










Servicios Personalizados
Revista

Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkIntersecciones en antropología
versión On-line ISSN 1850-373X
Intersecciones antropol. n.6 Olavarría ene./dic. 2005
Cambios en los patrones de representación ósea del guanaco en Cerro de los Indios 1 (Lago Posadas, Santa Cruz)
Mariana Eleonor De Nigris y María Paz Catá
Mariana E. De Nigris. CONICET, Sección Arqueología, Instituto de Ciencias Antropológicas, Facultad de Filosofía y Letras,
Universidad de Buenos Aires. 25 de Mayo 217, 3° piso. C1002ABE. Ciudad Autónoma de Buenos Aires, Argentina. E-mail: denigris@mail.retina.ar
María Paz Catá. CONICET, Sección Arqueología, Instituto de Ciencias Antropológicas, Facultad de Filosofía y Letras, Universidad de Buenos Aires. E-mail: paz@tasar-sa.com.ar
Recibido 30 de Junio 2004. Aceptado 24 de Noviembre 2004
RESUMEN
El análisis de los materiales óseos provenientes del bloque temporal tardío (ca. 1.800-900 años AP) de la localidad de Cerro de los Indios 1, emplazada en el noroeste de Santa Cruz, permite discutir una serie de tendencias en el aprovechamiento de los recursos faunísticos explotados por sociedades cazadoras-recolectoras que habitaron el interior de Patagonia. En este trabajo se analizan nuevas muestras con el objetivo de profundizar las conclusiones planteadas en estudios previos. Los resultados obtenidos señalan al guanaco (Lama guanicoe) como la especie dominante de toda la secuencia. Sus patrones de representación de partes varían a lo largo del tiempo, destacándose la mayor abundancia de extremidades inferiores en especial las falanges y la relativa ausencia de elementos del esqueleto axial hacia los momentos más tardíos.
Palabras claves: Zooarqueología; Cazadores-recolectores; Patagonia; Perfiles esqueletarios; Guanaco.
ABSTRACT
This study investigates hunter-gatherer faunal exploitation in inland Patagonia. The sample analyzed consists of a series of bone assemblages from the archaeological site Cerro de los Indios 1 (CI1), located in the northwest of Santa Cruz Province, which date from between 1800 and 900 years BP. The site is a large rock shelter located in the Lake Posadas-Pueyrredón Basin (270 m above-sea-level), a steppe environment. The results show the guanaco (Lama guanicoe) as the dominant species over the entire sequence. Close examination of skeletal part frequencies of this ungulate suggests changes through time: appendicular skeletal elements are more abundant than axial skeletal elements, particularly in more recent periods. The high frequency of lower limb and feet bones is also noteworthy. Moreover, it is important to note that high meat utility elements decrease in proportion towards the end of the CI1 occupations.
Keywords: Zooarchaeology; Hunter-gatherers; Patagonia; Skeletal profiles; Guanaco.
INTRODUCCIÓN
La localidad de Cerro de los Indios 1 (CI1), situada en el NO de Santa Cruz, ha sido intensamente estudiada en los últimos años (Aschero et al. 1999; De Nigris et al. 2004; Figuerero Torres 2000a, 2000b, 2004; Guráieb 1998, 1999, 2000a, 2000b; Mengoni Goñalons y Yacobaccio 2000). Entre las diversas líneas de investigación desarrolladas, la perspectiva zooarqueológica ha tenido particular relevancia, permitiendo discriminar ciertas tendencias generales (Catá 2003; De Nigris 1994, 2000, 2001; De Nigris y Mengoni Goñalons 2000; Mengoni Goñalons y De Nigris 1999; Mengoni Goñalons 1999). Hasta el momento, estos estudios señalan al guanaco (Lama guanicoe) como el recurso principalmente aprovechado por las sociedades cazadoras-recolectoras que ocuparon el alero entre ca. 3.800-900 años AP. En contraposición, sólo unos pocos especímenes de otras especies animales pudieron ser identificados.
A lo largo de la secuencia estratigráfica se observa que todas las partes esqueletarias del guanaco están presentes, no obstante predomina el esqueleto apendicular. En particular, para los momentos más tardíos de la ocupación es evidente el predominio, aún más destacado, de los huesos de las patas, mientras que los huesos del esqueleto axial si bien están presentes registran muy bajas proporciones. También es notable la incidencia de marcas de procesamiento que presentan los huesos -tanto apendiculares como axiales- atribuidos a esta especie.
Las diferencias observadas en los conjuntos óseos a través del tiempo, en especial los cambios registrados en los patrones de representación ósea en los últimos momentos de ocupación del alero, han motivado la incorporación de nuevas muestras para comprender, más acabadamente, las prácticas de aprovechamiento de presas que se estaban desarrollando en la localidad.
EL ALERO CERRO DE LOS INDIOS 1 Y SUS UNIDADES DE ANÁLISIS
La localidad arqueológica CI1 se encuentra localizada en la cuenca de los lagos Posadas-Pueyrredón, en el noroeste de la provincia de Santa Cruz (47° 35' LS, 71° 43' LO; 270 m snm). Se trata de un gran alero emplazado al pie de un farallón rocoso en el frente norte del cerro que lleva el mismo nombre, distante a unos 3,5 km del pueblo de Hipólito Irigoyen (ex Lago Posadas, IGM Hoja 4772-28; Figura 1) (Aschero 1996; Aschero et al. 1999; De Nigris et al. 2004; Gradin et al. 1979; Mengoni Goñalons y Yacobaccio 2000).

Figura 1. Mapa de localización de Cerro de los Indios 1.
Entre sus particularidades se puede mencionar que presenta una gran extensión de superficie ocupable (aproximadamente 240 m2); una alta densidad de materiales superficiales y en capa -tanto líticos como óseos-, observándose, asimismo, una alta redundancia de la ocupación acompañada por una marcada estructuración del espacio habitado; una importante concentración y diversidad de motivos de arte rupestre, característicos de los estilos más tardíos del Río Pinturas. Del mismo modo, es importante subrayar también su emplazamiento destacado dentro del paisaje circundante (Aschero 1996; Aschero et al. 1999; De Nigris et al. 2004; Gradin et al. 1979; Mengoni Goñalons y Yacobaccio 2000).
Hasta la fecha tres sectores han sido excavados en la localidad, denominados Área de Excavación 1, 2 y 3 (AE1, AE2 y AE3), respectivamente (Aschero et al. 1999; De Nigris et al. 2004). Los materiales óseos provenientes de la primer área (AE1) han sido intensamente estudiados (De Nigris 1994; Mengoni Goñalons 1999; Mengoni Goñalons y De Nigris 1999), mientras que los procedentes del AE2 si bien han recibido particular atención sólo se han presentado resultados parciales (Catá 2003; De Nigris 2000, 2001; De Nigris y Mengoni Goñalons 2000). Con la intención de seguir completando el panorama zooarqueológico de la localidad, este trabajo se concentra en los restos recuperados en esta última área.
En este sector han sido identificadas 17 unidades lito-estratigráficas, claramente definidas por su color, textura y estructura; su secuencia cronológica, hasta el momento, se extiende entre ca. 3.800-900 años AP. (Aschero et al. 1999; De Nigris et al. 2004). Este análisis abarca los materiales procedentes de las Capas 1 a 12; las tres primeras unidades (1, 2 y 3) no poseen fechados radiocarbónicos, mientras que las restantes unidades se ubican entre ca. 1.800-900 años AP en lo que fuera oportunamente denominado bloque temporal tardío (para una discusión detallada de las unidades estratigráficas y sus fechados ver De Nigris et al. 2004, Figuerero Torres 2004: Figura 2). Durante el estudio de los huesos recuperados se emplea el criterio de máxima distinción, respetando las divisiones naturales observadas entre las distintas unidades (Grayson 1984).

Figura 2. Representación anatómica axial en CI1.
RESULTADOS
Del análisis de los nuevos conjuntos óseos se desprende queel guanaco (Lama guanicoe) sigue siendo la especie dominante a lo largo de la secuencia, confirmando la tendencia ya observada (De Nigris y Mengoni Goñalons 2000). Si bien otras especies de mamíferos han sido identificadas -piche (Zaedyus pichiy), zorro (Pseudalopex sp.) y félidos (Felis sp.)- y algunas variedades de aves, entre las que se destaca el ñandú (Pterocnemia pennata), éstas nunca superan el 2% de la muestra considerada (Tabla 1). La presencia de marcas de procesamiento (e.g., corte, percusión) y la casi total ausencia de marcas de carnívoros en estos especímenes, señalaría su origen cultural.
Tabla 1. Taxones identificados en CI1 (valores de NISP).
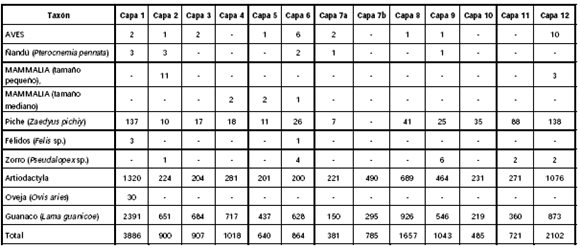
Cabe destacar, sin embargo, que en el caso de la Capa 1 fueron reconocidos también algunos huesos asignados a oveja (Ovis aries) -cuyo Número de Especímenes Identificados por Taxón (NISP) es igual a 30- (Tabla 1). En este caso su incorporación al conjunto zooarqueológico recuperado, sería el producto de muertes naturales ocurridas en el alero por lo menos desde la segunda década del siglo XX, fecha en la cual se introdujo esta especie en el área -cuenca de los lagos Posadas-Pueyrredón- (Catá 2003). Además, un importante número de huesos carecía de atributos diagnósticos que permitieran discriminarlos a nivel de especie, pero sus características generales posibilitaron su asignación a la categoría más amplia de Artiodactyla. Su representación en la muestra analizada resulta elevada y el conjunto está compuesto por fragmentos de dientes sueltos, porciones de cráneo, vértebras indeterminadas, superficies articulares no diferenciadas y diáfisis no diagnósticas de huesos largos. Asimismo, se han identificado una serie de especímenes pertenecientes a mamíferos indeterminados. Sin embargo, la consideración de sus rasgos morfológicos permitió realizar una distinción por tamaño: Mammalia de tamaño pequeño o menores a 5 kg y Mammalia de tamaño mediano entre 5 y 50 kg. En ambos casos se trata fundamentalmente de fragmentos de cráneo, trozos de dientes y diáfisis no diagnósticas de huesos largos.
Dada la preponderancia del guanaco por sobre el resto de los taxones identificados en toda la secuencia estudiada, este trabajo se concentra de aquí en adelante en su análisis detallado.
PATRONES DE REPRESENTACIÓN ANATÓMICA DE GUANACO
En relación con los valores de NISP calculados para cada una de las unidades se puede mencionar que, en general, oscilan entre los 200 y 900 especímenes, con la excepción de la Capa 1 que presenta un valor significativamente más elevado (NISP = 2391), señalando una mayor densidad de materiales óseos (Tabla 1). Si se estima la abundancia taxonómica relativa del Número Mínimo de Individuos (MNI)1 de la unidad 1 continúa siendo notable la diferencia en proporción al resto de las unidades (Tabla 2), dado que el valor calculado para este primer conjunto supera ampliamente los obtenidos para las demás capas.
Tabla 2. Representación del guanaco en CI1.

Cuando se evalúa la relación entre el NISP y el MNI (Tabla 2) se observa que en las Capas 1 y 2 hay menos especímenes óseos por individuo, es decir, que los guanacos de estos conjuntos están menos completos, situación que se repite en la unidad 10.2 Sin embargo, dado que esta última es la que posee el menor valor de NISP de toda la secuencia analizada, es posible que el resultado de la relación entre ambas medidas se vea afectado por el tamaño de la muestra (Grayson 1984; Lyman 1994; Ringrose 1993).
Los resultados de la razón entre el Número Mínimo de Elementos (MNE) y el MNI confirman la tendencia anteriormente descripta, sumándose en este caso la capa 11 (Tabla 2). En ésta última, la diferencia entre los resultados que arrojan ambas relaciones -NISP vs. MNI y MNE vs. MNI- podría explicarse por un mayor grado de fragmentación de los restos óseos de este conjunto, tal como se manifiesta en la relación entre el NISP y el MNE (Tabla 2).
Otra de las tendencias anteriormente descriptas marcaba el predominio del esqueleto apendicular sobre el axial, especialmente en las capas más recientes (De Nigris y Mengoni Goñalons 2000). La incorporación de los nuevos conjuntos, particularmente la Capa 1, refuerza esta observación e incluso la profundiza ya que en esta primera unidad se acentúa la primacía de la región apendicular, siendo la relación de 3 a 1. No obstante, las capas intermedias e inferiores de la secuencia analizada en esta oportunidad -7a a 12- exhiben un incremento en la representación de los elementos axiales, de forma tal que la relación entre ambas zonas del esqueleto tiende a equilibrarse cuando se considera los valores de NISP de cada región (Tabla 3).
Tabla 3. Regiones esqueletarias principales de guanaco en CI1.

Cuando se examinan los MNE (Tabla 3) se observa que la tendencia se mantiene, indicando que la segmentación de las unidades anatómicas no incidiría en la representación desigual de ambas zonas. La única excepción se evidencia en la Capa 11, en la cual la proporción del MNE axial resulta mucho menor en comparación con la del NISP, posiblemente debido a la fragmentación diferencial de este sector del esqueleto.
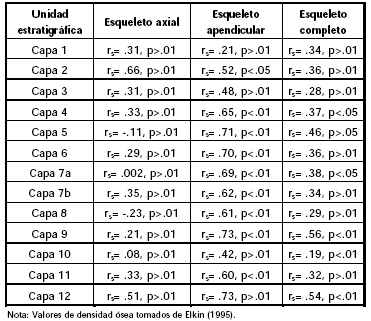
Es relevante destacar que los patrones de representación anatómica no se verían afectados por procesos dependientes de la densidad mineral ósea (Grayson 1989; Lyman 1994), tal como señalan las correlaciones efectuadas entre el MAU% y los valores de densidad ósea calculados para camélidos por Elkin (1995) (Tabla 4). Esto es especialmente evidente en el caso del esqueleto completo y de la región axial. Por el contrario, la covariación entre estas variables en la porción apendicular resulta, en algunas capas, elevada y estadísticamente significativa. Sin embargo, en las mismas unidades se registra la presencia de huesos con bajas densidades óseas, por lo tanto es posible que los resultados obtenidos se deban a un proceso de destrucción diferencial relacionado con el procesamiento y consumo de los huesos largos, corroborado a su vez por la presencia de fracturas de carácter intencional (e.g., negativos de impacto, fracturas transversales marcadas, huellas de percusión) y una muy baja incidencia -menor al 1%- de huellas de carnívoro.
Tabla 4. Coeficiente de correlación rho de Spearman para guanaco en CI1 (MAU% y densidad ósea).
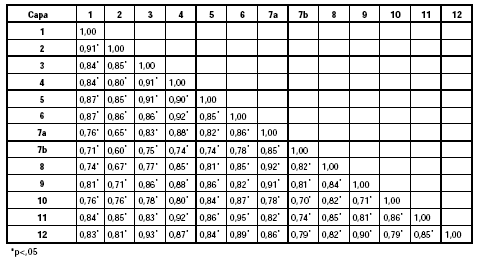
Se compararon entre sí los MNE de las distintas capas con el propósito de evaluar su grado de similitud. Los resultados obtenidos se presentan en la Tabla 5. De ella se desprende la semejanza existente entre las diversas unidades estratigráficas dado que los resultados alcanzan valores elevados y estadísticamente significativos. A pesar de las diferencias en el NISP -anteriormente comentadas- que presentan los conjuntos analizados, es destacable que la representación anatómica se mantiene constante a lo largo del tiempo, aunque como se verá más adelante, con ciertas variaciones.
Tabla 5. Matriz de correlación (rho de Spearman) basada en el MNE.
Afinando un poco más la escala, se pueden dividir las distintas porciones de los animales consumidos conforme al producto primario que predomina en cada una de ellas. De este modo, las partes anatómicas que componen una carcasa pueden segmentarse en las siguientes categorías: (a) huesos con carne solamente - vértebras, costillas, pelvis y escápula-; (b) huesos con abundante carne y médula -húmero y fémur; (c) huesos con poca carne y médula -radioulna y tibia-; (d) huesos con moderadas proporciones de médula solamente -metapodios-; y (e) huesos con bajas proporciones de médula -calcáneo y falanges-. Una categoría adicional correspondería al cráneo, el cual contiene órganos ricos en grasas (Stiner 1994).
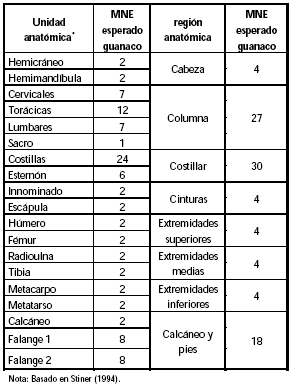
Siguiendo una propuesta originalmente planteada por Stiner (1994) se ha fraccionado el esqueleto del guanaco en ocho regiones esqueletarias diferentes. Para ello se centró, como se señala precedentemente, en los recursos que primordialmente proporcionan las diferentes partes esqueletarias de la carcasa. De esta forma, se midieron las diferentes unidades de consumo ya que se entiende que los elementos que brindan recursos similares deben tener secuencias de utilización afines. Los valores de MNE esperados para guanaco se presentan en la Tabla 6. Ciertos huesos fueron excluidos (vértebras caudales, carpianos y tarsianos -a excepción del calcáneo-, patella, sesamoideos y falange 3) puesto que no proporcionan ni carne ni médula. Además, en esta oportunidad, se ha subdivido la categoría de huesos ricos en carne en otras tres regiones -columna, costillares y cinturas- con el objetivo de evitar enmascarar la variabilidad que podría existir en el aprovechamiento de estas múltiples unidades.
Tabla 6. Valores de MNE esperados por unidad anatómica y por región esqueletaria para guanaco.
La relación entre el MNE observado y el esperado para guanaco permitió estandarizar las frecuencias de las diversas regiones anatómicas (Figuras 2 y 3). La Figura 2 ilustra la representación anatómica axial. La cabeza se encuentra presente en todas las unidades estratigráficas, predominando en alguna de ellas -e.g., Capas 9 y 12-. La columna vertebral si bien está presente, no alcanza proporciones demasiado elevadas, a excepción de la Capa 1. Por su parte, las frecuencias de los costillares son muy bajas en todas las capas; mientras que las proporciones de las cinturas son en general altas, destacándose en algunas de las unidades -e.g., Capas 2 y 6-.
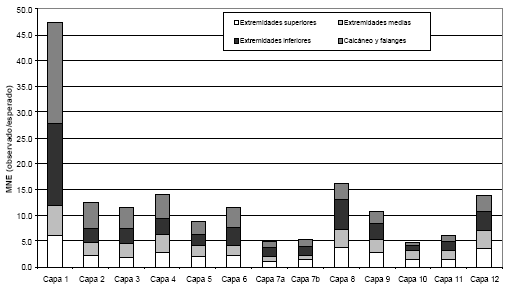
Figura 3. Representación anatómica apendicular en CI1.
En cuanto al esqueleto apendicular (Figura 3) se puede señalar que las extremidades superiores están pobremente representadas en la mayoría de las capas analizadas y de manera similar se comportan las extremidades medias. Los metapodios tienden a estar mejor representados que el resto de los huesos largos. Las falanges y los calcáneos, en las capas superiores -1 a 4- , presentan frecuencias más elevadas que las del resto del conjunto apendicular, en tanto que en las demás unidades si bien alcanzan valores importantes, no superan los calculados para las extremidades inferiores.
Se evalúa, entonces, en qué medida los patrones de representación de partes esqueletarias generales registrados podían responder al transporte selectivo en función del atractivo nutricional diferencial que poseen las diversas unidades anatómicas que conforman una carcasa. Se correlacionó el MAU% y el índice de utilidad de carne obtenido por Borrero (1990). En la correlación se consideraron 17 pares de observaciones siguiendo las recomendaciones sugeridas por Mengoni Goñalons (1999).
Los resultados de la correlación fueron negativos en todas las unidades estratigráficas, siendo significativos en algunos casos (Tabla 7). Ellos señalan la ausencia de selectividad de partes en función de la carne que brindan; indicando, por el contrario, que aquellas unidades anatómicas que poseen menores cantidades de este recurso son las más representadas.
Tabla 7. Coeficiente de correlación rho de Spearman para guanaco en CI1 (MAU% y utilidad de carne).

Asimismo, se consideró si las frecuencias de huesos largos registradas en las diversas unidades estratigráficas de CI1 respondían a una selección de partes en función de la carne que proporcionan. Se observó, en general, una correlación alta y negativa entre el MAU% y el índice de utilidad de carne (Borrero 1990), indicando que los huesos largos presentes son aquellos que poseen pocas cantidades de músculo asociado.
Del mismo modo, no se advierte covariación entre la representación de los huesos largos junto con la falange 1 y el volumen de la cavidad medular (Mengoni Goñalons 1996), mostrando que los huesos que poseen mayores proporciones de tuétano no son los que evidencian valores de MAU más elevados (Tabla 8). Por el contrario, parecería existir una selección inversa, es decir, aquellos huesos largos que poseen cavidades medulares más pequeñas se encuentran mejor representados (Figura 3).
Tabla 8: Coeficiente de correlación rho de Spearman para guanaco en CI1 (MAU% y volumen de la cavidad medular de huesos largos y falange 1).

En cuanto al esqueleto axial, la correlación entre el índice de utilidad de carne y el MAU% fue negativa en todos los casos y, en general, los valores son de bajos a moderados, con la excepción de la capa 7b que presenta un valor alto y estadísticamente significativo (Tabla 7). Estos resultados señalan, para esta región del esqueleto también, la ausencia de selectividad de aquellas partes que poseen más carne.
Los resultados de las correlaciones efectuadas indicarían un transporte selectivo de aquellas partes que poseen menos recursos (carne y médula). En términos tradicionales éstas se corresponderían con localidades de matanza y/o procesamiento inicial (e.g., Binford 1978). Como se ha visto, las características generales de la localidad de CI1 - fundamentalmente una alta densidad de materiales superficiales y en capa, una alta redundancia de la ocupación acompañada por una marcada estructuración del espacio habitado- difieren absolutamente de lo que podría considerarse como una localidad de ese tipo. Por otra parte, la densidad ósea tampoco sería la responsable de las curvas negativas observadas; cabría preguntarse, entonces, qué otros comportamientos podrían explicar los patrones de representación de partes observados.
A fin de responder este interrogante se evaluó en qué medida el secado de la carne con hueso podría ser responsable de los patrones identificados. Para ello se correlacionó el MAU% con el índice de secado de carne calculado por De Nigris y Mengoni Goñalons (2004). Los resultados de la covariación son negativos, de moderados a altos y estadísticamente significativos (Tabla 9), indicando que esta estrategia de conservación del recurso cárnico fue probablemente implementada por los grupos cazadores-recolectores que habitaron el alero entre los 900 y 1800 años AP. Es importante tener en cuenta que las diferentes formas de preparación de los productos animales se pueden ligar con un consumo inmediato o diferido de los mismos, las cuales se relacionan a su vez con la posibilidad de implementar técnicas de preservación adecuadas. Existen modos de conservación de los alimentos que se vinculan a la eliminación de la humedad presente en las diversas partes anatómicas, pudiéndose efectuar el secado de la carne con el hueso mediante la exposición al sol o a alguna otra fuente de calor (Binford 1978; De Nigris 2004; De Nigris y Mengoni Goñalons 2004; Friesen 2001).
Tabla 9. Coeficiente de correlación rho de Spearman para guanaco en CI1 (MAU porcentual e índice de secado de carne).

DISCUSIÓN Y CONSIDERACIONES FINALES
La incorporación de los nuevos conjuntos reforzó las tendencias que habían sido observadas con anterioridad. Continúa siendo destacado el predominio del guanaco por sobre las demás especies identificadas. Esta dominancia no resulta llamativa dado que en la zona geográfica donde se ubica la localidad de CI1 no existe una gran diversidad de especies animales y tampoco parece haberla habido en el pasado. De todas formas, para el período de ocupación humana del alero, podría esperarse que otras presas se incorporasen al registro, como por ejemplo el ñandú (Pterocnemia pennata) y el huemul (Hippocamelus bisulcus) presentes en otros sitios arqueológicos de Patagonia (ver por ejemplo Aschero et al. 1992; Cassiodoro et al. 2000; De Nigris 2004; Díaz 2000; Mena y Jackson 1991; Mena et al. 2000). Esta última especie aparece representada en otras localidades generalmente emplazadas en áreas boscosas y ecotonales. En el caso del ñandú, si bien se han recuperado algunos especímenes óseos su representación en la muestra analizada es prácticamente nula (Tabla 1), mientras que en el segundo caso, no se ha registrado ningún resto que pudiera ser asignado a este cérvido.
En cuanto a la explotación del guanaco ciertos aspectos pueden ser destacados. Primeramente, interesa resaltar que en el bloque temporal analizado se distingue la Capa 1 por la gran abundancia de especímenes que presenta, como así también por su elevado MNI. Es importante aclarar que esta unidad estratigráfica se comportaría de manera particular. A diferencia de las otras, presenta un bajo grado de preservación del registro y una baja conservación de rasgos que contribuyen a la estructuración del espacio ocupado (Figuerero Torres 2004); se ha planteado, además, que en su formación debieron haber intervenido un mayor número de eventos de acumulación ocurridos quizá de forma más rápida y seguida en el tiempo (Catá 2003).
Las capas superiores -1 y 2- de la secuencia estudiada presentan, como se ha observado, un menor número de huesos por individuo en comparación con las otras unidades. Este hecho resulta particularmente llamativo en el primer conjunto dada la densidad de restos óseos registrada. Cabe preguntarse, entonces, si este patrón respondería a una estrategia de selección y explotación de partes específica de los momentos más tardíos de la ocupación.
Los estudios efectuados señalan un marcado predominio del esqueleto apendicular por sobre el axial, que se acentúa en las capas más recientes. Cabe aclarar que esta preponderancia no respondería a procesos mediados por la densidad ni tampoco se vería afectada por la fragmentación diferencial. Por su parte, la selección de los elementos apendiculares no parece estar explicada por las cantidades de carne o médula que proporcionan. Contrariamente, las unidades anatómicas que poseen menores cantidades de estos recursos son las que aparecen con mayor frecuencia. Lo mismo puede plantearse para la región axial del esqueleto.
De los estudios realizados surge que esta ausencia de partes ricas en carne podría deberse a un consumo diferido en el tiempo de aquellas porciones que tienen mejores cualidades para ser secadas con el hueso -básicamente el esqueleto axial-. Al respecto, es destacable la escasez de costillas y esternebras observada en todos los conjuntos, precisamente las porciones más adecuadas para la deshidratación.
Con respecto a la región apendicular del esqueleto es notable la baja representación que tienen, en general, las extremidades superiores y medias, ricas en carne y médula. Mientras que los metapodios, que solamente proporcionan tuétano, tienden a presentarse en frecuencias más elevadas, especialmente en las capas más recientes -1 a 6-.
Las falanges 1 y 2 constituyen un caso peculiar, son muy abundantes en todas las capas pero es significativo el aumento de sus frecuencias desde la Capa 6 hasta la 1, llegando incluso a ser dominantes en los conjuntos superiores. Se encuentran, en la mayoría de los casos, segmentadas intencionalmente, registrando claras evidencias de la actividad humana (e.g., negativos de impactos y huellas de percusión). Sin embargo, es subrayable que las cavidades medulares de estos huesos resultan sumamente pequeñas en comparación con otros elementos óseos. Cabe preguntarse, entonces, cuáles serían las razones que llevarían a los cazadores a explotar huesos con tan bajos niveles de grasas.
Este notable incremento en el aprovechamiento de las falanges, así como también la disminución de las frecuencias de las extremidades superiores y medias, hacia los momentos más tardíos y, en particular en la Capa 1, señala un evidente cambio en la explotación del guanaco y sus recursos asociados. La consideración de las marcas de procesamiento y consumo y de las fracturas óseas, junto con la evaluación de otras líneas de evidencia permitirá comprender mejor la naturaleza de esta transformación.
Agradecimientos
La realización de este trabajo ha sido posible gracias a las Becas Posdoctoral y Doctoral del CONICET que tenemos en curso. Nuestro especial agradecimiento al Dr. Guillermo Mengoni Goñalons por habernos guiado en nuestro camino zooarqueológico. Todo lo vertido aquí es de nuestra exclusiva responsabilidad.
1 El MNI se obtuvo considerando el valor de la unidad anatómica más numerosa en la muestra, teniendo en cuenta el lado -izquierdo o derecho- y el estadio de fusión -no fusionado o fusionado- de los especímenes analizados (ver Grayson 1984; Lyman 1994; Mengoni Goñalons 1999; Ringrose 1993).
2 Para el cálculo de la relación entre el NISP y el MNI se han excluido, siguiendo a Mengoni Goñalons (1999), aquellos elementos óseos que no tienen valor nutricional asociado -vértebras caudales, rótula, carpianos, tarsianos y sesamoideos- y que no serían objeto de transporte en sí mismos sino por estar asociados a unidades anatómicas ricas en algún recurso (carne o médula). Si bien es cierto que algunos de estos elementos poseen mayores posibilidades de supervivencia en determinados contextos arqueológicos, debido a su mayor densidad ósea relativa, la representación anatómica observada en los conjuntos analizados en este trabajo no se encuentra afectada por procesos mediados por la densidad mineral. Asimismo, los elementos excluidos no presentan frecuencias de MAU% elevadas.
REFERENCIAS CITADAS
Aschero, C. A. 1996 El área Río Belgrano-Lago Posadas (Santa Cruz): problemas y estado de problemas. En Arqueología: Sólo Patagonia, editado por J. Gómez Otero, pp.17-26. CENPAT, Puerto Madryn. [ Links ]
Aschero, C. A., C. Bellelli y R. A. Goñi 1992 Avances en las investigaciones arqueológicas del Parque Nacional Perito Moreno, Provincia de Santa Cruz, Patagonia Argentina. Cuadernos del Instituto Nacional de Antropología y Pensamiento Latinoamericano 14: 143-170. [ Links ]
Aschero, C. A., M. E. De Nigris, M. J. Figuerero Torres, A. G. Guráieb, G. L. Mengoni Goñalons y H. D. Yacobaccio 1999 Excavaciones recientes en Cerro de los Indios 1, Lago Posadas (Santa Cruz): nuevas perspectivas. En Soplando en el viento, pp. 269-286. Instituto Nacional de Antropología y Pensamiento Latinoamericano y Universidad del Comahue, Neuquén-Buenos Aires. [ Links ]
Borrero, L. A. 1990 Fuego-patagonian bone assemblages and the problem of communal guanaco hunting. En Hunters of the Recent Past, editado por L. B. Davis y B. O. K. Reeves, pp. 373-399. Unwin Hyman, Londres. [ Links ]
Binford, L. R. 1978 Nunamiut Ethnoarchaeology. Academic Press, Nueva York. [ Links ]
Cassiodoro, G., G. Lublin, M. F. Piriz, y D. Rindel 2000 Los primeros pasos en el Alero Destacamento Guardaparque: análisis lítico y faunístico (NO Provincia de Santa Cruz, Argentina). En Desde el País de los Gigantes: perspectivas arqueológicas en Patagonia, pp. 369-384. Universidad Nacional de la Patagonia Austral, Río Gallegos. [ Links ]
Catá, M. P. 2003 Aspectos metodológicos relacionados con el concepto de grano. Un caso de aplicación al registro arqueofaunístico: Cerro de los Indios 1. Tesis de Licenciatura inédita. Departamento de Antropología, Facultad de Filosofía y Letras, Universidad de Buenos Aires. Buenos Aires. [ Links ]
De Nigris, M. E. 1994 Patrones de fragmentación de huesos largos en el sitio Cerro de los Indios 1, Lago Posadas. Tesis de Licenciatura inédita. Departamento de Antropología, Facultad de Filosofía y Letras, Universidad de Buenos Aires. Buenos Aires. [ Links ]
De Nigris, M. E. 2000 Procesando para el consumo: dos casos de Patagonia Meridional. En Desde el país de los gigantes: perspectivas arqueológicas en Patagonia, pp. 401-414. Universidad Nacional de la Patagonia Austral, Río Gallegos. [ Links ]
De Nigris, M. E. 2001 Patrones de procesamiento final y consumo del guanaco en Patagonia. En Zooarqueología de Camélidos 3: El Uso de los Camélidos a través del Tiempo, editado por G. L. Mengoni Goñalons, D. E. Olivera y H. D. Yacobaccio, pp. 9-28. Ediciones del Tridente. Buenos Aires. [ Links ]
De Nigris, M. E. 2004 El consumo en grupos cazadores-recolectores. Un ejemplo zooarqueológico de Patagonia meridional. Sociedad Argentina de Antropología, Buenos Aires. [ Links ]
De Nigris, M. E. y G. L. Mengoni Goñalons 2000 Patrones y tendencias generales de los conjuntos faunísticos en Cerro de los Indios 1. Arqueología 10: 227-243. [ Links ]
De Nigris, M. E. y G. L. Mengoni Goñalons 2004 El guanaco como fuente de carne y grasas en Patagonia. En Contra viento y marea. Arqueología de Patagonia, editado por T. Civalero, P. Fernández y A. G. Guráieb, pp. 469-476. Instituto Nacional de Antropología y Pensamiento Latinoamericano, Buenos Aires. [ Links ]
De Nigris, M. E., M. J. Figuerero Torres, A. G. Guráieb y G. L. Mengoni Goñalons 2004 Nuevos fechados radiocarbónicos en la localidad de Cerro de los Indios 1 (Santa Cruz) y su proyección areal. En Contra viento y marea. Arqueología de Patagonia, editado por T. Civalero, P. Fernández y A. G. Guráieb, pp. 537-544. Instituto Nacional de Antropología y Pensamiento Latinoamericano, Buenos Aires. [ Links ]
Díaz, N. I. 2000 El huemul (Hippocamelus bisulcus Molina, 1782): Una perspectiva histórica. En El huemul Patagónico: un misterioso cérvido al borde de la extinción, editado por N. I. Díaz y J. Smith-Flueck, pp. 1-32. L.O.L.A, Buenos Aires. [ Links ]
Elkin, D. C. 1995 Volume Density of South American Camelid Skeletal Parts. International Journal of Osteoarchaeology 5: 29-37. [ Links ]
Figuerero Torres, M. J. 2000a Análisis de la estructura del espacio en Cerro de los Indios 1 (Lago Posadas, Santa Cruz). En Desde el país de los gigantes. Perspectivas arqueológicas en Patagonia, pp. 385-400. Universidad Nacional de la Patagonia Austral, Río Gallegos. [ Links ]
Figuerero Torres, M. J. 2000b Tendencias en el uso del espacio en Cerro de los Indios 1. Arqueología 10: 203-214. [ Links ]
Figuerero Torres, M. J. 2004 La estructuración del espacio a través del tiempo en Cerro de los Indios 1 (Lago Posadas, Santa Cruz). En Contra viento y marea. Arqueología de Patagonia, editado por T. Civalero, P. Fernández y A. G. Guráieb, pp. 557-563. Instituto Nacional de Antropología y Pensamiento Latinoamericano, Buenos Aires. [ Links ]
Friesen, T. M. 2001 A Zooarchaeological Signature for Meat Storage: Re-Thinking the Drying Utility Index. American Antiquity 66: 315-331. [ Links ]
Gradin, C. J., C. A. Aschero y A. M. Aguerre 1979 Arqueología del Área Río Pinturas (Provincia de Santa Cruz). Relaciones de la Sociedad Argentina de Antropología XIII: 183-227. [ Links ]
Grayson, D. K. 1984 Quantitative Zooarchaeology. Academic Press, Orlando. [ Links ]
Grayson, D. K. 1989 Bone Transport, Bone Destruction and Reverse Utility Curves. Journal of Archaeological Science 16: 643-662. [ Links ]
Guráieb, A. G. 1998 Cuáles, cómo y dónde: tendencias temporales de selección de recursos líticos en el sitio Cerro de los Indios 1 (Lago Posadas, Santa Cruz). Arqueología 8: 77-99. [ Links ]
Guráieb, A. G. 1999 Análisis de la diversidad en los conjuntos instrumentales líticos de Cerro de los Indios 1 (Lago Posadas, Santa Cruz). Relaciones de la Sociedad Argentina de Antropología XXIV: 293-306. [ Links ]
Guráieb, A. G. 2000a Diversidad artefactual y selección de materias primas en contextos tardíos de Cerro de los Indios 1 (Lago Posadas, Santa Cruz). En Desde el país de los gigantes. Perspectivas arqueológicas en Patagonia, pp. 19-28. Universidad Nacional de la Patagonia Austral, Río Gallegos. [ Links ]
Guráieb, A. G. 2000b Características tecnológicas y composición de los conjuntos líticos en Cerro de los Indios 1. Arqueología 10: 215-225. [ Links ]
Lyman, R. L. 1994 Vertebrate Taphonomy. Cambridge University Press, Nueva York. [ Links ]
Mena, F. y D. Jackson 1991 Tecnología y subsistencia en Alero Entrada Baker, Región de Aisén, Chile. Anales del Instituto de la Patagonia(Serie Ciencias Sociales) 20: 169-203. [ Links ]
Mena, F., V. Lucero, O. Reyes, V. Trejo y H. Velásquez 2000 Cazadores tempranos y tardíos en la Cueva Baño Nuevo-1, margen occidental de la estepa centropatagónica (XI Región de Aisén, Chile). Anales del Instituto de la Patagonia (Serie Ciencias Históricas) 28: 173-195. [ Links ]
Mengoni Goñalons, G. L. 1996 La domesticación de los camélidos sudamericanos y su anatomía económica. En Zooarqueología de Camélidos 2, editado por D. C. Elkin, C. M. Madero, G. L. Mengoni Goñalons, D. E. Olivera, M. C. Reigadas y H. D. Yacobaccio, pp. 33-45. Grupo Zooarqueología de Camélidos, Buenos Aires. [ Links ]
Mengoni Goñalons, G. L. 1999 Cazadores de guanacos de la estepa patagónica. Sociedad Argentina de Antropología, Buenos Aires. [ Links ]
Mengoni Goñalons, G. L. y M. E. De Nigris 1999 Procesamiento de huesos largos de guanaco en Cerro de los Indios 1 (Santa Cruz). En Soplando en el viento, pp. 461-475. Instituto Nacional de Antropología y Pensamiento Latinoamericano y Universidad del Comahue, Neuquén-Buenos Aires. [ Links ]
Mengoni Goñalons, G. L. y H. D. Yacobaccio 2000 Arqueología de Cerro de los Indios y su entorno. Arqueología 10: 193-201. [ Links ]
Ringrose, T. J. 1993 Bone Counts and Statistics: a Critique. Journal of Archaeological Science 20: 121-157. [ Links ]
Stiner, M. C. 1994 Honor among Thieves: a Zooarchaeological Study of Neanderthal Ecology. Princeton University Press, Nueva Jersey. [ Links ]

