaluación de la diversidad fenotípica y genotípica de los rizobios simbiontes de M. erythroloma recuperados de suelos de las provincias de Corrientes y Santa Fe. La caracterización fenotípica permitió reconocer rizobios de crecimiento lento que mostraron un desarrollo óptimo en un amplio rango de pH, temperaturas entre 28 y 35°C, y en condiciones de 0,5% (p/v) de NaCl. Se han hallado rizobios con capacidad de crecimiento a pH 9 y a 40°C. Los perfiles de amplificación de ADN genómico (fingerprints) presentaron una marcada diversidad genética entre los aislamientos. La biodiversidad de rizobios noduladores de M. erythroloma y la tolerancia a factores abióticos constituyen una valiosa herramienta hacia la selección de potenciales cepas inoculantes.
Palabras clave. leguminosas nativas, rizobios noduladores, biodiversidad.
ABSTRACT
Macroptilium erythroloma is a perennial legume of wide distribution in the north and centre of Argentina, which high lights among native forage for its productive potential. Towards the sustainable use of this legume, the isolation and evaluation of the phenotypic and genotypic of symbionts rhizobia of M. erythroloma recovered from soils in the provinces of Corrientes and Santa Fe are presented. The phenotypical characterization allows to recognize rhizobia of slow growing up which show an optimus development among a widerange of pH, temperaturas among 28 and 35°C, and with conditions of NaCl 0.5% (p/v). Rhizobia with growing conditions at pH 9 and 40°C were found. The profiles of amplification of genomic DNA (fingerprints) showed a strong genetic diversity among the isolations. Nodulating rhizobia biodiversity of M. erythroloma and the tolerance to abiotic factors establish a valuable tool towards the choosing of potential inoculant strains.
Key words. native legumes, rhizobia nodulators, biodiversity.
INTRODUCCIÓN
En Argentina, el incremento de las zonas agrícolas cultivables ha impulsado a los sistemas ganaderos a desplazarse hacia áreas menos productivas. La implantación de pasturas forrajeras en zonas cada vez más marginales constituye uno de los principales desafíos que debe afrontar el sector ganadero (Fernández Cirelli & Volpedo, 2002). Una alternativa para mejorar la productividad de los sistemas agropecuarios podría ser la introducción a cultivo de especies leguminosas nativas con reconocido potencial forrajero y adaptadas a las diferentes condiciones edafoclimáticas de nuestro país.
Las leguminosas nativas herbáceas se caracterizan por su adaptación, calidad forrajera y capacidad de fijar nitrógeno en simbiosis con bacterias de distintos géneros denominadas colectivamente rizobios. Estas especies cumplirían una función esencial por su aporte de nitrógeno al sistema y contribución a la dieta de los rumiantes. En este sentido, en ambientes tropicales y subtropicales del mundo la adopción de especies leguminosas nativas o introducidas aumentó la disponibilidad de nitrógeno edáfico y la calidad de la dieta animal (Shelton et al., 2005; Nichols et al., 2007).
Entre las leguminosas forrajeras nativas con reconocido potencial productivo se encuentra el género Macroptilium (Benth.) Urban. Plantas herbáceas, anuales o perennes, de ciclo primavero- estivo-otoñal (Pensiero & Zabala, 2017). En particular, se destaca Macroptilium erythroloma (Mart. ex Benth.) Urb., "porotillo", como una planta voluble con raíz axonomorfa leñosa, tallo y hojas pubescentes, folíolos ovalados a romboidales, flores en racimos de color rosado y fruto en vaina levemente arqueado. Es una especie perenne de crecimiento estival muy susceptible a las heladas y moderadamente resistente a las sequías. En nuestro país crece en las Provincias Fitogeográficas Paranaense, Yungas y Chaqueña (Perez et al., 1999). Evaluaciones preliminares han destacado su plasticidad en requerimientos de suelos, alta producción de forraje y semillas, y buena competencia con las malezas (Fernandez et al., 1988). Investigaciones realizadas por Martin Montiel et al., (2012) revelaron que plantas de M. erythroloma presentaron raíces suculentas reservantes que permitieron un rápido rebrote luego de sobrevivir un estrés ambiental, indicando su potencialidad para ser cultivadas como forrajeras.
M. erythroloma, al igual que la mayoría de las leguminosas forrajeras perennes presentan un crecimiento inicial lento como limitante para su implantación (Cooper, 1977). Otro de los factores determinantes de un establecimiento exitoso es la ocurrencia de una simbiosis eficiente con rizobios noduladores (Armstrong et al., 1999; Thrall et al., 2005). El conocimiento y manejo agronómico de las interacciones simbióticas de poblaciones nativas y naturalizadas de rizobios es un factor clave en la dinámica de las leguminosas nativas en sistemas ganaderos, así como en el éxito de su introducción a cultivo y persistencia.
A pesar de los antecedentes mencionados, son escasos los estudios sobre el potencial forrajero de M. erythroloma en nuestro país. Pensiero & Zabala (2017) consideraron a M. erythroloma una especie nativa promisoria, que debería ser conservada, estudiada e introducida a cultivo como recurso forrajero para la región de los Bajos Submeridionales de la provincia de Santa Fe. En virtud de ello, en la Universidad Nacional del Litoral se ha desarrollado el cultivar "Don Augusto", de M. erythroloma (INASE, 2014), que en la actualidad se encuentra en proceso de producción de semillas en forma extensiva (Zabala et al., 2015).
La simbiosis rizobio-leguminosa presenta un papel muy importante en el crecimiento de las plantas y la adaptación a nuevos ambientes, particularmente en condiciones edáficas estresantes (Howieson & Ballard, 2004; Gerding et al., 2014). En los últimos años, el estudio de los simbiontes asociados a leguminosas nativas y naturalizadas como Astragalus spp, Calliandra grandiflora, Galega offcinalis, Indigofera spp., Sesbania sp., Vigna unguiculata, entre otras, ha incrementado notablemente el conocimiento sobre la taxonomía rizobial (Lemaire et al., 2015; Peix et al., 2015; Andrews & Andrews, 2017).
Desde el punto de vista práctico, la selección de rizobios nativos adaptados a las condiciones edafoclimáticas de nuestro país favorecería el establecimiento exitoso de la simbiosis. En este contexto, la caracterización funcional y molecular de las poblaciones de rizobios nativos constituirá una valiosa herramienta hacia la selección de potenciales cepas inoculantes que permitan un mejor establecimiento de las leguminosas y con ello un incremento en la producción agropecuaria y una reducción del empleo de fertilizantes nitrogenados. Hacia el uso sustentable de esta leguminosa, el objetivo de la presente investigación fue la caracterización fenotípica y el análisis de la diversidad genotípica de los rizobios simbiontes de M. erythroloma.
MATERIALES Y MÉTODOS
Se tomaron muestras de suelos de las localidades de Santo Tomé, Corrientes (28°07'S, 56°03'W) y General Obligado, provincia de Santa Fe (28°18'S, 59°16'W), con buena aptitud potencial para el cultivo de M. erythroloma.
Los sitios de muestreo correspondieron a Santo Tomé, Provincia Fitogeográfica Paranaense, Distrito de los Campos, con suelos ultisoles que presentaron un horizonte argílico de poco espesor, de coloración pardo rojizo oscuro y textura arcillosa, y a General Obligado, Provincia Fitogeográfica Chaqueña, Distrito Oriental, con suelos alfisoles que mostraron horizontes superficiales de colores claros y estructura masiva, seguido de horizontes muy arcillosos. Las Provincias Fitogeográficas presentan un clima cálido subtropical sin estación seca, con una temperatura media anual superior a 20ºC y abundantes precipitaciones distribuidas durante todas las estaciones del año. En la Tabla 1 se detallan las características químicas de los suelos en los sitios de origen.
Tabla 1. Propiedades químicas de los suelos en los sitios de muestreo.
Table 1. Chemical properties of soils in the sampling sites.
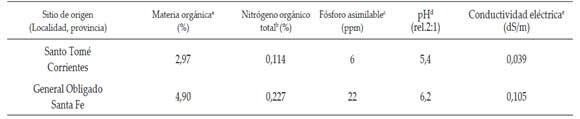
a: Walkey y Black, (1934). b: Kjeldhal, AOAC, (1990). C: Bray y Kurtz, (1945). d: potenciometría MAG, (1982) e: Conductimetría, MAG, (1982).
La colección de microsimbiontes se estableció mediante aislamientos de rizobios de nódulos radiculares de plantas trampas (plantas cultivadas en ensayos de laboratorio e inoculadas con muestras de suelo). A partir de los ensayos en cámara de crecimiento en condiciones controladas (26°C de temperatura, fotoperíodo de 16/8 horas luz/oscuridad), se colectaron nódulos del cv. Don Augusto de M. erythroloma empleando suelos provenientes de las localidades de Santo Tomé, Corrientes y General Obligado, provincia de Santa Fe.
Las semillas de M. erythroloma fueron escarificadas manualmente con lija, desinfectadas y germinadas en placas de Petri con agar-agua 1% (p/v) a 28°C. Las semillas que presentaban una radícula de 10 a 15 mm de longitud se colocaron en macetas de 500 cm3 de capacidad con drenaje conteniendo vermiculita estéril y suelo en relación 5:1. Las plántulas permanecieron a un nivel hídrico de capacidad de campo en cámara de cultivo durante 60 días y periódicamente se regaron con agua estéril.
Aislamiento y purificación de los rizobios
El procesamiento de los nódulos y aislamiento de rizobios se realizó según Vincent (1970). Los nódulos se desinfectaron con agua oxigenada 30 volúmenes durante 3 - 4 minutos y luego se realizaron cinco lavados con agua destilada estéril. Los nódulos se maceraron y el producto obtenido se sembró en cajas de Petri con Agar Extracto de Levadura Manitol (LMA) e indicador rojo Congo (Vincent, 1970). Posteriormente, fueron incubados a 28°C durante 10 días y repicados hasta la obtención de colonia pura. Los aislamientos fueron autenticados por su habilidad de formar nódulos en la planta huésped (Vincent, 1970) y conservados en caldo LMA + glicerol a -20°C.
Características fenotípicas de los aislamientos
Las cepas bacterianas se cultivaron en placas de LMA con agregado de rojo congo a 28°C y se observó la morfología macroscópica de las colonias: color, elevación, consistencia y bordes (CIAT, 1988). Las características morfológicas celulares y tinción diferencial de Gram se observaron a través de un microscopio óptico (Vincent, 1970). La velocidad de crecimiento se evaluó visualmente determinando el tiempo de aparición de las colonias según Jordan (1984). Para analizar la respuesta al cambio de coloración del medio, cada aislamiento se cultivó en caldo LMA (pH 6,8) e indicador azul de bromotimol (0,5% en NaOH 0,016N) a 28°C durante diez días y se observó el cambio de coloración a azul (alcalinidad) ó amarillo (acidez).
Posteriormente, los aislamientos se cultivaron en medio LMA a 28°C, 32°C, 35°C, 37°C, 40°C y 45°C. El crecimiento a diferentes niveles de pH y concentraciones de NaCl se determinó en placas con medio LMA con el pH ajustado a 4, 5, 6, 7, 8, 9 y 10; y conteniendo 0,01%; 0,5%; 1%; 2% y 3% (p/v) de NaCl. Para evaluar la resistencia intrínseca a antibióticos, se sembraron los aislamientos en medio LMA con adición de los siguientes antibióticos: ampicilina (20 y 40 μg/mL), kanamicina (10 y 40 μg/mL), gentamicina (5 y 20 μg/mL) y estreptomicina (20 y 100 μg/mL). El crecimiento de las cepas en diferentes condiciones de temperatura, pH, concentraciones de NaCl y con la adición de antibióticos fue evaluado por duplicado y se calificó según una escala numérica gradual desde 0 (ausencia de crecimiento) hasta 3 (desarrollo normal), comparándolo con el crecimiento obtenido en medio LMA en condiciones óptimas (pH 7, 28ºC y 0,01% p/v NaCl).
Se realizó un Análisis de Componentes Principales (ACP) de todas las variables analizadas para cada uno de rizobios de la colección. El análisis estadístico se efectuó utilizando el software InfoStat (Di Rienzo et al., 2016).
Características genotípicas de los rizobios
Las preparaciones y manipulaciones de las muestras de ADN se realizaron según las técnicas descriptas en Sambrook et al., (1989).
Las huellas digitales de amplificación de ADN total se realizaron con el cebador BOXA1R (Versalovic et al., 1994). La composición de las mezclas de reacción utilizadas fue: 50 mM Tris-HCl (pH 8,3); 3 mM MgCl2, 200 mM dNTPS; 1 U Taq DNA polymerasa (Promega Corp.) y 10 mM del cebador BOXA1R completando un volumen final de mezcla de reacción de 25 μl. Las condiciones de ciclado empleadas fueron: un ciclo de desnaturalización inicial a 94ºC 7 minutos y 30 ciclos de desnaturalización a 94ºC 10 segundos, annealing a 52ºC 60 segundos, y elongación a 72ºC 2 minutos. Al finalizar las reacciones de PCR, 10 μl de cada uno de los productos fueron separados por electroforesis en gel de agarosa.
Para el análisis de los productos de PCR se establecieron como presencia (1) o ausencia (0) de bandas para cada uno de los individuos analizados. Los datos fueron utilizados para generar un dendograma UPGMA (coeficiente de similitud Jaccard) el cual fue obtenido del análisis de los fingerprints derivados de BOXA1- PCR. Se utilizaron el software Cross Checker (para la construcción de la matriz binaria) y los programas FAMD (Fingerprint Analysis with Missing Data) versión 1.23 beta (Schluter & Harris, 2006) y MEGA 5 (Molecular Evolutionary Genetics Analysis) version 5 (Tamura et al., 2011). Para comparar la diversidad en la población de rizobios según el suelo de procedencia se empleó el índice de Shannon-Weaver (Shannon & Weaver, 1949) en función del número de aislamientos pertenecientes a cada grupo de perfiles en BOXA1R. Se consideran como individuos diferentes aquellos que presentan nivel de similitud menor al 70% (Alberton et al., 2006; Loureiro et al., 2007).
Para todos los estudios de autenticación y caracterización morfológica, fisiológica y genética se emplearon las cepas de referencia: Bradyrhizobium elkani USDA 76T, Bradyrhizobium yuanmingense CCBAU 10071T y Bradyrhizobium japonicum E 109.
RESULTADOS Y DISCUSIÓN
En el presente trabajo, los nódulos radiculares obtenidos en cámara de crecimiento evidenciaron que, en suelos de las Provincias Fitogeográficas Paranaense y Chaqueña, con características edáficas distintas y naturaleza química diferente, se encuentran poblaciones de rizobios simbiontes de M. erythroloma. A partir de los aislamientos realizados, se conformó una colección de 45 aislamientos: 27 recuperados de la provincia de Corrientes (MeC) y 18 de Santa Fe (MeS).
Las cepas bacterianas mostraron colonias circulares de bordes enteros y lisos, colores rosados o blanquecinos y translúcidas u opacas. La consistencia predominante fue mucosa, algunas presentaron un aspecto acuoso y muy pocas una apariencia seca. El aspecto de las colonias se ajustó a los fenotipos descriptos por Fuhrmann (1990) para bradyrizobios simbiontes de soja. Las células fueron bacilos pequeños, Gram negativos y no esporulados, correspondiendo a la descripción de bacterias que forman nódulos con leguminosas en el Phylum B12 Proteobacteria (Garrity et al., 2005). Los aislamientos mostraron crecimiento lento y extra-lento, siendo el tiempo de aparición de las colonias 5-6 días y 7-9 días respectivamente. Toniutti et al. (2015) obtuvieron resultados similares en la velocidad de crecimiento de rizobios simbiontes de M. atropurpureum y M. bracteatum. En este trabajo, el 67% de los rizobios alcalinizaron el medio de cultivo con el indicador azul de bromotimol, mientras que el 33% restante generó una acidificación leve ocasionando el viraje del color verde del medio a verde-amarillento.
Los aislamientos de la colección presentaron una morfología y tasa de crecimiento características del género Bradyrhizobium (Jordan, 1984). Se destaca que, algunos aislados produjeron una acidez ligera en el medio de cultivo, lo cual difiere con las características típicas de los bradyrizobios, que generalmente presentan reacción alcalina. Según Odee et al. (1997), las clasificaciones basadas en características fenotípicas estándar de morfología, velocidad de crecimiento y producción ácido-álcali no permitirían calificar a todos los rizobios nativos. Bécquer et al., (2000) informaron bradyrizobios aislados de leguminosas forrajeras de la provincia de Sancti Spíritus, Cuba, con reacción ácida. Asimismo, diversos autores han reportado rizobios de crecimiento lento que acidificaron el medio (Padmanabhan et al., 1990; Moreira et al., 1993; Bécquer et al., 2016). Esta característica que presentan algunos rizobios podría ser una ventaja competitiva, que les permitiría sobrevivir a condiciones adversas, y no se podría considerar como carácter taxonómico exclusivo (Moreno Conn, 2010).
El crecimiento de los rizobios noduladores de M. erythroloma evaluados bajo diferentes condiciones abióticas en medio LMA se presenta en la Figura 1. En relación a la temperatura, el 65% de los simbiontes recuperados de Corrientes presentó crecimiento a 35°C y solo dos aislamientos pudieron desarrollar a 37°C. Mientras que, aquellos provenientes de Santa Fe, mostraron una marcada aptitud a altas temperaturas, ya que el 67% de los aislados presentó crecimiento a 37°C y el 17% a 40°C. La variabilidad en el comportamiento de los rizobios podría responder a un proceso de adaptación natural debido a factores estresantes externos, que como la temperatura en los suelos, actúan de forma continua en los rizobios (Becquer, 2002). Según Giongo et al., (2010), la tolerancia a altas temperaturas podría ser útil en la selección de rizobacterias tolerantes a temperaturas oscilantes en condiciones de campo.
Figura 1. Crecimiento de rizobios simbiontes de M. erythroloma evaluados bajo diferentes condiciones de estreses abióticos en medio LMA. Escala de graduación: desde 3 hasta 0 indican la capacidad de los rizobios para crecer bajo la condición investigada (3=desarrollo óptimo, 0=sin crecimiento) .a: MeC: rizobios aislados de suelos de Corrientes, MeS: rizobios aislados de suelos de Santa Fe, B. japon: B. japonicum, B. yuan: yuanmingense.
Figure 1. Growth of rhizobial symbionts of M. erythroloma evaluated under different abiotic stressing conditions in LMA medium. Graduation scale: from 3 to 0 indicate the ability of rhizobia to grow under the investigated condition (3=full optimal development, 0=absence of growth). a: MeC: rhizobia isolated from soils of Corrientes, MeS: rhizobia isolated from soils of Santa Fe, B. japon: B. japonicum, B. yuan: yuanmingense.
El crecimiento óptimo para Bradyrhizobium sp. fue reportado por Jordan (1984) en un intervalo de pH 6 a 7, con cepas que manifiestan tolerancia a la acidez (De Souza et al., 2014). En relación a los microsimbiontes de M. erythroloma, el 89% de los aislamientos recuperados de Santo Tomé y el 94% de General Obligado presentaron crecimiento en un rango de pH entre 5 y 8. Sólo el 17% de los simbiontes de Santa Fe y el 26% de Corrientes desarrollaron a pH 9, mientras que ninguno pudo crecer en niveles de pH extremos. La tolerancia a diferentes niveles de pH es una característica deseable para las cepas de inoculantes potencialmente comerciales.
La presencia de sal puede afectar el crecimiento y la supervivencia de los rizobios en el suelo, disminuyendo la colonización de la raíz, el proceso de infección y con ello el desarrollo del nódulo y el funcionamiento activo en la fijación de nitrógeno (Zahran, 1991). En los rizobios, la sal provoca fundamentalmente un estrés por la concentración del ión cloruro más que por el efecto osmótico, ya que inhibe enzimas específicas (Elsheikh, 1998). Por lo tanto, el NaCl es considerado un buen indicador de la tolerancia de las bacterias al estrés salino (Abdelmoumen et al., 1999). En este trabajo, el 27% de los aislamientos presentaron desarrollo adecuado a 0,5% p/v de NaCl y sólo16% toleraron 1% p/v de NaCl, por el contrario ningún aislamiento fue capaz de crecer en niveles de salinidad superiores. Toniutti et al., (2015) obtuvieron resultados similares en la tolerancia a condiciones de salinidad de rizobios noduladores de M. atropurpureum y M. bracteatum.
En relación a la resistencia intrínseca a antibióticos, las bacterias simbiontes manifestaron resistencia a diferentes concentraciones de ampicilina y gentamicina. El 72% de los aislados de Santa Fe y el 18% de Corrientes tuvieron la capacidad de crecer en medios suplementados con 10 ug/ ml de kanamicina. Cabe destacar que, solo dos aislamientos mostraron resistencia a estreptomicina (20 μg/mL). La capacidad de los rizobios de crecer en presencia de diferentes antibióticos, le brindaría ciertas ventajas adaptativas que favorecerían la competencia por un sitio en la rizósfera y por tanto la colonización de las raíces de la leguminosa, impidiendo a su vez el establecimiento de microorganismos fitopatógenos.
Con el fin de caracterizar en más detalle la diversidad fenotípica de los rizobios noduladores de M. erythroloma y determinar los factores asociados a esa diversidad, se realizó un análisis de componentes principales (ACP) incluyendo el total de aislamientos de la colección y todas las variables analizadas para cada uno de ellos. Los resultados mostraron que los tres primeros componentes del ACP explicaron el 51% de la variabilidad total observada. En la Figura 2 se muestra la distribución de los rizobios simbiontes de M. erythroloma en el espacio de los componentes 1 y 3, que representan más de 36% de la variación total existente y el gráfico vectorial de la contribución de las variables originales a cada una de los componentes. La dispersión observada de los datos evidenció la diversidad fenotípica en la colección de germoplasma colectado. De acuerdo con la posición de los aislamientos en el plano comprendido por ambos ejes, hacia la derecha en el eje horizontal (CP1) se hallan los rizobios con mejor desempeño a diferentes temperaturas, ya sea los que desarrollaron a temperaturas de 32-35°C y generaron acidez localizados en la región positiva del CP3, o los aislamientos con mayor tolerancia a altas temperaturas y resistentes a gentamicina ubicados en la región negativa de este mismo componente.
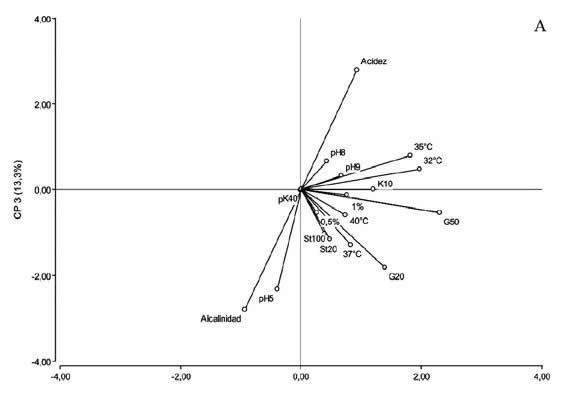
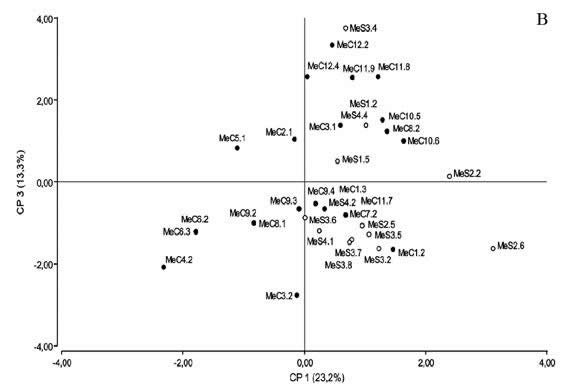
Figura 2. Análisis de Componentes Principales (ACP) basado en la separación de los aislamientos noduladores de M. erythroloma mediante sus diferentes tolerancias a estreses abióticos. A. Gráfico vectorial de correlación entre las variables de tolerancia a estreses y los componentes de variación 1 y 3. B. Diagrama de dispersión de los aislamientos en el espacio de las CP1 y CP3. Los aislamientos marcados en negro corresponden a rizobios recuperados de suelos de Corrientes y en blanco a rizobios de suelos de Santa Fe.
Figure 2. Principal-components-analysis (PCA) based separation of isolates nodulators of M. erythroloma according to their differences in tolerance to abiotic stresses. A: Vector-correlation plot between the variables examined of stress tolerances and the components of variation PC1 and PC3. B: Isolate distribution of isolates in the PC1 and PC3 spaces of variation. The isolates in black correspond to rhizobia retrieved from soils of Corrientes and in white to rhizobia from soils of Santa Fe.
El análisis de la diversidad genotípica del germoplasma asociado a M. erythroloma se realizó a través de la generación de huellas digitales de ADN, técnica que ha sido utilizada exitosamente por diversos autores para analizar la diversidad de poblaciones (Bizarro et al., 2011; Granada et al., 2014). Los resultados obtenidos del análisis revelaron once tipos de perfiles de amplificación BOXA1R diferentes evidenciando una marcada diversidad genética entre los aislamientos simbiontes. En la Figura 3 se presentan los resultados de un análisis UPGMA realizado en base a presencia/ausencia de bandas observadas en el gel para los productos de amplificación de cada uno de los aislamientos. El perfil tipo "D" está representado por 29% de las cepas simbiontes, mientras que siete perfiles BOXA1R se hallan representados solo por uno a tres aislamientos. La cepa de referencia B. yuanmingense mostró un perfil BOXA1R distinto a los observados en los aislamientos de la colección. La marcada diversidad genética de los rizobios simbiontes de M. erythroloma presentes en los suelos de Corrientes y Santa Fe, también ha sido reportada en el germoplasma asociado a las leguminosas nativas Desmanthus paspalaceus (Fornasero et al., 2014) y Desmodium incanun (Toniutti et al., 2017) procedentes del centro y norte de nuestro país. Asimismo, diversos estudios han revelado una amplia diversidad genotípica en las poblaciones de rizobios simbiontes de Dalea purpurea en pastizales de Minnesota (Martir et al., 2007), de Caragana spp. provenientes de China (Lu et al., 2009) y de Lens culinaris recuperados en diferentes localidades de Bangladesh (Rashid et al., 2012). De acuerdo a datos de la literatura, el tipo de suelo constituirá un factor determinante sobre la diversidad genética de las poblaciones de rizobios (Strain et al., 1994; Zribi et al., 2005).
Figura 3. Análisis UPGMA obtenido en base a la presencia-ausencia de bandas en los productos de amplificación BOXA1R de diferentes aislamientos noduladores de M. erythroloma recuperados de suelos de las provincias de Corrientes y Santa Fe. La figura muestra el análisis UPGMA (coeficiente de similitud de Jaccard) de los productos BOXA1R-PCR obtenidos con el ADN total de los aislamientos indicados. MeC: rizobios aislados de suelos de Corrientes, MeS: rizobios aislados de suelos de Santa Fe, B. yuan: yuanmingense.
Figure 3. UPGMA analysis obtained from the presence-absence of bands in the BOXA1R amplification products of different isolates nodulators of M. erythroloma retrieved from soils in the provinces of Corrientes and Santa Fe. The figure shows the UPGMA analysis (Jaccard similarity coefficient) derived from BOXA1R-PCR products obtained with total DNA from the indicated isolates. MeC: rhizobia isolated from from soils of Corrientes, MeS: rhizobia isolated from soils of Santa Fe, B. yuan: yuanmingense
Los valores del índice de Shannon estimados según el suelo de procedencia y un nivel de 70% de similitud en los perfiles BOXA1R fueron 3,99 para Santa Fe y 4,16 para Corrientes, mostrando que en ambos suelos existe una alta diversidad en la población de rizobios. Estos resultados coinciden con lo reportado en suelos del cono sur, en donde los valores variaron según el manejo del suelo y la región, entre 1,8 y 6, mientras que valores superiores a 3 fueron calculados para rizobios nativos (Alberton et al., 2006; Loureiro et al., 2007; Giongo et al., 2008). En este contexto, se reafirma la importancia del estudio de la biodiversidad de los rizobios noduladores de M. erythroloma, no sólo por su rol como microsimbiontes de leguminosas con potencial forrajero, sino también como un factor biótico potencialmente importante para el establecimiento de especies de Macroptilium con fines productivos.
Los criterios básicos a considerar en el proceso de selección de cepas son la capacidad de sobrevivencia en el suelo, la adaptación o tolerancia a situaciones de estrés, la infectividad y efectividad. Uno de los puntos más importantes para la selección de cepas de rizobios con potencial uso como inoculante es su capacidad de tolerancia a condiciones ambientales adversas. Así, con el propósito de identificar aislamientos que muestren tolerancia destacada a un estrés abiótico, se realizó un relevamiento de la colección de rizobios con los resultados que se presentan en la Tabla 2.
Tabla 2. Aislamientos de rizobios noduladores de M. erythroloma con tolerancia destacada a condiciones de estrés abiótico de importancia agronómica.
Table 2. Isolates of rhizobial nodulators of M. erythroloma with outstanding tolerance to abiotic stress conditions of agronomic importance.
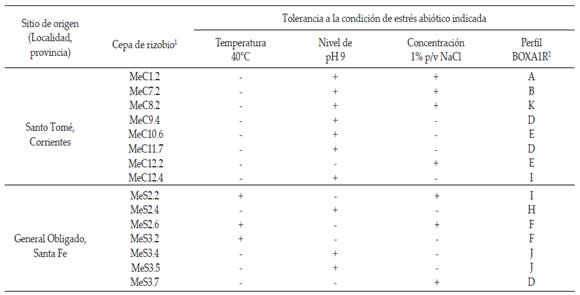
(+): Crecimiento óptimo (-): Sin crecimiento
1 seleccionadas en base a su tolerancia al menos a un factor abiótico adverso.
2 tipos BOXA1R según las huellas genómicas mostradas en la Figura 3.
Es interesante destacar que un número importante de rizobios presentaron tolerancia destacada a diferentes condiciones de estrés abiótico agrupados dentro de 9 tipos de perfiles de amplificación de ADN genómico distintos. Al mismo tiempo, cinco cepas mostraron crecimiento bajo más de un factor de estrés y los aislamientos MeC1.2 y MeS2.6 también evidenciaron resistencia a kanamicina (10 ug/ml) y estreptomicina (20 μg/mL). Los aislamientos seleccionados presentaron tolerancias marcadas a distintas condiciones de estrés frecuentemente presentes en los sitios del centro y norte de nuestro país en los que habitan las plantas de M. erythroloma y constituyen un material valioso para ser explorado de modo sistemático en programas de selección de rizobios inoculantes.
CONCLUSIONES
En suelos con características edáficas distintas y naturaleza química diferente de las provincias de Corrientes y Santa Fe se hallan bacterias nativas con capacidad de nodular plantas de M. erythroloma.
La colección de rizobios simbiontes caracterizada fenotípica y genotípicamente constituye un recurso valioso para la búsqueda y selección de cepas eficientes en fijación de nitrógeno para la elaboración de inoculantes bacterianos que estimulen la producción vegetal de M. erythroloma.
Este trabajo constituye el primer reporte de las características de diversidad feno-genotípicas de los simbiontes de M. erythroloma aislados en suelos de Argentina.
BIBLIOGRAFÍA
1. Abdelmoumen, H; A Filali-Maltouf; M Neyra; A Belabed & M Missbah El Idrissi. 1999. Effect of high salts concentrations on the growth of rhizobia and responses to added osmotica. J. Appl. Microbiol. 86: 889-898. [ Links ]
2. Alberton, O; G Kaschuk & M Hungria. 2006. Sampling effects on the assessment of genetic diversity of rhizobia associated with soybean and common bean. Soil Biol. Biochem. 38 (6): 1298-1307. [ Links ]
3. Andrews, M & ME Andrews. 2017. Specificity in legumerhizobia symbioses. Int. J. Mol. Sci. 18 (4): 705. [ Links ]
4. Armstrong, RD; K McCosker; SB Johnson; KB Walsh; G Millar; B Kuskopf; J Standley & ME Probert. 1999. Legume and opportunity cropping systems in central Queensland. 1. Legume growth, nitrogen fixation, and water use. Aust. J. Agr. Res. 50: 909-924. [ Links ]
5. Association of Official Analytical Chemists (AOAC). 1990. Official Methods of Analysis of the AOAC. Helrich, K. 15th Ed. AOAC. Arlington, USA. [ Links ]
6. Bécquer, CJ; D Prévost & A Prieto. 2000. Caracterización fisiológica-bioquímica de cepas de rizobios aislados en leguminosas forrajeras. Rev. Biol. 14: 57-65. [ Links ]
7. Bécquer, CJ. 2002. Caracterización y selección de rizobios, aislados de leguminosas nativas de Sancti Spíritus, Cuba. Ph.D. Thesis, Univ. La Habana, La Habana, Cuba, 140 pp. [ Links ]
8. Bécquer, CJ; Y Galdo; Y Ramos; MD Peña; N Almaguer; YF Peña; A Mirabal; M Quintana & A Puentes. 2016. Rhizobia isolated from forage legumes of an arid cattle rearing ecosystem in Holguín, Cuba. Morpho-cultural evaluation and nodulation (phase I). Cuban J. Agr. Sci. 50 (4): 607- 617. [ Links ]
9. Bizarro, MJ; A Giongo; LK Vargas; LFW Roesch; KA Gano; ELS De Sá; LMP Pasaglia & PA Selbach. 2011. Genetic variability of soybean bradyrhizobia populations under different soil managements. Biol. Fert. Soils. 47(3): 357-362. [ Links ]
10. Bray, RH & LT Kurtz. 1945. Determination of total, organic and available forms of phosphorus in soils. Soils Sc. 59: 39-45. [ Links ]
11. CIAT (Centro Internacional de Agricultura Tropical). 1988. Simbiosis leguminosa-rizobio. Manual de métodos de evaluación, selección y manejo agronómico. Proyecto CIAT-UNDP de evaluación, selección y manejo de la simbiosis leguminosa-rizobio para aumentar la fijación de nitrógeno (Ed.). Cali, Colombia. [ Links ]
12. Cooper, CS. 1977. Growth of the legume seedling. Adv. Agron. 29: 119-139. [ Links ]
13. De Souza, JAM; LMC Alves; A de Mello Varani & EG de Macedo Lemos. 2014. The Family Bradyrhizobiaceae. In: Rosenberg, E; EF. DeLong; S. Lory; E Stackebrandt & F Thompson (Eds.) The Prokaryotes– Alphaproteobacteria and Betaproteobacteria. Pp. 135-154. Springer-Verlag. Berlin Heidelberg.
14. Di Rienzo, JA; F Casanoves; MG Balzarini; L Gonzalez; M Tablada & CW Robledo. InfoStat versión 2016. Grupo InfoStat, FCA, Univ. Nac. Cba, Argentina. [ Links ]
15. Elsheikh, EAE. 1998. Effects of salt on rhizobia and bradyrhizobia: a review. Ann. Appl. Biol. 132:507-524. [ Links ]
16. Fernández Cirelli, A & AV Volpedo. 2002. Las Tierras secas de Iberoamérica. En: Fernández Cirelli, A & E Abraham (Eds.) El agua en Iberoamérica; De la escasez a la desertificación. Pp. 11-26. CYTED XVII, Programa Iberoamericano de Ciencia y Tecnología para el desarrollo. Buenos Aires, Argentina. [ Links ]
17. Fernandez, JG; CA Benitez; RM Pizzio & OR Pallarés.1988. Leguminosas Forrajeras Nativas del Este de la Provincia de Corrientes. Serie Téc. Nº 26, EEA Mercedes, Corrientes, 84 pp. [ Links ]
18. Fornasero, LV; MF Del Papa; JL López; FJ Albicoro; JM Zabala; MA Toniutti; JF Pensiero & A Lagares. 2014. Phenotypic, Molecular and Symbiotic Characterization of the Rhizobial Symbionts of Desmanthus paspalaceus (Lindm.) Burkart That Grow in the Province of Santa Fe, Argentina. PLoS ONE 9 (8): e104636. [ Links ]
19. Fuhrmann, J. 1990. Symbiotic effectiveness of indigenous soybean bradyrhizobia as related to serological, morphological, rhizobitoxine and hydrogenase phenotypes. Appl. Environ. Microb. 56 (1): 224-229. [ Links ]
20. Garrity, GM; JA Bell & T Lilburn. 2005. The revised road map to the manual. In: Brenner, DJ; NR Krieg; JT Staley & GM Garrity (Eds.) Bergey's Manual of Systematic Bacteriology. The Proteobacteria. Vol. 2. 2nd Ed. Pp. 159- 187. Springer, New York. [ Links ]
21. Gerding, M; GW O'Hara; JG Howieson & L Bräu. 2014. Overcoming nonselective nodulation of Lessertia by soilborne rhizobium in the presence of inoculant mesorhizobium. Plant Soil. 380: 117-132. [ Links ]
22. Giongo, A; A Ambrosini; LK Vargas; JRJ Freire; MH Bodanese- Zanettini & LMP Passaglia. 2008. Evaluation of genetic diversity of bradyrhizobia strains nodulating soybean [Glycine max (L.) Merrill] isolated from South Brazilian fields. Appl. Soil Ecol. 38(3): 261-269. [ Links ]
23. Giongo, A; A Beneduzi; A Ambrosini; L Kayser; MR Stroschein; FL Eltz; MH Bodanese-Zanettini & LM Pereira. 2010. Isolation and characterization of two plant growthpromoting bacteria from the rhizoplane of a legume (Lupinus albescens) in sandy soil. Rev. Bras. Cienc. Solo. 34 (2): 361-369. [ Links ]
24. Granada, CE; M Strochein; LK Vargas; M Bruxel; ELSD Sá & LM Passaglia. 2014. Genetic diversity and symbiotic compatibility among rhizobial strains and Desmodium incanum and Lotus spp. plants. Genet. Mol. Biol. 37(2): 396-405. [ Links ]
25. Howieson, J & R Ballard. 2004. Optimising the legume symbiosis in stressful and competitive environments within southern Australia - some contemporay thoughts. Soil Biol. Biochem. 36:1261-1273. [ Links ]
26. INASE. 2014. Cultivar de Macroptilium (Macroptilium erythroloma) Don Augusto. Ministerio de Economía y Producción, Sec. Agricultura, Ganadería, Pesca y Alimentos de la Nación, Inst. Nac. Semillas, Expte. en trámite Nro. 0069014/2014. [ Links ]
27. Jordan, DC. 1984. Bradyrhizobium, In: Kreig, NR & JG Holt (Eds.). Bergey's Manual of Systematic Bacteriology. Vol.1. Pp. 242-244. Williams & Wilkins, Baltimore, USA. [ Links ]
28. Lemaire, B; O Dlodlo; S Chimphango; C Stirton; B Schrire; J Boatwright; O Honnay; E Smets; J Sprent; E James & M Muasya. 2015. Symbiotic diversity, specificity and distribution of rhizobia in native legumes of the Core Cape Subregion (South Africa). FEMS Microbiol. Ecol. 9 (2): 1-17. [ Links ]
29. Loureiro, MDF; G Kaschuk; O Alberton & M Hungria. 2007. Soybean [Glycine max (L.) Merrill] rhizobial diversity in Brazilian oxisols under various soil, cropping, and inoculation managements. Biol. Fert. Soils. 43(6): 665-674. [ Links ]
30. Lu, YL; WF Chen; ET Wang; SH Guan; XR Yan & WX Chen. 2009. Genetic diversity and biogeography of rhizobia associated with Caragana species in three ecological regions of China. Syst. Appl. Microbiol. 32(5): 351-361. [ Links ]
31. MAG (Ministerio de Agricultura y Ganadería de la Provincia de Santa Fe). 1982. Toma de muestras y Determinaciones Analíticas en Suelos y Aguas de Santa Fe. MAG. Dirección General de Extensión e Investigaciones Agropecuarias. 152 pp. [ Links ]
32. Martín Montiel, DC; SM Pérez De Bianchi; A Etcheverry; C Camardelli; M Alemán & T Figueroa. 2012. Hábito de crecimiento y anatomía de la raíz en Macroptilium erythroloma y Macroptilium bracteatum (Fabaceae). Phyton (Buenos Aires). 81(2): 215-220. [ Links ]
33. Martir, MC; B Tlusty; P Van Berkum & PH Graham. 2007. The genetic diversity of rhizobia associated with Dalea purpurea Vent. in fragmented grasslands of west-central Minnesota. Can. J. Microbiol. 53 (3): 351-363. [ Links ]
34. Moreira, FMS; M Gillis; B Pot; K Kersters & AA Franco. 1993. Characterization of rhizobia isolated from different divergence groups of tropical Leguminosae by comparative polyacrylamide gel electrophoresis of their total proteins. Syst. Appl. Microbiol. 16: 135:146. [ Links ]
35. Moreno Conn, LM. 2010. Caracterización de las cepas ICA L9 e ICA J96, de bacterias simbióticas fijadoras de nitrógeno y pruebas de estabilidad de inoculantes elaborados para cultivos de arveja y soya. Thesis M.Sc. Pontificia Univ. Javeriana, Colombia, 126pp. [ Links ]
36. Nichols, P; A Loi; BJ Nutt; PM Evans; AD Craig; BC Pengelly; BS Dear; DL Lloyd; CK Revella; RM Nair; MA Ewinga; JG Howiesona; GA Auricht; JH Howie; GA Sandral; SJ Carra; CT de Koning; BF Hackney; GJ Crocker; R Snowballa; SJ Hughes; EJ Hall; KJ Fostera; PW Skinnera; MJ Barbetti & MP You. 2007. New annual and short-lived perennial pasture legumes for Australian agriculture-15 years of revolution. Field Crop Res. 104: 10-23. [ Links ]
37. Odee, DW; JM Sutherland; ET Makatiani; SG McInroy & JI Sprent. 1997. Phenotypic characteristic and composition of rhizobia associated with wood legumes growing in diverse Kenyan conditions. Plant Soil. 188: 65-75. [ Links ]
38. Padmanabhan, S; RD Hirtz & WJ Broughton. 1990. Rhizobia in tropical legumes: cultural characteristics of Bradyrhizobium and Rhizobium sp. Soil Biol. Biochem. 22: 23-28. [ Links ]
39. Peix, A; MH Ramírez-Bahena; E Velázquez & EJ Bedmar. 2015. Bacterial associations with legumes. Crit. Rev. Plant Sci. 34 (1-3): 17-42. [ Links ]
40. Pensiero, JF & JM Zabala. 2017. Recursos Fitogenéticos Forrajeros Nativos y Naturalizados para los Bajos Submeridionales: prospección y priorización de especies para planes de introducción a cultivo. FAVE Sección Ciencias Agrarias. 16 (1): 67-98. [ Links ]
41. Perez, SM; MC Camardelli; F Juarez; AR Bianchi & R Newman. 1999. Geographical distribution of Macroptilium species in Argentina. Trop. Grasslands. 33: 22-33. [ Links ]
42. Rashid, MHO; H Schäfer; J Gonzalez & M Wink. 2012. Genetic diversity of rhizobia nodulating lentil (Lens culinaris) in Bangladesh. Syst. Appl. Microbiol. 35(2): 98-109. [ Links ]
43. Sambrook, J; EF Fritsch & T Maniatis. 1989. Molecular Cloning: A Laboratory Manual (Second Ed.). Cold Spring Harbor, Lab. Press.New York. [ Links ]
44. Schluter, PM & SA Harris. 2006. Analysis of multilocus fingerprinting data sets containing missing data. Mol. Ecol. Notes: 6: 569-572. [ Links ]
45. Shannon, CE & W Weaver. 1949. The Mathematical Theory of Communication. University Illinois Press, Urbana, IL. [ Links ]
46. Shelton, HM; S Franzel & M Peters. 2005. Adoption of tropical legume technology around the world: analysis of success. Trop. Grasslands. 39: 198-209. [ Links ]
47. Strain, SR; K Leung; TS Whittam; FJ de Bruijn & PJ Bottomley. 1994. Genetic structure of Rhizobium leguminosarum biovar trifolii and viciae population found in two Oregon soils under different plant communities. Appl. Environ. Microbiol. 60: 2772-2778. [ Links ]
48. Tamura, K; D Peterson; N Peterson; G Stecher; M Nei & S Kumar. 2011. MEGA5: molecular evolutionary genetics analysis using maximum likelihood, evolutionary distance, and maximum parsimony methods. Mol. Biol. Evol. 28: 2731-2739. [ Links ]
49. Thrall, PH; DA Millsom; AC Jeavons; M Waayers; GR Harvey; DJ Bagnall & J Brockwell. 2005. Seed inoculation with effective root-nodule bacteria enhances revegetation success. J. Appl. Ecol. 42: 740-751. [ Links ]
50. Toniutti, MA; LV Fornasero; BS Trod; NE Zuber & MS Córdoba. 2015. Caracterización fenotípica y funcional de rizobios noduladores de dos especies del género Macroptilium. Rev. FAVE. Sección Ciencias Agrarias.14 (1): 107-119. [ Links ]
51. Toniutti, MA; LV Fornasero; FJ Albicoro; MA Martini; W Draghi; F Alvarez; A Lagares; JF Pensiero & MF Del Papa. 2017. Nitrogen-fixing rhizobial strains isolated from Desmodium incanum DC in Argentina: phylogeny, biodiversity and symbiotic ability. Syst. Appl. Microbiol. 40 (5): 297-307. [ Links ]
52. Versalovic, M; FJ Schneider; J De Bruijn & J Lupski. 1994. Genomic fingerprinting of bacteria using repetitive sequence- based polymerase chain reaction. Methods Mol. Cell Biol. 5: 25-40. [ Links ]
53. Vincent, JM. 1970. A Manual for the Practical Study of the Root-Nodule Bacteria. IBP Handbook No. 15, Blackwell Scientifc, Oxford. [ Links ]
54. Walkey, A & IA Black. 1934. An examination of the Degtjareff method for determining soil organic matter and proposed modification of the chromic acid titration method. Soil Sci. 37: 27-38. [ Links ]
55. Zabala, JM; JF Pensiero; M Forni; N Sosa; M Testa; J Giavedoni; F Aiello; A Yost & P Quarin. 2015. Valorización de los recursos fitogenéticos a través de pequeñas empresas productoras de semillas de forrajeras nativas: evaluación de algunos factores que afectan la producción de semillas en leguminosas forrajeras. En: XVII Foro de Decanos de Facultades de Agronomía del Mercosur, Bolivia y Chile. Red de Cultivos no Tradicionales de Agricultura Familiar. 1ra Ed. Pp. 65-73. Universidad Nacional de Entre Ríos. Argentina. [ Links ]
56. Zahran, HH. 1991. Conditions for successful Rhizobiumlegume symbiosis in saline environments. Biol. Fertil. Soils. 12: 73-80. [ Links ]
57. Zribi, K; R Mhamdi; T Huguet & M Aouani. 2005. Diversity of Sinorhizobium meliloti and S. medicae nodulating Medicago truncatula according to host and soil origins. World J. Microb. Biot. 21: 1009-1015. [ Links ]

