










Servicios Personalizados
Revista

Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkIntersecciones en antropología
versión On-line ISSN 1850-373X
Intersecciones antropol. n.7 Olavarría ene./dic. 2006
Anatomía económica de Rheidae
Miguel A. Giardina
Miguel A. Giardina. CONICET. Departamento de Antropología,. Museo de Historia Natural de San Rafael. Parque Mariano Moreno s/n. (5.600) San Rafael, Mendoza. E-mail: miguelgiardina@hotmail.com
Recibido 30 de Junio 2005. Aceptado 17 de Enero 2006
RESUMEN
En el siguiente trabajo se presentan los Índices de Rendimiento Económico para Rheidae (Clase: Aves), los cuales incluyen los cálculos del Índice de Utilidad General Modificado (Modified General Utility Index o MGUI) y de los índices que lo componen: el Índice de Utilidad de Carne (Meat Utility Index o MUI), Índice Medular (Marrow Index o MI), Índice General de Grasa (White-Grease Index o WGI) y el Índice de Utilidad General (General Utility Index o GUI). Se presenta además la metodología implementada para la obtención de cada uno de los mismos y se discuten las implicancias de los resultados a la luz de las expectativas generadas para los conjuntos arqueofaunísticos. Los resultados obtenidos muestran valores netos de rendimiento de carne, asociados a las partes anatómicas correspondientes al esqueleto apendicular, así como altos índices de grasa en las porciones axiales (cintura pélvica y esternón principalmente) y médula en tibiotarso y tarsometatarso. Por último se analiza la correlación existente entre los Índices de Rendimiento Económico y el Índice de Densidad Mineral Ósea, la cual muestra un problema de equifinalidad.
Palabras clave: Zooarqueología; Anatomía económica; Ratites Sudamericanas; Rheidae.
ABSTRACT
Economic anatomy of Rheidae. In this paper, the Economic Indexes for Rheidae (Class Aves), which include the calculation of the Modified General Utility Index (MGUI) and its component indexes, Meat Utility Index (MUI), Marrow Index (MI), White Grease Index (WGI) and General Utility Index (GUI), are presented. Moreover, the methodology implemented for obtaining each index and the discussion of the results in an archeofaunistic setting are also presented. The results obtained show net values of meat performance associated with the anatomical portions belonging to the appendicular skeleton, as well as a high grease index in the axial portions (mainly pelvic girdle and sternum), and a high marrow index in tibiotarsus and tarsometatarsus. Finally, the correlation that exists between the Economic Utility Index and the Structural Bone Density Index is analyzed, showing a problem of equifinality.
Keywords: Zooarchaeology; Economic anatomy; South American Ratites; Rheidae.
INTRODUCCIÓN
Los estudios de rendimiento económico han mostrado su utilidad en la generación de expectativas arqueológicas referidas a la frecuencia diferencial de partes anatómicas en sitios arqueológicos. A partir de la década del 80 y motivados por los trabajos de (Binford 1978), se han realizado estudios de rendimiento económico para grandes mamíferos en diversas partes del mundo (Binford 1978; Metcalfe y Jones 1988; Outram y Rowley-Conwy 1998). Particularmente en Sudamérica se han realizado estudios para animales como guanaco (Lama guanicoe) (Borrero 1990), llama (Lama glama) (Mengoni Goñalons 1991) y huemul (Hippocamelus bisulcus) (Belardi y Gómez Otero 1998). Existen los trabajos de Kooyman (1990) hechos para moa (Aves: Dinornithidae, Anomalopterygidae) referidos a estrategias de cacerías individuales o colectivas. Este autor realiza además un índice de utilidad en kiwi (Apteryx sp.) y a partir de este modelo examina el problema de la cacería de moa en detalle. Pese a la importancia que han tenido los rheidos para la subsistencia humana, hasta el momento no existe este tipo de estudio en aves corredoras sudamericanas de gran porte, solamente se cuenta con trabajos realizados sobre ñandú común (Rhea americana) en cautiverio con propósitos comerciales (Garriz et al. 2003; Sales et al. 1996).
Durante la década del 80, se enfatizaron los estudios sobre aspectos taxonómicos y biogeográficos, referidos a este tipo de aves (Ratites sudamericanos). Estos trabajos en general fueron realizados por paleontólogos y arqueólogos (Caviglia y Figuerero Torres 1976; Miotti y Salemme 1999; Miotti y Tonni 1996; Tambussi y Tonni 1985; Tonni y Laza 1980). El objetivo del presente trabajo es presentar la metodología y los resultados obtenidos a partir del cálculo de los índices de utilidad económica de Pterocnemia pennata, para utilizarlos luego como una herramienta metodológica, que permita una mejor evaluación de la representación anatómica diferencial de los Rheidae en los conjuntos arqueológicos. En primer lugar se presenta la metodología utilizada, para luego calcular los índices de rendimiento económico y, finalmente, se discuten los valores obtenidos.
LA IMPORTANCIA DE LOS RHEIDOS EN EL CONSUMO ABORIGEN
Los estudios etnográficos y fuentes etnohistóricas muestran el gran interés que el consumo de rheidos ha tenido para las poblaciones aborígenes en gran parte de América del Sur (Carman 1973). Las crónicas de numerosos viajeros mencionan aspectos de la explotación de rheidos por comunidades aborígenes del siglo XIX, como así también por parte de pobladores de las regiones pampeana y patagónica de principios del siglo XX (Bond Head 1997; Claraz 1988; Muñiz 1916; Musters 1997 [1871]; entre otros). Estos viajeros en muchos casos relatan la importancia que tenía la cacería de aves silvestres (principalmente Pterocnemia pennata y Rhea americana), como forma de obtener recursos importantes, específicamente grasa y plumas para la subsistencia de los pobladores de Pampa y Patagonia .
Es reiterativa en los relatos etnohistóricos la descripción de "boleadas" colectivas (Agüero Blanch 1969; San Martín 1930). Describen como se reúnen todos los que participan en esta tarea y el papel que realiza cada uno de los integrantes en esta cacería. Al respecto es ilustrativa la frase de Francisco Muñiz "El objetivo que se proponen es: bolear avestruces, sacar las plumas, comer su carne y sus huevos, traer de éstos consigo cuanto más se pueda" (Muñiz 1916:176). George Chaworth Musters (Musters 1997 [1871]) en el libro "Vida entre los Patagones", describe la técnica de la preparación de las aves una vez terminada la cacería; cómo son tratadas las distintas partes del animal; la cocción de la misma (colocando piedras calientes en el interior de la presa) y cómo aprovechaban sus huevos cocinándolos en las brasas.
Los motivos de la caza de los rheidos pueden haber diferido de acuerdo a la estación del año en que ocurría el evento de caza. En Patagonia, según mencionan las crónicas, cuando capturaban estas especies en períodos en los que sus reservas de grasa habían disminuido (comienzo de la primavera), el motivo de la caza era únicamente para la obtención de la piel y plumas. George Chaworth Musters describe en su viaje por la Patagonia "cuando la hembras están flacas se las mata para sacarle la piel, abandonando la carne; y cosiendo estas pieles, los indios hacen mantas que venden en las colonias" (Musters 1997 [1871]:156).
Las excavaciones y análisis de distintos conjuntos de vertebrados en el registro arqueológico muestran la aparición recurrente de ciertos elementos y restos óseos perteneciente a rheidos (Belardi 1999; Caviglia y Figuerero Torres 1976; Cruz y Elkin 2003; Fernández 2000; Fernández et al. 2001; Tonni y Laza 1980; Tambussi y Tonni 1985; Miotti y Salemme 1999; Miotti y Tonni 1996; Miotti et al. 1999; Neme y Gil 2002; Neme et al. 1998, entre otros). Sin embargo, la interpretación de las partes representadas ha estado limitada por la falta de estudios actualísticos que mejoren las predicciones u observaciones.
LOS ÍNDICES DE UTILIDAD ECONÓMICA
En cualquier partida de caza un dilema que enfrentan sus integrantes es el tamaño de los animales y la cantidad de presas capturadas. Si son demasiado grandes, deben decidir si transportarlos enteros o despostados. En este segundo caso también deben decidir qué descartar en el sitio de caza. La toma de decisiones estará afectada por numerosas variables a tener en cuenta, tales como comodidad del transporte, necesidad de materias primas (tendones, huesos, plumas, etc.), número de personas participando en la cacería y el consumo inmediato o futuro de cada parte. Tales decisiones conductuales llevarán a generar patrones arqueofaunísticos diferenciales de representación de partes esqueletales (Binford 1978). Ante distintas estrategias se esperan diferencias en la frecuencias de las partes esqueletales.
Los trabajos de Binford (1978) fueron pioneros en la creación de un marco de referencia para discutir uno de los aspectos de la toma de decisiones, a través de la elaboración de los índices de utilidad económica para diferentes unidades anatómicas en oveja doméstica (Ovis aries) y caribú (Rangifer tarandus). Estos índices fueron diseñados para predecir la estrategia durante cacerías, transporte, o técnicas de almacenaje de alimento. El principal supuesto del uso de estos índices de utilidad para interpretar conjuntos de fauna prehistórica es que los huesos asociados a la carne siempre serán transportados o abandonados como una unidad. Por lo tanto, la presencia de huesos particulares en un sitio residencial es interpretada como reflejo de las partes anatómicas asociadas que fueron transportadas hacia el sitio (Metcalfe y Jones 1988). Los índices están basados sobre cantidades de carne, médula ósea y grasa ósea y estos representan medidas cuantitativas de utilidad del alimento.
Estos trabajos pioneros de Binford (1978) marcaron una línea de investigación seguida por distintos autores quienes han generado índices de utilidad económica para otros taxones. Es así que se cuenta con los trabajos sobre león marino (Zalophus californianus) (Savelle et al. 1996), foca (Phoca hispida) (Diab 1998) y de cetáceos (Phocoena phocoena) (Savelle y Friesen 1996). Para ungulados se conocen los mencionados trabajos de Binford (1978) en caribú (Rangifer tarandus) y oveja doméstica (Ovis aries), los cuales son reexaminados y simplificados por (Metcalfe y Jones 1988), en llama (Lama glama) (Mengoni Goñalons 1991), en guanaco (Lama guanicoe) (Borrero 1990), en caballo (Equus caballus) (Outram y Rowley-Conwy 1998), y en huemul (Hippocamelus bisulcus) (Belardi y Gómez Otero 1998), entre otros.
Como un aporte a esa línea de investigación, e intentando incorporar a las aves, en este trabajo se calculan los índices de rendimiento económico para Rheidae (Rhea americana y Pterocnemia pennata). En este sentido, se realizaron los estudios de utilidad económica propuestos por Binford (1978), calculándose el "Índice de Utilidad General Modificado" (Modified General Utility Index o MGUI) y los índices que lo componen: "Índice de Utilidad de Carne" (Meat Utility Index o MUI), "Índice Medular" (Marrow Index o MI), "Índice General de Grasa" (White-Grease Index o WGI) y el "Índice de Utilidad General" (General Utility Index o GUI).
MATERIALES Y MÉTODOS
Los datos aquí presentados se obtuvieron de una hembra, subadulto de Pterocnemia pennata capturada en un ambiente natural del sur mendocino correspondiente fitogeográficamente a la Provincia del Monte (Cabrera 1976). Fue atrapada en el mes de abril de 2004 por cazadores furtivos. Se encontraba en buen estado nutricional con gran cantidad de la grasa acumulada. La presa fue inmediatamente introducida en un freezer, comenzando con su faenamiento un día después de la captura.
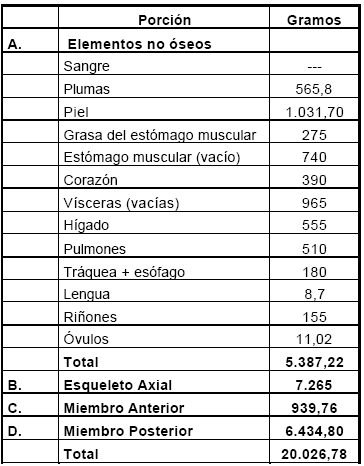
Primero se extrajeron las plumas de las alas, luego las del resto del cuerpo y finalmente se separó la piel del animal que se desprendió con facilidad dado que el cuero aún estaba relativamente congelado. Luego se evisceró el animal, pesando cada uno de los órganos internos del tórax y de la cavidad abdominal (Tabla 1).
Tabla 1. Peso para partes sin hueso y de porciones completas.
La segunda etapa consistió en el desposte primario, dividiendo la carcasa en unidades menores. En ningún caso el desposte implicó roturas de huesos, ya que durante la segmentación inicial del animal se siguieron las articulaciones. Los huesos largos fueron divididos en porciones proximales y distales. El propósito de esta subdivisión es generar información compatible con distintas formas de trozamiento primario donde no necesariamente se desposta cada hueso como una unidad (Binford 1978; Metcalfe y Jones 1988).
Los tejidos comestibles fueron extraídos del hueso separando la carne, la grasa y los tendones, siguiendo el método utilizado por Outram y Rowley-Conwy (1998). Este método difiere del empleado por Binford (1978) quien no separa tejidos comestibles de los tejidos conectivos. El peso de los tejidos comestibles fue usado para calcular los distintos índices de utilidad económica: el de utilidad de carne (MUI), el medular (MI) y el de grasa (WGI) los que, a su vez, permiten derivar el índice de utilidad general (GUI) y el índice de utilidad general modificado (MGUI). Se calculó el peso de las distintas partes con una precisión de 0,1 g. Para medir el peso total de la carne de cada parte anatómica se siguió la metodología de Binford (1978). Esto implicó la división de la carcasa en una serie de unidades que incluyen las distintas partes anatómicas.
Los huesos largos fueron seccionados en el punto medio de sus diáfisis y para extraer la médula se utilizó una variedad de implementos de metal y sometiendo al hueso a hervor. La remoción fue muy dificultosa, especialmente para los casos del fémur y de los huesos de los miembros anteriores, debido a la presencia de gran cantidad de hueso trabecular en la cavidad medular en el primero y por la angosta cavidad medular, en los segundos. Este problema fue resuelto pesando los huesos una vez extraída la médula de forma mecánica y luego hirviéndolos por el lapso de 4 horas. Una vez finalizado este proceso se pesaron nuevamente los huesos y se tomaron las diferencias, dando de esta forma el valor de médula total de cada hueso.
Para la realización de este índice, se utilizó solamente los huesos de las extremidades anteriores y posteriores. Su cálculo toma en cuenta el contenido medular y la grasa que los tejidos óseos contienen en su interior. A diferencia de Binford (1978), en este trabajo por cuestiones metodológicas no se consideró separadamente los distintos componentes de la cavidad medular sino que se los incluyó como una única fuente de energía. De esta forma, este índice incluye todo aquel contenido que se ha podido obtener de la cavidad medular y de los componentes grasos que se encuentran en los tejidos óseos.
El método de extracción del tejido graso fue similar al empleado para la carne, separándolo de forma manual de la carne con cuchillos y bisturí. Una significativa cantidad de tejido estaba adherido a la piel y para que su extracción fuera más sencilla se mantuvo a esta porción a bajas temperaturas.
RESULTADOS
El peso del animal vivo (completo con vísceras y plumas) fue de 23,410 kg y el de la carcasa eviscerada y sin plumas de 14,785 kg. Los valores obtenidos de plumas para las alas son de 356,5 g y para el resto del cuerpo de 208,3 g.
Índice de carne (MUI)
Para calcular la cantidad de carne de cada unidad anatómica, las unidades fueron divididas en porciones y los distintos tejidos fueron separados de forma manual, obteniendo los valores de carne de cada unidad. Para el caso de los huesos largos, este proceso se realizó separando la porción proximal de la distal. Los valores resultantes son presentados en la Tabla 2 la cual muestra los valores brutos en gramos de las distintas unidades anatómicas y los índices estandarizados de los pesos de carne para Rheidae (MUI).
Tabla 2. Peso total, peso de la carne e índice de carne (MUI).
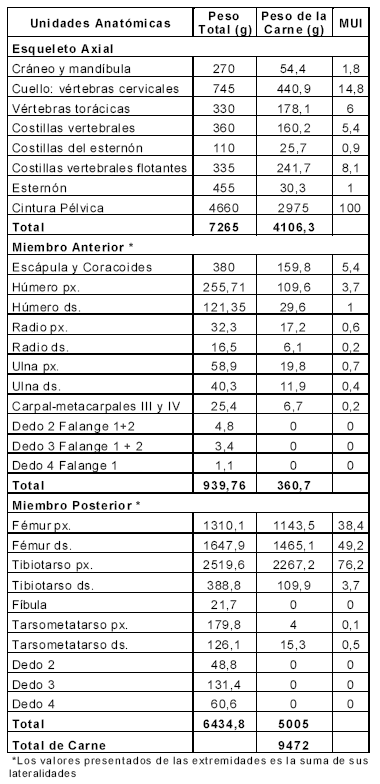
Considerando la propuesta de Binford (1978) la estandarización fue realizada expresando cada valor como un porcentaje del valor más alto registrado. En el caso de este estudio ese valor corresponde a la cintura pélvica y es de 2.975 g. En la Figura 1 se presentan los valores de carne estandarizados, se observa allí que la mayor proporción de carne se encuentra distribuida en la cintura pélvica. Siguiendo en importancia se encuentran el fémur y el tibiotarso en el miembro posterior, las vértebras y las costillas.
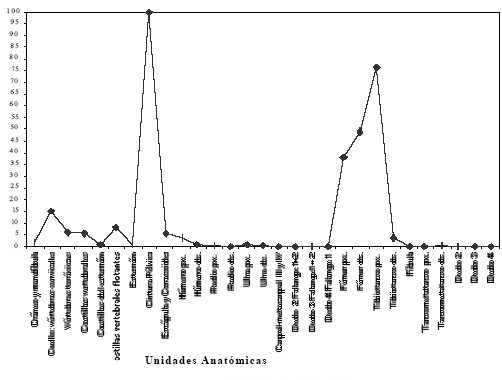
Figura 1. Valores estandarizados de carne (MUI).
Por último, es importante destacar que durante el proceso de descongelado, la carne pierde una cantidad mínima de agua, lo que debe haber alterado el peso original. De todos modos, se considera que las proporciones de las distintas unidades anatómicas se han mantenido constantes.
Índice de médula (MI)
Además de la carne se evaluó el rendimiento económico de la médula, ya que ésta es importante para los grupos humanos al proporcionar nutrientes que no son obtenidos durante el consumo de la carne (Jones y Metcalfe 1988). La proporción de médula en este grupo de animales es muy importante y se encuentra distribuida en diferentes estados físicos dentro de los huesos. Puede ser compacta en algún hueso (fémur y tibiotarso) como así también se puede encontrar en ciertos casos en forma líquida, sobre todo en el tarsometatarso, cuando el animal cazado ha estado corriendo.
En la Tabla 3 se muestran los valores de médula absolutos y estandarizados del contenido de médula de las distintas unidades anatómicas. El valor más alto corresponde a la porción proximal del tibiotarso, siguiendo en importancia la porción distal del mismo hueso. Presentan valores medios las porciones proximales y distales del fémur, y ambas porciones del tarsometatarso. Con valores menos del 10% se encuentran húmero, fíbula y falanges del miembro posterior. La Figura 2 muestra los valores del peso medular y del peso de médula total.
Tabla 3. Peso de la médula e índice medular (MI).
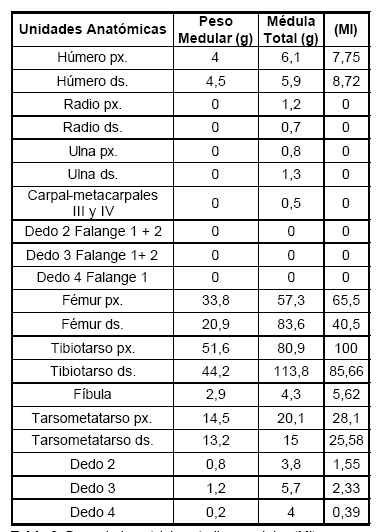
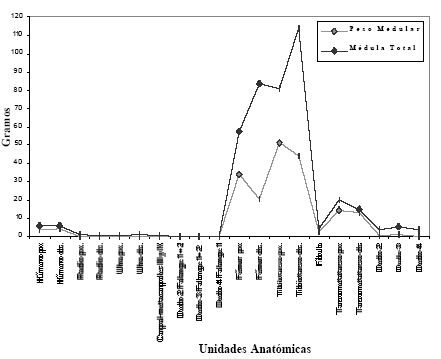
Figura 2. Cantidad de Médula.
Índice de grasa (WGI)
Los valores de grasa que se presentan en este trabajo corresponden a todo aquel tejido graso que se encuentra en el cuerpo del animal. Como se mencionó anteriormente, la proporción de este recurso puede variar de un mes a otro, por lo que el momento de su captura es muy importante. Para el cálculo de los valores de grasa Binford (1978) utiliza sólo aquella grasa contenida dentro de los huesos. Este autor incluye los tejidos grasos del cuerpo junto al peso de la carne. A diferencia de lo realizado por Binford (1978), para este trabajo se tomaron los valores del tejido graso, a partir de la grasa contenida entre los músculos y debajo de la piel.
En la Tabla 4 se presentan los valores absolutos en gramos para cada una de las unidades anatómicas, y su valor estandarizado a partir del valor más alto que en este caso corresponde a la cintura pélvica. Se puede ver que más del 70% de la grasa se encuentra en el esqueleto axial especialmente en la cintura pélvica, seguida ésta por el esternón (Figura 3).
Tabla 4. Peso de la grasa e índice de grasa (WGI).
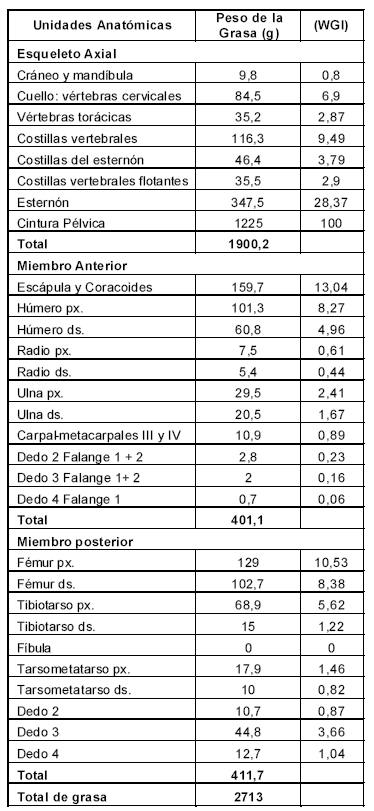

Figura 3. Valores Estandarizados de Grasa (WGI).
Índice de utilidad general (GUI)
Binford (1978) combinó los valores de carne, médula y de grasa ósea para crear un Índice de Utilidad General (GUI), el cual ha sido simplificado por Metcalfe y Jones (1988). Estos autores presentan un método de clasificación de la utilidad económica de partes anatómicas que es de menor complejidad que la realizada por Binford (1978). Ellos crean un índice idéntico llamado FUI (Food Utility Index) el cual muestra escalas simples de variación en la cantidad de carne, médula y grasa ósea asociada con diferentes partes anatómicas. En realidad, el índice de utilidad de carne de (Binford 1978) basado en la sustracción del peso del hueso seco del peso total de la parte anatómica, no corresponde a un índice de carne sino a un índice de todos los tejidos que se encuentran asociados a esa parte anatómica, incluyendo médula y grasa (Metcalfe y Jones 1988).
Los índices de carne, médula y grasa calculados aquí, reflejan directamente cantidades absolutas de carne, médula y grasa. Los resultados de este proceso son mostrados en la Tabla 5 y el GUI obtenido es la suma del peso de la carne, médula y grasa que se obtuvo por separado de la misma unidad anatómica (Figura 4).
Tabla 5. Índice de Utilidad General, basado en la suma del peso de la carne. (Tabla 2), peso de la médula (Tabla 3) y peso de la grasa (Tabla 4).
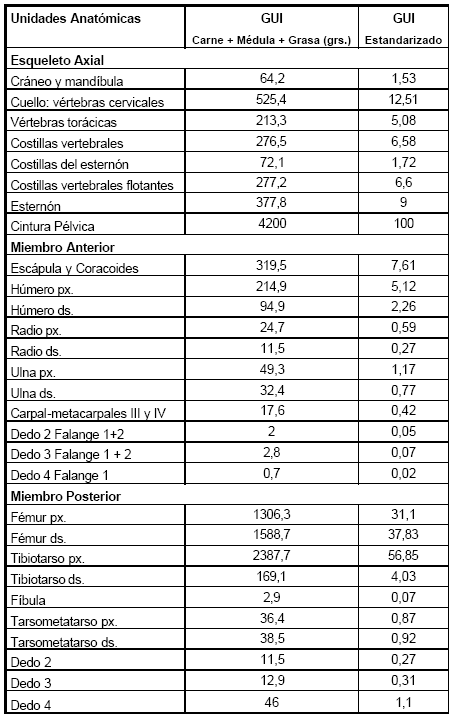
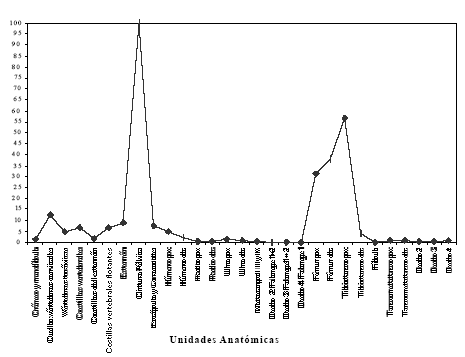
Figura 4. Valor estandarizado del Índice General de Utilidad (GUI).
De este cálculo se observa que los valores más altos se encuentran en la cintura pélvica, teniendo una distribución homogénea de carne y grasa. Un punto a destacar en el miembro posterior, es que la suma de los valores del fémur y tibiotarso son mayores al que se encuentran en la pelvis, y poseen una distribución heterogénea en relación a las proporciones de carne y grasa. De los valores obtenidos se resalta que la mayor proporción de tejidos comestibles se encuentran en el esqueleto axial sumando todos los huesos que componen esa zona.
Índice de utilidad general modificado (MGUI)
Binford (1978) observó que los Nunamiut cazan grandes animales y transportan hacia otro lugar partes de ellos, las cuales incluyen tanto porciones de bajo como de alto rendimiento según la escala de GUIs (Índice de Utilidad General Estandarizado). Además, demostró que el transporte de elementos de baja utilidad está relacionado a su proximidad anatómica con partes de valores de rendimiento alto. Binford (1978) denominó a estos elementos riders y notó que su efecto es importante fundamentalmente en los miembros apendiculares (ver también O´Connell et al. 1990; Outram y Rowley-Conwy 1998). Binford (1978) compensó este problema midiendo los valores de las porciones proximales y distales por separado y haciendo un promedio entre las porciones adyacentes de los miembros. Como resultado, cuando una parte de baja utilidad está anatómicamente adyacente a una parte que es comparativamente de alto rendimiento, entonces sus valores se incrementan. Este procedimiento sienta las bases del MGUI (Índice General de Utilidad Modificado) (Binford 1978), el cual es seguido en este trabajo.
Los valores para el MGUI fueron derivados del "Índice de Utilidad General" estandarizado siguiendo el procedimiento propuesto por Metcalfe y Jones (1988: nota 3), ya que las explicaciones de Binford (1978) son poco claras (Ecuación 1).
Ecuación 1
A diferencia de otros trabajos esta corrección se realizó en el miembro anterior desde el húmero y no desde la escápula y coracoides, debido a que en las aves estas porciones se encuentran insertas en el esqueleto axial. Cuando se realizó el trozamiento primario la parte más sobresaliente era el húmero, lo que sugiere que su desposte se realiza en forma separada de la escápula y coracoides. Los resultados son presentados en la Tabla 6 y Figura 5.
Tabla 6. Índice de Utilidad General Modificado.
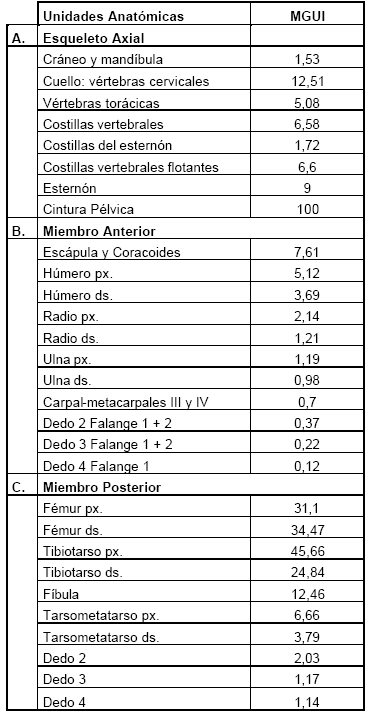
Figura 5. Valor estandarizado del Índice General de Utilidad Modificado (MGUI).
Al comparar los valores MGUI con GUI se notan algunas tendencias. Es así como se nota un incremento en los elementos del miembro posterior, específicamente tibiotarso y tarsometatarso. El MGUI disminuye las diferencias existentes entre la porción distal y la proximal del tibiotarso y aumenta los valores del tarsometatarso. En el miembro anterior no se observan diferencias entre el GUI y MGUI ya que posee valores bajos en todos sus elementos.
Índices de rendimiento económico y densidad mineral ósea
Numerosos autores han señalado los problemas de equifinalidad en la formación de conjuntos óseos, relacionados a la alta correlación existente entre los valores de rendimiento económico y los de densidad mineral ósea (Lyman 1994). Por este motivo se utilizaron los valores de densidad obtenidos para Pterocnemia pennata (Fernández et al. 2001; Cruz y Elkin 2003) para ver el grado de correlación con los valores de rendimiento obtenidos en el presente trabajo.
Para hacer comparable los valores de rendimiento económico obtenido dividiendo los huesos largos en proximal y distal con los datos de densidad mineral ósea, que dividen a los huesos largos en proximal, diáfisis y distal, se promedió el valor de densidad de la porción proximal con el valor de la diáfisis y de igual forma la parte distal con la diáfisis. El valor obtenido fue correlacionado con los valores de GUI. De esta forma, el valor de fémur proximal que se utilizó (0,77) es el promedio de los valores de fémur proximal (0,34) con el de la diáfisis (1,20). Este mismo cálculo se realizó para tibiotarso, cráneo mandíbula, vértebras cervicales, escápula y coracoides, húmero, tarsometatarso y fíbula. Siguiendo las sugerencias de (Lyman 1994), se empleó el test estadístico Spearman a partir del software Systat 11 para calcular la correlación entre el GUI y la densidad mineral ósea. La misma es de 0,354 y los resultados son mostrados en la Figura 6. Hay que considerar que aunque el grado de correlación no es fuerte, tampoco es despreciable por lo que señala un cierto grado de equifinalidad entre las partes de mayor rendimiento y las de mayor densidad mineral.
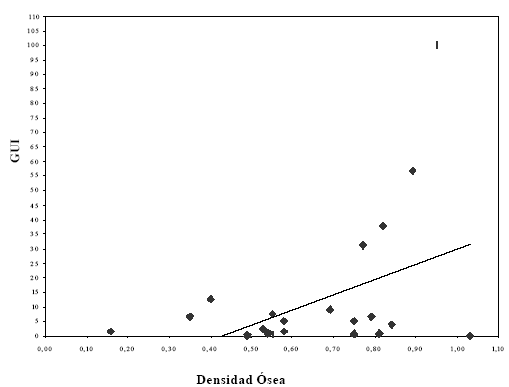
Figura 6. Relación entre GUI y densidad mineral ósea.
DISCUSIÓN
El rendimiento de carne y grasa tanto de Rhea americana y Pterocnemia pennata es similar (Sales et al. 1996), por lo que se considera que los índices pueden ser utilizados al nivel taxonómico de familia (Rheidae). Si bien para el cálculo de los índices se utilizó este ejemplar de Pterocnemia pennata, las proporciones en las que se hallan distribuidas las partes anatómicas en el individuo son muy similares a la de Rhea americana.
Los resultados de este trabajo han mostrado diferencias importantes en cuanto a la distribución de carne, grasa y médula en la anatomía de los rheidos. El modelo implementado originalmente por Binford (1978) para oveja doméstica (Ovis aries) y caribú (Rangifer tarandus) y posteriormente usado en otras especies, permite generar expectativas de hallazgo diferencial en contextos con distinta funcionalidad. Este modelo, predice que las partes de mayor rendimiento serían transportadas a los campamentos base donde son procesadas y consumidas, mientras que por el contrario, aquellas partes de bajo rendimiento permanecen en los lugares de matanza y procesamiento primario. Algunas de estas expectativas se ven modificadas por el traslado en forma conjunta de unidades de bajo rendimiento junto a unidades de alto rendimiento económico, para lo cual el autor desarrolló el índice de utilidad modificado. En este caso unidades de moderado y bajo rendimiento (tarsometatarso) pueden estar siendo transportadas en forma articulada junto con unidades de mayor rendimiento (tibiotarso), aumentando de este modo los valores relativos de aquellos huesos que se pueden encontrar en un sitio arqueológico.
En general, la disposición de los tres recursos principales contenidos en los rheidos (carne, grasa y médula) muestra que su distribución en el cuerpo es heterogénea. En este sentido, mientras que los valores más altos de rendimiento de carne se encuentran en las unidades anatómicas que forman el miembro posterior (fémur y tibiotarso) y así también la cintura pélvica presenta valores altos. La mayor proporción de grasa se ubica en la cintura pélvica y esternón y los de médula se encuentran en el tibiotarso, fémur y tarsometatarso.
Los resultados de este trabajo han mostrado que un aspecto importante a tener en cuenta es la gran cantidad de grasa que presenta este taxón. A excepción de los dasipódidos, el resto de los animales que habitan el sur de Mendoza, especialmente el guanaco, son muy magros. De allí que su captura podría haber estado orientada a la obtención de este recurso vital para la fisiología humana (Speth 1983, 1992).
Los valores de médula obtenidos muestran que los principales reservorios de este recurso se encuentran en el tibiotarso, que cuenta con la mitad del valor total de la médula de todo el animal. Es importante destacar la diferencia que existe entre la médula extraída del tibiotarso y la del tarsometatarso, ya que en esta última se encuentra en un estado mucho más líquido que la médula del resto del cuerpo. Según los puesteros1 de áreas rurales del sur mendocino, que alguna vez participaron en la cacería de ñandú o choique con métodos tradicionales de captura, cuentan que una vez cazado el animal, antes de extraer sus plumas ellos fracturan el tarsometatarso consumiendo la porción de médula contenida en él, en forma líquida, sin proceso de cocción previo. Por lo tanto, es importante considerar que si bien los mayores índices de médula de los huesos se encuentran en el tibiotarso, la extracción inmediata de la médula es mucho más sencilla en el tarsometatarso. Dada esta característica, podría esperarse la presencia de dichos huesos fracturados, asociados a los lugares de captura del animal, siempre y cuando estos no estén funcionando como instrumentos óseos. Es importante tener en cuenta esta última observación, dado que estos huesos se cuentan entre los más densos del esqueleto (Cruz y Elkin 2003) y por lo tanto es esperable que sean conservados para otro tipo de usos (Miotti, comunicación personal 2002).
La incorporación de los dos valores de médula ósea (peso medular y médula total) puede ser un índice muy útil para abordar temas de rendimiento en aquellos grupos que han incorporado la tecnología cerámica. En estos casos el aprovechamiento de la médula puede ser mayor que en aquellos grupos cazadores- recolectores que no han incorporado esta tecnología y que por lo tanto no pueden hervir los huesos, aprovechando parcialmente los contenidos de la misma. Sin embargo, es necesario tener en cuenta que, como observó Binford (1978) algunos grupos etnográficos de cazadores-recolectores acceden a los contenidos de médula calentando directamente el hueso al fuego, sin utilizar tecnología cerámica: "He normally moves to the edge of a hearth where a fire has been kindled and begins the processing of the bone. Processing consists of placing the bone or articulated leg section, if unskinned, along the margins of the fire where the hair is singed and the bone is warmed" (Binford 1978:152). Las mayores diferencias obtenidas entre los valores de médula previos y posteriores al procesamiento por hervido ocurren en los huesos con mayor tejido trabecular (huesos de las extremidades), donde se observa que su valor de médula ha aumentado considerablemente (Tabla 3, valor de médula total).
En cuanto a los huesos de bajo valor de rendimiento económico como las falanges, donde las cantidades de carne son nulas, su valor está dado por la cantidad de tendones y grasa. Para el caso del miembro anterior ocurre algo parecido, ya que presenta proporciones muy bajas de carne pero, por el contrario, presenta altos valores de grasa y tendones en sus falanges. Por último, el aprovechamiento de los huesos puede haber sido otro incentivo para la caza de ñandú. Es necesario considerar que algunos de los huesos del esqueleto de estas aves pudieron haber sido transportados no sólo por su utilidad económica en términos de alimento, sino como materia prima para la confección de instrumentos y/o adornos. En este sentido, en sitios de Patagonia se han encontrado agujas o punzones confeccionados a partir de fíbulas de rheidos, (Miotti comunicación personal 2002). Así mismo Anderson (1983) menciona también la utilización de huesos de moa para la confección de anzuelos y arpones de pesca en Nueva Zelanda. De esta forma, estas partes anatómicas podrían entrar al sitio independientemente de su rendimiento económico en términos de carne y/o grasa.
Expectativas generadas
Sobre la base de los resultados obtenidos exclusivamente de los índices de rendimiento económico elaborados para Rheidae, es esperable la presencia de las partes correspondientes a la cintura pélvica y fémur en los lugares de consumo dado los altos valores de rendimiento (especialmente en carne) asociados a éstas. Por el contrario, es esperable que se encuentren las vértebras cervicales, el miembro anterior y las falanges del miembro posterior en sitios de caza que se encuentren lejos de los lugares de consumo y/o con problemas de accesibilidad.
Si los grupos de cazadores están explotando a estos animales por su contenido en grasa, entonces se esperaría en los sitios de consumo encontrar las partes del esqueleto que corresponden al esternón, cintura pélvica y escápula-coracoides (Figura 3). Si estas partes son las que están presentes en el campamento, entonces también sería posible sostener que el momento de captura elegido haya sido el otoño. Por el contrario, la captura de rheidos para utilizar sólo la médula, tendones y piel es esperable en otras estaciones del año. En este caso las partes anatómicas que deberían estar presentes en los sitios son los miembros posteriores, incluyendo las falanges.
En el caso del aprovechamiento de la médula es esperable encontrar huesos fracturados, quemados y/o hervidos; correspondiendo a tibiotarso, tarsometatarso y fémur y, en menor proporción, húmero, radio y ulna.
Por último la correlación entre densidad mineral ósea y GUI alertan sobre la posibilidad de que existan problemas de equifinalidad en los conjuntos arqueológicos recuperados, donde no sólo pueden estar jugando decisiones de los grupos humanos, sino también problemas tafonómicos que pueden oscurecer la forma de entender por qué se registra un patrón arqueofaunístico determinado.
CONCLUSIÓN
Tanto las crónicas de viajeros como los trabajos arqueofaunísticos han mostrado la importancia que los rheidos han tenido en la subsistencia prehispánica (Belardi 1999; Caviglia y Figuerero Torres 1976; Cruz y Elkin 2003; Fernández 2000; Fernández et al. 2001; Miotti y Salemme 1999; Miotti y Tonni 1996; Miotti et al. 1999; Tambussi y Tonni 1985; Tonni y Laza 1980). Sin embargo, no existían estudios previos de rendimiento económico para estas aves, lo que debilita las interpretaciones emanadas de los conjuntos arqueofaunísticos analizados en el extremo sur de Sudamérica. Aquí se detallan los resultados del rendimiento de la utilidad económica de partes esqueletales sobre el peso de los tejidos aprovechables, presentando los valores absolutos y estandarizados, modificados y no modificados. Se siguió la metodología propuesta por Binford (1978) para la elaboración de los índices de rendimiento económico. Se generaron algunas expectativas de registro arqueofaunístico a partir del rendimiento económico de las diferentes partes anatómicas de los rheidos y sobre la base de la relatos informales de pobladores locales se esbozaron algunas cuestiones sobre la caza y costumbres del consumo de estas aves corredoras.
Los resultados presentados en este trabajo muestran que las decisiones de los cazadores pudieron haber estado afectadas por el momento elegido para su captura ya que la estacionalidad modificaría los valores de rendimiento económico de estas aves. La diversa cantidad de recursos que puede brindar este animal (piel, plumas, carne, grasa, tendones y huesos) abre un abanico de expectativas en cuanto a los conjuntos esperados en sitios arqueológicos. Los mismos no sólo dependen de la distancia existente entre los lugares de captura y los campamentos base, sino también depende del tipo de recurso del animal que haya sido explotado.
Los datos obtenidos muestran que las unidades anatómicas de mayor rendimiento en carne se encuentran en los miembros posteriores, mientras que su proporción en relación con la grasa es muy baja. Por el contrario, en el esqueleto axial se encuentran los máximos valores de grasa mostrando una fuerte correspondencia con los valores de carne. Es necesario tener en cuenta que las proporciones de grasa contenida en el esqueleto axial sufren fuertes variaciones según la época de captura del animal.
Por último, el cruce de los datos de rendimiento económico con los de densidad mineral ósea muestra una correlación media, lo que implica un problema potencial de equifinalidad en los conjuntos arqueológicos, con una tendencia positiva entre rendimiento y densidad ósea.
1 Puesteros: Denominación que se les da a los habitantes de las zonas rurales del sur de Mendoza, son transhumantes. Aprovechan la época de invernada para realizar boleadas (Agüero Blanch 1971).
Agradecimientos
Este estudio se pudo realizar gracias a la colaboración de la Dirección de Recursos Naturales del Gobierno de Mendoza, Delegación San Rafael, quienes brindaron el espécimen analizado de Pterocnemia pennata. A Gustavo Neme, Adolfo Gil, Luis A. Borrero y Víctor Durán por su asesoramiento y lecturas críticas del manuscrito. Las sugerencias y correcciones de los evaluadores ayudaron significativamente a mejorar este trabajo. Este trabajo fue posible realizar con la ayuda de la Fundación Antorchas (Proyecto N° 14116-182 y N° 14116-116) y FONCYT (PICT 04-14695). La metodología seguida fue expuesta en las VI Jornadas de Jóvenes Investigadores en Ciencias Antropológicas desarrollado en la Ciudad de Buenos Aires y en el XV Congreso Nacional de Arqueología Argentina, llevado a cabo en la ciudad de Río Cuarto. También quiero agradecer la colaboración de Miriam Ayala y Adrián Atencio quienes ayudaron en los trabajos de laboratorio y por último a J. B. Sterli por sus minuciosas correcciones.
REFERENCIAS CITADAS
1. Agüero Blanch, V. 1969 Supervivencia Aborigen en la Alimentación, en el departamento de Malargüe (Mendoza). Anales de Arqueología y Etnología 22: 93-100. [ Links ]
2. Agüero Blanch, V. 1971 Malargüe Pueblo Trashumante. Anales de Arqueología y Etnología 24-25: 209-223. [ Links ]
3. Anderson, A. 1983 When all the moa-ovens grew cold: nine centuries of changing forture for the Southern Maori. Heritage Books, Dunedin Otago. [ Links ]
4. Belardi, J. B. 1999 Hay choiques en la terraza. Información tafonómica y primeras implicaciones arqueofaunísticas para Patagonia. Arqueología 9: 163-187. [ Links ]
5. Belardi, J. B. y J. Gómez Otero 1998 Anatomía económica del huemul (Hippocamelus bisulcus): una contribución a la interpretación de las evidencias arqueológicas de su aprovechamiento en patagonia. Anales del Instituto de la Patagonia 26: 195-207. [ Links ]
6. Binford, L. 1978 Nunamiut Ethnoarchaeology. Academic Press, Nueva York. [ Links ]
7. Bond Head, F. 1997 Las Pampas y Los Andes. El Elefante Blanco, Buenos Aires. [ Links ]
8. Borrero, L. A. 1990 Fuego - Patagonia bone assemblages and the problem of comunal guanaco hunting. En Hunters of the recent past, editado por L. B. Davis y B. O. K. Reeves, pp. 373-406. Unwin Hyman, Londres. [ Links ]
9. Cabrera, A. 1976 Regiones Fitogeográficas Argentinas. Enciclopedia Argentina de Agruicultura y Jardinería, Tomo II. Acme S.A.C.I, Buenos Aires. [ Links ]
10. Carman, R. 1973 De la Fauna Bonaerense. s/e, Buenos Aires. [ Links ]
11. Caviglia, S. y M. J. Figuerero Torres 1976 Materiales faunísticos de la cueva "La Buitrera" (Dpto. UER Aike, Prov. de Santa Cruz). Relaciones de la Sociedad Argentina Antropología (N.S.) 10: 315-319. [ Links ]
12. Claraz, J. 1988 Diario de viaje de exploración al Chubut 1865- 1866. Marymar, Buenos Aires. [ Links ]
13. Cruz, I. y D. Elkin 2003 Stuctural bone density of the lesser Rhea (Pterocnemia pennata) (Aves: Rheidae). Taphonomic and archaeological implications. Journal of Archaeological Science 30: 37-44. [ Links ]
14. Fernández, P. 2000 Rendido a tus pies: acerca de la composición anatómica de los conjuntos arqueofaunísticos con restos de Rheiformes de Pampa y Patagonia. En Desde el País de los Gigantes, Perspectivas arqueológicas en Patagonia, Tomo 2, pp. 572-586. Universidad Nacional de la Patagonia Austral, Río Gallegos. [ Links ]
15. Diab, M. 1998 Economic Utility of the Ringed Seal (Phoca hispida): Implications for Arctic Archaeology. Journal of Archaeological Science 25: 1-26. [ Links ]
16. Fernández, P., I. Cruz y D. Elkin 2001 Densidad mineral ósea de Pterocnemia pennata (Aves: Rheidae). Una herramienta para evaluar frecuencias anatómicas en sitios arqueológicos. Relaciones de la Sociedad Argentina de Antropología 26: 243-260. [ Links ]
17. Garriz, C., M. Urioste, S. Delarada, M. Della Croce, J. Isequilla y H. Albera 2003 Ñandúes Argentinos (Rhea americana). I - rendimiento de res y derivados de faena. Trabajo presentado en el 1° Congreso Latinoamericano sobre Conservación y Cría de Ñandues, Buenos Aires. MS. [ Links ]
18. Jones, K. y D. Metcalfe 1988 Bare bones archaeology: bone marrow index and efficiency. Journal of Archaeological Science 15: 415-423. [ Links ]
19. Kooyman, B. 1990 Moa Procurement: Communal or Individual Hunting? En Hunters of the recent past, editado por L. B. Davis y B. O. K. Reeves, pp. 327-351. Unwin Hyman, Londres. [ Links ]
20. Lyman, R. 1994 Vertebrate Taphonomy. Cambridge Manuals in Archaeology. Cambridge University Press, Cambridge. [ Links ]
21. Mengoni Goñalons, G. 1991 La llama y sus productos primarios. Arqueología 1: 179-196. [ Links ]
22. Metcalfe, D. y K. Jones 1988 A reconsideration of animal body-part utility index. American Antiquity 53(3): 486-504. [ Links ]
23. Miotti, L. y M. Salemme 1999 Biodiversity, Taxonomic Rechness and Specialists-Generalists during Late Pleistocene/Early Holocene Times in Pampa and Patagonia (Argentina, Southern South America). Quaternary International 53-54: 53-68. [ Links ]
24. Miotti, L. y E. Tonni 1996 Rheidos del sitio arqueológico AEP-1 de Piedra Museo y la meseta central de Santa Cruz y sus implicancias paleoambientales. Póster presentado en las 3ras. Jornadas de Arqueología de Patagonia, Bariloche. MS. [ Links ]
25. Miotti, L., M. Vázquez y D. Hermo 1999 Piedra Museo, un Yamnagoo pleistocénico de los colonizadores de la meseta de Santa Cruz. El estudio de la arqueofauna. Soplando en el Viento. Acta de las III Jornadas de Arqueología de la Patagonia, pp 113-136. Facultad de Humanidades, Universidad Nacional del Comahue e INALP, Neuquén. [ Links ]
26. Muñiz, F. 1916 El Ñandú o avestruz americano. Escritos Científicos seleccionados y ordenados por Domingo F. Sarmiento. Obras de Sarmiento XLII: 109-237. [ Links ]
27. Musters, G. 1997 [1871] Vida entre los Patagones. Un año de excursiones por tierras no frecuentadas desde el Estrecho de Magallanes hasta Río Negro. El Elefante Blanco, Buenos Aires. [ Links ]
28. Neme,G., A. Gil y V. Durán 1998 El Registro Arqueofaunístico del Alero Puesto Carrasco (Malargüe-Mendoza). Soplando en el Viento. Acta de las III Jornadas de Arqueología de la Patagonia, pp 491-514. Facultad de Humanidades, Universidad Nacional del Comahue e INALP, Neuquén. [ Links ]
29. Neme, G. y A. Gil 2002 La explotación faunística y la frecuencia de partes esqueletarias en el registro arqueológico del sur mendocino. En Entre Montañas y Desiertos: Arqueología del Sur de Mendoza, editado por A. F. Gil y G. A. Neme, pp. 101-118. Sociedad Argentina de Antropología, Buenos Aires. [ Links ]
30. O´Connell, J., K. Hawkes y N. Blurton Jones 1990 Reanalysis of Large Mammal Body Part Transport among the Hadza. Journal of Archaeological Science 17: 301-316. [ Links ]
31. Outram, A. y P. Rowley-Conwy 1998 Meat and Marrow Utility Indices for Horse (Equus). Journal of Archaeological Science 25: 839- 849. [ Links ]
32. San Martín, F. 1930 Neuquén. Biblioteca del Suboficial, Buenos Aires. [ Links ]
33. Sales, J., A. Manero y J. Navarro 1996 Carcass and fabrication yields of Lesser Rhea (Pterocnemia pennata). Report for National Institute of Agricultural Technology (INTA), Santa Cruz Experimental Station, Argentina. MS. [ Links ]
34. Savelle, J., T. Frisen y R. Lyman 1996 Derivation and Application of an Otariid Utility Index. Journal of Archaeological Science 23: 705-712. [ Links ]
35. Savelle, J. y T. Friesen 1996 An Odontocete (Cetacea) Meat Utility Index. Journal of Archaeological Science 23: 713-721. [ Links ]
36. Speth, J. 1983 Energy Source, Protein Metabolism, and Hunter-Gatherer Subsistence Strategies. Journal of Anthropological Archaeology 2: 1-31 [ Links ]
37. Speth, J. 1992 Protein selection and avoidance strategies of contemporary and ancestral foragers: unresolved issues. En Foraging Strategies and Natural Diet of Monkeys, Apes and Humans, editado por A. Whiten y E. M. Widdowson, pp. 105-110. Claredon Press, Oxford. [ Links ]
38. Tambussi, C. y E. Tonni 1985 Aves del sitio arqueológico Los Toldos, Cañadón de las Cuevas, Provincia de Santa Cruz (República Argentina). Ameghiniana 22(1-2): 69-74. [ Links ]
39. Tonni, E. y J. Laza 1980 Las aves de la fauna local de Paso Otero (Pleistoceno tardío) de la Provincia de Bs. As. Su significación ecológica; climática y zoogeográfica. Ameghiniana 17(4): 313-322. [ Links ]

