










Services on Demand
Journal

Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO
Related links
-
 Similars in
SciELO
Similars in
SciELO
Share

 Permalink
PermalinkCiencia del suelo
On-line version ISSN 1850-2067
Cienc. suelo vol.38 no.1 Ciudad Autónoma de Buenos Aires July 2020
BIOLOGÍA DEL SUELO
Estructura de la comunidad de la mesofauna edafica en dos suelos con distinta intensidad de uso
Mesofauna community structure in two soilswith different use intensity
Salvador Nicosia1-2; Liliana Beatriz Falco2-3; Ricardo Castro Huerta4; Rosana Veronica Sandler3; Carlos Eduardo Coviella2-3*
1 Instituto de Biotecnología, INTA Castelar, Buenos Aires, Argentina
2 Departamento de Ciencias Basicas e INEDES (CONICET-UNLu). Argentina
3 Programa de Investigaciones en Ecología Terrestre. Departamento de Ciencias Básicas e INEDES (CONICET-UNLu) Universidad Nacional de Lujan. Argentina
4 Escuela de Agronomía, Facultad de Cs. Agrarias y Forestales, Universidad Católica del Maule. Chile
* Autor de contacto: carlosecoviella@yahoo.com
Recibido: 21/05/2019
Recibido con revisiones: 27/11/2019
Aceptado: 27/11/2019
RESUMEN
El aumento en la intensidad del uso de los suelos es un factor del cambio global de importancia en la degradación de los agroecosistemas que pone en riesgo su uso sustentable y afecta la pérdida de los servicios ecosistémicos provistos por los suelos. La fauna edáfica tiene un rol relevante en la provisión de estos servicios, tales como descomposición de la materia orgánica, la actividad microbiológica y el ciclado de nutrientes. Las prácticas agrícolas intensivas afectan la fauna edáfica, modificando su composición y riqueza. El estudio de la fauna edáfica resulta entonces un útil descriptor del estado de conservación del suelo. El objetivo de este trabajo, fue determinar la existencia de cambios en la estructura de la mesofauna edáfica asociados a dos intensidades de uso de los suelos. Se trabajó con muestras de mesofauna asociada a la hojarasca de dos suelos argiudoles con distinta intensidad de uso: suelos con 40 años de agricultura intensiva bajo siembra directa y suelos de pastizales naturalizados con al menos 50 años. Se tomaron muestras de mesofauna a intervalos regulares durante 8 meses. Los artrópodos presentes en las muestras se extrajeron mediante embudo de Berlesse y se identificaron al mayor nivel taxonómico posible. Los resultados muestran que la estructura de la comunidad edáfica entre los sistemas contrastados difiere en riqueza, composición y diversidad, y que ésta última varía según la fecha de muestreo. Mediante análisis de componentes principales se determinó la existencia de diferencias entre ambas comunidades y un análisis de similitud por permutaciones, ADONIS, confirma las diferencias en la estructura de las comunidades. La detección de diferencias estructurales entre ambas reafirma la sensibilidad de los artrópodos de suelo a distintas intensidades de uso. Los resultados de este trabajo también sugieren que el sistema de siembra directa, cuando es correctamente implementado, es una práctica sustentable.
Palabras clave: Impacto antrópico, pérdida de servicios ecosistémicos, uso sustentable, siembra directa.
ABSTRACT
The increase in soil use intensity is a part of global change and it is relevant for agroecosystem degradation, risking soil sustainable use and threatening the loss of ecosystem services provided by soils. The soil fauna has a relevant role in providing those ecosystem services, such as organic matter decomposition, microbiological activity, and nutrient cycling. Intensive agricultural practices affect soil fauna, modifying its composition and richness. Its study can then be a useful descriptor of soil conservation status. The objective of this work was to test the existence of structure changes in the soil mesofauna associated to two different soil use intensities. Soil mesofauna samples associated to leaflitter of two argiudoil soils with different use intensity were collected: soils with 40 years of intensive agriculture under no-tillage, and soils of naturalized pastures for at least 50 years. Decomposition bags were placed under the leaflitter, and one bag was removed at regular intervals during 8 months. Arthropods present in the samples were separated by Berlesse funnel and identified at the highest possible taxonomic milevel. The results show that the structure of the soil community between the contrasted systems differs and that this difference varies with sampling moment. By means of main component analysis, the existence of differences between both communities was determined and an analysis of similarity by permutations, ADONIS, confirms the differences in community structure. The detection of structural differences between systems confirms the sensitivity of soil arthropods to different use intensities. These results also suggest that no-tillage system, when properly implemented, is a sustainable practice.
Keywords: Anthropic impact, ecosystem services loss, sustainable use, no-tillage
INTRODUCCIÓN
En el perfil de un suelo se alojan diversas comunidades relacionadas en variadas y complejas formas (Schaefer, 1995; Altieri, 1999; Lavelle & Spain, 2003). Según el tamaño del animal adulto, la fauna del suelo se divide en: microfauna, mesofauna y macrofauna. La macrofauna está compuesta por organismos mayores a 2 mm, usualmente pertenecientes distintos estadíos de invertebrados, mientras que la microfauna está compuesta por organismos menores a 0,2 mm y se compone esencialmente de micorrizas, hongos, nematodes y bacterias. La mesofauna edáfica está compuesta por organismos cuyo tamaño oscila entre los 0,2 y 2 mm. Pertenecen a ésta los microartrópodos (ácaros, colémbolos, proturos, dipluros y sinfílos) y los enquitréidos. Estos grupos forman parte de un componente sustancial para la regulación de las condiciones edáficas ya que modulan las condiciones físicas, químicas y biológicas del perfil (Hendrix et al.,1990; Cabrera et al., 2001; Gizzi, 2009; Menta et al., 2011; Benckiser, 2018).
En particular, los artrópodos que forman parte de la mesofauna edáfica son un componente importante del ecosistema debido a su relevante rol como catalizadores de la actividad microbiológica en la descomposición de la materia orgánica y en el ciclado de nutrientes (Wallwork, 1983; Cortet et al., 1999; Dominguez et al., 2018). También influyen en el mantenimiento de factores físicos del suelo tales como la aireación, retención de agua y circulación vertical de materia orgánica. En este mismo sentido, la biota edáfica juega un papel importante en la estructura del suelo, por ejemplo, los exudados bioquímicos de los microorganismos y la actividad física de los animales edáficos son los principales factores en la formación de materia orgánica y en el mantenimiento de la estructura porosa (Benckiser, 2018). Como principales integrantes de la mesofauna edáfica, los Colémbolos (Insecta, Collembola) y los ácaros oribátidos (Acari, Cryptostigmata), constituyen entre el 72 y el 98% de los individuos de la fauna total de artrópodos del suelo (Lindo & Viser, 2003).
La dinámica de la materia orgánica en el suelo es el resultado de una compleja cadena trófica (Labrador, 1996; Barrios 2007), siendo la formación del humus consecuencia de un proceso fundamentalmente biológico en el que intervienen la mayor parte de los organismos que allí viven. Según Cabrera & Crespo (2001) la principal contribución de la mesofauna a la dinámica de la materia orgánica, es la de promover el crecimiento y la distribución de microorganismos, hongos y transporte de los productos de descomposición a los estratos inferiores y a las zonas radicales del perfil del suelo. Estos a su vez pueden favorecer la velocidad de descomposición. En este sentido, Gupta (1994) señala que la descomposición de la materia orgánica puede ser acelerada en 1,7 a 1,9 veces en presencia de ácaros.
Las prácticas agrícolas intensivas afectan fuertemente la estructura de la fauna edáfica, llevando a la extinción local a algunos grupos de organismos y a cambios en su riqueza, diversidad y abundancia (Bedano & Ruf, 2007). Dicho impacto depende del sistema de siembra, la diversidad y rotación de cultivos, los insumos utilizados y las condiciones edafoclimáticas locales (Aquino et al., 2008). Domínguez et al. (2011), señalan que la densidad de los principales grupos de la fauna edáfica disminuye según el siguiente gradiente de intensidad de manejo: sitios naturales, sitios bajo agricultura orgánica, sitios con agricultura química bajo siembra directa y sitios con agricultura química y labranza convencional.
El reciente periodo de expansión de la agricultura en la Argentina es motivo de preocupación debido a perturbaciones crecientemente asociadas a la degradación de los agroecosistemas. Así, de los cincuenta millones de hectáreas que abarca la región pampeana húmeda, se estima que más de doce millones manifiestan en mayor o menor grado efectos erosivos (hídricos y eólicos), lo cual genera degradación física, química y biológica (Lenardon, 2010).
En este contexto, la hipótesis de este trabajo fue que diferencias en la intensidad de uso de dos suelos se verán reflejados en diferencias en la estructura de la mesofauna edáfica.
El objetivo de este trabajo, fue el de evaluar la existencia de dichas diferencias en la estructura de la mesofauna edáfica asociadas a dos intensidades de uso de los suelos.
MATERIALES Y MÉTODOS
El diseño experimental fue de 5 réplicas para cada una de las dos situaciones o tratamientos: 1.- Sitios de agricultura intensiva bajo siembra directa y 2.- Sitios de pastizal naturalizado.
Las muestras sobre las que se trabajó fueron tomadas de suelos Argiudoles típicos (Carta de suelos INTA, 2013), con diferente intensidad de uso ubicados en distintos potreros contiguos y pertenecientes al mismo productor, en la localidad de Chivilcoy (35º 03' S; 59º 41' O), de la provincia de Buenos Aires, Argentina.
En la superficie del suelo de cada sitio de muestreo, se colocaron bolsas de descomposición de 20 x 20 cm y 4 mm de tamaño de malla, conteniendo 5 gramos de hojarasca de soja (Glycinemax L. Merrill). Todas las bolsas se colocaron al inicio de la experiencia, y durante un periodo de 8 meses se extrajeron 5 bolsas al azar de cada tratamiento en los intervalos de tiempo 17, 53, 94, 126,171 y 252 días.
En el laboratorio, el material de cada bolsita se colocó en recipientes para la extracción de los organismos de la mesofauna presentes en cada muestra mediante la técnica de Berlese-Tullgren (Berlese, 1905; Tullgren, 1918). Los organismos extraídos se conservaron en alcohol 70% hasta su identificación taxonómica.
Cada una de las muestras se analizó bajo microscopio binocular para la separación y cuantificación de los individuos presentes. Para la determinación de la mesofauna edáfica se utilizaron las claves de Kethley (1990), Krantz & Ainscough (1990), Norton (1990), Philips (1990), Eisenbeis (2006), Walter & Proctor (2008) & Bellinger et al. (2012).
De cada muestreo se calcularon riqueza específica (S); abundancia como número de individuos por cada grupo taxonómico, abundancia relativa, como el número de individuos por grupo taxonómico sobre el total de individuos y diversidad según el índice de Shannon-Wiener (H') (Shannon & Wiener, 1949; Magurran, 1988; Krebs, 1998), descriptores de la mesofauna de cada tratamiento.
La riqueza de los sitios de ambos usos se tomó como el total de grupos taxonómicos encontrados en todos los muestreos y sitios según el uso.
La biodiversidad se calculó mediante el índice de Shannon-Wiener (H'), para ambos tratamientos y para cada fecha, según la fórmula: H' = -Σ pi * log2 pi,. Donde pi es la abundancia relativa de cada grupo taxonómico, ni/Ni. La comparación de la diversidad entre sistemas se analizó mediante ANOVA de una vía.
La equitatividad se estimó mediante el índice de Pielou (J) según la fórmula: H'/log (S), donde S es la riqueza específica (Magurran, 1988), como una medida de la homogeneidad de distribución de los individuos entre los taxa. Se realizó un PCA para explorar la posible existencia de diferencias entre ambas comunidades. La similitud entre ambas comunidades se comparó utilizando el índice de Jaccard, según la fórmula: Ij = c / (a + b + c), donde a y b son el número de grupos en el sitio a y en el sito b y c es el número de grupos comunes a ambos sitios.
Para comprobar la existencia de posibles diferencias en la estructura de ambas comunidades debido al uso, se determinó el grado de estructuración por unidad muestreal mediante el análisis de varianza multivariado con permutaciones usando matrices de distancia (ADONIS) que prueba la hipótesis de igualdad de medias multivariada entre grupos (Oksanen et al., 2013) y su significancia es evaluada con permutaciones. Esta prueba transforma las abundancias por sitio y por fecha en distancias Bray y Curtis y es análoga al análisis de varianza multivariado basado en la distribución normal y al análisis de varianza multivariado (MANOVA) propuesto por Anderson (2001) que se basa en permutaciones y es similar al análisis de redundancia (Legendre & Anderson, 1999). ADONIS analiza y particiona la suma de cuadrados usando matrices de distancia y disimilitudes basado en un ajuste de modelos lineales y utilizando prueba de permutaciones con seudo-F (Oksanen et al., 2013).
Los análisis estadísticos fueron realizados con el paquete Vegan de R y con el software Past.
RESULTADOS
Abundancia y riqueza de la mesofauna asociada a la hojarasca según uso del suelo
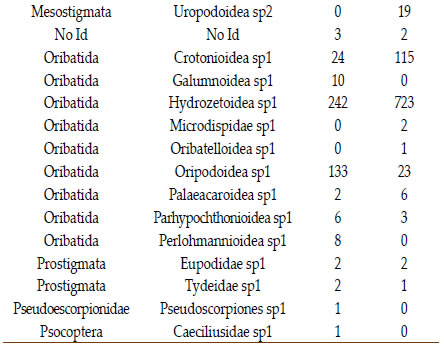
La Tabla 1 muestra los grupos taxonómicos identificados y sus abundancias a lo largo de todos los muestreos.La abundancia total de individuos capturados fue mayor en los sitios de uso agrícola respecto de los sitios de pastizal (1333 vs 533 individuos).
En ambos ambientes dos grupos participan con más del 70% de los individuos. La superfamilia Hydrozetoidea es la más abundante tanto en el pastizal como en el sistema agrícola. La superfamilia Oripodoidea sp1 es la segunda en importancia en el pastizal en tanto que la segunda en importancia para el sistema agrícola es Onychiuridae. Tanto ácaros como colémbolos fueron menos numerosos en el pastizal. En los sitios agrícolas se encontraron un total de 21 grupos taxonómicos, mientras que en los sitios de pastizal se encontraron un total de 26 grupos taxonómicos. Del total de grupos identificados en el pastizal, 12 de ellos no fueron registrados en el otro sistema
Tabla 1.Composicion y abundancia de la mesofauna del suelo para los dos usos de suelo evaluados.
Table 1. Soil mesofauna composition and abundance of the two soil uses tested.

Biodiversidad
El análisis de los índices de diversidad mediante ANOVA entre sitios para las distintas fechas de muestreo arrojó diferencias significativas (Tabla 2) para los muestreos 1, 3 y 4 (p = 0,018; 0,0019 y 0,0031 respectivamente), aunque no mostró diferencias entre sitios cuando se lo calculó para todo el período de muestreos, H' agrícola = 2,36 y H' pastizal = 2,51 (p = 0,6).
Tabla 2. Comparación de los índices de diversidad entre los dos usos de suelo para cada fecha de muestreo. Los valores de H' son el promedio de 5 réplicas por fecha.
Table 2. Diversity index comparisson between soil uses for each sampling date. H'values are the average of 5 replicates per date.

Equitatividad
La equitatividad en ambos sitios se calculó mediante el índice de Pielou (J), para cada sistema, resultado 0,54 para el sistema agrícola y 0,53 para el pastizal.
Análisis de similitud
El índice de Jaccard utilizado, arrojó una similitud del 44,4% entre las comunidades de mesofauna de los suelos de ambos sistemas.
Análisis de Componentes Principales
Los resultados del Análisis de Componentes Principales (PCA) se muestran en la Figura 1. Las dos principales variables sintéticas explican el 64,4% de la varianza total y muestran una muy baja superposición. Mientras que la variable sintética 1 acumula todos los puntos del sistema pastizal en valores negativos, la variable 2 hace lo propio con el sistema agrícola.
Análisis multivariado de varianza con permutaciones (ADONIS)
El análisis ADONIS mostró diferencias en la estructura entre ambas comunidades (p = 0,034) y por fecha de muestreo (p = 0,044) sin interacción significativa entre ambos factores (p = 0,302), Tabla 3.
Figura 1: Gráfico del análisis de componentes principales (PCA) para todos los muestreos de ambos sistemas. Ag: Agricultura, Pa: Pastizales.
Figure 1. Principal Components analysis graph, for all samplings in both systems.
Tabla 3. Comparación de la estructura entre ambas comunidades utilizando ADONIS (1000 permutaciones).
Table 3. Structure comparison between both communities using ADONIS (1000 permutations).


DISCUSIÓN
Los resultados de este trabajo muestran claramente que, sometidas a distinta intensidad de uso, las comunidades de mesofauna edáfica asociadas a la hojarasca no solo difieren en su composición sino que dichas diferencias tienen además una elemento temporal. Por una parte, el análisis de la riqueza de los grupos taxonómicos identificados y de sus abundancias relativas indica la existencia de diferencias entre ambas comunidades edáficas.
Es interesante notar que los índices de Shannon- Wiener resultaron significativamente distintos entre ambas comunidades para tres de las seis fechas de muestreos lo que podría indicar que la dinámica temporal asociada a los cambios estacionales en la temperatura y humedad puede ser un factor que influya diferencialmente en la riqueza y la abundancia de la mesofauna edáfica de ambos sistemas.
Estos resultados, junto con los de las diferencias en la riqueza específica y las abundancias relativas, sugieren que las mismas podrían ser consecuencia en parte, de una distinta dinámica temporal modulada por la distinta intensidad de uso entre ambos sistemas. El análisis PCA separa claramente ambas comunidades. La primera variable sintética agrupa todos los puntos del sitio Agrícola, mientras que la segunda variable hace lo propio con los puntos del sitio Pastizal.
El Análisis multivariado de varianza con permutaciones (ADONIS), muestra claramente que las comunidades de la mesofauna edáfica de dos sitios de un mismo suelo Argiudol sometidos a distinta intensidad de uso son estructuralmente distintas.
Las diferencias encontradas son al menos parcialmente atribuibles a una distinta dinámica temporal asociada a la intensidad de uso y a la oferta de alimento disponible. Esta distinta estructura de las comunidades de artrópodos estudiadas, coincide además, con diferencias significativas en los factores físico-químicos analizados para los mismos sitios por Sandler et al. (2018) y en el funcionamiento de la biota edáfica de los mismos sitios estudiados en este trabajo, tanto en la velocidad de descomposición de la materia orgánica (Castro-Huerta et al., 2015) como en la actividad microbiológica (Di Ciocco et al., 2014).
Los resultados coinciden con los obtenidos por Fraile & Serafino (1978) y de Takeda (1987), que han informado que cambios estacionales en las condiciones ambientales pueden promover explosiones demográficas en los colémbolos. Por otro lado, Arbea & Blasco (2001), asocian estos cambios a diferencias estacionales en la humedad y la temperatura, como factores determinantes del hábitat óptimo de los colémbolos, que influyen en la tasa de reproducción, crecimiento de los individuos y en su distribución vertical a lo largo del perfil. Los resultados obtenidos pueden deberse a que el sitio agrícola estuvo sometido a un sistema de siembra directa desde 18 años antes del inicio de este trabajo. Este sistema de labranza deja típicamente gran cantidad de residuos de cosecha en superficie, los cuales constituyen una fuente de alimento constante a lo largo del año por incorporación gradual de materia orgánica. Asimismo, la cobertura vegetal constante en el sistema intensivo bajo siembra directa, amortigua las variaciones temporales tanto de temperatura como de humedad. Así, los resultados obtenidos pueden deberse a la gran acumulación de restos vegetales en superficie en los sitios agrícolas bajo siembra directa, los cuales crean un ambiente propicio para el desarrollo de mesofauna (Arbea & Blasco, 2001).
Los resultados son consistentes también con lo expresado por Weideman et al. (1982), quienes encontraron que la existencia de un área con cobertura vegetal produce un alto valor de humedad y una reducción de la temperatura, lo que favorece altos valores de densidad y abundancia de la mesofauna. En especial, se puede apreciar que las mayores densidades de ácaros pertenecen al orden Oribatida. Estos ácaros son más abundantes cuando existe una mayor acumulación de hojarasca, debido a que la mayoría de las especies de este grupo son laterícolas y se alimentan de hojarasca, hongos y otros materiales orgánicos (Andrén & Lagerlöf, 1983).
Como se ha dicho, los sitios agrícolas de los cuales se colectaron las muestras de este trabajo, tienen un historial de uso de 40 años de agricultura, siendo los últimos 18 con manejo de siembra directa. Los sistemas conservacionistas tienden a preservar la estructura del suelo, mejorar la disponibilidad de nutrientes y, debido a la presencia de gran cantidad de rastrojo en superficie, a reducir el efecto de la erosión y la pérdida de agua del perfil (Studdert & Echeverría, 2002). Adicionalmente, estas condiciones influyen sobre la dinámica de los microinvertebrados. La presencia de una cubierta vegetal proporciona un ambiente especial continuo y de alimento esencialmente ilimitado tanto en el espacio como en el tiempo para los organismos del suelo. Esto afecta la descomposición de los residuos vegetales directamente a través de la ingestión, trituración y redistribución de la materia orgánica (Castro-Huerta et al., 2015) e indirectamente, actuando sobre la dinámica de los hongos y de las bacterias (Gizzi, 2009; Di Ciocco et al., 2014). De esta manera, los resultados obtenidos apoyan la interpretación de que la acumulación de restos vegetales tuvo una influencia positiva en la diversidad y equitatividad del sistema agrícola bajo siembra directa.
Los resultados aquí presentados son también consistentes con los resultados presentados por Marasas et al. (2001) quienes en un estudio sobre trigo comparando labranza convencional y siembra directa en Argentina, encontraron un porcentaje mayor de detritívoros en cultivos agrícolas con respecto a pastizales naturales y argumentaron que estos resultados responden a sistemas que disponen de mejor calidad y cantidad de materia orgánica la cual se incorpora de manera sostenida en el tiempo.
Los resultados presentados en este trabajo apoyan el hecho de que la siembra directa, realizada correctamente, minimizando el uso de agroquímicos y particularmente dejando gran cantidad de residuo sobre el suelo, es una herramienta agronómica conservacionista de los parámetros físicoquímicos del suelo y su correlato biológico. La protección brindada por la gran cantidad de rastrojo acumulado en superficie, unida al continuo ingreso de materia orgánica al perfil puede beneficiar a las poblaciones de microartrópodos que habitan en los primeros centímetros del perfil del suelo.
Es posible que el aumentar la resolución taxonómica permita también una mejora en los análisis de diversidad e identificar aquellos grupos biológicos que puedan ser utilizados como bioindicadores. Es de destacar sin embargo, que el nivel taxonómico al que se llegó en este trabajo permitió identificar diferencias entre los sistemas. Esto refuerza la creciente idea que la estructura de la comunidad de artrópodos de suelo constituye una respuesta integrada que puede ser utilizada como indicadora de impacto antrópico y que lograr resoluciones taxonómicas mayores podría permitir aumentar el poder de discriminación.
AGRADECIMIENTOS
Este trabajo fue parcialmente financiado por dos subsidios de la ANPCyT (PICT 2006- 02293 y PRH 2007-19), una beca de Iniciación de la CIC y el Departamento de Ciencias Básicas, Universidad Nacional de Luján. Los autores agradecen a los Sres. Edgardo Ferrari, Pablo Peretto y Romina de Luca por facilitar sus campos como sitios de muestreos. Un agradecimiento especial a Loreta Gimenez por su ayuda en los trabajos de campo y de laboratorio, y a la Dra. Gabriela Campana por su ayuda con el análisis ADONIS.
REFERENCIAS
1. Altieri, MA. 1999. Agroecología: Bases Científicas para una Agricultura Sustentable. Editorial Nordam. Montevideo. 10-56. [ Links ]
2. Anderson, MJ. 2001. A new method for non-parametric multivariate analysis of variance. Austral Ecology 26, pp 32-46. [ Links ]
3. Andren, O & L Lagerlöf. 1983. Soil fauna (microarthropods, enchytraeids, nematods) in Swedish agricultural cropping systems. Acta. Agric. Scand., 33: 29-42 [ Links ]
4. Aquino, AM; ME Fernandez & MV Alves. 2008. Diversidade da macrofauna edafica no Brasil. 143-170 En: Moreira FJO Siqueira, L Brussard (Eds.) Biodiversidade do solo em ecossistemas brasileiros. Ed UFLA, Lavras, Brasil. 768 pp. [ Links ]
5. Arbea, J & J Blasco. 2001. Ecología de los Colémbolos (Hexapoda, Collembola) en los Monegros (Zaragoza, España). Aracnet, 7 28: 35-48. [ Links ]
6. Barrios, E. 2007. Soil biota, ecosystem services and land productivity. Ecological Economics 64:269-285. [ Links ]
7. Bedano, JC & A Ruf. 2007. Soil predatory mite communities (Acari: Gamasina) in agroecosystems of central Argentina. USA. Applied soil ecology 36: 22-31. [ Links ]
8. Bellinger, P; K Christiansen & F Janssens. 2006-2012. Checklist of the Collembola of the World. 18/11/19 [ Links ]
9. Benckiser, G. 2018. Fauna in soil ecosystems. Recycling Processes, Nutrient Fluxes, and Agricultural Production. CRC Press, Taylor & Francis Group, Boca Raton, FL. USA [ Links ]
10. Berlese, A. 1905. Apparachio per raccogliere presto ed in gran numero piccoli Artropodi. Redia 2: 85-9. [ Links ]
11. Cabrera G & G Crespo. 2001 Influencia de la biota edáfica en la fertilidad de los suelos en ecosistemas de pastizales. Revista Cubana de Ciencia Agrícola, 1 (35): 3-9. [ Links ]
12. Castro-Huerta, R; LB Falco; RV Sandler, & CE Coviella. 2015. Differential contribution of soil biota groups to plant litter decomposition as mediated by soil use. PeerJ3:e826; DOI 10.7717/peerj.826. [ Links ]
13. Cortet, J; A Gomot-De Vauflery; N Poinsot-Balaguer; L Gomot; C Texier, C & D Cluzeau. 1999. The use of invertebrate soil fauna in monitoring pollutant effects. Eur J Soil- Biol 35: 115-134. [ Links ]
14. Di Ciocco, CA; RV Sandler; LB Falco & CE Coviella. 2014. Actividad microbiológica de un suelo sometido a distintos usos y su relación con variables físico-químicas. Revista de la Facultad de Ciencias Agrarias UNCUYO, 46(1): 73-85. [ Links ]
15. Domínguez, A; JJ Jiménez; CE Ortiz & JC Bedano. 2018. Soil macrofauna diversity as a key element for building sustainable agricultura in Argentne Pampas. Acta Oecologica 92:102-116. [ Links ]
16. Domínguez, A.; JC Bedano & A Becker. 2011. La macrofauna edáfica en agroecosistemas del centro de Córdoba: Agricultura convencional vs. agricultura química. II Congreso Nacional de Ecología y Biología del suelo. [ Links ]
17. Eisenbeis, G. 2006. Biology of soil invertebrates. En: KÖNIG, H & A VARMA. (Eds.) Soil Biology vol. 6: Intestinal microorganism of soil invertebrates. Berlin. Springer-Verlag. 3 - 53. [ Links ]
18. Fraile, M & A Serafino. 1978.- Variaciones mensuales en la densidad de microartrópodos edáficos en un cafetal de Costa Rica. Revista Biología Tropical, 26: 291-301. [ Links ]
19. Gizzi, A. 2009. Caracterización de la meso y macrofauna edáfica en sistemas de cultivos del sudeste bonaerense. Ciencia del Suelo 27(1): 1-9. [ Links ]
20. Gupta, VVSR. 1994. The impact of soil and crop management practices on the dynamics of soil microfauna and mesofauna. En: CE Pankhurst; BM Doube; VVSR Gupta & PR Grace (eds.). Soil biota: management in sustainable farming systems. 107-124. CSIRO, East Melbourne. [ Links ]
21. Hendrix, PF; JR Crossley; JM Blair & DC Coleman. 1990. Soil Biota as components of sustainable agroecosystems. Sustainable Agricultural Systems, Soil and Water Conservation Society, Ankeny, Iowa. [ Links ]
22. INTA (Instituto de Tecnología Agropecuaria). 2014. Carta de suelos de la Provincia de Buenos Aires. Fuente de internet: https://inta.gob.ar/documentos/carta-de-suelos- de-la-provincia-de-buenos-aires. 10/05/19. [ Links ]
23. Kethley, J. 1990. Acarina: Prostigmata (Actinedida). En: DINDAL, D. (Editor). Soil Biology Guide. New York. John Wiley & Sons. 667 - 756. [ Links ]
24. Krantz, G & B Ainscough. 1990. Acarina: Mesostigmata (Gamasida). En: DINDAL,D. (ed). Soil Biology Guide. New York. John Wiley & Sons. 583 - 665. [ Links ]
25. Krebs, C. 1998. Ecological methodology. Wiley and Sons. New York, USA. 620 pp. [ Links ]
26. Labrador, J. 1996. La Materia Orgánica en los Agroecosistemas. Ministerio de Agricultura, Pesca y Alimentación. Mundi-Prensa. España. 29-35. [ Links ]
27. Lavelle, P & A Spain. 2003. Soil Ecology. Kluwer Academic Publishers. New York. 25 - 100. [ Links ]
28. Legendre, P & MJ Anderson. 1999. Distance-based redundancy analysis: testing multispecies responses in multifactorial ecological experiments. Ecol. Monographs 69(1): 1-24. [ Links ]
29. Lenardon, A. 2010. Agriculturización. Efectos sobre el suelo y el agua. Revista ECO ciencia y naturaleza. 21: 10-12. [ Links ]
30. Lindo, Z & S Visser. 2003. Microbial biomass, nitrogen and phosphorus mineralization, and mesofauna in boreal conifer and deciduous forest floors following partial and clear-cut harvesting. Canadian Journal of Forest Research. 33 (9): 1610-1620. [ Links ]
31. Magurran A. 1988. Ecological diversity and its Measurement. Princeton University Press. Princeton, 192 p. [ Links ]
32. Marasas, ME; SJ Sarandon & AC Cicchino. 2001. Changes in soil arthropod functional group in a wheat crop under conventional and no tillage systems in Argentina. Applied Soil Ecology 18: 61-68. [ Links ]
33. Menta, C; A Leoni; C Gardi & FD Conti. 2011. Are grasslands important habitats for soil microarthropod conservation? Biodiversity Conservation 20:1073-1087. [ Links ]
34. Norton, R. 1990. Acarina: Oribatida. In: DINDAL, D. (Editor). Soil Biology Guide. New York. John Wiley & Sons. 779 - 803. [ Links ]
35. Oksanen, J; FG Blanchet; R Kindt; P Legendre; PR Minchin; RB O'Hara; GL Simpson; P Solymos; MH; Stevens & HH Wagner. 2013. Vegan: Community Ecology Package. R PackageVersion. 2.0-10. https://github.com/vegandevs/ vegan. 10/05/19. [ Links ]
36. Sandler, RV; LB Falco; CA Di Ciocco; R Castro-Huerta & CE Coviella. 2018. Change of collembolan (Hexapoda: Collembola) community structure related to antrhopic soil disturbance. Revista de la Facultad de Ciencias Agrarias UNCUYO 50(1):217-231. [ Links ]
37. Schaefer, M. 1995. Interspecific interactions in the soil community. Acta Zoologica Fennica 196: 101 - 106. [ Links ]
38. Shannon, C & W Weaver. 1949. A Mathematical Model of Communication. University of Illinois Press, Urbana, IL. 117 [ Links ]
39. Studdert, GA & HE Echeverría. 2002. Soja, girasol y maíz en los sistemas de cultivo del sudeste bonaerense. 413- 443 en Andrade, FH & V Sadras (eds) Bases para el manejo del maíz, el girasol y la soja (2ed.). INTA - Facultad de Ciencias Agrarias (UNMP). Balcarce. [ Links ]
40. Takeda, H. 1987. Dynamics and maintenance of collembolan community structure in a forest soil system. Res. Pop. Ecol., 29: 291-346. [ Links ]
41. Tullgren, A. 1918. Einsehreinfacher AusleseapparatfurterricoleTierfaunen. Z. angew. Ent. 4: 149-50. [ Links ]
42. Wallwork, JA. 1983. Oribatids in forest ecosystems. Annual Review of Entomology 28: 109 - 130. [ Links ]
43. Walter, D & H Proctor. 2008. Soil Microarthropods. Lucid Keys, http://keys.lucidcentral.org/keys/cpitt/public/Mites/ Microarthropods/Index.htm. 10/05/19. [ Links ]
44. Weidemann, G; H Koehler & T Schriefer. 1982. Recultivation: a problem of stabilization during ecosystem development. En: R. Bornkamm, J. A. Lee & M. R. D. Seaward (eds.), Urban Ecology, Blackwell, Oxford, 305-313. [ Links ]

