










Servicios Personalizados
Revista

Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkIntersecciones en antropología
versión On-line ISSN 1850-373X
Intersecciones antropol. n.4 Olavarría ene./dic. 2003
Pastoreo y caza de camélidos en el Temprano de la Puna de Salta: Datos osteométricos del sitio Matancillas 2
Gabriel E. J. López
Gabriel E. J. López. Sección Prehistoria, ICA-FFyL (UBA), 25 de Mayo 217, 3er piso, Buenos Aires, Argentina.
RESUMEN
Los camélidos fueron los recursos faunísticos más importantes para las poblaciones puneñas a lo largo de toda su historia cultural. Así lo indican los registros arqueofaunísticos de la mayoría de los sitios analizados en el Noroeste argentino. Para el Temprano (2500-1350 AP.), la existencia de una diversificación de las estrategias económicas no habría impedido seguir consumiendo mayoritariamente camélidos; más aún, las estrategias pastoriles y cazadoras se habrían centrado en estos recursos. Los datos osteométricos obtenidos de los conjuntos arqueofaunísticos provenientes del sitio Matancillas 2 están de acuerdo con estas consideraciones, ya que se registró la presencia de especímenes correspondientes a llama y vicuña, lo que indicaría una economía con pastoreo y caza. Siguiendo las predicciones de modelos de manejo del riesgo, es posible que la diversificación citada minimizara las fluctuaciones ambientales a las que estaban sometidas las poblaciones de la Puna.
ABSTRACT
Camelids were the most important faunal resource for the human populations of the Puna highlands throughout their cultural history in the Argentinean Northwest, as is demonstrated by the archaeofaunal record. During the Temprano period (2500- 1350 BP.), the existence of diversification in economic strategies did not prevent the consumption of camelids predominating. Even most pastoralists and hunters' strategies were centered on these resources. The osteometrical data obtained from the archaeofaunal assemblages of Matancillas 2 site shows the presence of specimens pertaining to llama (a domesticated species) and vicuña (a wild species), suggesting both pastoralism and hunting strategies for the supply of meat. Following the predictions of risk-sensitive models, the combining of these strategies can be explained as a risk-minimization tactic based on the diversification of economic activities to minimize fluctuations in the resource base to which the human populations of this region were exposed.
INTRODUCCIÓN
El objetivo de este trabajo es discutir las estrategias económicas implementadas en cuanto al consumo de camélidos por parte de los pobladores del sitio Matancillas 2 durante el Temprano de la Puna de Salta, contribuyendo al estudio de la diversidad de las adaptaciones humanas para este período. El Temprano abarca un lapso que va del 2500 al 1350 AP. aproximadamente y se caracteriza por la presencia de grupos agropastoriles y alfareros (González 1963, 1977).
La Puna, como todo desierto de altura (más de 3000 m snm.) es un ambiente de alto riesgo (sensu Winterhalder et al. 1999), debido a las fluctuaciones impredecibles en la disponibilidad de los recursos. Esta región se caracteriza por la bajísima productividad primaria; la distribución heterogénea en el espacio de los nutrientes; la alta estacionalidad y, principalmente, la variación impredecible de la caída de precipitaciones (Muscio 1998-1999; Yacobaccio et al. 1994). Cada uno de estos factores pueden tener incidencia en las adaptaciones humanas a los entornos locales puneños.
La vegetación está compuesta especialmente de tolar (Parastrephia) y gramíneas de distintos géneros (Ruthsatz y Movia 1975). Por su parte, la diversidad de recursos faunísticos es muy baja, destacándose los camélidos por su alto rendimiento económico.
Los camélidos pueden ser domesticados como la llama (Lama glama) y la alpaca (Lama pacos) y silvestres como la vicuña (Lama vicugna) y el guanaco (Lama guanicoe). La alpaca no habita en la actualidad en el Noroeste Argentino y su presencia no ha sido determinada en épocas prehispánicas, quizás debido a la preferencia por sectores ambientales más húmedos (Olivera 1997).
Durante el Temprano, las evidencias arqueológicas sobre las estrategias económicas desarrolladas en la Puna registran una marcada variabilidad de escala local. En este sentido, existen indicadores del consumo de recursos a través de la caza, la recolección, el pastoreo y la agricultura, ya sea con el predominio de determinadas estrategias, o directamente con la presencia/ausencia de algunas de ellas. La existencia de una estrategia económica que predomina sobre el resto (Yacobaccio et al. 1997-1998) es la que determina la organización de todas las actividades, entre ellas, tecnología, movilidad, asentamiento, etc. De este modo, por ejemplo, en los grupos donde el pastoreo predomina, la movilidad y las tácticas de asentamiento se relacionan a la disponibilidad de pasturas, mientras que para el desarrollo agrícola es fundamental poseer tierras fértiles aptas para tal fin.
En la Puna Meridional, los casos de Casa Chávez (Olivera 1991) y Tebenquiche (Haber 2001a, 2001b; Krapovickas 1955), evidencian la presencia de grupos con estrategias agropastoriles. Una situación similar, se registra en la Puna Norte, en los sitios próximos al río de las Burras (Fernández Distel 1998). Sin embargo, en Susques -también Puna Norte-, hacia el 2000 AP., la estrategia que predomina sería el pastoreo con un aporte importante de la caza, pero sin agricultura (Yacobaccio et al. 1997-1998).
A partir de la lógica de la ecología evolutiva (Smith y Winterhalder 1992), la diversidad local en las estrategias económicas de la Puna, se relacionaría con la incidencia de factores ecológicos tales como la heterogeneidad en la distribución espacial de los nutrientes necesarios para la subsistencia humana, la calidad de los recursos y las fluctuaciones impredecibles en la caída de precipitaciones que afectan la abundancia de los mismos. En otras palabras, la variabilidad ambiental de la Puna explicaría las distintas estrategias económicas desarrolladas por las poblaciones locales.
Los estudios presentados en este trabajo provenientes de la Quebrada de Matancillas, en la Puna de Salta, realizan el aporte de un nuevo caso de estudio a la diversidad de las adaptaciones humanas puneñas durante el Temprano. La Quebrada de Matancillas, se ubica lateralmente al valle de San Antonio de los Cobres (SAC), con una altitud aproximada de 3600 m snm. En esta región, las quebradas laterales como Matancillas, ofrecen la mayor productividad ecológica y protección contra agentes metereológicos (Muscio 2003a). Matancillas se encuentra surcada por el curso de agua principal del mismo nombre, que actualmente permite la horticultura con riego (agricultura de baja escala) y la disponibilidad de pasturas para el pastoreo de camélidos, ovicápridos y vacunos. La Quebrada de Matancillas puede ser considerada una Zona de Concentración de Nutrientes (sensu Yacobaccio 1994) debido a la alta potencialidad que posee para la subsistencia humana (en este caso vinculada al desarrollo hortícola pastoril), derivada de la oferta altamente localizada de agua.
El sitio arqueológico Matancillas 2, fechado en ca 2000 AP. es el foco de análisis en este trabajo. Se encuentra en una ladera de piedemonte, elevada 3,70 m del nivel del curso de agua y está compuesto por estructuras de formas mayormente circulares y subcirculares, conformadas por lajas paradas. Las unidades arquitectónicas que componen el sitio se encuentran agrupadas formando un patrón aglomerado en la ladera de piedemonte, mientras que en el fondo de quebrada se ubican los terrenos aluvionales aptos para la horticultura.
Los datos obtenidos de plantas cultivadas, artefactos de molienda, azadas líticas y fundamentalmente la ubicación del sitio Matancillas en una quebrada protegida con tierras fértiles para el desarrollo agrícola, permiten plantear que la horticultura (agricultura de baja escala) podría ser la estrategia principal, a partir de la cual se seleccionó y organizó la ocupación y el asentamiento en la Quebrada de Matancillas (Muscio 2003b). Sin embargo, aquí se aborda el estudio de otras estrategias económicas de suma importancia, específicamente pastoreo y caza, que en conjunto con la horticultura conformarían el nicho económico de la población local del Temprano de Matancillas. Al respecto, existen evidencias contextuales en el sitio, tales como estructuras de gran diámetro (superiores a 10 m) adscribibles a corrales para el cautiverio de los rebaños y una marcada diversidad de puntas de proyectil líticas, que podrían estar relacionadas a la caza de animales silvestres. Pero más allá de estos elementos contextuales el principal indicador del uso del pastoreo y la caza está dado por la existencia de camélidos domesticados y silvestres en los registros arqueofaunísticos.
Por este motivo, el aporte a la discusión sobre este tema, se realiza fundamentalmente a través de estudios osteométricos en las arqueofaunas del sitio Matancillas 2, para diferenciar de una manera preliminar las especies de camélidos representadas. En consecuencia, los datos obtenidos en este trabajo contribuyen al conocimiento de las estrategias de uso de recursos faunísticos que formaron parte del nicho económico de la población local de Matancillas.
Algunas consideraciones teóricas
Desde la ecología evolutiva se postula que los factores ecológicos tienen una alta influencia sobre la toma de decisión humana en cuanto a las estrategias de subsistencia desarrolladas (Smith y Winterhalder 1992). En este sentido, en la región de estudio -la Punadebe considerarse especialmente el alto riesgo ambiental, debido principalmente a las fluctuaciones impredecibles de las precipitaciones, que pueden producir sequías con resultados catastróficos para los recursos animales y vegetales. Para los casos en los cuales está involucrada la producción de alimentos en los Andes centro-sur, diversos autores señalaron la importancia de diferentes estrategias que darían seguridad en el manejo del riesgo (Browman 1994; Escola 1996; Gobel 1994). Entre ellas, pueden mencionarse el incremento de la capacidad de sustentación, que incluye sistemas de irrigación y estrategias especializadas de pastoreo para cada tipo de ganado; diversificación de las actividades productivas a través del uso de distintos cultígenos, parcelas y especies animales; movilidad para el acceso a diferentes pasturas o parcelas; almacenamiento a través de una tecnología adecuada para llevarlo adelante y, por último, mecanismos sociales de cooperación basados generalmente en lazos parentales de reciprocidad, vinculados especialmente al riesgo laboral.
En consecuencia, las distintas estrategias desarrolladas a partir del manejo del riesgo podrían reflejar una alta variabilidad de combinaciones posibles en cada una de las diferentes poblaciones locales. En la Puna, como se señaló en la introducción, existen sobradas evidencias de esta variabilidad local. Sin embargo, en casi todos los casos, tal como dicen Winterhalder et al. (1999), las estrategias diversificadoras serían las más importantes en la minimización del riesgo. Al respecto pueden considerarse estrategias tales como ampliar la dieta; diversificar el uso del espacio en cuanto a pasturas, parcelas, etc.; diversificar las actividades económicas (por ejemplo a través de la utilización del pastoreo, la agricultura y la caza en forma conjunta) y compartir e intercambiar fuentes de recursos no sincrónicas en el espacio. Es importante recalcar que estas estrategias no tienen por qué darse todas juntas, aunque la mayoría de ellas parecen haber sido empleadas durante el Temprano de la Puna.
Estas predicciones pueden derivarse de modelos formales como el Z score (Winterhalder y Goland 1997; Winterhalder et al. 1999). No es el objetivo de este trabajo realizar una descripción detallada del mismo (para su formulación y aplicación al caso de estudio ver López 2002a, 2002b) sino simplemente aclarar que este modelo considera niveles de energía mínimos y máximos, umbrales a partir de los cuales se establecen las distintas posibilidades para adaptarse a un ambiente (Lanata y Borrero 1994). A su vez, predice dos estrategias fundamentales de manejo del riesgo: ser propenso (risk prone) o adverso al riesgo (risk averse) (Stephens y Krebs 1986). La primera desarrolla estrategias que tienden hacia la especialización y no dan la seguridad de superar el requerimiento mínimo necesario para adaptarse a un ambiente; mientras que la segunda desarrolla generalmente estrategias diversificadoras, que llevan a minimizar el riesgo, dando la seguridad de no caer debajo del umbral adaptativo, aunque no sean las de mayor retorno (Winterhalder et al. 1999). Esto significa que la diversificación en sus distintas formas, tendería a minimizar el riesgo. Por ejemplo, el uso conjunto del pastoreo y la caza por parte de una población humana, sería una estrategia adversa al riesgo, que promovería la disminución de la varianza (el desvío estándar) en las variables del Z score (ver Winterhalder et al. 1999).
Por lo tanto, a partir del modelo Z score puede plantearse que la especialización no es una buena estrategia en ambientes tan fluctuantes como el de la Puna sino, por el contrario, tiene muy bajas expectativas de superar el umbral adaptativo. Mucho menos cuando existe una alta dependencia de estrategias agrícolas, tal como parecería en el caso de estudio (Muscio 2003b). En las estrategias productoras de alimentos, este umbral debe calcularse para la unidad doméstica en su conjunto, ya que según Winterhalder y Goland (1997), es la unidad de producción y consumo sobre la que el riesgo operaría. De esta forma, el manejo del riesgo se produce a partir de la toma de decisiones en la unidad doméstica, más que en el individuo. Para el caso de Matancillas, en este nivel de análisis, estudios actualísticos comenzaron a estimar el requerimiento adaptativo mínimo en relación a las kilocalorías diarias (Muscio 2003a).
Siguiendo el modelo Z score, la expectativa es que el manejo del riesgo de los pobladores de Matancillas durante el Temprano habría implicado una diversificación de las estrategias económicas. Entre ellas, el uso conjunto del pastoreo y la caza habría sido fundamental para asegurar el consumo de la proteína animal necesaria. Por lo tanto, la relación patoreo-caza se entiende dentro de un nicho económico diversificador, orientado a la minimización del riesgo del conjunto de la subsistencia.
Como se señaló anteriormente, para el Temprano del Noroeste Argentino y, más específicamente en la Puna, existen fuertes evidencias del uso conjunto del pastoreo y la caza por parte de las poblaciones locales. En casi todos los casos, los camélidos representan los recursos faunísticos principales. Estos artiodáctilos habrían sido protagonistas de una relación coevolutiva con los humanos (Olivera 1998), entendiendo por coevolución a la relación entre dos especies genéticamente no relacionadas que evolucionan conjuntamente (Rindos 1980).
Respecto del pastoreo y la caza debe plantearse claramente que son estrategias económicas con objetivos diferentes. En una estrategia pastoril, el objetivo es la conversión de biomasa vegetal (pasturas) en biomasa animal para el consumo humano y el pastor considera los recursos como un capital que acumula y reproduce (ver Chang y Koster 1986). Por el contrario, la estrategia cazadora tiene como objetivo la obtención directa de biomasa animal y el cazador considera a los animales como un alimento ya presente en el ambiente que explota.
Los animales domesticados requieren el control humano de su reproducción, alimentación y organización del territorio, al contrario de los animales silvestres (López y Medina 2001). En consecuencia, el pastoreo suma costos de manejo con respecto a la caza, pero a su vez anula los costos de búsqueda (ver López 2002b). Desde un punto de vista teórico, estas variables que afectan a la relación costo-beneficio del aprovisionamiento de proteína animal, deben ser tenidas en cuenta para el estudio de la convivencia entre pastoreo y caza en un contexto socioecológico particular (Layton et al. 1991; Winterhalder y Goland 1997). El objetivo de este trabajo no es determinar la incidencia de cada una de estas estrategias económicas sino identificar en el registro arqueofaunístico la presencia o ausencia de las mismas.
Evidencias arqueofaunísticas del consumo de camélidos y estrategias económicas relacionadas al mismo
En la cueva de Huachichocana III, Puna de Jujuy (Fernández Distel 1974, 1986), la capa del Precerámico Temprano registró un predominio absoluto de camélidos (Yacobaccio y Madero 1992); mientras que el sitio Quebrada Seca 3 en la Puna Sur (Prov. de Catamarca), para esos momentos también mostró una mayoría de especímenes de estos artiodáctilos (Aschero et al. 1991). Esto indica que desde comienzos del Holoceno, el hombre empezaba a mostrar cierta preferencia sobre los camélidos, obtenidos a través de la caza.
Entre los 5000 y los 3000 AP., las frecuencias arqueofaunísticas de casi todos los sitios de la Puna, registran alrededor de un 90% de especímenes de camélidos sobre otros taxones (ver Olivera 1997;Yacobaccio 1997). Este período marca las primeras evidencias claras de la domesticación de camélidos; tal es el caso de Huachichocana III, que hacia los 3400 AP., presenta una inhumación humana, asociada a una cabeza de llama (Elkin et al. 1991).
Para el Temprano (2500 a 1350 AP.), la información arqueofaunística es abundante. En la Puna Sur (Antofagasta de la Sierra, Prov. de Catamarca), el sitio Casa Chávez Montículos, datado entre 2400 y 1300 AP., fue caracterizado como una base residencial de actividades múltiples, con una economía principalmente agropastoril (Olivera 1991). Los conjuntos arqueofaunísticos evidenciaron un 90% de especímenes de camélidos, con un predominio de llamas, (Olivera 1991, 1997), lo que indicaría el fuerte desarrollo del pastoreo. Sin embargo, en una quebrada de altura (también en Antofagasta de la Sierra), el sitio Real Grande I (alero), registró una preponderancia de especímenes de vicuñas entre los camélidos, los cuales representaron más del 90% en NISP (Olivera y Elkin 1994). Esto demostraría que la caza continuó siendo importante enel Temprano de la Puna Sur, a pesar del establecimiento del pastoreo como estrategia económica.
En la Puna Norte, más específicamente en la zona de Susques (Prov. de Jujuy), la economía habría sido pastoril-cazadora, con la primera como estrategia predominante y no habría existido un desarrollo agrícola como en Antofagasta de la Sierra. Aproximadamente hacia el 2040 AP., se habría producido la estabilización del pastoreo como estrategia de subsistencia en la región (ver Yacobaccio et al. 1997-1998). Los sitios de Susques, mayormente aleros como Unquillar y Huirunpure, registraron una mayoría de especímenes de camélidos tanto por NISP como por MNI. A su vez, los especímenes de Huirunpure medidos osteométricamente correspondieron en un 50% a llamas y otro 50% a vicuñas, lo que implicaría la existencia de pastoreo y caza como estrategias económicas (ver Yacobaccio et al. 1997-1998).
A los mencionados sitios de la Puna, pueden agregarse otros que son próximos temporal y espacialmente. En el Valle Calchaquí Norte (Prov. de Salta), se encuentra una aldea del Temprano, denominada Campo Colorado (Tarragó 1980). Allí, el registro arqueofaunístico también mostró una predominancia de camélidos en NISP (más del 95%). Asimismo, en la Quebrada del Toro, una quebrada de acceso a la Puna de Salta, las arqueofaunas de aldeas como Las Cuevas, Cerro El Dique y Potrero Grande, registraron una mayoría absoluta de camélidos, ya que más de un 75% de los individuos representados pertenecieron a esta familia, determinándose también la presencia de llama y vicuña (ver Raffino et al. 1977).
A la mención precedente sobre los estudios realizados por distintos investigadores, se aportará un nuevo trabajo con datos sobre el consumo de camélidos y las estrategias económicas desarrolladas al respecto durante el Temprano de la Puna de Salta.
ARQUEOFAUNAS DE MATANCILLAS
Las unidades de análisis consideradas aquí corresponden a sectores de basural, recinto e interrecinto del sitio Matancillas 2 (para más detalles ver López 2002a; Muscio 2001; Muscio y López 2002). Las arqueofaunas recuperadas de cada uno de estos sectores, metodológicamente, fueron asignadas a conjuntos distintos (Muscio y López 2002).
En primer lugar, se considerará brevemente la integridad de los conjuntos arqueofaunísticos provenientes de estas unidades. En cuanto a la meteorización, siguiendo la escala de Behrensmeyer (1978), los restos óseos presentaron un desigual estado de preservación. Los huesos del basural y del recinto tuvieron una baja meteorización, con más del 80% no meteorizados; mientras que en el sector interrecinto casi la mitad de los huesos estuvieronmeteorizados y más de un 8% en estadios de más de 2.
Por otra parte, las marcas de carnívoros y roedores fueron mínimas en todos los conjuntos y la preservación de los mismos tampoco se vio afectada por la densidad global ósea, según la propuesta de Elkin y Zanchetta (1991) (para un análisis más extenso ver López 2002a).
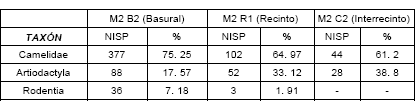
En relación al número y frecuencia de los taxones representados en los tres conjuntos arqueofaunísticos analizados, se registró una predominancia absoluta de especímenes de camélidos. En el conjunto del basural M2 B2, el NISP de camélidos superó el 75%; mientras que en los conjuntos del recinto 1 (M2 R1) y del sector interrecinto (M2 C2), los camélidos representaron más del 60% del NISP total (ver Tabla 1). Debe aclararse que los roedores tuvieron una bajísima presencia y que muchos especímenes fueron asignados a artiodáctilos, aunque en realidad serían camélidos, ya que no se detectó la presencia de cérvidos en los conjuntos.
Tabla 1. Cálculo del NISP en los tres conjuntos arqueofaunísticos del sitio Matancillas 2.
A partir de estos resultados, cabe preguntarse qué especies de camélidos están representadas. Esta no es una pregunta más, ya que en la introducción se señaló la importancia de conocer las estrategias económicas desarrolladas en cuanto al uso de camélidos. La posibilidad de identificar camélidos domesticados y/o silvestres se vincula directamente con dicha cuestión. Una vía posible para responder a la pregunta planteada podría brindarla la realización de análisis osteométricos.
OSTEOMETRÍA
Los análisis osteométricos son fundamentales para determinar la adscripción de especímenes óseos arqueológicos a distintas especies, principalmente dentro de una misma familia (e.g., Camelidae). Para el caso de estudio, la osteometría es una vía de análisis que permite diferenciar entre especies de camélidos domesticados y silvestres. Esto resulta sumamente importante para determinar la existencia de pastoreo y caza.
Como la osteometría se basa en el tamaño de los huesos de las distintas especies, se deben comparar las medidas de camélidos actuales (tomadas como estándar) con las realizadas en especímenes arqueológicos. Por eso, debe considerarse que el gradiente de tamaño en los camélidos que habitan el Noroeste Argentino (de menor a mayor) es vicuña-guanaco-llama (Elkin et al. 1991). La alpaca, como se dijo anteriormente, es una especie de camélido domesticado que no habita ni habría habitado la región. Por lo tanto, la llama sería el único animal domesticado presente en la Puna Argentina hace 2000 años.
Para ser sometidos a osteometría los huesos deben poseer los puntos de medición necesarios; tener buen de estado de conservación y mostrar madurez osteológica (estar fusionados) (ver Elkin et al. 1991).
En el sitio Matancillas 2, debido a estos condicionantes, sólo pudo ser medida una porción de especímenes de los tres conjuntos. Los elementos medidos corresponden a falanges, calcáneos y escápulas, todos del esqueleto post-craneal. La determinación de los puntos osteométricos y la forma de medirlos se obtuvo de la lista elaborada por Elkin (1996, apéndice 5.1), basada principalmente en los criterios de Kent (1982) y Von den Driesch (1976).
Los valores obtenidos para cada especimen fueron comparados a través de la técnica de Meadow (1987), basada en la diferencia de logaritmos, cuya fórmula es:
(dl) = log x - log z = (log x/z)
La fórmula expresa las distancias logarítmicas entre la medida (una sola) del especimen arqueológico (x) y la medida estándar del camélido actual (z). A pesar de haberse realizado la mayoría de las mediciones en los distintos puntos osteométricos de cada especimen, para poder comparar por las distancias logarítmicas se seleccionó una sola medida de cada hueso, que intentó ser discriminante y representativa a la vez. Esto también permitió evitar los sesgos que generan el desigual grado de fragmentación y la variabilidad entre las distintas mediciones de un mismo hueso. Para las falanges 1 y 2 se tomó el ancho máximo de la superficie articular proximal; para la escápula, el ancho de la cavidad glenoidea; y para el calcáneo, el largo máximo.
Sin embargo, la técnica de Meadow y los estudios osteométricos en general, no carecen de problemas. El principal de ellos es no poder controlar adecuadamente la variabilidad intra e interespecie debido a que se compara con el estándar de un sólo individuo. Por lo tanto, los resultados obtenidos con esta técnica, por el momento, deben ser considerados como una aproximación probable respecto de la distinción de especies de camélidos.
RESULTADOS
Tal como se observa en la Figura 1, los valores obtenidos de las mediciones osteométricas sobre los especímenes arqueológicos fueron comparados con el estándar de guanaco actual, a través de la diferencia de logaritmos. Se eligió como medida comparativa al guanaco, debido a que tiene un tamaño intermedio entre la vicuña y la llama. El ejemplar de guanaco que sirve como estándar actual proviene de las Cumbres Calchaquíes y fue medido por D. Elkin y G. Mengoni Goñalons (com. pers., marzo 2003). Este mismo estándar ya fue utilizado en otros trabajos (Elkin et al. 1991; Elkin 1996;Yacobaccio et al. 1997-1998).

Figura 1. Comparación de los valores obtenidos de la diferencia de logaritmos sobre las mediciones osteométricas realizadas (n=20), en los tres conjuntos analizados, M2 B2 (Basural), M2 R1 (Recinto) y M2 C2 (Interrecinto).
En la Figura 1 se observa la comparación entre los resultados logarítmicos para cada conjunto arqueofaunístico, M2 B2 (Basural), M2 R1 (Recinto) y M2 C2 (Sector Intenrrecinto). Las mediciones corresponden al ancho máximo de la cara articular proximal de la primer falange, en los tres conjuntos; los resultados de la misma medida, pero en las segundas falanges, provenientes de los conjuntos del basural y del recinto 1; el largo máximo del calcáneo (hueso del tarso) también en el basural y el recinto 1; y el valor del ancho de la cavidad glenoidea de la escápula, en el conjunto del basural.
Resultó oportuno seleccionar una sola dimensión osteométrica por especimen, para poder comparar a través de las distancias logarítmicas y evitar la variabilidad que generan los valores tomados en todos los puntos osteométricos disponibles. Los más alejados hacia la izquierda se corresponderían a vicuñas; mientras que los más alejados hacia la derecha, podrían ser asignados a llamas (ver Elkin 1996). Sin embargo, existen valores que oscilan entre 0.01 y 0.02, los cuales son bastante aproximados al estándar de guanaco. Pero estos valores podrían superponerse con los de llama. Por lo tanto, en estos casos se prefirió asignar a los especímenes como "llama-guanaco". Si bien se tiene en cuenta que ésta no es una morfoespecie reconocida, esta categoría sirve para expresar el rango en el que se superponen los valores obtenidos de estas mediciones, pudiendo ser llamas o guanacos, por lo cual su significado no es taxonómico.
Así, en el conjunto arqueofaunístico del basural, siguiendo los resultados de la diferencia de logaritmos, de 14 especímenes medidos, 7 corresponderían a llamas, 1 a llama-guanaco y 6 a vicuñas. En forma más específica, la falange 1 mostró 2 especímenes con estándares de llama, 1 en el que se superponía llama y guanaco y otro de vicuña. La falange 2, registró 5 especímenes de llama y 3 de vicuña; mientras que el calcáneo y la escápula pertenecerían a vicuñas. En porcentajes, sobre un número de 14 especímenes medidos, se estableció la presencia de un 50% de llamas, un 7.1% de llama-guanaco y 42.9% de vicuñas (ver Figura 2).
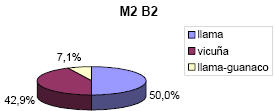
Figura 2. Cuantificación de especies de camélidos determinadas por osteometría en el conjunto Matancillas 2 Barranca 2 (Basural).
En el conjunto del recinto 1, sólo 5 especímenes pudieron ser sometidos a osteometría. Las falanges primeras fueron asignadas una a llama y otra a vicuña; mientras que las falanges segundas, corresponderían una a llama y otra a llama-guanaco, esta última igual que el calcáneo. Por lo tanto, siguiendo la técnica de Meadow (1987), se registró un 40% de especímenes correspondientes a llama, un 40% a llama-guanaco y un 20% a vicuña (ver Figura 3).

Figura 3. Cuantificación de especies de camélidos determinadas por osteometría en el conjunto Matancillas 2 Recinto 1.
Del sector interrecinto, se pudo medir sólo una falange primera. Evidentemente, los valores se acercaron claramente a los de llama.
Por el momento, en Matancillas, la osteometría permitió asignar tentativamente especímenes a llamas y a vicuñas, o sea camélidos domesticados y silvestres, respectivamente. La presencia de guanaco no ha sido determinada certeramente, lo cual no significa descartarla.
Al sumar los especímenes medidos en los tres conjuntos, la llama predomina sobre la vicuña. En total se evidencia 55% de llamas, 35% de vicuñas y 15% de llama-guanaco, asignada así por su carácter dudoso (ver Figura 4).

Figura 4. Cuantificación de especies de camélidos determinadas por osteometría en los tres conjuntos analizados.
DISCUSIÓN Y CONCLUSIONES
La principal limitación en el uso de la técnica de Meadow para los valores osteométricos es la escasa cantidad de mediciones realizadas sobre especímenes de camélidos actuales para determinar la variación en el tamaño de los huesos correspondientes a individuos de una misma especie (específicamente el caso de guanaco que sirve como estándar), como así también de especies distintas. En consecuencia, los estándares actuales necesitan ampliarse para analizar la variación poblacional. Sin embargo, aún con estas limitaciones la técnica es ilustrativa de una tendencia en la representación de distintas especies de camélidos.
Bajo los límites de la técnica aquí seguida, a partir de los resultados osteométricos registrados en los conjuntos arqueofaunísticos del sitio Matancillas 2, se infiere que la población que ocupó este sitio durante el Temprano, consumió preferentemente camélidos, obtenidos a través del pastoreo y la caza. La probable presencia de distintas especies de estos artiodáctilos, una domesticada -la llama- y otra silvestre -la vicuña apoya fuertemente la existencia de una diversificación de las estrategias económicas en cuanto al consumo de recursos faunísticos.
A pesar de que las llamas están cuantitativamente más representadas que las vicuñas, no se puede afirmar aún que el pastoreo era más importante que la caza, debido a que una porción pequeña (n=20) de los especímenes cumplió con los requisitos para ser medidos osteométricamente. Este tamaño de la muestra es comparable con el de otros sitios analizados en la Puna Norte, por ejemplo los de la zona de Susques (Yacobaccio et al. 1997-1998). Sin embargo, es factible que el pastoreo haya sido una estrategia económica fundamental en esos momentos, quizás de mayor importancia que la caza, lo cual debería seguir investigándose en el futuro, mediante técnicas que permitan medir la incidencia de ambas estrategias.
En Matancillas, la horticultura pudo haber sido la estrategia principal, a partir de la cual se organizó la ocupación de la quebrada (ver Muscio 2003b). Por lo tanto, considerando el consumo de vegetales y los resultados obtenidos de las arqueofaunas, que indican el desarrollo de estrategias pastoriles y cazadoras, se podría plantear para Matancillas la existencia de un nicho económico compuesto de horticultura-pastoreo-caza.
En el marco de la diversidad de las adaptaciones de la Puna para el Temprano, en donde precisamente lo que más varía es el desarrollo de la agricultura, el caso de Matancillas se correspondería con las particularidades del hábitat local, apto para el desarrollo de las distintas estrategias diversificadoras de la base económica. Más generalmente el ambiente puneño es de alto riesgo, por lo que el manejo del mismo, podría haber implicado un diseño orientado a la diversificación de las actividades económicas, más allá de cual haya sido la estrategia que predominó en cada caso. Es probable que en cada hábitat local el manejo del riesgo se desarrollara a partir del uso de distintas tácticas y en todos ellos se puede plantear que la especialización no habría sido una estrategia viable. Esto se desprende del modelo Z score, el cual predice la diversificación como principal estrategia para minimizar el riesgo en adaptaciones humanas (Winterhalder et al. 1999).
Tanto en Matancillas como en la mayoría de los sitios de la Puna Norte y de la Puna Sur, los camélidos domesticados y silvestres fueron los recursos faunísticos indispensables en la dieta. Esto se explicaría por su alto rendimiento económico, medido principalmente por el retorno energético. Al respecto, se puede plantear una alta eficiencia en el consumo de fauna, tal como predicen modelos de la ecología del comportamiento, especialmente el modelo de amplitud de la dieta (López 2002b). Sin embargo, resta considerar en qué medida los costos de ambas estrategias combinadas afectan la eficiencia económica. Para esto es necesario contar con información actualística orientada a determinar principalmente los costos de la estrategia pastoril.
Finalmente, se puede concluir que en Matancillas la diversificación de las estrategias económicas pastoriles y cazadoras, que en conjunto permitirían minimizar el riesgo, no implicarían una pérdida importante de la eficiencia en cuanto al consumo de fauna. Por el contrario, es notable la muy alta representación de recursos faunísticos de alto rendimiento, los camélidos.
Agradecimientos
Agradezco al Lic. Hernán Muscio por la lectura y los comentarios del trabajo. Por el mismo motivo, agradezco al Dr. Guillermo Mengoni Goñalons. También expreso mi agradecimiento a Natalia López por la traducción del resumen al inglés.
REFERENCIAS CITADAS
Aschero, C.; D. Elkin y E. Pintar 1991 Aprovechamiento de recursos faunísticos y producción lítica en el precerámico tardío. Un caso de estudio: Quebrada Seca 3 (Puna Meridional Argentina). Actas del XI Congreso Nacional de Arqueología Chilena vol. 2., pp. 101-114. Santiago de Chile. [ Links ]
Behrensmeyer, A. K. 1978 Taphonomic and ecologic information from bone weathering. Paleobiology 4: 150-62. [ Links ]
Browman, D. 1994 Información y manejo del riesgo de los fleteros de llamas en los Andes Centro-Sur. En Zooarqueología de camélidos 1, editado por D. Elkin, C. Madero, G. Mengoni Goñalons, D. Olivera, M. Reigadas y H. Yacobaccio, pp. 23-42. Grupo de Zooarqueología de Camélidos, Buenos Aires. [ Links ]
Chang, C. y H. Koster 1986 Beyond bones: toward an archaeological of pastoralism. En Advances in Archaeological Methods and Theory 9: 97-147. Academic Press, Nueva York. [ Links ]
Elkin, D. 1996 Arqueozoología de Quebrada Seca 3: Indicadores de Subsistencia Humana Temprana en la Puna Meridional Argentina. Tesis doctoral inédita. Facultad de Filosofía y Letras (UBA), Buenos Aires. [ Links ]
Elkin, D. y J. Zanchetta 1991 Densitometría ósea de camélidos. Aplicaciones arqueológicas. Shinkal 3, Tomo 1: 195-204. [ Links ]
Elkin, D.; C. Madero; G. Mengoni; D. Olivera y H.Yacobaccio.1991 Avances en el estudio arqueológico de los camélidos del noroeste argentino. Trabajo presentado en la VII Convención Internacional de Especialistas en Camélidos Sudamericanos, S. S. de Jujuy. Ms. [ Links ]
Escola, P. 1996 Riesgo e incertidumbre en economías agropastoriles: Consideraciones teórico-metodológicas. Arqueología 6: 9-24. [ Links ]
Fernández Distel, A. 1974 Excavaciones arqueológicas en las cuevas de Huachichocana, Dpto. Tumbaya, Pcia. de Jujuy. Relaciones de la Sociedad Argentina de Antropología VIII: 101-127. [ Links ]
Fernández Distel, A 1986 Las Cuevas de Huachichocana, su posición dentro del precerámico con agricultura incipiente del Noroeste Argentino. Beitrage zur Allgemeinen und Vergleichenden Archaologie, Band 8: 353-430. Bonn. [ Links ]
Fernández Distel, A 1998 Arqueología del Formativo en la Puna Jujeña (1800 AC al 650 DC). Colección Mankacén, Buenos Aires. [ Links ]
Gobel, B.1994 El manejo del riesgo en la economía pastoril de Susques. En Zooarqueología de camélidos vol.1, editado por D. Elkin, C. Madero, G. Mengoni Goñalons, D. Olivera, M. Reigadas y H. Yacobaccio, pp. 43-56. Grupo de Zooarqueología de Camélidos, Buenos Aires. [ Links ]
González, A. R. 1963 Las Tradiciones Alfareras del Período Temprano del N.O. Argentino y sus Relaciones con las de las Áreas Aledañas. Anales de la Universidad del Norte de Antofagasta 2: 49-69. [ Links ]
González, A. R 1977 Arte precolombino en la Argentina, introducción a su historia cultural. Filmediciones Valero, Buenos Aires. [ Links ]
Haber, A. 2001 El oasis en la articulación del espacio circumpuneño. Actas del XIII Congreso Nacional de Arqueología Argentina, pp. 251-274. Córdoba. [ Links ]
Haber, A 2001 La domesticación del oasis. Actas del XIII Congreso Nacional de Arqueología Argentina, pp.451-466. Córdoba. [ Links ]
Kent, J.1982 The domestication and exploitation of the South American camelids: methods of analysis and their application to circum-lacustrine archaeological sites in Bolivia and Peru. Tesis Doctoral inédita. Washington University. St Luois. [ Links ]
Krapovickas, P. 1955 El yacimiento de Tebenquiche (Puna de Atacama). Publicaciones del Instituto de Arqueología III: 1-40 Facultad de Filosofía y Letras (UBA), Buenos Aires. [ Links ]
Lanata, J. L. y L. A. Borrero 1994 Riesgo y Arqueología. En Arqueología de Cazadores-Recolectores. Límites, Casos y Aperturas. Arqueología Contemporánea 5, compilado por J. L. Lanata y L. A. Borrero, pp 129-143. [ Links ]
Layton, R.; R. Foley y E. Williams 1991 The Transition between Hunting and Gathering and the Specialized Husbandry of Resources. A Socio-ecological Approach. Current Anthropology 32(3): 255-274 [ Links ]
López, G.2002 Análisis de conjuntos arqueofaunísticos del Período Formativo de la Puna de Salta: Aportes teóricos para el caso del sitio Matancillas 2. Tesis de Licenciatura inédita. Facultad de Filosofía y Letras, Universidad de Buenos Aires, Buenos Aires. [ Links ]
López, G 2002 La ecología del comportamiento como marco explicativo del consumo de recursos faunísticos del Temprano de la Puna salteña. En Perspectivas integradoras entre arqueología y evolución. Teoría, método y casos de aplicación, editado por G. Martínez y J. L. Lanata, pp. 55-76, INCUAPA (UNCPBA), Olavarría. [ Links ]
López, G. y M. Medina 2001 Optimización y riesgo desde las arqueofaunas: su aplicación en el análisis de adaptaciones con estrategias de producción de alimentos de la Puna de Salta y del Sector Central de Sierras Centrales.Trabajo presentado en el XIV Congreso Nacional de Arqueología Argentina. Rosario, septiembre de 2001. [ Links ]
Meadow, R.1987 Techniques for comparing bone measurement data from small samples. Trabajo presentado a la Northeastern Faunal Analysis Conference, Connecticut. Ms. [ Links ]
Muscio, H.1998-1999 Tendencias en la variabilidad ambiental de la Puna Argentina: Implicancias para la ecología humana prehistórica y para los paisajes arqueológicos. Cuadernos del Instituto Nacional de Antropología y Pensamiento Latinoamericano 18: 271-296. [ Links ]
Muscio, H. 2001 Arqueología de la Quebrada de Matancillas, Puna de Salta. Trabajo presentado en el XIVCongreso Nacional de Arqueología Argentina. Rosario, septiembre de 2001. [ Links ]
Muscio, H. 2003 Etnografía de agricultores y pastores de Matancillas, Puna de Salta: Contribución al análisis arqueológico-evolutivo de las adaptaciones sensitivas del riesgo en las Tierras Altas Surandinas. Ms. [ Links ]
Muscio, H. 2003 Nicho y Estrategia Predominante: Dos Conceptos Útiles en el Estudio Evolutivo de la Diversidad Arqueológica del Noroeste de Argentina.Ms. [ Links ]
Muscio, H. y G. López 2002 Unidades de análisis arqueológicas en el estudio evolutivo de adaptaciones con economías de producción de alimentos: Un examen de las arqueofaunas de la Quebrada de Matancillas (Puna de salta). Ms.Geertz, C. 1966 Religion as a Cultural System. En Anthropological Approaches to the Study of Religion, editado por M. Banton. Asa Monographs 3: 1-46. Tavistock, Londres. [ Links ]
Olivera, D.1991 El Formativo en Antofagasta de la Sierra (Puna Meridional Argentina). Análisis de sus posibles relaciones con contextos arqueológicos agroalfareros Tempranos del Noroeste Argentino y Norte de Chile. Actas del XI Congreso de Arqueología Chilena, II, pp. 61-78. Santiago de Chile. [ Links ]
Olivera, D.1997 La importancia del Recurso Camelidae en la Puna de Atacama entre los 10.000 y 500 años AP. Estudios Atacameños 14: 29-41. [ Links ]
Olivera, D.1998 Cazadores y Pastores Tempranos de la Puna Argentina. En Past and Present in Andean Prehistory and Early History, editado por S. Ahlgren, A. Muñoz, S. Sjodin y P. Stenborg. Etnologiska Studier 42: 153-180. Etnografiska Museet, Gotemburgo. [ Links ]
Olivera, D. y D. Elkin. 1994 De cazadores y pastores: el proceso de domesticación de camélidos en la Puna Meridional Argentina. En Zooarqueología de camélidos 1: editado por D. Elkin, C. Madero, G. Mengoni Goñalons, D. Olivera, M. Reigadas y H. Yacobaccio, pp. 95-124. Grupo de Zooarqueología de Camélidos, Buenos Aires. [ Links ]
Raffino, R.; E. Tonni y A. Cione 1977 Recursos alimentarios y economía en la región de la Quebrada del Toro, Provincia de Salta, Argentina. Relaciones de la Sociedad Argentina de Antropología XI: 9-30. [ Links ]
Rindos, D. 1980 Symbiosis, Instability and the Origins and Spread of Agriculture: a New Model. Current Anthropology 21(6): 751-772. [ Links ]
Ruthsatz, B. y C. Movia 1975 Relevamientos de las estepas andinas del noreste de la Provincia de Jujuy, República Argentina. Fundación para la Educación, la Ciencia y la Cultura, Buenos Aires. [ Links ]
Smith, E. A. y B. Winterhalder 1992 Natural Selection and Decision Making: some Fundamental Principles. En Evolutionary Ecology and Human Behavior, editado por E. A. Smith y B. C. Winterhalder, pp. 25-60. Aldine de Gruyter, Nueva York. [ Links ]
Stephens, D. y J. Krebs 1986 Foraging Theory. Princeton University, Princeton. [ Links ]
Tarragó, M. 1980 Los Asentamientos Aldeanos Tempranos en el Sector Septentrional del Valle Calchaquí, Pcia. de Salta y el Desarrollo Agrícola Posterior. Estudios Arqueológicos 5: 29-53, Antofagasta. [ Links ]
Von den Driesch, A. 1976 A guide to the measurement of animal bones from archaeological sites. Peabody Museum Bulletins 1, Harvard University, Cambridge. [ Links ]
Winterhalder, B. y C. Goland 1997 An Evolutionary Ecology Perspective on Diet Choice, Risk, and Plant Dometication. En People, Plants, and Landscapes Studies in Paleoethnobotany, editado por Kristen J. Gremillion, pp. 123-160. The University of Alabama Press, Alabama. [ Links ]
Winterhalder, B.; F. Lu y B. Tucker 1999 Risk-Sensitive Adaptive Tactics: Models and Evidence from Subsistence Studies in Biology and Anthropology. Journal of Archaeological Research 7 (4): 301-348. [ Links ]
Yacobaccio, H.1994 Biomasa Animal y Consumo en el Pleistoceno-Holoceno Surandino. Arqueología 4: 43-71 [ Links ]
Yacobaccio, H.1997 Sociedad y Ambiente en el NOA Pre-colombino. En De Hombres y Tierras, una Historia Ambiental del Noroeste Argentino, compilado por C. Reboratti, pp. 26-38. Salta. [ Links ]
Yacobaccio, H. y C. Madero 1992 Zooarquelogía de Huachichocana III (Jujuy, Argentina). Arqueología 2: 149-188. [ Links ]
Yacobaccio, H.; D. Elkin y D. Olivera 1994 ¿El fin de las sociedades cazadoras? El proceso de domesticación animal en los Andes Centro Sur. Arqueología contemporánea, vol. 5, compilado por J. L. Lanata y L. A. Borrero, pp. 23-32. [ Links ]
Yacobaccio, H.; C. Madero; M. Malmierca y M.Reigadas 1997-1998 Caza, domesticación y pastoreo de camélidos en la Puna Argentina. Relaciones de la Sociedad Argentina de Antropología XXII- XXIII: 389-428. [ Links ]

