










Services on Demand
Journal

Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO
Related links
-
 Similars in
SciELO
Similars in
SciELO
Share

 Permalink
PermalinkIntersecciones en antropología
On-line version ISSN 1850-373X
Intersecciones antropol. no.6 Olavarría Jan./Dec. 2005
El consumo de moluscos en la Costa Norte de Santa Cruz
Miguel Ángel Zubimendi, Alicia Susana Castro y Julián Eduardo Moreno
Miguel A. Zubimendi. Departamento Científico de Arqueología, Museo de La Plata, Buenos Aires, Argentina. Diag. 77 N° 550, 1ro C. E-mail: mikel@lpsat.net
Alicia S. Castro. Departamento Científico de Arqueología, Museo de La Plata, Buenos Aires, Argentina. E-mail: acastro@museo.fcnym.unlp.edu.ar
Julián E. Moreno. CENPAT, Puerto Madryn, Chubut, Argentina. E-mail: julianemoreno@yahoo.com.ar
Recibido 30 de Junio 2004. Aceptado 24 de Noviembre 2004
RESUMEN
En este trabajo se discute el valor de los moluscos en las dietas indígenas de los grupos que ocuparon la Costa Norte de Santa Cruz durante el Holoceno tardío, por medio del potencial alimenticio de los mismos, inferido a partir del análisis del registro arqueológico superficial. Se discute, de esta manera, la imagen arqueológica construida a partir de análisis previos, a la luz de los nuevos resultados centrados en los restos malacológicos. Se considera que la densidad de restos de moluscos en los sitios arqueológicos refleja la importancia dietaria relativa de los mismos. Por ello, se tiene en cuenta la variabilidad de especies de moluscos representados arqueológicamente, cuya presencia presumiblemente, responde a decisiones antrópicas orientadas al consumo alimenticio. Por último, se analiza el aporte calórico estimado por especie en el nivel areal. Dado que es imposible establecer valores absolutos para el aporte calórico total representado en cada sitio, se trabaja con porcentajes relativos como aproximación al aporte diferencial de los moluscos a la dieta indígena. Esta metodología exploratoria y experimental, es una herramienta sencilla y operativa de análisis, que permitirá hacer un aporte inicial, para tratar de responder parcialmente a los interrogantes, así como para discutir algunas conclusiones a las que se han arribado previamente.
Palabras claves: Recursos alimenticios; Moluscos; Distribución de sitios; Densidad de sitios.
ABSTRACT
In this paper the value of mussels in the hunter-gatherer diet of the northern coast of Santa Cruz in the Late Holocene is discussed through the alimentary potential inferred from the archaeological record. As such, the archaeological image constructed from previous analyses is discussed in light of new results obtained from the analysis of the mussel remains. The density of mussel shell reflects the relative dietary importance of this resource. In relation to that idea we considered the variability of species represented archaeologically, the presence of which presumably represents the result of human decisions taken in relation to food consumption. Given that it is impossible to establish absolute values for the total calorific contribution of each site relative percentages are used to approximate the differential contribution of these mollusks to the indigenous diet. This exploratory and experimental method is a simple and operative analytical tool that enables an initial response, partial to the questions raised, as well as to discuss some conclusions previously passed over.
Keywords: Economical resources; Mussels; Site distribution; Site density.
ACERCA DE LA ARQUEOLOGÍA DE COSTAS
A juzgar por la literatura surgen muchos problemas cuando se trata de estudiar a los cazadores-recolectores costeros y sus formas de vida (Wasselkov 1987). En las últimas décadas los estudios de los cazadoresrecolectores costeros se han intensificado, impulsados por el interés en las diferentes adaptaciones humanas y, especialmente, por los estudios ecológicos derivados del paradigma ecológico-sistémico. Muchos son los trabajos realizados, pero quizá el problema que ha provocado más controversias entre los investigadores es el referido al valor de los recursos alimenticios y el de los hábitats costeros para los grupos humanos. La productividad costera Las zonas costeras, donde ocurre el encuentro de recursos marinos y terrestres en un área geográfica limitada, son en general altamente productivas. Suelen caracterizarse por la abundancia y concentración de recursos alimenticios junto a una fauna típica sólo presente en la zona intermareal (i.e., Bailey y Parkington 1988:1; Yesner 1980:728). De manera muy general es posible definir a los recursos costeros como aquellos recursos orgánicos e inorgánicos que están localizados en esas aguas o que trascienden el límite entre tierra y agua (Perlman 1980:259).
Para muchos investigadores, los ambientes costeros, en especial ciertas costas favorecidas, proporcionan una diversidad de recursos extremadamente ricos que habrían sido explotados en momentos tempranos en diferentes partes alrededor del mundo (i.e., Moseley 1975; Rick y Erlandson 2000). Algunos autores incluso postulan que estos ambientes han conducido a altas densidades de población y a comunidades más sedentarias (Perlman 1980), así como al desarrollo de sociedades de cazadores-recolectores complejos (i.e., Lig htfoot 1993:170). A pesar de esto, para otros investigadores los hábitats costeros son solo estacionalmente ricos (Parkington 1972). Mientras que otros opinan que los ambientes costeros y los recursos marinos, en especial los moluscos, son relativamente inferiores en cuanto a su valor frente a los recursos y hábitat terrestres. Para estos, la ocupación costera sería consecuencia de situaciones de estrés medioambiental o poblacional en sectores mediterráneos considerados más favorecidos ecológicamente (Bailey 1975; Beaton 1985; Broughton y O'Connell 1999; Osborn 1977, entre otros).
En realidad, los estudios arqueológicos han avalado todas las posiciones planteadas más arriba. Pero, más allá de estas discusiones, el valor de los recursos costeros es muy variable y situacional, dependiendo entre otras cosas de la latitud, el tipo de costa, la productividad de la zona terrestre adyacente y la dinámica oceánica (Jones y Hildebrandt 1995:80). Además de otros factores como la temperatura del agua, el sustrato de la plataforma costera, la salinidad del agua, la disponibilidad de alimentos y oxígeno, etc. (Claassen 1998:126; Pastorino, 2000:152; Perlman 1980:260). Estos factores provocan en los ambientes costeros una pauta muy estructurada y visible de disponibilidad de recursos, siendo el más importante para el análisis aquí realizado, los intermareales, donde se pueden formar bancos de moluscos o restingas.
La existencia y productividad de los intermareales se ve condicionada por la topografía, la sedimentología y la hidrología local. La pendiente del terreno determina la extensión de la cobertura de marea alta; las altas pendientes reducen el área de mareas y la cantidad de restingas productivas. El tipo de substrato condiciona en forma local la presencia de especies, existiendo una correlación entre el tipo de organismo y el sedimento en que se desarrolla, pudiendo éste ser separado en dos grupos: de fondo blando y de fondo duro o rocoso (Pastorino 2000:152). La batimetría local afecta las productividades acuáticas al influir sobre la tensión de energía del oleaje. Estas variables locales determinan una estructuración de los intermareales llamada zonación vertical (Claassen 1998:128). Cada especie tiene sus requerimientos específicos; los mejillones (Mytilus edulis platensis) son bivalvos sésiles, tienen un rango de hábitat limitado y una distribución espacial estable, suelen habitar costas rocosas con alto estrés de ola donde crecen más lentamente y con valvas más gruesas (Jones y Richman 1995:34-35). Las lapas (Nacella sp.) (=Patinigera) son gasterópodos y también tienen un rango de habitabilidad limitado con valvas más altas a mayor estrés de ola (Morriconi y Calvo 1993; Zuschin y Stanton 2001).
Como ya se mencionó, el estrés de ola es un factor fundamental que participa en la delimitación de la zonación de las restingas, ya que promueve algunas especies y puede determinar una mayor productividad, mientras que a su vez condiciona negativamente el desarrollo de otras. Pero, básicamente, el desarrollo de amplios bancos de moluscos en la región intermareal depende de dos cosas: biomasa marina, dentro de ella de la producción primaria, y del contexto geológico. El soporte para la zona intermareal es fundamental, ya que determina una particular ecología que será sustento para las especies. Los mejillones y las lapas suelen estar asociadas a un substrato rocoso, mientras que las almejas prefieren un substrato más limoso. A su vez los rodados inhiben el desarrollo de estas especies.
El consumo de moluscos en las dietas indígenas
El tratamiento de los moluscos en los cazadoresrecolectores también ha sido discutido durante los últimos 30 años (Bailey y Parkington 1988; Orquera 1999; Orquera y Piana 2000; Osborn 1977; Parmalee y Klippel 1974; Yesner 1980). Hay quienes ven a este recurso como marginal en comparación con las tasas de rendimiento de otros recursos, especialmente los terrestres. Hay que recordar que la función del recurso malacológico no era únicamente la nutricional, sino que también podían cumplir otras funciones: adornos (con o sin previa formatización), instrumentos para cortar, desbastar, recipientes o platillos, instrumentos musicales (Claassen 1998) e inclusive para Patagonia se afirmó que servían como "instrumento de depilación" (Deodat 1967). Más allá de estos usos diversos, no hay duda de que en su gran mayoría fueron empleados como recurso alimenticio. Los moluscos constituyen una excelente fuente de calcio, yodo, electrolitos y otros minerales, aunque es probable que parte de estos elementos se pierdan en su procesamiento como alimentos (Jones y Richman 1995:42). Sin embargo, siempre se ha enfatizado su rol como proveedores de proteínas (Erlandson 1988), especialmente en el momento previo al desove.
Algunos autores consideran a los moluscos como suplementarios en momentos de relativa escasez de alimentos y consideran que estos no son atractivos para las poblaciones humanas (Bailey 1975; Bailey y Parkington 1988; Osborn 1977; Parmalee y Klippel 1974; Yesner 1980), constituyendo una estrategia óptima frente a los recursos terrestres bajo ciertas circunstancias y períodos de tiempo. Otros, los consideran complementarios en determinados momentos (Claassen 1986:33; Perlman 1980:286-287) cuando servirían como "válvula de seguridad para superar situaciones de insuficiente disponibilidad de otros recursos más sustanciosos" (Orquera 1999:320). Cualquiera fuese el caso, una dieta fundada solamente sobre mariscos no es adecuada y es peligrosa por ser pobre en calorías, aunque sea apropiada en proteínas y minerales. A su vez, el volumen de carne necesario para satisfacer las necesidades mínimas del ser humano con una dieta exclusivamente constituida por mariscos es inaceptable para la capacidad normal del estómago humano (Orquera 1999:314-315).
Los moluscos también están indicados como un recurso altamente concentrado (con un mínimo esfuerzo en su búsqueda), fácilmente recolectable, sin inversión de energía y por todos los segmentos de la población humana (mujeres, niños y ancianos); por lo que quizás su mayor valor sea su fácil y homogénea accesibilidad para los miembros de movilidad restringida dentro los grupos cazadores-recolectores (Bailey y Parkington 1988:2 y 6; Claassen 1998; Hildebrandt y Jones 1992:360; Jones 1991:436; Yesner 1980). También se han estudiado las causas que provocan el consumo de diferentes especies en los sitios, por ejemplo, la sobreexplotación (Cabral y da Silva 2003; Jerardino 1997; Mannino y Thomas 2001), el método de recolección (Bird y Bird 1997, 2000), o causas culturales y/o simbólicas (Ezzughayyar et al. 1996:3). En consecuencia, los moluscos son un recurso potencial importante en las dietas prehistóricas y en las interpretaciones que surgen del análisis arqueológico de los mismos porque pueden vincular múltiples aspectos de las sociedades humanas (Claassen 1998). Todas estas discusiones han contribuido con interesantes ideas para el debate, pero aún los datos son incompletos o inadecuados para resolver el problema y, según algunos autores, dada la imposibilidad de hacerlo desde la perspectiva teórica la mejor manera de abordarlo es en forma particular en cada ambiente y latitud, desde una perspectiva arqueológica (Jones 1991).
LA ARQUEOLOGÍA EN LA COSTA NORTE DE SANTA CRUZ
Lo dicho hasta aquí se aplica perfectamente al caso de la Costa Norte de Santa Cruz. Se cree que pequeños aportes pueden enriquecer una discusión más general. La Costa Norte de Santa Cruz ha sido descripta y definida en otros trabajos (Castro et al. 1999, 2001, 2003; Moreno 2003; Moreno y Castro 1999). Se denomina costa, a la franja de territorio replegada sobre la actual línea de mareas, donde se tiene acceso a la productividad de los recursos de ambos ambientes, limitada a los primeros 100 m a partir de la línea de tormenta (Castro et al. 2003:72).
En trabajos previos se planteó una estrategia de análisis diagnóstica amplia y no intrusiva que permitiera alcanzar un conocimiento general del universo dentro del cual deberían desarrollarse los estudios de arqueología costera (Castro et al. 2003). El objetivo de dichos estudios fue conocer, desde una escala regional, parámetros relativos a la distribución de sitios arqueológicos en el espacio; las relaciones entre estos y los distintos tipos de costa así como los recursos presentes; finalmente, establecer los patrones de asentamiento.
El litoral del área de estudio se caracteriza por una geomorfología muy variada donde se pueden reconocer distintos tipos de costas. En el sector sur del Golfo San Jorge entre Caleta Olivia y el límite interprovincial con la provincia del Chubut se presentan amplias playas de arena con amplios intermareales, aunque sin restingas. Otras playas del mismo sector son de rodados con gran inclinación, expuestas al oleaje y sin restingas (playa céntrica de Caleta Olivia). A su vez, hay sectores de playas de poca inclinación e intermareales amplios de restingas formadas por la litificación de substratos sedimentarios (bahía Lángara). Finalmente, en algunos puntos, las terrazas altas se acercan a éstas formando costas acantiladas.
Hacia el sur, se encuentra un sector de costa abierto hacia el Atlántico, el cual se extiende desde Cabo Blanco hasta Puerto Deseado con playas de rodados y de marcada inclinación. Estas playas configuran costas acantiladas y regulares, con playas angostas, con escasos intermareales y sin restingas desarrolladas. El sector de costa al sur de la ría Deseado presenta características contrastantes. El mismo posee un perfil recortado con un área de gran cantidad de islas e islotes y pequeñas playas de arena que se intercalan entre afloramientos rocosos (bahía de los Nodales). Allí donde afloran rocas volcánicas (Punta Medanosa y bahía Laura) se desarrollan restingas muy ricas en especies comestibles. Al sur y al norte de la misma, se hallan playas bajas de arena o de rodados y acantiladas alternadas.
Las investigaciones arqueológicas realizadas han permitido alcanzar algunas conclusiones relativas a los patrones de distribución de sitios. Se observa una correlación entre la densidad de sitios y la productividad costera. Aquellos sectores de costa con mesolitorales más amplios y con desarrollo de colonias de moluscos en las restingas son los de mayor densidad de restos arqueológicos (ver Castro et al. 2003). Por lo que las colonias de moluscos serían gravitantes en la elección de los espacios ocupados dentro de una estrategia y una planificación en el uso del ambiente y sus recursos.
En trabajos previos (Castro et al. 2001) los sitios hallados en la costa han sido clasificados en tres tipos: concheros, enterratorios y sitios en general. Posteriormente, se han definido tipologías más especificas para dar cuenta de la variabilidad interna de cada tipo (enterratorios en Castro y Moreno 2000 y en Zubimendi et al. 2004). Los concheros conforman la mayoría de los sitios en la Costa Norte. Están estructuralmente compuestos por restos de valvas asociados a artefactos líticos, restos arqueofaunísticos y fogones antrópicos, producto de acumulaciones de descarte de diversas actividades.
Zonación vertical y patrón de presencia de especies de moluscos en la Costa Norte
Las especies de moluscos presentes en la Costa Norte de Santa Cruz de interés y analizadas en este trabajo pertenecen a la Provincia Biogeográfica Magallánica y son las lapas (Nacella sp.) (=Patinigera), los mejillones (Mytilus edulis platensis), los caracoles (Adelomedon sp.) y las almejas (Ameghinomya antiqua). También se hallan en el registro arqueológico cholgas (Aulacomya ater ater), "picolocos" (Balanus sp.) y mejillines (Brachiodontes purpuratus) (=Perumytilus purpuratus). Estas especies se presentan en las restingas en una zonación en horizontes y cada especie se distribuye de manera diferencial. A las lapas se las encuentran preferentemente en el mesolitoral medio e inferior de las costas rocosas (Otaegui 1974) y son más abundantes en el inferior (Otaegui y Zaixso 1974). Mientras que los mejillones constituyen parte de la comunidad de mejillín (Ringuelet et al. 1962), tienen muy pocos individuos en el mesolitoral superior y aumentan en cantidad en el medio si las condiciones son favorables y más aún en el inferior (Olivier et al. 1966; Zaixso y Pastor 1978). Cuando el mesolitoral está formado por substratos limosos, el mejillinar desaparece (Zaixso 1975). También están ausentes en aquellas paredes del substrato duro, protegidas de la agitación de las aguas (Olivier et al. 1966).
Puede existir una zona de transición entre el meso e infralitoral según la pendiente. Esta zona de transición caracteriza por la presencia de lapas, mejillones y la aparición de cholgas (Otaegui y Zaixso 1974; Zaixso y Pastor 1978). En el infralitoral se pueden hallar lapas, mejillones y cholgas de mayor tamaño que en el mesolitoral (Callebaut Cardu y Borzone 1979), pero fuera del alcance humano, excepto durante las bajantes excepcionales.
OBJETIVOS Y METODOLOGÍA
El objetivo principal de este trabajo es explorar si la imagen arqueológica obtenida (Castro et al. 2003) y definida como un uso intenso de la costa tiene un correlato significativo, en relación al valor como sustento alimenticio de los conjuntos malacológicos relevados, considerando que la densidad de restos de moluscos puede reflejar la importancia dietaria relativa de los mismos (sensu Glassow y Wilcoxon 1988:40-41). Para tal fin se estudiaron las especies presentes, su distribución y rinde a partir de conjuntos malacológicos relevados en muestreos intrasitios realizados en campañas sucesivas (para más detalle ver Castro et al. 1999, 2001, 2003). En estos trabajos se tomaron como variables de interés la presencia, el número y la densidad de sitios asociados a la línea de costa, así como su relación a los recursos marinos. Se considera a los sitios como una muestra representativa de todas o la mayoría de las actividades llevadas a cabo por un grupo humano en un determinado punto del espacio (ver Rossignol y Wandsnider 1992 y los trabajos allí citados).
En este trabajo el universo estadístico lo constituye la franja costera tal como fue definida más arriba, a lo largo de aproximadamente 420 km de costa. Se muestreó un kilómetro cada 20 km, por lo que las Unidades de Muestreo (denominadas de ahora en más UM) tuvieron una superficie de 1000 m por 100 m, sumando en total 21 UM. En estas se ubicó cada uno de los sitios presentes.
Dentro de cada sitio identificado, se realizó un muestreo intrasitio, según unidades de muestreo de tamaño operativo para trabajar en el campo, cada 5 m a lo largo de una transecta sobre el eje mayor del sitio. Se realizó una lectura y registró todo el material arqueológico presente: moluscos (lapas, mejillones, caracoles y almejas), artefactos líticos y otros restos arqueofaunísticos (para más detalles ver Castro et al. 1999, 2001, 2003). De las 21 UM muestreadas se hallaron sitios arqueológicos en 15 de ellas y se registraron un total de 123 sitios.
Procesamiento y análisis de los conjuntos malacológicos
A partir del conteo de la cantidad de valvas de lapas, mejillones, caracoles y almejas en cada sitio se procedió a hacer un análisis de la diversidad de estas especies. Se entiende a la diversidad en términos de riqueza como la cantidad de clases de moluscos presentes por sitio (Claassen 1998:117), lo que brindaría una idea estimativa de la variabilidad de especies de moluscos y permitiría identificar tendencias generales en el nivel areal.
Para el conteo de las valvas se usó el concepto de nonrepetitive elements (NRE), lo que da una frecuencia del Número Mínimo de Individuos (MNI, Claassen 1998:106). Este se basa en el conteo de elementos no repetitivos para estimar el número de valvas de individuos por especie representado. Los elementos no repetitivos son las partes de las valvas que poseen más posibilidad de preservarse en la matriz malacológica (Claassen 2000; Glassow 2000; Mason et al. 1998, 2000). Los NRE en bivalvos son el umbo (mejillones y almejas) y en univalvos el ápex (lapas) o la columela (caracoles).
Primeramente se trabajó con la variación en las proporciones entre las valvas de las distintas especies y con las proporciones entre las valvas y otros artefactos (líticos y restos arqueofaunísticos). La intención de estos análisis fue ver la diferente representación de los distintos componentes dentro del registro arqueológico. Esto permitió realizar una clasificación de sitios sobre la base de la presencia o ausencia de restos en su estructura (Tabla 1). Se diferenciaron sitios que poseían sólo artefactos líticos y restos óseos, así como otros sólo con valvas y por último sitios con los tres tipos de restos. Otros sitios no brindaron datos cuantitativos debido a su pequeño tamaño o porque no se hallaron restos de ningún tipo. Cabe aclarar que se diferenciaron sitios con muy bajos valores de hallazgos (menos de 5 valvas por sitio) que no se utilizaron para el análisis nutricional, pero sí para el análisis de variedad de especies.

Tabla 1. Cantidad de sitios de acuerdo a la presencia o no de restos en su estructura.
Para poder incorporar en el análisis las diferentes superficies de los sitios fue necesario realizar una corrección a fin de utilizar los datos de una manera comparativa a lo largo de las 21 UM, dado que el tamaño de la grilla utilizada en el muestreo intrasitio varió en las distintas campañas. Esta corrección se realizó con la estimación de la superficie de cada una de las grillas extrapolándolas a la superficie de 1 x 1 m, a pesar de que el uso de una grilla de pequeño tamaño puede alterar la muestra haciendo que no aparezcan representadas especies de baja frecuencia. Estos valores fueron luego corregidos para estimar la densidad de restos de valvas por m2 y por sitio, la densidad de artefactos (valvas sumadas a líticos y óseos) total por m2. A su vez y como medida de control de la validez de los datos empleados, se obtuvo un índice de la superficie muestreada de los sitios calculando la superficie total del mismo y dividiéndola por la del total de las grillas muestreadas de un sitio.
A partir del número estimativo de valvas se trató de establecer la cantidad de individuos que estos valores representaban. Se procedió dividiendo la cantidad de valvas de mejillones y de almejas por 2 y en caso de dar fracciones se tomó el número entero superior. Para lapas y caracoles no se realizó ninguna operación matemática adicional. Posteriormente, se multiplicaron estos valores por la superficie del sitio para obtener así un estimativo del número de individuos representados.
Hasta aquí se trabajó con las 4 especies de invertebrados de interés en este trabajo; pero el análisis de aporte calórico se centró en las lapas y los mejillones, por su representatividad estadística como producto de mayor importancia en la dieta indígena. Se parte del supuesto de que los restos del sitio son representativos de las proporciones en que los recursos -malacológicos- contribuirían a la dieta (Perlman 1980:288) y no que los restos son representativos del consumo real que se produjo en el sitio. Para este cálculo es necesario establecer los tamaños de individuos y utilizar un tamaño promedio. Este análisis se basó en las mediciones que se realizaron sobre muestras obtenidas en la localidad de Punta Medanosa en particular. Estos datos se proyectaron como modelo al resto de las muestras de la Costa Norte, hasta tanto se complete este estudio para otros sectores (Castro et al. 2001).
En cuanto al aporte calórico se toman los valores usados por Orquera (1999:311 y Cuadro 1) teniendo en cuenta que pueden existir diferencias entre las especies de un lugar y otro, es decir, los moluscos del Canal Beagle y de la costa continental. En virtud de ello, se analizaron comparativamente el tamaño promedio de distintas muestras de mejillones y lapas del Canal Beagle (provincia de Tierra del Fuego) y de Punta Medanosa (Costa Norte de Santa Cruz). En estos análisis se observaron que las valvas de mejillón enteras provenientes de Punta Medanosa son más grandes que todas las halladas en el Canal Beagle y sólo menores a las de tamaño óptimo de la muestra actual. Para las lapas se observa una tendencia similar siendo sólo los hallados en el sitio Imiwaia I más grandes que las de Punta Medanosa.
Si bien el aporte calórico individual sería diferente entre una muestra y la otra, debido a que el promedio de longitud de las valvas es distinto, se extrapolaron de manera tentativa y experimental los valores calóricos dados por Orquera (1999: Cuadro 1) para ambos recursos del Canal Beagle, a los de Punta Medanosa. Se considera que esta comparación es útil a los fines de observar la diferencia en el aporte calórico relativo entre especies, no así si se quisiera hacer un análisis del aporte real de las especies en cada sitio.
El valor usado para mejillones es de 1,59 kcal por individuo, valor más alto dado por Orquera (1999), cuya longitud promedio es de 43,8 ± 3,4 mm; siendo el de Punta Medanosa de 54,96 ± 8,75. Mientras que para lapas es de 1,16 kcal por individuo, cuya longitud promedio es 32,5 ± 2,3 mm en el Beagle y de 39,65 ± 4,24 para Punta Medanosa.
El número de individuos de lapas y mejillones por sitio se multiplicaron por el valor de aporte calórico. Así se obtuvo el porcentaje del aporte relativo para cada especie, como el aporte del sitio. Este valor es considerado en términos relativos para la comparación entre sitios y cada sector de costa. Se analizó la distribución de frecuencias del aporte calórico, para estimar si brindan información sobre patrones diferenciales en el uso de las costas en los dos sectores analizados.
A los fines de resumir la información y hacerla más clara se presentan los resultados en una única tabla (Tabla 2), obviando la descripción detallada de los resultados por UM y sitios (para la descripción de las UM, ver Castro et al. 1999, 2003).
Tabla 2. Resultados de los análisis para todos los sitios y todas las UM.
DISCUSIÓN
De los análisis realizados se desprende que hubo un uso de los recursos marinos invertebrados no homogéneo a lo largo de la Costa Norte de Santa Cruz. Esto implicaría un uso diferencial de ciertos sectores de la misma y un aprovechamiento planificado, según las características y la disponibilidad de estos recursos litorales en cada sector. En cuanto a la variedad de especies, se observa que al sur de la ría Deseado están ausentes las almejas, las cuales aparecen, aunque en poca cantidad, en la zona del Golfo. Esto se podría deber a que las mismas necesitan un substrato blando, de arenas finas, para vivir, el cual se presenta intermitentemente entre Caleta Olivia y el límite interprovincial. Contrariamente, los caracoles se hallan presentes casi en la totalidad de las UM, aunque en mucha menor proporción que las lapas y mejillones (ver Tabla 2).
Existe además una diferencia en cuanto a la proporcionalidad numérica de valvas en cada uno de los dos sectores. En el Golfo, se observa una proporción relativamente alta tanto para lapas como para mejillones, mientras que los restos de caracoles y almejas tienen una muy baja representatividad. Al sur del Deseado dominan ampliamente los mejillones, le siguen en importancia las lapas y aumentan levemente los caracoles. Estos valores se modifican si los analizamos por individuos, las lapas dominan numéricamente en el Golfo en una relación de 2:1 frente a los mejillones. Al sur del Deseado la relación se invierte (ver Tabla 2). Esta diferencia se debería a condiciones ecológicas que favorecen el desarrollo de determinadas especies. Las UM en que predominan las lapas presentan mesolitorales duros (Castro et al. 2003: Tabla 1), mientras que en las UM 10 y 14 el intermareal está compuesto por rodados y rocas. Otra razón de esta diferencia podría ser que, allí donde se hallen juntos las lapas y los mejillones, y si los mismos van a ser transportados a cierta distancia, se elijan las primeros que tendrían un mayor rinde por unidad de peso vivo y que al ser recolectados retienen menor cantidad de agua (E. Piana, comunicación personal, 2003).
En las UM donde predomina el mejillón, los intermareales podrían definirse como mixtos, debido a la presencia de substratos duros y blandos a intervalos, excepto la UM 2, en la cual el intermareal es de substrato duro. En las UM que presentan proporciones similares de mejillones y lapas sus intermareales pueden ser duros o mixtos. Mientras que el resto de las UM, donde no se halló ningún sitio, están asociadas a playas de rodados y por lo tanto, sin restingas.
Si se considera el aporte calórico estimado para cada UM, se desprende que hay dos subsectores que sobresalen por su productividad relativa (ver Figura 1a. y 1b.):
a) el centro-sur del Golfo San Jorge (UM 4, 5 y 6), con el 66% del aporte del sector;
b) bahía de los Nodales, al sur del Deseado (UM 16, 17 y 18), con el 93% del aporte calórico del sector.
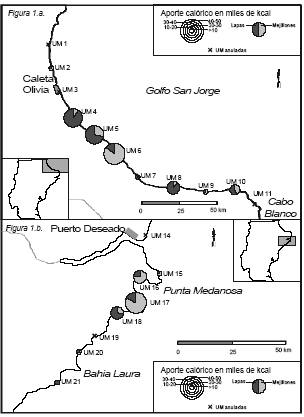
Figura 1. Aporte calórico: (a) en el Golfo San Jorge; (b) al Sur de la ría Deseado.
No obstante, en el Golfo San Jorge existen dos UM (las 8 y 10) con una productividad en kcal relativamente importante. Aunque no es posible definirlas como subsectores aparte, ya que aparecen como casos aislados al existir, colindantes a ellos, UM de menor productividad. En el Golfo San Jorge no se presenta una concentración tan marcada de los sitios con mayor aporte calórico como al sur del Deseado. Por ejemplo, el sitio con mayor aporte calórico no se halla en el subsector centro-sur descrito más arriba, sino en la UM 8. Es de notar que no hay ninguna UM, sin evidencias de consumo de moluscos, lo que implica que probablemente la totalidad de la costa del Golfo ha sido usada en algún momento. Esta característica del registro arqueológico de este sector, estaría en relación a la configuración del litoral, el cual es lineal, con restingas, donde los bancos de moluscos se extienden a todo lo largo de la costa, excepto en aquellos sectores de playas de rodados, los cuales no suelen ser muy extensos.
En este sector del Golfo San Jorge los recursos alimenticios estarían distribuidos homogéneamente. La presencia de estos recursos (especialmente los moluscos) sería altamente segura y predecible, lo que favorecería un uso indistinto de la costa. A su vez, no se registran históricamente grandes colonias de lobos marinos (Carrara 1952) que es el recurso nutricional más importante (Moreno 2003) para los visitantes humanos en esas localidades, por lo que la posibilidad de encontrar lobos marinos en la costa, sería prácticamente igual en cualquier sector de la misma. Así, la captura de lobos marinos sería una actividad impredecible y aleatoria. Esto permite pensar que si bien la costa del Golfo presenta el recurso seguro en abundancia, la posibilidad de captura del recurso principal al quedar librada al azar, aumentaría el riesgo y condicionaría el uso de la misma.
En el sector de costa al sur del Deseado se da una clara concentración espacial de los sitios con mayor aporte calórico a favor del subsector bahía de los Nodales, donde no sólo está concentrado el 93% del aporte calórico del sector, sino que los 6 sitios con un aporte calórico mayor a 100.000 kcal se hallan dentro del mismo. El litoral de este sector está caracterizado por grandes afloramientos de rocas volcánicas ignimbritas, los cuales configuran costas muy recortadas e irregulares con características particularmente sobresalientes (ver especialmente Castro et al. 2001).
Esta configuración recortada hace que aumente el largo de la costa en una menor distancia lineal y por lo tanto estén disponibles desde una misma locación un mayor número de bancos de moluscos. Además, las restingas son más cortas pero más ricas en cuanto a algunas especies de moluscos, como los mejillones y, en consecuencia, más productivas. Estas costas están asociadas a otros recursos alimenticios marinos, en especial a lobos marinos y gran cantidad de aves (Carrara 1952; Castro et al. 2004).
La costa al sur del Deseado, por lo tanto es visualizada como más heterogénea, con áreas muy productivas (bahía de los Nodales) y otras no (i.e., bahía Desvelos). En estas últimas, con playas de rodados, el recurso seguro no estaría disponible. Lo que habría condicionado un menor uso y aprovechamiento de los otros recursos presentes, ya que implica un aumento en el riesgo por ausencia de los bancos de moluscos. Mientras que en bahía de los Nodales los moluscos estarían disponibles en abundancia dada su distribución espacial, accesibilidad y su mayor aporte cárnico. Estas características la convierten en un área muy segura, donde se podría volver con recurrencia, o donde se podría permanecer por más tiempo. Además, la posibilidad de encuentro del recurso principal costero (los lobos marinos) sería mayor en este subsector ya que han existido históricamente gran número de colonias de reproducción.
Dada esta configuración de los recursos litorales, la distribución de los sitios podría ser producto de una reocupación genérica en el caso del Golfo San Jorge, con un patrón distribucional más homogéneo; y una reocupación especifica en el caso al sur del Deseado, con un patrón más heterogéneo de distribución de sitios. Los subsectores donde habría una mayor densidad de sitios serían aquellos donde el recurso complementario, los moluscos, estarían asociados a otros recursos como lobos marinos.
Dentro de este esquema, si bien los moluscos no serían el recurso que aportara los nutrientes principales en la dieta indígena, sí serían la "válvula de seguridad" (Orquera 1999:320) necesaria para el asentamiento en la costa. En tal sentido es que serían un recurso gravitante en la elección del lugar de asentamiento en el espacio.
En la Figura 2a se ha representado la distribución de sitios según rangos de 10.000 kcal en los dos sectores. Se observa que las diferencias del aporte calórico de los moluscos entre los dos sectores producen un patrón distinto en cada uno. En el Golfo San Jorge se daría una tendencia decreciente en la distribución de frecuencias de sitios según aumenta el aporte calórico (ver también Figura 2b). Al sur del Deseado se observa un patrón alternante con una tendencia general decreciente. En la Figura 2c existen tres claras agrupaciones de sitios según su aporte calórico, separados por "saltos". El primer grupo tiene un aporte calórico menor a 10.000 kcal (distribuido uniformemente en todas las UM), el segundo grupo tiene un aporte calórico entre 20.000 y 32.000 kcal, mientras que el tercero entre 115.000 y 130.000 kcal (ver Figura 2c). Los sitios dentro de estos dos últimos grupos se ubican en bahía de los Nodales.

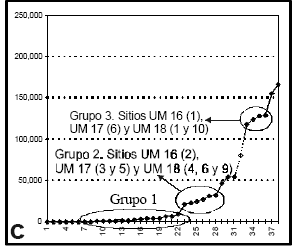
Figura 2. Aporte calórico en la Costa Norte de Santa Cruz: (a) distribución de frecuencias de sitios según su aporte calórico en rangos de 10.000 kcal; (b) gráfico acumulativo de sitios según su aporte calórico en el Golfo San Jorge; (c) gráfico acumulativo de sitios según su aporte calórico al Sur del Deseado.
La concentración de sitios en el subsector de bahía de los Nodales ya había sido objeto de un análisis particular (ver Castro et al. 2001). Así, la imagen que se había construido de dicho subsector, en base al tipo de sitios que allí se presentan, concuerda con los resultados alcanzados en este trabajo. Sin embargo, los estudios en el subsector ubicado en el centro-sur del Golfo San Jorge, brindaron una imagen arqueológica diferente, ya que los sitios concheros no "impresionaban" como los de bahía de los Nodales. Pero, los resultados del presente trabajo llevan a la necesidad de cambiar la imagen que se había construido y asignarles una mayor importancia en términos de uso del espacio.
Los resultados obtenidos en este trabajo podrían también permitir discutir el significado de las ocupaciones que estos sitios representan en términos de cantidad de personas, tiempo o formas de consumo. De esta manera se debería discutir la ocurrencia de eventos de consumo intensivo u ocasional, así como de ocupación esporádica o estacional, etc. Pero el análisis particularizado de un recurso resulta insuficiente para ello, siendo necesario contextualizar estos datos con información proveniente del análisis de otros recursos, como físicos, biológicos y análisis cronológicos. No obstante, a fines ilustrativos se puede observar que, por ejemplo, el 92% de los sitios analizados tienen un aporte calórico menor al equivalente a un único lobo marino macho adulto (Orquera 1999:316), mientras que el 70% no alcanza a cubrir 10 días/hombre. Estos datos podrían ser considerados como indicadores de un consumo de moluscos asociado a estadías de corta duración o por parte de pocos individuos. Pero, para poder completar esta imagen arqueológica y corregirla, se debería, como ya se mencionó, analizar el uso de otros recursos económicos y otros condicionantes no económicos en el uso de la costa como aspectos sociales y/o simbólicos.
CONCLUSIONES
El estudio estadístico de las muestras malacológicas de los sitios nos permite afirmar que el valor alimenticio de los moluscos en las dietas indígenas de la Costa Norte de Santa Cruz fue aprovechado por las poblaciones que la habitaron. La capacidad de los moluscos para proveer un recurso alimenticio abundante, sencillo de explotar y fácilmente localizable fue reconocido por las poblaciones aborígenes, evidenciado por los más de 120 sitios que han sido objeto de este análisis.
Se observa que hay una variabilidad relativa entre las especies representadas en los sectores del Golfo San Jorge y al sur de la ría Deseado. En ambos predominan los mejillones y las lapas, pero la tercer especie representada en el primer sector son las almejas, y en el segundo sector, los caracoles.
Respecto a las especies más aprovechadas, las lapas son más numerosas en el Golfo San Jorge, mientras que los mejillones lo son al sur del Deseado. Se pudo demostrar una correlación entre el tipo de intermareal y las especies representadas en las UM. En aquellos intermareales de substrato duro se dio una predominancia de lapas, mientras que en aquellos de tipo mixto dominan los mejillones. Los análisis indican que en términos de individuos se produce un relativo emparejamiento frente a los valores porcentuales de las valvas, aunque se mantiene la tendencia por especies.
Con respecto al aporte calórico de los moluscos representado de manera proporcional en las UM se desprende que hubo dos subsectores de la costa que fueron explotados con mayor intensidad. Estos subsectores corresponden al centro-sur del Golfo San Jorge y bahía de los Nodales, al sur de la ría Deseado. En el primer sector, en todas las UM se registró algún aporte calórico de los moluscos, mientras que al sur del Deseado, en una UM no se hallaron sitios y en otra el aporte fue casi insignificante.
El aporte calórico de los sitios según una distribución de frecuencias nos brinda un patrón decreciente en el Golfo San Jorge a mayor aporte. Mientras que al sur del Deseado el patrón es alternante, con una tendencia general decreciente y agrupaciones de sitios de similar aporte calórico. Los sitios con mayor aporte calórico corresponden al subsector de bahía de los Nodales. Se infiere que los sitios con mayor aporte se encuentran en las zonas de la costa donde se hallan aquellos recursos que proporcionan el grueso de los requerimientos energéticos de subsistencia, como los lobos marinos; junto con los moluscos, los cuales actuarían como amortiguadores en caso de no obtener el recurso principal algún día, o como aporte energético extra. Por lo tanto, la importancia del recurso moluscos no estaría dada sólo por su aporte a la dieta, sino también por todas aquellas características de los bancos de moluscos (disponibilidad, predictibilidad, accesibilidad), que hacen que influyan en la elección del lugar de asentamiento de un grupo humano.
En respuesta a los objetivos principales de este trabajo, se puede decir que el potencial alimenticio de los moluscos en las dietas indígenas de las poblaciones que habitaron la Costa Norte de Santa Cruz es el de proveer el recurso alimenticio de mínima necesario para sobrevivir, y que su valor está dado por la seguridad que brindan en cuanto a su localización en el espacio y a su abundancia.
En términos de sustento de la población, la imagen de un uso intensivo de la costa proporcionada por los sitios concheros deberá ser rediscutida a partir de los datos surgidos de este trabajo, integrando el análisis de otras variables y relativizándolas en el tiempo y el espacio. En el tiempo, ya que se debería hablar de ocupaciones poco densas pero reiteradas para gran parte de la costa, lo que daría arqueológicamente una imagen de alta densidad de sitios. En el espacio, porque los distintos sectores de costa fueron ocupados de manera diferencial en relación, posiblemente, a decisiones basadas en criterios de reducción de riesgo a partir de la disponibilidad de recursos marinos.
Finalmente, este trabajo ha sido útil para aportar nuevos datos sobre el uso de la Costa Norte de Santa Cruz, pero principalmente para generar nuevas preguntas, las cuales deberán ser encaradas en futuras investigaciones y para comprender mejor las interpretaciones sobre el uso de la costa. Por ello, se cree que la construcción de modelos sencillos, operativos y que simplifican la realidad del registro arqueológico es útil y necesario para las investigaciones que se vienen realizando en este ámbito particular de la arqueología patagónica.
Agradecimientos
Estos trabajos fueron realizados en el marco de un subsidio del FONCyT, PICT 98-04-04112, y de la UNPA, acreditación 2002. Agradecemos especialmente al Dr. Ricardo Bastida por la orientación que nos proporcionó sus comentarios científicos y en especial por las conversaciones amenas, al Sr. Juan Heupel por el apoyo local que siempre brinda, y a Lucía Mazzitelli por la colaboración en la elaboración de este trabajo.
REFERENCIAS CITADAS
Bailey, G. N. 1975 The role of molluscs in Coastal Economies: The results of midden analysis in Australia. Journal ofArchaeological Science 2: 45-62. [ Links ]
Bailey, G. N. y J. Parkington 1988 The archaeology of Prehistoric Coastlines: an introduction. En The archaeology of prehistoric coastlines, editado por G. Bailey y J. Parkington, pp. 1-10. Cambridge University Press, Nueva York. [ Links ]
Beaton, J. M. 1985 Evidence for a Coastal occupation time-lag at Princess Charlotte Bay (North Queensland) and implications for coastal colonization and population growth theories for aboriginal Australia. Archaeology in Oceania 20: 1-20. [ Links ]
Bird, D. W. y R. B. Bird 1997 Contemporary shellfish gathering strategies among the Meriam of the Torres Strait Islands, Australia: testing predictions of a central place foraging model. Journal of Archaeological Science 24: 39-63. [ Links ]
Bird, D. W. y R. B. Bird 2000 The ethnoarchaeology of juvenile foragers: shellfishing strategies among Meriam children. Journal of Anthropological Archaeology 19: 461-476. [ Links ]
Broughton, J. M. y J. F. O'Connell 1999 On evolutionary ecology, selectionist archaeology, and behavioral archaeology. American Antiquity 64: 153-165. [ Links ]
Cabral, J. P. y C. A. F. da Silva 2003 Morphometric analysis of limpets from an Iron- Age shell midden found in northwest Portugal. Journal of Archaeological Science 30: 817-829. [ Links ]
Callebaut Cardu, J. y C. A. Borzone 1979 Observaciones ecológicas del infralitoral de Puerto Deseado (Prov. de Santa Cruz, Argentina). I. Península foca. Ecosur 6(11): 45-54. [ Links ]
Carrara, I. S. 1952 Lobos marinos, pingüinos y guaneras del litoral marítimo e islas adyacentes de la República Argentina. Ministerio de Educación. Facultad de Ciencias Veterinarias (publicación especial), UNLP. La Plata. [ Links ]
Castro, A. y E. Moreno 2000 Noticia sobre enterratorios humanos en la Costa Norte de Santa Cruz. Anales del Instituto de la Patagonia, (Serie Ciencias Humanas) 28: 225-233. [ Links ]
Castro A., E. Moreno, K. Martinelli, F. Pepe, V. Díaz y M. Zubimendi 1999 Los asentamientos indígenas tardíos en la Costa Norte de Santa Cruz y su relación con los recursos marinos. Actas del XIII Congreso Nacional de Arqueología Argentina, Córdoba. En prensa. [ Links ]
Castro, A., E. Moreno, M. Andolfo y M. Zubimendi 2001 Distribución espacial de sitios en la localidad de Punta Medanosa, Santa Cruz (Argentina). Relaciones de la Sociedad Argentina de Antropología XXVI: 303-321. [ Links ]
Castro, A., J. Gomez Otero, G. Arrigoni y E. Moreno 2004 Prospección macrorregional a las loberías de la costa atlántica de Patagonia. Algunas claves sobre el uso del espacio y los recursos. En Contra Viento y Marea. Arqueología de Patagonia, editado por M. T. Civalero; P. M. Fernandez y A. G. Guráieb. Instituto Nacional de Antropología y Pensamiento Latinoamericano. pp. 197-215. Buenos Aires [ Links ]
Castro, A., E. Moreno, M. Andolfo, R. Gimenez, C. Peña, L. Mazzitelli, M. Zubimendi y P. Ambrústolo 2003 Análisis distribucionales en la costa de Santa Cruz (Patagonia Argentina): alcances y resultados. Anales del Instituto de la Patagonia, (Serie Ciencias Humanas) 31: 69-94. [ Links ]
Claassen, C. 1986 Shellfishing seasons in the prehistoric Southeastern United States. American Antiquity 51: 21-37. [ Links ]
Claassen, C. 1998 Shells. Cambridge University Press, Cambridge. [ Links ]
Claassen, C. 2000 Quantifying shell: comments on Mason, Peterson and Tiffany. American Antiquity 65: 415-418. [ Links ]
Deodat, L. S. M. 1967 Una antigua manufactura valvácea en el golfo San Matías (Argentina). Runa X: 319-353. [ Links ]
Erlandson, J. 1988 The role of shellfish in prehistoric economies: A protein perspective. American Antiquity 53: 102- 109. [ Links ]
Ezzughayyar, A., M. Al-Zawahra y H. Salem 1996 Molluscan fauna from Site 4 of Tell Jenin (northern west bank, Palestine). Journal of Archaeological Science 23: 1-6. [ Links ]
Glassow, M. 2000 Weighing vs. counting shellfish remains: a comment on Mason, Paterson and Tiffany. American Antiquity 65: 407-414. [ Links ]
Glassow, M y L. Wilcoxon 1988 Coastal adaptations near Point Conception, California, with particular regard to shellfish explotation. American Antiquity 53: 36-51. [ Links ]
Hildebrandt, W. R. y T. L. Jones 1992 Evolution of marine mammal hunting: a view from the California and Oregon coasts. Journal of Anthropological Archaeology 11: 360-401. [ Links ]
Jerardino, A. 1997 Changes in shellfish species composition and mean shell size from a late-Holocene record of the west coast of southern Africa. Journal of Archaeological Science 24: 1031-1044. [ Links ]
Jones, T. L. 1991 Marine-resource value and the priority of coastal settlement: a California perspective. American Antiquity 56: 419-443. [ Links ]
Jones, T. L. y W. R. Hildebrandt 1995 Reasserting a prehistoric tragedy of the commons: reply to Lyman. Journal of Anthropological Archaeology 14: 78-98. [ Links ]
Jones, T. L. y J. Richman 1995 On mussels: Mytilus californianus as a prehistoric resource. North American Archaeologist 16: 33-58. [ Links ]
Lightfoot, K. G. 1993 Long-term developments in complex huntergatherer societies: recent perspectives from the Pacific Coast of North America. Journal of Archaeological Research 1: 167-201. [ Links ]
Mannino, M. y K. D. Thomas 2001 Intensive mesolithic explotation of coastal resources? evidence from a shell deposit on the Isle of Portland (southern England) for the impact of human foraging on populations of intertidal rocky shore molluscs. Journal of Archaeological Science 28: 1101-1114. [ Links ]
Mason, R., M. Peterson y J. Tiffany 1998 Weighing vs. counting: measurement realiability and the California school of midden analysis. American Antiquity 63: 303-324. [ Links ]
Mason, R., M. Peterson y J. Tiffany 2000 Weighing and counting shell: a response to Glassow and Claassen. American Antiquity 65: 757- 761. [ Links ]
Moreno, E. 2003 El uso indígena de la Costa Patagónica Central en el Periodo Tardío. Tesis Doctoral inédita. Facultad de Ciencias Naturales y Museo, Universidad Nacional de La Plata. La Plata. [ Links ]
Moreno, E. y A. Castro 1999 Costa Norte de Santa Cruz como excepción al modelo de cazadores de guanaco. Anales de Arqueología y Etnología 50-51: 13-22. [ Links ]
Morriconi, E. y J. Calvo 1993 Influencia ambiental sobre el crecimiento alométrico de la valva en Nacella (Patinigera) deaurata (Gmelin, 1791) del Canal Beagle. Malacología 35: 135-140. [ Links ]
Moseley, M. E. 1975 The Maritime Foundation of Andean Civilizations. Cummings, Menlo Park, California. [ Links ]
Olivier, S. R., I. Kreibhom de Paternoster y R. Bastida 1966 Estudios biocenóticos en las costas de Chubut (Argentina) I. Zonación biocenológica de Puerto Pardelas (Golfo Nuevo). Boletín N° 10 del Instituto de Biología Marina: 5-74. Mar del Plata. [ Links ]
Orquera, L. A. 1999 El consumo de moluscos por los canoeros del extremo sur. Relaciones de la Sociedad Argentina de Antropología XXIV: 307-327. [ Links ]
Orquera, L. A. y E. L. Piana 2000 Composición de conchales de la costa del Canal Beagle (Tierra del Fuego, República Argentina) - Primera parte. Relaciones de la Sociedad Argentina de Antropología XXV: 249-274. [ Links ]
Osborn, A. J. 1977 Strandloopers, mermaids and other Fairy tales: Ecological determinants of marine resources utilization - the Peruvian case. En For Theory Building in Archaeology, editado por L. Binford, pp. 157-205. Academic Press, Nueva York. [ Links ]
Otaegui, A. V. 1974 Las especies del género Patinigera (Dall 1905) en la provincia Magallánica (Mollusca, Gastropoda, prosobranchiata). Physis 33(86): 173-184. [ Links ]
Otaegui, A. V. y H. E. Zaixso 1974 Distribución vertical de los moluscos marinos del litoral rocoso de la Ría de Puerto Deseado (Santa Cruz). Una guía para reconocer los diferentes pisos y horizontes litorales. Physis 33(86): 321-334. [ Links ]
Parkington, J. E. 1972 Seasonal mobility in the late Stone Age. African Studies 31: 223-243. [ Links ]
Parmalee, P. y W. Klippel 1974 Freshwater mussels as a prehistoric food resource. American Antiquity 39: 421-434. [ Links ]
Pastorino, G. 2000 Asociaciones de moluscos de las terrazas marinas cuaternarias de Río Negro y Chubut, Argentina. Ameghiniana 37(2): 131-156. [ Links ]
Perlman, S. M. 1980 An optimum diet model, coastal variability and hunter-gatherer behaviour. En Advances in Archaeological Method and Theory 3, editado por M. B. Schiffer, pp. 257-310. Academic Press, Nueva York. [ Links ]
Rick, T. C. y J. M. Erlandson 2000 Early Holocene fishing strategies on the California Coast: evidence from CA-SBA-2057. Journal of Archaeological Science 27: 621-633. [ Links ]
Ringuelet, R. A., A. Amor, N. H. Magaldi y R. E. Pallares 1962 Estudio ecológico de la fauna intercotidal de Puerto Deseado en Febrero de 1961 (Santa Cruz, Argentina). Physis 23(64): 35-54. [ Links ]
Rossignol, J. y L. Wandsnider (eds) 1992 Space, time and archaeological landscapes. Plenum Press, Nueva York. [ Links ]
Wasselkov, G. A. 1987 Shellfish gathering and shell midden archaeology. En Advances in Archaeological Method and Theory 10, editado por M. B. Schiffer, pp. 93- 210. Academic Press, Nueva York. [ Links ]
Yesner, D. R. 1980 Maritime hunter-gatherers: ecology and prehistory. Current Anthropology 21: 727-750. [ Links ]
Zaixso, H. E. 1975 Distribución vertical de los moluscos marinos de la Ría Deseado (Santa Cruz, Argentina). Sustrato con fracción limosa. Physis, sección A, 34(89): 229- 243. [ Links ]
Zaixso, H. E. y C. T. Pastor 1978 Observaciones sobre la ecología de los mitílidos de Puerto Deseado. I. Distribución y análisis biocenótico. Ecosur 4: 1-46. [ Links ]
Zubimendi, M. A., A. Castro y E. Moreno 2004 Hacia la definición de modelos de uso de la Costa Norte de Santa Cruz. Magallania 32: 85-98. [ Links ]
Zuschin, M. y R. J. Stanton, Jr. 2001 Experimental measurement of shell strength and its taphonomic interpretation. Palaios 16: 161- 170. [ Links ]

