










Servicios Personalizados
Revista

Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkIntersecciones en antropología
versión On-line ISSN 1850-373X
Intersecciones antropol. n.8 Olavarría ene./dic. 2007
Análisis paleoambiental sobre la base del estudio malacológico del sitio Paso Otero 1 (región pampeana, Argentina)
Pamela G. Steffan
Pamela Steffan. CONICET, INCUAPA. Facultad de Ciencias Sociales (UNCPBA). Avda. del Valle 5737, B7400JWI Olavarría, Argentina. E-mail: psteffan@soc.unicen.edu.ar
Recibido 8 de Junio 2005. Aceptado 21 de Diciembre 2005
RESUMEN
Se infieren las condiciones paleoclimáticas y paleoambientales en Paso Otero 1 durante el Holoceno, particularmente desde los ca. 4800 AP a 2900 años AP. Paso Otero 1 está ubicado en el área del curso medio del río Quequén Grande (área Interserrana Bonaerense, región pampeana). El análisis de la composición de la fauna de gasterópodos comprende distintos aspectos: diversidad de especies representadas en el muestreo, medidas de abundancia relativa de individuos por especie y distribución de frecuencias por niveles muestreados. Los resultados indican una mayor humedad hacia los 4800 años AP y un ascenso en la temperatura hacia los 2900 años AP. Estos análisis permitieron evaluar la información paleoambiental de los gasterópodos fósiles y aportar datos acerca de las condiciones ambientales locales.
Palabras clave: Paleoambientes; Holoceno; Area Interserrana Bonaerense; Paso Otero 1.
ABSTRACT
Paleoenvironmental analysis of the malacological studies conducted at Paso Otero 1 Site (pampean region, Argentina). In this paper, paleoclimatic and paleoenvironmental conditions at Paso Otero 1 are inferred for the Holocene, particularly from ca. 4800 to 2900 BP. Paso Otero 1 is located in the area of the middle basin of the Quequén Grande river (Interserrana Bonaerense area, Pampean region). Analysis of the faunal composition of gastropod fauna included different aspects: diversity of represented species in the sample, measurements of relative abundance of individuals by species, and the distribution of frequencies for sampled levels. The results indicate greater humidity toward ca. 4800 BP and an increase in temperature toward 2900 BP. The analyses enable the environmental information of the fossil gastropods to be evaluated, contributing data on local environmental conditions.
Keywords: Paleoenvironment; Holocene; Interserrana Bonaerense Area; Paso Otero 1.
INTRODUCCIÓN
Los restos de gasterópodos ingresados al registro estratigráfico por depositación natural, han sido reconocidos como un apéndice relacionado con su presencia. Sin embargo, el potencial de información que brinda es mucho mayor debido a su interés biológico, ecológico intrínseco y para reconstruir el ambiente del pasado, tanto local como regional (Bobrowsky 1984). Asimismo, permitiría resolver problemas de las condiciones de depositación de los restos producto de la actividad humana y/o peculiaridades de los sitios arqueológicos.
Se considera que la presencia de gasterópodos en Paso Otero 1 (38° 34' LS y 58° 42' LO) requiere de una mayor atención. El objetivo del trabajo es evaluar la información obtenida a partir de los gasterópodos fósiles dulceacuícolas y terrestres obtenidos del muestreo realizado en la columna estratigráfica del perfil del sitio, previamente estudiado por Politis et al. (1991), Gutiérrez (2001, 2004), Johnson et al. (1997, 1998), Martínez (1999).
El presente análisis considera a la totalidad de la fauna malacológica, incluyendo la diversidad de especies representadas en el muestreo, medidas de abundancia relativa de individuos por especie y la distribución de frecuencias por niveles muestreados. Estas variables permitirán discutir aspectos relacionados con las condiciones de depositación, como así también de la evolución del paleoclima durante el Holoceno.
REFERENCIA DEL ÁREA
El sitio Paso Otero 1 está ubicado sobre la margen izquierda del curso medio del río Quequén Grande (Provincia de Buenos Aires, Argentina) (Politis et al. 1991). En el perfil estratigráfico del sitio se identificaron tres superficies de estabilización (SE) u horizontes A de suelos enterrados, -superficie de estabilización Media (SEM) y superficie de estabilización Superior (SES)- asociados a acumulaciones de huesos de guanaco (Lama guanicoe) y una tercera, denominada Inferior (SEI) (Martínez 1999). Los primeros estudios de la acumulación de una gran cantidad de huesos de guanaco (ca. 3800), concentrados en cuatro pilas, fue relacionada a eventos diacrónicos de ocupación humana relacionados a actividades específicas de caza y procesamiento inicial de guanaco (Lama guanicoe) (Gutiérrez 2004; Martínez 1999). A partir de un estudio geoarqueológico en la cuenca media del río Quequén Grande, Favier Dubois (2006) evaluó la situación estratigráfica y paleoambiental en Paso Otero 1 y consideró que la ocurrencia de las pilas óseas se vincula con la presencia de discordancias erosivas relacionadas a la acción de flujos turbulentos. Los estudios tafonómicos llevados a cabo por (Gutiérrez y Kaufmann 2007), sugieren un origen fluvial para dichas acumulaciones óseas, resultando en una mezcla de materiales de origen natural y antrópico.
Desde el punto de vista ambiental estos horizontes A representan condiciones ambientales húmedas muy localizadas, con gran cantidad de materia orgánica bajo condiciones pobremente drenadas (Holliday 1995). Estos períodos de estabilidad se habrían producido en planicies de inundación o charcas, en alternancia con períodos de aluviación, desarrollados en depósitos lacustres del Platense o Miembro Río Salado de la Formación Luján sensu Fidalgo et al. (1973) (Johnson et al. 1998; Martínez 1999) (Figura 1). Favier Dubois (2006: 112) reconoce cuatro unidades litoestratigráficas. En la base, Unidad I, representa facies fluviales a fluvio-lacustres arenosas y se corresponde con el Miembro Guerrero de la Formación Luján, sobre el cual se desarrolla el suelo Puesto Callejón Viejo. La Unidad II referible al Miembro Río Salado de la Formación Luján, presenta dos bancos de origen lacustre con claras evidencias de cambios en la profundidad del antiguo cuerpo de agua. El sector inferior consiste en un depósito de diatomeas, representa facies lacustres limosas. Entre ambos bancos, se desarrolló un suelo palustre, (SEM) formado por somerización del cuerpo de agua. El banco superior representa una diatomita masiva con facies más someras. En la parte superior del suelo palustre y la base de la unidad superior, se observa un contacto discordante con morfología de canal, con acción de un flujo erosivo. Con registro de facies muy finas de contenido bioclático que indica un nuevo crecimiento del cuerpo de agua. Por encima de la Unidad II se desarrolla un paleosuelo referible al Suelo Puesto Berrondo (SES) decapitado en variable grado por una evidente discordancia erosiva. La Unidad III, depósitos franco arenosos masivo intercalado con un total de seis paleosuelos de diferente desarrollo. La Unidad IV es un depósito pequeño que no presenta pedogénesis hacia el techo de la sección estratigráfica.
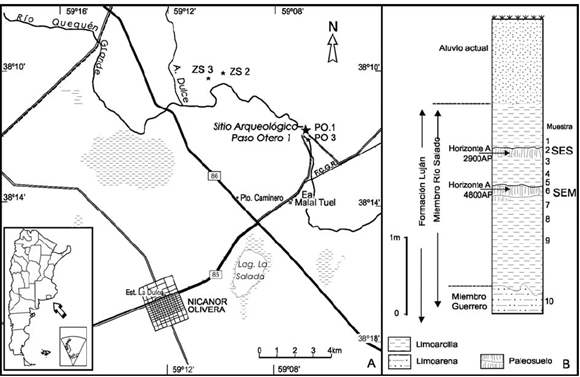
Figura 1 A: ubicación de Paso Otero 1. B: perfil estratigráfico de muestreo (modificada de Miquel y Steffan 2005).
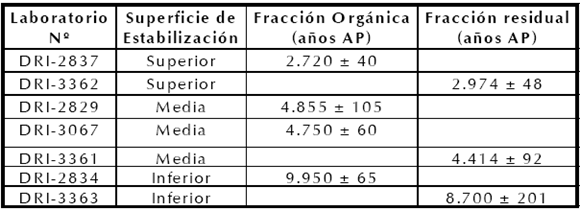
La cronología de los horizontes A se obtuvo a partir de fechados radiocarbónicos de sedimento orgánico provenientes de cada uno de los horizontes A de suelo enterrado (Johnson et al. 1998). Tomando el criterio de la edad más antigua, las edades corregidas de la superficie de estabilización Superior es de 2974 ± 48 años AP (DRI-3362), la superficie de estabilización Media es de 4855 ± 105 años AP (DRI-2837) y la superficie de estabilización Inferior con una edad de 9950 ± 65 años AP (DRI- 2831) (Johnson et al. 1998) (Tabla 1).
Tabla 1. Edades radiocarbónicas del sitio Paso Otero 1 (Johnson et al. 1998).
Los datos faunísticos y polínicos utilizado como proxy para la reconstrucción paleoambiental de la región pampeana sugieren diferencias entre las condiciones climáticas durante ca. 4800 AP y los ca. 2900 años AP en Paso Otero 1. Teniendo en cuenta la información obtenida desde los modelos regionales (i.e., Prado et al. 2001; Tonni 1992; Tonni et al. 1999), las condiciones húmedas y cálidas habrían prevalecido cuando la asociación ósea de la superficie de estabilización Media (ca. 4800 años AP) fue depositada en el sitio. Progresivamente, las condiciones fueron más frías y áridas durante la depositación de la asociación ósea en la superficie de estabilización Superior (ca. 2900 años AP). Con respecto a la temperatura, los datos proxy indican que los cambios en las temperaturas fueron suficientes como para producir la migración y reemplazo de especies animales y vegetales (Prieto 1996; Tonni 1992).
OBJETIVOS E HIPÓTESIS
En relación con la situación planteada previamente, en este trabajo se propone comprender las condiciones paleoclimáticas y específicamente, el paleoambiente en relación con horizontes A de suelos enterrados, -superficie de estabilización Media (SEM) y superficie de estabilización Superior (SES)- vinculados a acumulaciones de huesos de guanaco (Lama guanicoe). En este sentido, el análisis malacológico permitirá conocer las condiciones climáticas y ambientales en una escala temporal y espacial más acotada que los modelos climáticos regionales. Esto permitirá contrastar los resultados obtenidos por estudios previos respecto del ambiente de depositación mediante una línea de evidencia independiente.
La hipótesis de trabajo plantea que "las condiciones paleoambientales (temperatura y humedad) durante el desarrollo de ambos paleosuelos en Paso Otero 1 fueron similares". Así mismo, se espera observar si las condiciones cálidas y húmedas del Holoceno medio pudieron afectar a las especies de moluscos. La relación presencia - ausencia de las especies terrestres y de agua dulce a lo largo del perfil permitirá inferir la intensidad de las variaciones climáticas registradas durante la depositación de los sedimentos en el sitio Paso Otero 1.
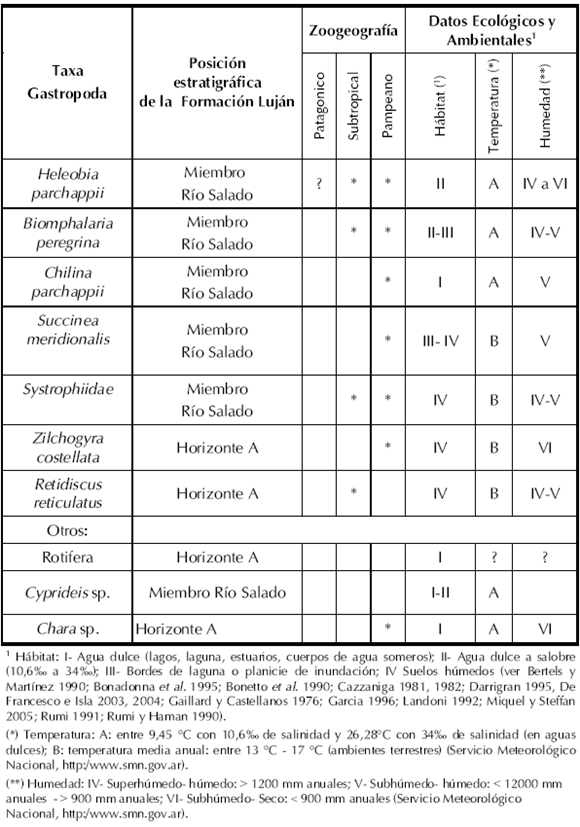
En las aproximaciones paleoambientales basadas en el uniformitarismo con gasterópodos, se parte de la premisa de que la ecología de los taxones fósiles y modernos no ha cambiado en el lapso temporal que se considera. Específicamente, en el tratamiento de la hipótesis se parte del conocimiento de la capacidad de resistencia de los organismos a sobrevivir en un ambiente sujeto a constantes cambios. No obstante, cada especie manifiesta capacidades de adaptación que son propias de las dinámicas de población particulares. Dichos supuestos, surgen de la información zoogeográfica proveniente de varios estudios realizados sobre la especie Biomphalaria peregrina -d'Orbigny 1835- (Rumi 1991; Rumi et al. 1997) y de Heleobia parchappii (D'Orbigny 1835) sobre la base de muestras fósiles y modernas (i.e., Aguirre y Farinati 2000; Castellanos y Miquel 1991; De Francesco e Isla 2003).
METODOLOGÍA
Los gasterópodos se recolectaron a partir de la extracción de muestras de sedimento del perfil estratigráfico, correspondiente a la pared oeste de la excavación del sitio Paso Otero 1. Las muestras fueron tomadas siguiendo la orientación y disposición de los estratos naturales. La cantidad promedio de sedimento por muestra fue de ca. 9000 cm3.
En el campo se dividió el muestreo en dos etapas. En la primera se realizó un muestreo en todo el perfil con el objeto de ser utilizado como modelo de referencia y observar la evolución de la diversidad de especies a través del tiempo. En el segundo muestreo, se puntualizó en las superficies de estabilización Superior y Media, asociadas a las acumulaciones óseas.
En el laboratorio se llevó a cabo la preparación de las muestras, determinación taxonómica de los gasterópodos, identificando los especímenes por especies según el conjunto de caracteres preservables en las conchillas al estado fósil (contorno, características de la espira, tamaño, forma de los anfractos, suturas y aberturas) que permiten asignarlos taxonómicamente y efectuar inferencias relacionadas con la clasificación adecuada. En la cuantificación de los individuos, se contabilizaron las conchillas enteras, el criterio seguido se debió a la necesidad de evitar conteos duplicados, de tal modo que cada individuo fuera ingresado sólo una vez en la tabla general. En el caso que la conchilla estuviera muy fragmentada, se recuperó el último anfracto como es el caso de la especie Heleobia parchappii (y otras de morfología similar) y la especie Biomphalaria peregrina y otras (se recuperaron las partes correspondientes al ombligo, típico en la estructura de la conchilla en dicha especie). Los materiales examinados fueron incorporados en la colección de la Facultad de Ciencias Sociales (Olavarría), bajo la sigla FCS-IN.
De acuerdo con la problemática planteada, para contrastar la hipótesis de trabajo se observaron una serie de valores de diversidad taxonómica y de abundancia relativa de especies. Estas medidas son utilizadas como un indicador de la productividad del ambiente en el cual se depositaron. Ninguno de estos métodos está libre de limitaciones, sin embargo, estas medidas se utilizaron para analizar la fauna malacológica en un área dada durante un intervalo de tiempo específico. La distribución de las frecuencias de individuos por especies en cada nivel estratigráfico arbitrario permite analizar su alternancia en relación con las variaciones climáticas de períodos húmedos o secos. Al mismo tiempo, se analizó el comportamiento de la relación presencia-ausencia de especies terrestres y de agua dulce representada en ambos muestreos, lo cual brinda una dimensión respecto de la intensidad de las variaciones climáticas registradas en el perfil estratigráfico del sitio Paso Otero 1.
En cuanto a la cronología del muestreo fue necesario realizar una serie de consideraciones respecto de la edad estimada para el mismo. Estratigráficamente, el muestreo general en el perfil del sitio Paso Otero 1 comprende la porción superior del Miembro Guerrero denominado por Prado et al. (1987) "verde superior" y abarca la totalidad del depósito asignable al Miembro Río Salado de la Formación Luján. En dicho Miembro, se formaron los paleosuelos reconocidos como superficies de estabilización Media y Superior (Johnson et al. 1997). Teniendo en cuenta las dataciones realizadas previamente para el límite entre ambos miembros (Carbonari et al. 1992; Johnson et al. 1998), se estima que la cronología comprendida por el muestreo se ubica entre 15.370 + 780 - 710 años AP a 10.840 ± 380 años AP como límite inferior y los 2720 ± 40 años AP según el fechado obtenido para la superficie de estabilización Superior. No obstante, se reconoce la necesidad de contar con fechados para establecer con mayor precisión el sector del perfil analizado.
ANÁLISIS DE LOS RESULTADOS
En el muestreo general del sitio Paso Otero 1 la malacofauna presenta una importante diversidad taxonómica representada por individuos de siete especies y de distintos ambientes, tanto de agua dulce como por especies terrestres (Figura 2, Tabla 2). La asociación de especies es el resultado de una serie de procesos tafonómicos actuantes de acumulación, fracturación y redepositación. La presencia de varias taxones de moluscos acuáticos, de ostrácodos adultos y juveniles, asociados con estructuras reproductivas de algas de la familia Characeae (Chara sp.) sobre las que viven los individuos de la especie Heleobia parchappii, indican una asociación de ambientes de aguas calmas y de depositación in situ de elementos autóctonos. Adicionalmente se agregaron abundantes conchillas fracturadas y con evidencias de abrasión y pulido en el periostraco indicando un transporte por alta energía. Esta dinámica de acumulación propia de la planicie de inundación ha dejado como resultado una importante abundancia de especies.

Figura 2. Diferentes especies de gasterópodos de agua dulce y terrestres: 1- Biomphalaria peregrina, 2- Heleobia parchappii, 3- Systrophiidae, 4- Succinea meridionalis, 5- Retidiscus reticulatus.
Tabla 2. Características de las especies malacológicas.
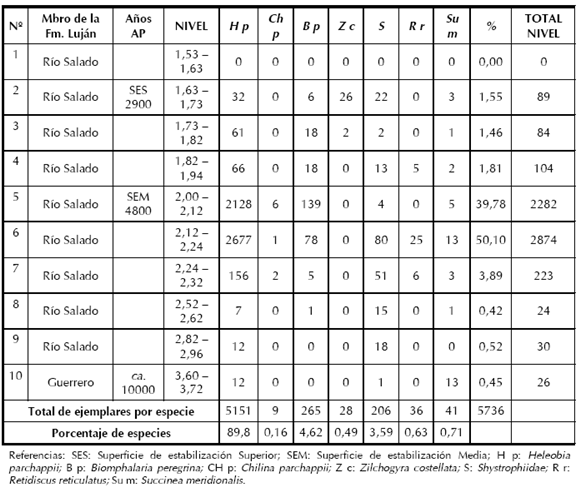
A lo largo del perfil del sitio Paso Otero 1, fue posible observar variaciones en la frecuencia de aparición de la malacofauna representada en el mismo. Estadísticamente, se pudo observar que los diez niveles muestreados presentan diferencias estadísticamente significativas para todas las especies. Las medidas de abundancia relativa de especies por niveles muestreados indican porcentajes muy bajos en la base del muestreo, con valores entre 0,45 - 0,42% -nivel 10 a 8- (Tabla 3). La mayor abundancia de la fauna malacológica se registra en el nivel 6 (2,24 m - 2,12 m) con un máximo porcentaje del 50,10%, el cual podría estar identificando el optimo climático (ver discusión). Desde este nivel, comienza a descender nuevamente la representatividad de las especies presentes en el sitio desde un 39,78% a 1,55%, en relación con el total de individuos recuperados en el muestreo. En cuanto a la abundancia relativa de especies (Tabla 3), el mayor porcentajes es de Heleobia parchappii (89,80%), en menor medida Biomphalaria peregrina (4,62%) y Chilina parchappii -D'Orbigny, 1835- (0,16%). Dentro de las especies terrestres, la mayor representación le corresponde a Systrophiidae -Baker, 1925- (3,59%), Succinea meridionalis -D'Orbigny, 1835- (0,71%), Retidiscus reticulatus -Fonseca y Thomé, 1995- (0,63%) y Zilchogyra costellata -D'Orbigny, 1835- (0,49%). Es notable la abundancia de Heleobia parchappii en relación con las otras especies presentes.
Tabla 3. Gasterópodos en Paso Otero 1. Muestreo general del perfil.
En el caso del muestreo puntual de las dos superficies de estabilización se observa la misma tendencia reflejada en el muestreo general (Tabla 4). Es notable la presencia de la especie Heleobia parchappii (91,71%) y en una proporción mucho menor Systrophiidae (4,57%), Biomphalaria peregrina (1,96%), Succinea meridionalis (0,94%), Retidiscus reticulatus (0,75%) y Chilina parchappii (0,03%). En la superficie de estabilización Superior la representatividad de especies malacológicas está dada en orden decreciente por Heleobia parchappii, Systrophiidae, Succinea meridionalis, Retidiscus reticulatus, Biomphalaria peregrina y Zilchogyra costellata.
Tabla 4. Superficie de estabilización Media (SEM) y Superficie de estabilización Superior (SES).
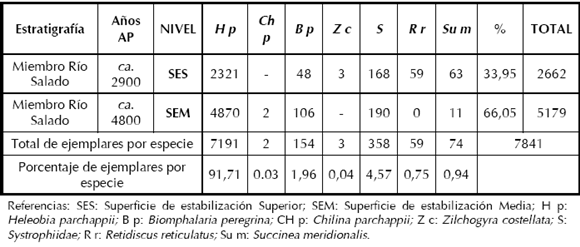
Por otra parte, en el muestreo de las superficies de estabilización Media y Superior, la mayor abundancia relativa de gasterópodos se registra en la superficie de estabilización Media con un 66,05% y coincide con el máximo porcentaje registrado de algas (Charophytas) en el sitio. Estos sectores con mayor presencia de gasterópodos y algas, podrían estar indicando un ambiente con abundancia de nutrientes tales como, nitrógeno, fósforo y otros elementos utilizados por este tipo de organismos. Esta situación observada coincide con las inferencias previas del ambiente de depositación del sitio (Johnson et al. 1997). Por lo tanto, durante el desarrollo de los paleosuelos estos microfósiles han favorecido condiciones reductoras en el ambiente de depositación debido al aporte de gran cantidad de materia orgánica. Por lo tanto, dicha situación es un elemento más de apoyo a lo interpretado indirectamente respecto de ambas superficies de inundación (Holliday 1995), lo cual permitiría apoyar la inferencia de que los microorganismos actuaron en un ambiente anaeróbico sobre los conjuntos óseos de Paso Otero 1 (Gutiérrez 2001).
Para analizar los resultados obtenidos y contrastar las hipótesis planteadas en el presente trabajo, se realizaron una serie de pruebas estadísticas para comparar las diferencias entre ambas superficies (SEM y SES) y entre los distintos niveles estratigráficos analizados. Dado que el muestreo que se utilizó genera distribuciones multinomiales se aplicó la comparación de proporciones de varias muestras según el test paramétrico chi-cuadrado. La hipótesis nula que se planteó considera que las proporciones de gasterópodos eran iguales para cada especie aunque estuvieran a distinta profundidad (p1=p2). En contraposición, una hipótesis alternativa suponía que en alguna profundidad la proporción de gasterópodos era diferente (p1≠p2).
En aquellos casos, en que el valor c2 observado (c2 o) fuese mayor que el c2 crítico (c2 c) se rechazó la igualdad de medias. Considerando cada especie para las diferentes profundidades (Tabla 3): Hp (c2 o 553,31 >c2 c 15,06); S (c2 o 519,11 > c2 c 15,06); Bp (c2 o 730,4 >c2 c 15,06); Sum (c2 o 937,15 >c2 c 18,47). En el caso de las SEM y la SES para cada especie: Hp (c2 o 118,5 >c2 c 6,63); S (c2 o 18,5 >c2 c 6.63); Bp (c2 o 53,65 >c2 c 6,63); Sum (c2 o 89,90 >c2 c 6,63). En el caso de Rr, Zc y Chp debido a su escasa presencia en el perfil en todos los casos c2 o fue mayor que c2 c.
Los resultados obtenidos indican que las muestras presentan diferencias cuantitativas estadísticamente significativas en lo referente al valor de medias. Es por ello que para todas las especies se rechazó la hipótesis nula (p1= p2), por lo tanto se considera que existen diferencias de proporciones entre los niveles muestreados del perfil estratigráfico de Paso Otero 1.
DISCUSIÓN
La diversidad taxonómica y la distribución de las frecuencias de las especies malacológicas en ambas superficies de estabilización no presentan una correspondencia de variación proporcional a los valores obtenidos a través del análisis de los isótopos estables del oxígeno y el carbono (d O18 y d C13) (Steffan 2000). Por un lado, los valores isotópicos indican variaciones de O18 de -0,03‰ y de C13 -0,76‰ entre ambas superficies, que resultan leves e insuficientes como para modificar el ambiente entre la SEM y la SES. Sin embargo, la fauna malacológica indica variaciones de temperatura y humedad en el ambiente que ocuparon. Esto puede ser explicado si se tiene en cuenta que dicha fauna es muy sensible a las variaciones en el microambiente, especialmente considerando los períodos de marcadas fluctuaciones como inviernos muy fríos y veranos muy intensos. El ambiente de depositación habría estado fuertemente influenciado por la estacionalidad, según se infiere de los datos malacológicos.
El otro aspecto a considerar en el presente trabajo es si las condiciones cálidas y húmedas del Holoceno medio, tal como lo reflejan las especies representadas en el muestreo del sitio Paso Otero 1, afectó a la malacofauna cualitativamente y/o cuantitativamente. Para la interpretación de los resultados obtenidos se parte de tres supuestos, teniendo en cuenta los condicionamientos ecológicos de cada especie: a) una alta frecuencia de las especies dulceacuícolas asociada a una muy baja frecuencia de especies de ambientes terrestres, indicarían un intenso pulso húmedo; b) la disminución de especies dulceacuícolas evidenciaría un cambio hacia condiciones más secas, desde la superficie de estabilización media a la superior; o bien, c) una alta frecuencia de especies terrestres sería un indicador de condiciones áridas a semiáridas registradas en el perfil de Paso Otero 1.
Con respecto a la abundancia de individuos dentro de las especies dulceacuícolas representadas en el muestreo de la SEM y SES (Tabla 5), tanto Biomphalaria peregrina como Heleobia parchappii, están presentes en ambas superficies y evidencian una marcada disminución desde la SEM hacia la SES. Al mismo tiempo, se observa la ausencia de la especie Chilina parchappii en la superficie de estabilización Superior. Por su parte, las especies terrestres evidencian diferencias en la presencia y frecuencia de algunos taxones, pudiendo responder esta situación a la dinámica de población propia de cada especie. En el caso de Succinea meridionalis está presente en ambas superficies, incrementándose su frecuencia hacia la SES. El porcentaje de abundancia relativa para la familia Systrophiidae es inversamente proporcional al de la especie Succinea y su presencia disminuye hacia la superficie de estabilización Superior en un 6%. La presencia y/o aparición en el registro de las especies terrestres Retidiscus reticulatus y Zilchogyra costellata, representarían un elemento más a favor de establecer diferencias ambientales entre ambas superficies de estabilización.
Tabla 5. Porcentaje de abundancia de individuos por especie.
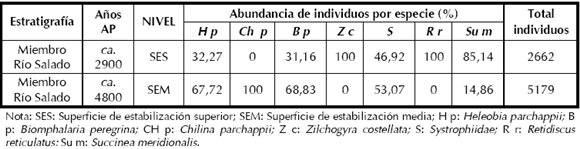
En suma, cabe concluir que de acuerdo al supuesto a); una alta frecuencia de especies de agua dulce y una baja proporción de especies terrestres, se registra en la superficie de estabilización Media (ca. 4800 años AP) en el sitio Paso Otero 1. Esto evidencia un pulso húmedo con concentración de precipitaciones y eventos de inundación. Dicha situación parece apoyar el supuesto b) según el cual, la disminución de especies dulceacuícolas evidenciaría un cambio hacia condiciones paleoclimáticas más secas, debido a una disminución en el porcentaje de individuos para las especies de agua dulce desde la SEM a la SES.
Por otra parte, el supuesto c) hace hincapié en el aumento de especies terrestres como un indicador de condiciones áridas a semiáridas en los sectores muestreados. Debido a la abundancia relativa de Systrophiidae, luego Succinea meridionalis y con un porcentaje mucho menor Retidiscus reticulatus y Zilchogyra costellata, asociadas con especies dulceacuícolas, redunda en un elemento más de apoyo en favor del supuesto c) y a la tendencia general planteada en el supuesto b).
Con la referencia al pulso cálido y húmedo del Holoceno medio, se hace alusión a las características generales del evento climático denominado Hyspistermal en la década de 1970. La evidencia actual generada indica que no necesariamente un aumento de la temperatura debería estar acompañado por un aumento de la humedad, en todos los sectores del mundo (i.e., Blunier y Brook 2001; Delcourt et al. 2002). A escala global, la información geoquímica demuestra que las temperaturas se mantuvieron elevadas entre ca. 7000 y 3000 años AP (ver Thompson 2000). En algunos sectores del sudeste del área Interserrana se registraron diferentes eventos para el Holoceno medio (i.e., Aguirre y Whatley 1995; Isla 1989; Tonni 1992; Zanchetta 1995). En el estuario del río Quequén Grande, se registró una creciente influencia marina asociada con el óptimo climático, desde los ca. 6700 años AP hasta aproximadamente los 5300 años AP (i.e., De Francesco y Zárate 2001; Golfieri et al. 1998). Sin embargo, este evento climático parece registrarse en el sitio Paso Otero 1 hacia los ca. 4800 años AP, considerando la edad mínima de la pedogénesis del paleosuelo datado.
Desde un punto de vista paleoambiental la situación observada en la SEM (ca. 4800 años AP) registra básicamente condiciones húmedas o al menos de una mayor humedad que en la superior. Sin embargo, el pulso cálido estaría mejor registrado hacia los ca. 2900 años AP, el cual no había sido registrado previamente. Este aumento de la temperatura estaría indicado en el registro paleoecológico por la disminución de Biomphalaria peregrina en relación con su frecuencia a lo largo del perfil muestreado. Por otra parte, se encuentran individuos de tamaño pequeño lo que indicaría una temprana edad de mortalidad de esta especie y dicha situación ocurre cuando se provocan ascensos de la temperatura, acompañado por fuertes vientos y prolongados períodos de desecación del cuerpo de agua en el que habitan (Bonetto et al. 1990; Rumi y Haman 1990). Tales aproximaciones corresponden a la observación de hechos actuales que permite una extrapolación para inferir condiciones ambientales ocurridas en el pasado.
Al mismo tiempo, se registra la presencia de las especies Retidiscus reticulatus y Zilchogyra costellata. La primera de ellas, se distribuye hacia el sur del Brasil (Rio Grande do Sul) y no tiene registro actual en la provincia de Buenos Aires. En cambio, la especie Zilchogyra costellata presenta una distribución moderna similar a la del pasado. La presencia de estas especies estaría indicando condiciones de clima húmedo y cálido entre los ca. 4800 AP a los ca. 2900 años AP en el sitio Paso Otero 1 (Miquel y Steffan 2005). En un nivel de generalización mayor, esto podría indicar que hacia los ca. 2900 años AP, se habría dado la reinstauración de un nuevo período tendiente a la aridización indicado por la disminución de Heleobia, Biomphalaria y la desaparición de Chilina. Por otro lado, evidencia la capacidad de la fauna malacológica a sobrevivir ante las presiones paleoclimáticas impuestas en estos momentos de cambios. En este sentido, es notable el comportamiento similar de la familia Systrophiidae, con las especies de agua dulce, ya que disminuye la cantidad de individuos desde la superficie de estabilización Media hacia la Superior. Cabe destacar que su presencia porcentual es relevante con relación al porcentaje total de la muestra. Pero en términos generales, Systrophiidae está indicando la misma tendencia hacia un ambiente más árido que fue observada para las otras especies.
CONCLUSIONES
Durante los ca. 4800 AP a los ca. 2900 años AP en el sitio Paso Otero 1, las condiciones paleoambientales, básicamente de temperatura y de humedad, presentan diferencias estadísticamente significativas, por lo tanto se rechaza la hipótesis planteada en el presente trabajo. La misma había supuesto condiciones ambientales similares. Sin embargo el análisis aquí realizado permite inferir que además de las variaciones estacionales, ocurrieron eventos climáticos a escala local asociados a anomalías en las precipitaciones sin precedentes en la actualidad, capaces de producir eventos de inundación y desplazamientos zoogeográficos de las especies malacológicas.
En relación al pulso cálido y húmedo registrado en otras regiones del área Interserrana entre los ca. 6700 y 5300 años AP, en el sitio Paso Otero 1 se observa un pulso de mayor humedad y un incremento sustancial en la temperatura desde los ca. 4800 años AP. Si bien se reconoce la necesidad de contar con fechados radiocarbónicos que permitan ubicar con mayor precisión este evento climático, por el momento la edad mínima de la pedogénesis de la superficie de estabilización Media puede ser considerada como una edad de referencia para el área y el sitio estudiado aquí.
Luego de este período de mejoramiento climático, habría continuado un momento signado por un descenso de la humedad ambiental, mientras la temperatura habría permanecido constante. El registro malacológico revela que las especies dulceacuícolas habrían sido las más afectadas por las variaciones ambientales. Al mismo tiempo, la mayor abundancia relativa de las especies terrestres indicaría la reinstauración de un nuevo período tendiente a la aridización. La presencia de la especie Retidiscus reticulatus acotada entre ca. 4800 y 2900 años AP estaría reflejando el mantenimiento de condiciones cálidas y posteriormente habría ocurrido un descenso de la temperatura.
A partir del análisis malacológico del sitio Paso Otero 1, se observa una adecuada correspondencia entre la interpretación puntual del ambiente y las variaciones climáticas inferidas para los últimos diez mil años. En el lapso temporal comprendido entre la base del Miembro Río Salado, se habrían dado condiciones áridas, con un período de mayor humedad relativa del ambiente. Dicha situación estaría indicada particularmente por la especie Heleobia parchappii. Luego de este evento de mayor humedad continuaría la instauración de una nueva etapa marcada por la aridez, evidenciada por la disminución de los géneros de Heleobia y Biomphalaria.
Si bien el análisis llevado a cabo en este trabajo estuvo acotado al sitio Paso Otero 1, es posible proyectar las conclusiones alcanzadas aquí a los sitios arqueológicos localizados en el área adyacente a Paso Otero 1 (i.e., Paso Otero 3, Zanjón Seco 2 y 3 de la Localidad arqueológica Zanjón Seco). Las conclusiones aquí obtenidas brindan información para comprender el patrón de variabilidad paleoclimática para el área durante el Holoceno y permite tener una escala de referencia para inferir el patrón en la distribución de los recursos disponibles para el área durante el lapso temporal analizado.
Con respecto a las aproximaciones paleoambientales basadas en datos cuantitativos de gasterópodos cabe destacar su utilidad de aplicación como "proxy" para detectar variaciones microambientales que no fueron registradas a partir de las variaciones en los valores de los isótopos estables medidos desde las conchillas. Al mismo tiempo, permiten aportan inferencia basadas en evidencias del incremento de temperatura y de la humedad durante el Holoceno medio, muchas veces interpretados como una asociación contingente. Sin embargo, la señal de un elemento no implica la ocurrencia del otro con igual intensidad.
Agradecimientos
Agradezco la valiosa orientación y discusiones de M. Gutiérrez y J. Prado durante la realización del trabajo, quienes impulsaron y estimularon esta investigación. La revisión crítica y los enriquecedores aportes de E. Tonni y M. Aguirre. La colaboración de S. Miquel, A. Rumi, C. De Francesco en la identificación de especies. A G. Gómez, P. Messineo, M. Cocconi y S. Salomon. El trabajo fue realizado con subsidios del INCUAPA otorgados por la SeCyT (UNCPBA) y beca doctoral del CONICET.
REFERENCIAS CITADAS
1. Aguirre M. y E. Farinati 2000 Aspectos sistemáticos, de distribución y ambientales de Littoridina australis (d'Orbigny, 1835) (Mesogastropoda) en el Cuaternario marino de Argentina (Sudamérica). Geobios 33: 569-597. [ Links ]
2. Aguirre M. y R. Whatley 1995 Late Quaternary marginal marina deposits from north-eastern Buenos Aires province, Argentina: A review. Quaternary Science Review 14: 223-254. [ Links ]
3. Baker, H. B. 1925 The mollusca collected by the University of Michigan-Williamson Expedition in Venezuela. Part III. Ocassional Papers of the Museum of Zoology 156, University of Michigan, Michigan. [ Links ]
4. Bertels, A. y D. Martinez 1990 Quaternary ostracodes of continental and transitional littoral-shallow marine environments. Courier Forschungs-instut senckenberg 132: 141-159. [ Links ]
5. Blunier, T. y E. Brook 2001 Timing of millennial-scale climate change in Antarctica and Greenland during the last glacial period. Science 291: 109-114. [ Links ]
6. Bonadonna, F., G. Leone G. y G. Zanchetta 1995 Composición isotópica de los fósiles de gasterópodos continentales de la provinca de Buenos Aires. Indicadores paleoclimáticos. En Evolución biológica y climática de la región pampeana durante los últimos 5 millones de años. Un ensayo de correlación con el Mediterráneo occidental, editado por M. T. Alberdi, G. Leone y E. P. Tonni, pp. 77-104. Monografías del Museo Nacional de Ciencias Naturales, Madrid. [ Links ]
7. Bonetto, A., A. Rumi y M. Tassara 1990 Notas sobre el conocimiento limnológico de los gasterópodos paranaenses y sus relaciones tróficas. II Planorbidae, con aspectos distribucionales y sanitarios. Ecosur 16 (28): 69-84. [ Links ]
8. Bobrowsky, P. 1984 The history and science of Gastropods in archaeology. American Antiquity 49 (1): 77-93. [ Links ]
9. Carbonari, E., R. Huarte, A. Figini 1992 Miembro Guerrero Formación Luján (Pleistoceno, Pcia de Buenos Aires) edades C14. Actas de las Terceras Jornadas Geológicas Bonaerenses, pp. 245-247. La Plata. [ Links ]
10. Castellanos, Z. y S. Miquel 1991 Distribución de los Pulmonata Basommatophora. Fauna de agua dulce de la República Argentina 15 (8): 1-11. [ Links ]
11. Cazzaniga, N. 1981 Notas sobre Hidróbidos argentinos. III (Gastropoda Rissoidea). Strobeliella, un nuevo género de la Patagonia. Neotropica(La Plata) 27 (77): 3-10. [ Links ]
12. Darrigran, G. 1995 Aspectos ecológicos de la malacofauna litoral del Río de la Plata, República Argentina. Tesis Doctoral inédita. Facultad de Ciencias Naturales y Museo, Universidad Nacional de La Plata, La Plata. [ Links ]
13. De Francesco, C. y F. Isla 2003 Distribution and abundance of Hydrobiid snails in a mixed estuary and a Coastal Lagoon, Argentina. Estuaries 26 (3): 790-797. [ Links ]
14. De Francesco, C. y F. Isla 2004 Reproduction and growth of the freshwater snail Heleobia parchappii (d'Orbigny, 1835) (Gastropoda: Rissooidea) in a shallow brackish habitat Buenos Aires Province, Argentina. Malacologia 45 (2): 443- 450. [ Links ]
15. De Francesco, C. y M. Zárate 2001 Bioturbaciones de Tagelus plebeius (Lingtfoot, 1785) (MOLLUSCA: BIVALVIA) en un ambiente estuárico Holoceno (Río Quequén Grande, Buenos Aires, Argentina). Ameghiniana 38 (4): 467-470. [ Links ]
16. Delcourt, P., P. Nesterm, H. Delcourt, C. Mora y K. Orvis 2002 Holocene lake effect precipitation in Northern Michigan. Quaternary Research 59: 225-233. [ Links ]
17. D'Orbigny, A. 1835 Sinopsis terrestrium et fluviatilium molluscorum, in suo per American meridionalen itinere. Magazine Zoology 5 (61): 1-44. [ Links ]
18. Favier Dubois, C. 2006 Dinámica fluvial, paleoambientes y ocupaciones humanas en la localidad Paso Otero, río Quequén Grande, Pcia. de Buenos Aires. Intersecciones en Antropología 7: 109-127. [ Links ]
19. Fidalgo, F., F. De Francesco y U. Colado 1973 Geología Superficial en la Hojas Castelli, J. M. Cobo y Monasterio (Pcia. de Buenos Aires). Actas del Quinto Congreso Geológico Argentino IV, pp. 27-39. Buenos Aires. [ Links ]
20. Fonseca, Á. y J. Thomé 1995 A new genus and species of Charopidae (Gastropoda, Endodontoidea, Stylommatophora), Retidiscus reticulatus, from southern Brazil. Walkerana 7 (17/18): 55-62. [ Links ]
21. Gaillard, M. y Z. Castellanos 1976 Hydrobiidae. Fauna de Agua Dulce de la República Argentina 15 (2): 1-40. [ Links ]
22. Garcia, A. 1996 Charophyta y Ostracoda asociados de cuatro localidades holocenas de Argentina: Evidencias paleoambientales. Ameghiniana 33 (4): 409-420. [ Links ]
23. Golfieri, G., L. Ferrero y M. Zárate 1998 Tafonomía y paleoecología de Tagelus plebeius (Lightfoot, 1786) (MOLLUSCA, BIVALVIA) en sedimentos holocenos del Río Quequén Grande, Provincia de Buenos Aires, Argentina. Ameghiniana 33 (3): 255-264. [ Links ]
24. Gutiérrez, M. 2001 Bone diagenesis and taphonomic history of the Paso Otero 1 bone bed, Pampas of Argentina. Journal of Archaeological Science 28: 1277-1290. [ Links ]
25. Gutiérrez, M. 2004 Análisis tafonómico en el Área Interserrana (Provincia de Buenos Aires). Tesis Doctoral inédita. Facultad de Ciencias Naturales y Museo, Universidad Nacional de La Plata, La Plata. [ Links ]
26. Gutiérrez, M. y C. Kaufman 2007 Criteria for the Identification of Formation Processes in Guanaco (Lama guanicoe) Bone Assemblages in Fluvial-Lacustrine Environments. Journal of Taphonomy 5 (4): 151-175. [ Links ]
27. Holliday, V. 1995 Stratigraphy and Paleoenvironments of Late Quaternary Valley fills on the Southern High Plains: Geological Society of America, Memoir 186: 135. [ Links ]
28. Isla, F. 1989 Holocene sea level fluctuation in the Southern Hemisphere. Quaternary Science Reviews 8: 359- 368. [ Links ]
29. Johnson, E., M. A. Gutiérrez, G. Politis, G. Martinez y W. Hartwell 1997 Holocene taphonomy at Paso Otero 1 on the eastern Pampas of Argentina. En: Proceedings of the 1993 Bone Modification Conference, Hot Spring, South Dakota, editado por L. A. Hannus, L. Rossum y R. P. Winham, pp. 105-121. Occasional Publication Nro. 1, Archaeology Laboratory, Augustana College, Sioux Fall. [ Links ]
30. Johnson, E., G. Politis, G. Martinez, W. Hartwell, M. Gutiérrez y H. Hass 1998 The Radiocarbon chronology of Paso Otero 1 in the Pampean Region of Argentina. Quaternary of South America Antarctic Peninsula 11: 15-25. [ Links ]
31. Landoni, N. 1992 Inventario de los moluscos de agua dulce de la Provincia de Buenos Aires. En Situación ambiental de la Provincia de Buenos Aires. Recursos y Rasgos Naturales en la evaluación ambiental, editado por H. Lopez y E. P. Tonni, pp. 3-57. Vol. II (17). Ministerio de la Producción y Comisión de Investigaciones Científicas, La Plata. [ Links ]
32. Martínez, G. 1999 Tecnología, subsistencia y asentamiento en el curso medio del Río Quequén Grande: un enfoque arqueológico. Tesis Doctoral inédita. Facultad de Ciencias Naturales y Museo, Universidad Nacional de La Plata, La Plata. [ Links ]
33. Miquel, S. y P. Steffan 2005 Hallazgo de Retidiscus reticulatus (Gastropoda Pulmonata Charopidae) en la Argentina, un posible indicador de paleoclima subtropical húmedo en el Holoceno de la provincia de Buenos Aires. Ameghiniana 42 (2): 495-500. [ Links ]
34. Politis, G., M. Gutiérrez y G. Martínez 1991 Informe preliminar de las investigaciones en el Sitio Paso Otero 1 (Pdo. de Necochea, Pcia. de Buenos Aires). Boletín del Centro de Registro Arqueológico y Paleontológico 2: 80-90. [ Links ]
35. Prado, J. L., A. Menegaz, E. P. Tonni y M. Salemme 1987 Los mamíferos de la fauna local Paso Otero (Pleistoceno tardío), provincia de Buenos Aires. Aspectos paleoambientales y bioestratigráficos. Ameghiniana 24: 217-233. [ Links ]
36. Prado, J. L., B. Azanza, M. T. Alberdi y G. Gómez 2001 Mammal community and global change during the Late Pleistocene-Holocene in the Pampean Region (Argentina). Intenationale Archäeologie. Studia honoraria - Band 11: 362-375. [ Links ]
37. Prieto, A. 1996 Late Quaternary vegetation and climatic changes in the Pampa Grassland of Argentina. Quaternary Research 45: 75-88. [ Links ]
38. Rumi, A. 1991 La familia Planorbidae Rafinisque, 1815 en la República Argentina. Fauna de agua dulce de la República Argentina 15 (8): 5-51. [ Links ]
39. Rumi, A. y M. Hamann 1990 Potential Schistosome vector snails and associated Trematodes in ricefields of Corrientes Province, Argentina. Preliminary results. Memorias Instituto Osvaldo Cruz 85 (3): 321-328. [ Links ]
40. Rumi, A., M. Tassara y A. Bonetto 1997 Distribución de las especies de planorbidae en Argentina y su relación con el riesgo de esquistosomiasis. Ecosur 17 (28): 1-19. [ Links ]
41. Steffan, P. 2000 Implicancias paleoambientales en el sitio Paso Otero 1 (Pcia. de Buenos Aires). Análisis de isótopos estables de 13C y 18O y de la composición de la fauna malacológica. Tesis de Licenciatura inédita. Facultad de Ciencias Sociales, Universidad Nacional del Centro de la Provincia de Buenos Aires, Olavaria. [ Links ]
42. Thompson, L. 2000 Ice core evidence for climate change in the tropics: implications for our future. Quaternary Science Review 19: 19-35. [ Links ]
43. Tonni, E. P. 1992 Mamíferos y clima en la provincia de Buenos Aires. CADINQUA 1: 64-78. [ Links ]
44. Tonni, E. P., A. Cione y A. Figini 1999 Predominance of arid climates indicated by mammals in the pampas of Argentina during the late Pleistocene and Holocene. Palaeogeography, Palaeoclimatology, Paleoecology 147: 257-281. [ Links ]
45. Zanchetta, G. 1995 Estado actual de la geología y estratigrafía de los depósitos plio-pleistocenos de la Región Bonaerense. En Evolución biológica y climática de la Región Pampeana durante los últimos cinco millones de años, editado por M. T. Alberdi, G. Leone y E. P. Tonni, pp. 29-45. Monografías del Museo Nacional de Ciencias Naturales de Madrid, Madrid. [ Links ]

