










Serviços Personalizados
Journal

Artigo
 Português (pdf)
Português (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares em
SciELO
Similares em
SciELO
Compartilhar

 Permalink
PermalinkCiencia del suelo
versão On-line ISSN 1850-2067
Cienc. suelo vol.36 no.2 Ciudad Autónoma de Buenos Aires ago. 2018
FÍSICA, QUÍMICA Y FÍSICO-QUÍMICA DE LOS SUELOS
Alterações físicas e biológicas de um eutrustox (latossolo) sob plantio direto e preparo convencional no semiárido brasileiro
Physical and biological changes of a eutrustox (latosol) under no-till and conventional tillage in the brazilian semi-arid
Cleiton Fernando Barbosa Brito1*, Varley Andrade Fonseca1, Natanael Pereira da Silva1, Stephanie Simões Braga1, Fernando Lopes Godinho1, Marcos Koiti Kondo1, Arley Figueiredo Portugal2, Michele Xavier Vieira Megda1
1. Universidade Estadual de Montes Claros- UNIMONTES, campus Janaúba-MG, Brasil
2. Empresa Brasileira de Pesquisa Agropecuária (EMBRAPA Milho e Sorgo)
*Autor de contacto: cleiton.ibce@hotmail.com
Recibido: 04/07/2018
Recibido con revisiones: 14/08/2018
Aceptado: 28/09/2018
RESUMO
A sustentabilidade dos sistemas de plantio direto e preparo convencional do solo em regiões semiáridas deve considerar as variações dos atributos do solo ao longo do tempo. Assim, objetivou-se avaliar as alterações nos atributos físicos e biológicos de um Typic Eutrustox (Latossolo) sob diferentes usos (plantio direto, preparo convencional e mata nativa) na região semiárida. O experimento foi realizado em unidades com 18 x 18 m dentro de sistema de plantio direto (SPD), preparo convencional (SPC) e mata nativa (MN). Para cada uso determinou-se a densidade de partículas, densidade do solo, porosidade, resistência à penetração, respiração microbiana e carbono orgânico nas profundidades 0,00-0,05; 0,05-0,10; 0,10-0,20 e 0,20-0,40 m. O SPD e SPC alteram os atributos físicos do solo superficial em relação à MN, com maior compactação na camada arável do SPC. O solo sob MN, além das vantagens físicas, também possui atributos biológicos superiores ao SPD e SPC em todas as profundidades avaliadas. O SPD é o uso mais próximo das condições de MN em relação à qualidade biológica, representada pela respiração microbiana. Após nove anos de implantação do SPD e SPC, destaca-se a necessidade de aporte contínuo de matéria orgânica para manutenção das vantagens do SPD em relação ao SPC nas condições do semiárido.
Palavras-Chave. Manejo do solo, plantio direto, sustentabilidade.
ABSTRACT
The sustainability of no-till and conventional tillage systems in semi-arid regions should assess soil property changes over time. In this way, the objective was to evaluate the changes in the physical and biological attributes of a Typic Eutrustox (Latosol) under different uses (no-tillage, conventional tillage and native forest) in the semi-arid region. The experiment was carried out in 18 x 18 m plots under no-till system (NT), conventional tillage (CT) and native forest (NF). Particle density, bulk density, porosity, penetration resistance, microbial respiration and soil organic carbon were determined at 0.00-0.05; 0.05-0.10; 0.10-0.20 and 0.20-0.40 m depths. The NT and CT changed soil physical properties of the surface layer, compared with NF, besides soil compaction increased in the arable layer (0.00-0.20 m) of CT. Soil under NF had superior biological properties compared to NT and CT in all evaluated depths. NT is the closest tillage compared to the NF conditions, regarding biological quality, represented by microbial soil respiration. After nine years of NT and CT, a continuous organic matter input is essential to maintain NT advantages over CT in the semi-arid conditions.
Key words: No-tillage, soil management, sustainability.
INTRODUÇÃO
O sistema de plantio direto (SPD) foi amplamente difundido nas principais regiões brasileiras produtoras de grãos (Sul, Centro-Oeste, Cerrado) como uma ferramenta importante para melhorar a qualidade do solo e a produtividade das culturas (Sá et al., 2014).
Embora consolidado nas regiões tradicionalmente produtoras de grãos, a expansão do SPD no semiárido apresenta limitações, com viabilidade associada à irrigação, devido à baixa e irregular precipitação pluviométrica. Nessas condições, o SPD apresenta inúmeros benefícios ao solo, principalmente com o aumento do carbono orgânico proveniente da decomposição de resíduos vegetais e sua mineralização (Giubergia et al., 2013) e melhora as condições físicas do solo, proporcionando um ambiente mais favorável para crescimento das raízes, quando comparado às práticas de plantio convencional (Blanco-Moure et al., 2012).
Além dos aspectos físico-químicos, os atributos biológicos do solo são importantes indicadores de alterações causadas por práticas agrícolas, úteis para o monitoramento e também orientam o planejamento e a avaliação das práticas de manejo (Ferreira et al., 2017). Notadamente, no SPD a biomassa e a atividade microbiana do solo são indicadores efetivos e consistentes de mudanças induzidas pelo cultivo e, portanto, devem ser consideradas ao avaliar o impacto do cultivo sobre a qualidade do solo (Raiesi & Kabiri, 2016).
Apesar dos benefícios descritos sobre a utilização do sistema de plantio direto em regiões semiáridas (Blanco-Moure et al., 2012, Gozubuyuk et al., 2014, Raiesi & Kabiri, 2016, Yazdanpanah et al., 2016, Kabiri et al., 2016), pesquisas no semiárido brasileiro ainda são incipientes (Sales et al., 2016 e 2017), havendo muitas lacunas no conhecimento.
Em regiões semiáridas a melhoria das propriedades físicas, químicas e biológicas do solo em SPD não ocorrem nos primeiros anos de implantação e dependem da classe de solo, clima e sucessão de culturas (Moussa-Machraoui et al., 2010). Portanto, o acompanhamento das alterações nos atributos de qualidade do solo em SPD deve ser realizado ao longo do tempo.
Além disso, é importante salientar que mudanças de uso do solo são mecanismos de ação antropogênica que afetam a biodiversidade e o funcionamento dos ecossistemas, como as que ocorrem no semiárido, com transformações de floresta nativa em áreas agrícolas, cujos efeitos são pouco conhecidos, principalmente, em relação aos atributos físicos e biológicos do solo (Ferreira et al., 2014). Neste sentido, objetivou-se avaliar as alterações nos atributos físicos e biológicos de um Typic Eutrustox (Latossolo) sob diferentes usos (plantio direto, preparo convencional e mata nativa) na região semiárida brasileira.
MATERIAL E MÉTODOS
O experimento foi realizado no Campo Experimental da Embrapa Milho e Sorgo, situado no Projeto Gorutuba, Nova Porteirinha, Minas Gerais (latitude 15°45'01" S, longitude 43°17'29" W e altitude 524 m), onde o clima, segundo Köppen, é Aw (savana com inverno seco). Os dados registrados no período de janeiro de 2009 a junho de 2017 de temperatura e precipitação média foram de 26,05 °C e 618 mm, respectivamente (Figura 1).
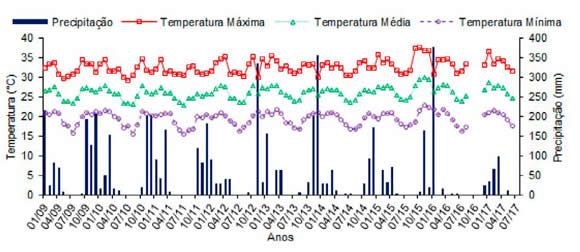
Figura 1. Temperatura (máxima, média e mínima) e precipitação durante o período de 2009 a 2017 na área experimental.
Figure 1. Temperature (Maximum, medium and minimum) and rainfall during the 2009 to 2017 period in the experimental area.
O solo foi o Typic Eutrustox (Soil Survey Staff, 1999) (Latossolo Vermelho-Amarelo Eutrófico - LVAe, segundo Santos et al., 2014), um solo oxídico, com alta saturação por bases e naturalmente fértil, embora a maior parte dos sítios de troca sejam ocupados por cátions básicos (Ca, Mg, K), tem-se nesses solos uma baixa CTC, que resulta em reserva limitada de nutrientes para as plantas. O material de origem dominante na área experimental é aluvional com coberturas detrito- -coluvionares em superfícies de aplainamento do Quaternário/Terciário, associado à metacalcilutito e metacalcarenito da Formação Lagoa do Jacaré, Grupo Bambuí (Neoproterozoico) (Mourão et al., 2012).
O solo da área experimental apresentou na camada de 0,0-0,2 m, a seguinte granulometria: 120 g kg-1 de areia grossa, 370 g kg-1 de areia fina, 210 g kg-1 de silte e 300 g kg-1 de argila; na profundidade de 0,2-0,4 m: 118 g kg-1 de areia grossa, 372 g kg-1 de areia fina, 215 g kg-1 de silte e 295 g kg-1 de argila; sendo o solo classificado como textura média, equivalente a franco-argilo- -arenosa (Sales et al., 2016).
A área foi cultivada de forma convencional alternadamente com milho e sorgo desde o ano de 1984, havendo pousio com capim colonião (Panicum maximum) entre 2004 a 2008. Entre 2009 e 2012 a área foi dividida em SPC e SPD cultivando-se nesse período três espécies (milho, sorgo e girassol) na primavera-verão que foram consorciadas - apenas no primeiro ano - com Brachiaria decumbens, para maior produção de cobertura vegetal, sendo sempre cultivado o feijão jalo rajado em sucessão no outono-inverno (Sales et al. 2016). A partir de 2013 foi conduzido com SPD e SPC os cultivos de milho/sorgo continuamente até 2015, quando se estabeleceu o pousio até a coleta das amostras, em maio de 2017.
Além disso, foi utilizado como controle a mata nativa (MN) localizada a 60 m da área experimental, na mesma classe de solo, sem qualquer ação antrópica desde a década de 1970, caracterizada como Floresta Estacional Decidual, conhecida como "mata seca". No SPC, o preparo do solo ao longo do tempo foi com arado de discos seguido por uma gradagem leve, sendo todos os restos culturais remanescentes incorporados, e a semeadura realizada com uma semeadora pneumática, a qual também foi utilizada no SPD, em cada cultivo.
Os tratamentos foram constituídos por três usos: sistema de plantio direto (SPD), sistema de preparo convencional (SPC) e mata nativa (MN), sendo cada cultivo realizado em três unidades experimentais com 18 x 18 m (324 m2), distanciadas 12 m entre si, com quatro repetições.
Em maio de 2017, após dois anos de pousio nas áreas de SPC e SPD, coletaram-se amostras de solo deformadas e indeformadas em quatro trincheiras (repetições), abertas aleatoriamente dentro de cada unidade experimental (tratamento). Para realização da coleta das amostras indeformadas foram utilizados anéis volumétricos com 0,05 m de altura e 0,054 m de diâmetro interno, nas profundidades de 0,00-0,05; 0,05- 0,10; 0,10-0,20 e 0,20-0,40 m. Desta forma, adotou-se o arranjo em esquema fatorial 3 (usos) x 4 (profundidades), disposto em delineamento experimental inteiramente casualizado, com quatro repetições, totalizando 48 amostras.
As amostras deformadas foram secas ao ar, destorroadas, passadas em peneiras de malha de 2 mm para obtenção da terra fina seca ao ar (TFSA), determinando-se a densidade de partículas (Dp) pelo método do balão volumétrico. As amostras indeformadas foram usadas para a determinação da densidade do solo (Ds) pelo método do anel volumétrico, onde também se determinou a porosidade total equivalente à umidade saturação (PT), microporosidade após extração de água em mesa de tensão, aplicando-se a sucção de 6 kPa e macroporosidade pela diferença entre a porosidade total e a microporosidade. Todas as determinações foram conduzidas segundo Donagema et al. (2011).
As amostras deformadas também foram utilizadas para a determinação do carbono orgânico total (Raij et al., 2001) e da respiração microbiana (por meio da evolução do C-CO2) conforme Curl & Rodriguez-Kabana (1972). A determinação do carbono orgânico foi realizada conforme metodologia de Walkley & Black (1934) modificado. Foram determinados os estoques de carbono das camadas para os diferentes usos do solo multiplicando-se o teor de C (dag kg-1) x densidade do solo (Mg m-3) x espessura da camada (cm).
Determinou-se também a resistência do solo à penetração, na profundidade de 0-0,4 m, com utilização do penetrômetro de impacto modelo IAA/Planalsucar/Stolf em quatro pontos representativos de cada uso do solo. Realizou- -se a equalização dos dados para as profundidades 0,00-0,05; 0,05-0,10; 0,10-0,15; 0,15-0,20; 0,20-0,25; 0,25-0,30; 0,30-0,35 e 0,35-0,40 m, conforme Stolf et al. (2014). Determinou-se a umidade do solo em cada ponto amostrado nas profundidades de 0,00- 0,10; 0,10-0,20 e 0,20-0,40 m.
Os dados foram submetidos inicialmente ao teste de normalidade e homogeneidade, em seguida procedeu-se à análise de variância e, havendo significância, comparou-se as médias pelo teste de Tukey a 5% de probabilidade. Para a análise de respirometria, onde se considerou o fator quantitativo "Dias após incubação", foram realizados ajustes das equações de regressão com base na adequação do modelo ao fenômeno estudado, significância dos parâmetros da regressão pelo teste t a 5% de probabilidade e no coeficiente de determinação ajustado (R2) e, além disso, também foi utilizado teste de Tukey para os tipos de uso dentro de cada dia da respirometria e de cada profundidade da resistência à penetração.
RESULTADOS E DISCUSSÃO
Os valores de densidade de partículas não apresentaram diferença significativa nos usos do solo para as profundidades avaliadas (Tabela 1). A densidade de partículas tem relação principalmente com a composição mineralógica nos solos não orgânicos, assim, diferenças não observadas estão relacionadas ao domínio da mesma classe de solo em todos os tratamentos.
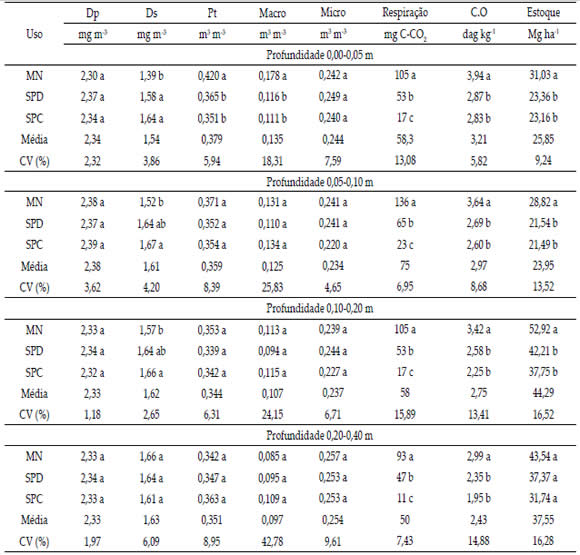
Tabela 1. Densidade de partículas (Dp), densidade do solo (Ds), porosidade total (Pt), macroporosidade (Macro), microporosidade (Micro), respiração microbiana acumulada 14 dias após a incubação, carbono orgânico (C.O) e estoque de carbono de um Eutrustox, em mata nativa (MN), sistema de plantio direto (SPD) e convencional (SPC), em diferentes profundidades.
Table 1. Particle density (Pd), bulk density (Bd), total porosity (Tp), macroporosity (Macro), microporosity (Micro), microbial respiration and soil organic carbon 14 days after incubation, organic carbon (OC) and carbon stock of a Eutrustox under native forest (NF), no-till system (NT) and conventional tillage (CT) at different depths.
Na profundidade 0,00-0,05 m os maiores valores de densidade do solo foram encontrados no SPC e SPD, superando a MN em 14 e 18%, respectivamente. Essa compactação resulta do movimento de máquinas após o preparo do solo no SPC, intensificando a aplicação de pressões acima da capacidade de suporte de carga do solo, enquanto que a o revolvimento mínimo no SPD contribui para os processos de agregação e coesão entre partículas, sendo amenizado pelo maior acúmulo de matéria orgânica.
Na profundidade 0,05-0,10 e 0,10-0,20 m a densidade do solo foi semelhante entre o SPD e SPC, sendo o SPD também semelhante a MN. O SPC superou a MN em 9,87 e 5,73% para as profundidades 0,05-0,10 e 0,10-0,20 m, respectivamente. Há uma tendência dos valores de densidade do solo no SPD se equipararem aos valores encontrados na MN nestas duas profundidades. Isto está relacionado ao não revolvimento do solo preservando sua estruturação durante nove anos (2009 a 2017) referentes ao período após a instalação dos sistemas de cultivo. A maior Ds até 0,05 m no SPD resulta do tráfego de máquinas, sendo essa compactação reduzida em profundidade, possibilitando o crescimento das raízes das plantas dentro dos bioporos e espaços entre agregados (Fernández-Ugalde et al., 2009). Para a profundidade 0,20-0,40 m os valores de densidade do solo não apresentaram diferença entre usos.
O maior valor de porosidade total na profundidade 00-0,05 m foi encontrado na MN, havendo redução desse atributo em 13,09 e 16,43% para o SPD e SPC, respectivamente. Para as demais profundidades os valores encontrados não apresentaram diferença nos diferentes usos do solo.
Os valores de macroporosidade do solo seguiram a tendência da porosidade total, em que sóhouve diferença para a profundidade de 00-0,05 m, sendo maior na MN, diferindo do SPD e SPC (Tabela 1). A redução para esse parâmetro do solo foi de 34,83 e 37,64% do valor encontrado respectivamente no SPD e SPC em relação a MN. Para as demais profundidades não foi observada diferença significativa de macroporosidade nos diferentes usos do solo.
Geralmente, os valores inferiores de macroporosidade resultam do processo de compactação superficial do solo, como observado nas maiores densidades do solo para o SPD e SPC, com 1,58 e 1,64 Mg m-3 respectivamente, na camada de 0,00-0,05 m.
A microporosidade do solo não foi influenciada pelos usos em nenhuma das profundidades estudadas, confirmando os resultados dos anos iniciais do estudo (Sales et al., 2016). Este fato ocorre em virtude da microporosidade do solo ser intensamente influenciada pela textura, teor de carbono orgânico e, muito pouco influenciada pelo aumento da densidade do solo, originada do tráfego de máquinas e implementos (Silva & Kay, 1997).
Houve efeito dos diferentes usos sobre a resistência à penetração nas profundidades 0,00- 0,05; 0,05-0,10 e 0,10-0,15 m, onde os maiores valores foram encontrados no SPD e SPC (Figura 2 A). Além disso, verificou-se diferença para as umidades apenas na camada superficial de 0,00-0,10 m, no qual a MN e o SPD apresentaram os maiores valores, diferindo do SPC. Para as profundidades de 0,10-0,20 e 0,20-0,40 m o solo apresentou uniformidade em relação à umidade (Figura 2B).
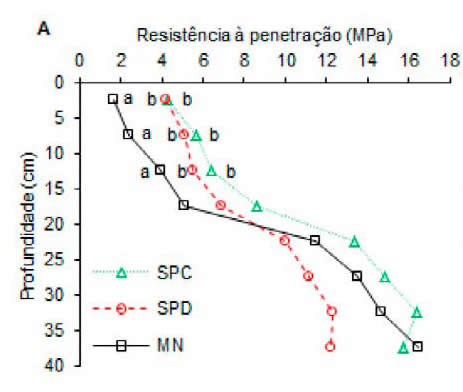

Figura 2. Resistência à penetração (A) e umidade (B) de um Eutrustox, em mata nativa (MN), sistema de plantio direto (SPD) e convencional (SPC), em diferentes profundidades. Letras iguais, na horizontal, para os usos do solo, não diferem entre si pelo teste de Tukey a 5% de probabilidade.
Figure 2. Penetration resistance (A) and humidity (B) of a Eutrustox under native forest (NF), no-till system (NT) and conventional tillage (CT) at different depths. Same letter, horizontally, for soil uses, do not differ from each other by the Tukey test at 5% probability.
Estes resultados estão coerentes, pois, na profundidade de 0,2-0,4 m não houve diferença entre os diferentes usos para a porosidade e densidade (Tabela 1) que possuem relação direta com os valores de resistência à penetração, principalmente a densidade do solo (Andrade et al., 2013).
A menor resistência à penetração verificada para a MN nas camadas superficiais do solo é resultado possivelmente da melhor qualidade física e biológica (Tabela 1) provenientes da contínua deposição e decomposição de restos vegetais (Cunha et al., 2011). Essa matéria orgânica também contribui para aumentar a retenção de água no solo e reduzir sua resistência à penetração, devido ao efeito lubrificante do filme de água formado sobre as partículas do solo, aumentando sua plasticidade, conforme observa-se na figura 2.
Verificou-se também um indicativo de uma camada compactada ou de adensamento natural, através da coesão das partículas, a partir de 0,20 m. Essas camadas compactadas são comuns em áreas com intenso uso de máquinas no preparo do solo (Teixeira et al., 2016). No presente estudo essa tendência é mais evidenciada devido à umidade do solo não diferir a partir dessa profundidade.
Os valores elevados de resistência à penetração devem-se a baixa umidade do solo nas condições de campo (Figura 2B) e corroboram com pesquisas que verificaram valores superiores a 20 Mpa, para a profundidade se 20-40 cm, em condições de umidade de 0,025 kg kg-1(Silveira et al., 2010). Esse fato fica evidenciado devido aos valores de densidade do solo não se alterarem com as diferentes formas de uso.
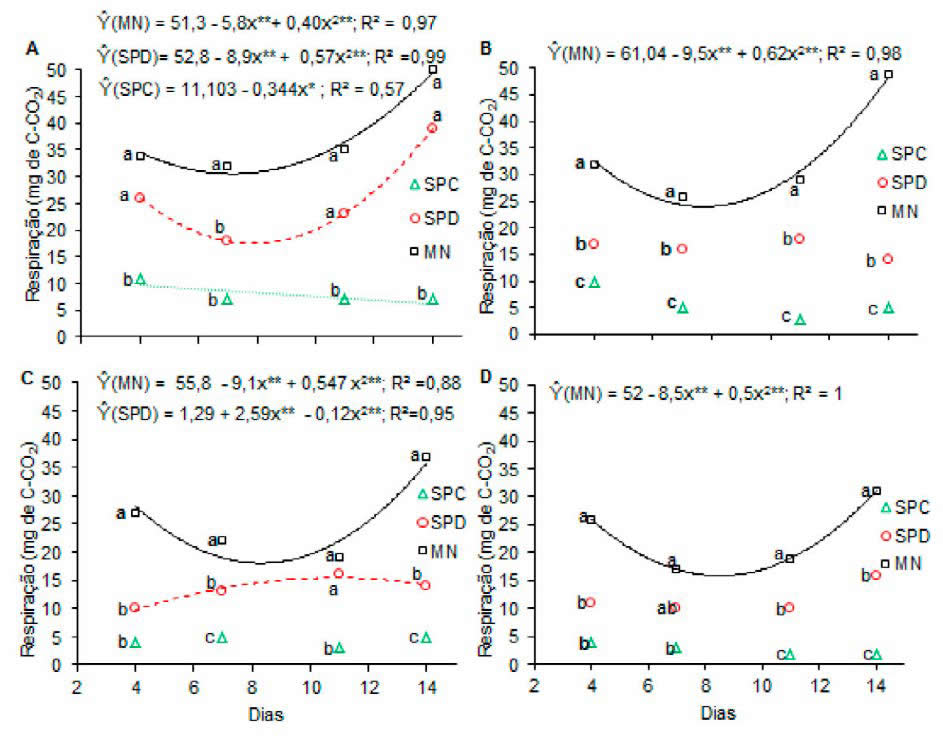
A respiração microbiana apresentou efeito significativo para a interação usos versus dias após a incubação (P < 0,05) em todas as profundidades avaliadas (Figura 3). Na profundidade 0,00- 0,05 m verificou-se que a evolução da respiração microbiana (C-CO2) ajustou-se ao modelo de regressão quadrático para o SPD e MN e linear para SPC. Já em relação aos diferentes usos verificou- se que, de forma geral, considerando os dias avaliados, não houve diferença significativa para a respiração microbiana entre a MN e SPD, onde ambas diferiram do SPC que apresentou os menores valores de (C-CO2) (Figura 3A).
Figura 3. Respiração microbiana nas profundidades 0,00-0,05 (A), 0,05-0,10 (B), 0,10-0,20 (C) e 0,20-0,40 m (D) de um Eutrustox, sob mata nativa (MN), sistema de plantio direto (SPD) e convencional (SPC), ao longo de 14 dias de incubação. Letras iguais, na vertical, para os usos do solo dentro de um mesmo tempo, não diferem entre si pelo teste de Tukey a 5% de probabilidade.
Figure 3. Microbial respiration at 0.00-0.05 (A), 0.05-0.10 (B), 0.10-0.20 (C) and 0.20-0.40 m depths (D) of a Eutrustox under native forest (NF), no-till system (NT) and conventional tillage (CT), during 14 days of incubation. Same letters, vertically, for the land uses within the same time, do not differ among themselves by the Tukey test at 5% probability.
Os valores de C-CO2 se ajustaram ao modelo de regressão quadrática para o solo da MN nas profundidades 0,05-0,10; 0,10-0,20 e 0,20- 0,40 m e SPD em 0,10-0,20 m. Para as demais profundidades em relação ao SPD e SPC não houve ajuste significativo de nenhum modelo que se adequasse ao fenômeno biológico estudado. Ao contrário do que ocorreu na profundidade de 0,00-0,05 m para os diferentes usos, nas profundidades de 0,05-0,10; 0,10-0,20 e 0,20-0,40 m a MN ocorreram maiores emissões de C-CO2, diferindo do SPD e SPC. Já entre estes, o SPD apresentou o maior valor (Figura 3 B, C e D).
A utilização de Panicum maximum, nos cinco anos anteriores à implantação, o plantio de milho, sorgo e girassol consorciados com Brachiaria decumbens no primeiro ano, a presença de plantas espontâneas durante o pousio dos tratamentos, a cobertura morta produzida pelas sucessões e o possível aumento na densidade radicular que favoreceu uma maior alocação de carbono nas camadas superficiais do solo (Sá et al. 2014), foram os fatores que contribuíram para uma evolução de C-CO2 no SPD semelhante às condições de MN nacamada de 0,00-0,05 m. Este resultado confirma a maior intensidade dos processos de transformação da matéria orgânica pelos microrganismos do solo em sistemas com aporte contínuo de carbono, como no SPD (Cunha et al., 2011).
No entanto, em sistemas convencionais de cultivo onde as práticas culturais de revolvimento desestruturam o solo e contribuem para reduzir a quantidade e, possivelmente, a diversidade de microrganismos, observam-se reflexos negativos na microbiota do solo. Além disso, presume-se que a quantidade de C nesses sistemas não seja suficiente para atender à demanda para a manutenção da biomassa existente (Rui et al., 2016) e, como o C orgânico é o nutriente que mais limita o crescimento microbiano, as taxas respirométricas tendem à estabilização após seu consumo. Estas mesma tendência está de acordo Rui et al. (2017). Assim, justificam-se os menores valores de C-CO2 em SPC e o ajuste linear na camada 0,00-0,05 m do presente estudo.
Para a MN e SPD nas profundidades avaliadas, com exceção para 0,10-0,20 m em SPD, verificou- se a mesma tendência com uma queda da evolução C-CO2 até um ponto mínimo e posterior recuperação até o final da avaliação (14 dias). No início da incubação a ação da atividade microbiana resulta numa rápida decomposição da matéria orgânica e liberação de C-CO2 (Cunha et al., 2011). Em seguida ocorre uma redução gradativa do quarto ao sétimo dia após incubação que pode ser decorrente da morte de alguns microrganismos. No entanto, estes podem servir de alimento para a microbiota remanescente e assim elevar novamente a liberação do C-CO2 oriundo da atividade microbiana (Loss et al., 2013) como foi observado do sétimo até o 14º dia de incubação.
Os valores médios de acúmulo de respiração microbiana, 14 dias após a incubação, foram em ordem decrescente: MN > SPD > SPC, com diferença significativa entre as médias para três usos do solo em todas as profundidades avaliadas. Para o carbono orgânico houve efeito semelhante, com o maior valor para a MN, porém, em nenhuma das profundidades observou-se diferença entre os valores médios referentes ao SPD e SPC. Por fim, verificou-se que a MN apresentou o maior estoque de carbono, com exceção da profundidade 0,20-0,40 m que não houve diferença entre os usos (Tabela 1).
Os resultados de respiração microbiana encontram- se coerentes com pesquisas já realizadas com diferentes usos do solo (Dadalto et al., 2015; Ferreira et al., 2017). Em condições de MN a maior liberação de CO2 é em função da constante incorporação e ciclagem de resíduos vegetais, aumentando a biomassa microbiana e a atividade biológica sobre esse material (Fang et al., 2016). Assim, a adição de fontes de carbono ao solo, como as utilizadas no presente estudo (palhada de milho, sorgo, Panicum maximum, Brachiaria decumbens) estimulam a respiração microbiana (Loss et al., 2013).
O CO2 liberado na respiração é produto do fornecimento de energia para manutenção microbiana do solo (Fang et al., 2016). Portanto, a maior liberação de CO2, geralmente, está associada à maior atividade biológica, que, por sua vez, está relacionada diretamente com a quantidade de carbono no solo (Cunha et al., 2011). Assim, como no presente estudo a MN apresentou maior carbono orgânico e maior respiração microbiana acumulada é possível inferir que a diferença em relação ao SPD e SPC, está ligada ao estado de conservação em que se encontra a mata, o que também foi verificado para as características físicas de densidade, porosidade total e macroporosidade.
O SPD e o SPC não diferiram em relação ao teor de carbono em nenhuma das profundidades. No entanto, as quantidades de carbono semelhantes entre os dois sistemas, proporcionaram diferentes valores de respiração microbiana. Portanto, o SPC contribui para reduzir a quantidade e, possivelmente, a diversidade de microrganismos, com grande impacto na microbiota do solo. Já no SPD, onde a comunidade microbiana do solo é pouco perturbada, há maior estabilidade, contribuindo para maior sustentabilidade do sistema (Ferreira et al., 2017).
Em pesquisa realizada por Raiesi & Kabiri (2016) sob condições de clima semiárido verficou- se que o preparo convencional, com aração com arado de disco, reduziu significativamente a atividade microbiana do solo. Estes resultados confirmam que a biomassa microbiana é fortemente afetada pelas atividades de manejo do solo, principalmente em condições de SPC (Pandey et al., 2014), onde o revolvimento do solo resulta em alta desordem, com efeitos negativos como a ruptura dos agregados do solo e exposição da matéria orgânica, antes protegida fisicamente no interior dos agregados (Loss et al., 2013).
Na área em estudo as sucessões de cultivo foram encerradas em 2012, sem aportes significativos de carbono nos últimos cinco anos. Soma-se a este fato, a rápida decomposição da matéria orgânica, pela ação da microbiota associada a umidade do solo no verão e às altas temperaturas do clima semiárido (Rui et al., 2017). Como o acúmulo de carbono é proporcional ao tempo de implantação do SPD (Moussa-Machraoui et al., 2010), não houve condições para a manutenção desse acúmulo, igualando o carbono orgânico do SPD ao SPC. Em consequência disso o estoque de carbono considerando a profundidade total de 0,00-0,40 m foi de 156,31; 124,48 e 114,14 Mg ha-1 para MN, SPD e SPC, respectivamente.
Verifica-se que em condições semiáridas que precipitações com menor intensidade e bem distribuídas propiciam menor perda de carbono orgânico quando comparada com chuvas concentradas e com maior intensidade, pois mudanças súbitas e intensas na umidade do solo associadas a chuvas pouco frequentes são estressantes para os microrganismos, uma vez que estes devem gastar energia para regular a pressão osmótica para o seu microambiente (Rui et al., 2017). Neste contexto, as características de irregularidade e concentração de chuvas verificadas na região semiárida do Norte de Minas Gerais (Figura 1) também favoreceram a diminuição do carbono orgânico no SPD ao longo dos cinco anos de pousio.
Outro fator relevante que pode ter contribuído para a estabilização do teor de carbono entre o SPD e SPC foi à associação do carbono com a argila e a estabilização dos materiais orgânicos dentro dos agregados, o que é comum em solos com elevados teores de argila e silte (Moussa-Machraoui et al., 2010) como o do presente estudo.
Apesar da ausência de diferenças entre os dois sistemas para os valores de carbono orgânico e características físicas do solo, os valores de respiração microbiana, nos primeiros centímetros de profundidade do solo, indicam que o SPD é o que mais se aproxima das condições da mata e, assim, infere-se que esse sistema causa menor impacto do que o SPC. Esta tendência foi verificada em pesquisas realizadas na mesma área experimental (Sales et al., 2016 e 2017) e, também em outras regiões tradicionais com o uso desses dois tipos de sistema (Ferreira et al., 2017).
CONCLUSÕES
O SPD e SPC alteram os atributos físicos do solo superficial em relação à MN, com maior compactação na camada arável do SPC. O solo sob MN, além das vantagens físicas, também possui atributos biológicos superiores ao SPD e SPC em todas as profundidades avaliadas. O SPD é o uso mais próximo das condições de mata nativa (MN) em relação à qualidade biológica, representada pela respiração microbiana.
AGRADECIMENTOS
Os autores agradecem à FAPEMIG e CAPES pela concessão de bolsas, e à EMBRAPA, pela disponibilidade da área experimental e auxílio logístico. O presente trabalho foi realizado com apoio da Coordenação de Aperfeiçoamento de Pessoal de Nível Superior - Brasil (CAPES) - Código de Financiamento 001.
BIBLIOGRAFÍA
1. Andrade, RS.; LF Stone & LG Godoy. 2013. Estimativa da resistência do solo à penetração baseada no índice S e no estresse efetivo. Rev. bras. eng. agríc. ambient. 17 (9): 932-937. [ Links ]
2. Blanco-Moure, N; LA Angurel; D Moret-Fernández & MV López. 2012. Tensile strength and organic carbon of soil aggregates under long-term no tillage in semiarid Aragon (NE Spain). Geoderma 18: 423-430. [ Links ]
3. Cunha, EQ; LF Stone; EPB Ferreira; AD Didonet; JAA Moreira & W Leandro. 2011. Sistemas de preparo do solo e culturas de cobertura na produção orgânica de feijão e milho. II - Atributos biológicos do solo. R. Bras. Ci. Solo 35: 603-611. [ Links ]
4. Curl, EA & R Rodriguez-Kabana. 1972. Microbial interactions. In: RE Wilkinson (ed.). Research methods in weed science. Pp. 162-194. Southern Weed Society. [ Links ]
5. Dadalto, JP; Fernandes, HC; Teixeira, MT; Cecon, PR; Matos, AT. 2015. Sistema de preparo do solo e sua influência na atividade microbiana. Eng. Agríc. .35 (3): 506-513. [ Links ]
6. Donagema, GK; DVB Campos; SB Calderano; WG Teixeira & JHM Viana (eds). 2011. Manual de métodos de análise do solo. Embrapa Solos. [ Links ]
7. Fang, X; D Yu; W Zhou; L Zhou & L Dail. 2016. The effects of forest type on soil microbial activity in Changbai Mountain, Northeast China. A. For. Science 73 (3): 473-482. [ Links ]
8. Fernández-Ugalde, O; I Virto; P Bescansa; MJ Imaz; A Enrique & DL Karlen. 2009. No-tillage improvement of soil physical quality in calcareous, degradation-prone, semiarid soils. Soil Til. Res. 106(1): 29-35. [ Links ]
9. Ferreira, ACC; LFC Leite; ASF Araújo & N Eisenhauer. 2014. Land-use type effects on soil organic carbon and microbial properties in a Semi-arid Region of Northeast Brazil. Land. Degrad. Dev. 27(2): 01-08. [ Links ]
10. Ferreira, EPB; LF Stone & CCG Martin-Didone. 2017. População e atividade microbiana do solo em sistema agroecológico de produção. Rev. Ciênc. Agron. 48(1): 22-31. [ Links ]
11. Giubergia, JP; E Martellotto & RS Lavado. 2013. Complementary irrigation and direct drilling have little effect on soil organic carbon content in semiarid Argentina. Soil Till. Res. 134:147-152. [ Links ]
12. Gozubuyuk, Z; U Sahin; I Ozturk; A Celik & MC Adiguzel. 2014. Tillage effects on certain physical and hydraulic properties of a loamy soil under a crop rotation in a semiarid region with a cool climate. Catena 118 (2): 195- 205. [ Links ]
13. Kabiri, V; F Raiesi & MA Ghazavi. 2016. Tillage effects on soil microbial biomass, SOM mineralization and enzyme activity in a semi-arid Calcixerepts. Agric. Ecosyst. Environ. 232 (1): 73-84. [ Links ]
14. Loss, A; MG Pereira; SJ Beutler; A Perin & LHC Anjos. 2013. Carbono mineralizável, carbono orgânico e nitrogênio em macroagregados de Latossolo sob diferentes sistemas de uso do solo no Cerrado Goiano. Semina: Ciênc. Agrár. 34 (5): 2153-2168. [ Links ]
15. Mourão, MAA; Oliveira, MJR; Noce, CM; Sad, JH. 2012. Sad. Mapa Geológico da Folha Janaúba Gerais, Brasil. Folha SD.23-Z-D-IV. Escala 1:100.000. CODEMIG. Projeto Criação de Banco de Dados Geográfico para os Mapas Geológicos do Projeto Espinhaço, Minas Gerais. [ Links ]
16. Moussa-Machraoui, SB; F Errouissi; M Ben-Hammouda & S Nouira. 2010. Comparative effects of conventional and no-tillage management on some soil properties under Mediterranean semi-arid conditions in northwestern Tunisia. Soil Till. Res. 106: 247–253.
17. Pandey, D; M Agrawal & JS Bohra. 2014. Effects of conventional tillage and no tillage permutations on extracellular soil enzyme activities and microbial biomass under rice cultivation. Soil Till. Res. 136 (4): 51-60. [ Links ]
18. Raiesi, F & V Kabiri. 2016. Identification of soil quality indicators for assessing the effect of different tillage practices through a soil quality index in a semi-arid environment. Ecol. Indic. 71(3): 198-207. [ Links ]
19. Raij, B.V; JC Andrade; H Cantarella & JA Quaggio. 2001. Determinação da Matéria Orgânica. In: BV Raij; JC Andrade; H Cantarella; JA Quaggio (eds). Análise química para avaliação da fertilidade de solos tropicais. pp.189- 199. Instituto Agronômico de Campinas. [ Links ]
20. Rui, Y; DB Gleeson; DV Murphy & FC Hoyle. 2017. Response of microbial biomass and CO2-C loss to wetting patterns are temperature dependent in a semiarid soil. Scien. Rep. 7: 01-12. [ Links ]
21. Rui, Y; DV Murphy; X Wang & FC Hoyle. 2016. Microbial respiration, but not biomass, responded linearly to increasing light fraction organic matter input: Consequences for carbon sequestration. Scien. Rep. 6: 01-09. [ Links ]
22. Sá, JCM; F Tivet; R Lal, R; C Briedis; DC Hartman; JZ Santos & JB Santos. 2014. Long-term tillage systems impacts on soil C dynamics, soil resilience and agronomic productivity of a Brazilian Oxisol. Soil Till. Res. 136: 38-50. [ Links ]
23. Sales, RP; RF Pegoraro; AF Portugal; JAA Moreira & MK Kondo. 2017. Organic matter fractions of an irrigated Oxisol under no-till and conventional tillage in the brazilian semi-arid region. Rev. Caatinga 30 (2): 303-312. [ Links ]
24. Sales, RP; AF Portugal; JAA Moreira; MK Kondo & RF Pegoraro. 2016. Qualidade física de um Latossolo sob plantio direto e preparo convencional no semiárido. Rev. Ciênc. Agron. 47(3): 429-438. [ Links ]
25. Santos, HG; Jacomine, PKT; Anjos, LHC; Oliveira, VA; Lumbreras, JF; Coelho, MR; Almeida, JA; Cunha, TJF & Oliveira, JB (eds). 2014. Sistema brasileiro de classificação de solos. Embrapa Solos. [ Links ]
26. Silva, AP & BD Kay. 1997. The sensitivity of shoot growth of corn to the least limiting water range of soils. Plant Soil 184(2): 323-329. [ Links ]
27. Silveira, DC; Melo Filho, JF; Sacramento, JAAS; Silveira, ECP. 2010. Relação umidade versus resistência à penetração para um Argissolo Amarelo distrocoeso no recôncavo da Bahia. R. Bras. Ci. Solo, 34 (20):659-667. [ Links ]
28. Soil Survey Staff. Soil taxonomy: A basic system of soil classification for making and interpreting soil surveys. 1999. 2nd edition. Natural Resources Conservation Service. U.S. Department of Agriculture Handbook 436. [ Links ]
29. Stolf, R; JH Murakami; C Brugnaro; LG Silva; LCF Silva & LAC Margarido. 2014. Penetrômetro de impacto Stolf - programa computacional de dados em Excel-VBA. R. Bras. Ci. Solo 38 (3): 774-782. [ Links ]


{kind=link}