










Services on Demand
Journal

Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO
Related links
-
 Similars in
SciELO
Similars in
SciELO
Share

 Permalink
PermalinkIntersecciones en antropología
On-line version ISSN 1850-373X
Intersecciones antropol. no.9 Olavarría Jan./Dec. 2008
Evidencias bioarqueológicas en patagonia meridional: el sitio Orejas de Burro 1 (Pali Aike, provincia de Santa Cruz)
Gabriela L. L'Heureux y Ramiro Barberena
Gabriela L. L'Heureux. CONICET, IMHICIHU, Departamento de Investigaciones Prehistóricas y Arqueológicas. Saavedra 15, piso 5, (C1083 ACA) Buenos Aires, Argentina. E-mail: lorenalheureux@yahoo.com.ar
Ramiro Barberena. CONICET, IMHICIHU, Departamento de Investigaciones Prehistóricas y Arqueológicas. Saavedra 15, piso 5, (C1083 ACA) Buenos Aires, Argentina. E-mail: ramidus28@fibertel.com.ar
Recibido 21 de Diciembre 2007.
Aceptado 9 de Abril 2008
RESUMEN En este trabajo se presentan los resultados de las excavaciones realizadas recientemente en el sitio Orejas de Burro 1, localizado en el campo volcánico Pali Aike, provincia de Santa Cruz, Argentina. En el mismo se recuperó un entierro humano compuesto por cinco individuos de diferentes edades depositados en un único evento de inhumación. La información desarrollada incluye datos estratigráficos, contextuales, bioarqueológicos e isotópicos, que contribuyen a la evaluación de temas mortuorios y biogeográficos discutidos en el Proyecto Magallania.
Palabras clave: Pali Aike; Entierro múltiple; Holoceno tardío.
ABSTRACT Bioarchaeological Evidence In Meridional Patagonia. The Orejas De Burro 1 Site (Pali Aike, Santa Cruz Province). The results from recent excavations at the Orejas de Burro 1 site (Santa Cruz Province, Argentina) are presented in this paper. A human burial was recovered with the remains of five individuals from different age groups that were deposited simultaneously. Stratigraphic, contextual, bioarchaeological, and isotopic data are provided. This data contributes to discussions of mortuary behavior and human biogeography currently being developed within the Magallania Project.
Keywords: Pali Aike; Multiple burial; Late Holocene.
EL SITIO OREJAS DE BURRO 1
En este trabajo se presenta la información correspondiente a un entierro humano múltiple recuperado en el sitio Orejas de Burro 1 (OB1), ubicado en el campo volcánico Pali Aike (CVPA), sur de la provincia de Santa Cruz, Argentina (Figura 1). La integración de datos bioarqueológicos y geoarqueológicos permite evaluar la génesis de este conjunto y contribuir de esta manera con el estudio de las prácticas mortuorias en Patagonia meridional. OB1 se localiza en el sector sureste del campo de lava (52º 07.769' S, 69º 33.151' O), en una cueva ubicada en la ladera interna de un cono volcánico. Fechados de ![]() realizados sobre las rocas que conforman estos rasgos volcánicos proveen una edad de 0,36 millones de años AP para su formación (Corbella 1999). La cueva tiene aproximadamente 12 m de largo por 5 m de ancho, y su boca se encuentra orientada al noreste (rango de visión ubicado entre 50ºy 90º). Presenta una posición topográfica destacada, dado que el sitio se ubica en un sector elevado del cono, aunque existen restricciones a la visibilidad impuestas por el borde opuesto del mismo. Desde el oeste se puede apreciar la forma característica del rasgo que da origen a su nombre (Figura 2).
realizados sobre las rocas que conforman estos rasgos volcánicos proveen una edad de 0,36 millones de años AP para su formación (Corbella 1999). La cueva tiene aproximadamente 12 m de largo por 5 m de ancho, y su boca se encuentra orientada al noreste (rango de visión ubicado entre 50ºy 90º). Presenta una posición topográfica destacada, dado que el sitio se ubica en un sector elevado del cono, aunque existen restricciones a la visibilidad impuestas por el borde opuesto del mismo. Desde el oeste se puede apreciar la forma característica del rasgo que da origen a su nombre (Figura 2).

Figura 1. El Campo Volcánico Pali Aike. Localización del sitio Orejas de Burro 1.

Figura 2. El cono volcánico Orejas de Burro y la boca de entrada de la cueva ubicada en la ladera sur del volcán.
Durante los primeros trabajos en el sitio se observaron montículos de sedimento removido en la entrada de la cueva con abundantes restos óseos y valvas de Mytilus sp., y en el centro de la misma un área alterada bien delimitada con forma pseudo circular (Figura 3). Estas modificaciones se reconocieron como el producto de viejas excavaciones realizadas por aficionados en la década de 1970, que permitieron la recuperación de un entierro doble (Guerra de Fretes 1977), actualmente perdido1. Al sur de esta área y en un sector libre de rocas de derrumbe, se realizó un sondeo inicial de 0,25 m2 que alcanzó 95 cm de profundidad. La excavación se detuvo al hacer contacto con un cráneo humano en el perfil sur del sondeo. Los trabajos prosiguieron al año siguiente, cuando se plantearon cuatro cuadrículas para recuperar el individuo detectado. En esta instancia se recuperó un entierro compuesto por cinco individuos de diferentes edades, que es el objeto de este trabajo. La excavación alcanzó la roca de caja (ca. 91 cm) y se completaron 1,75 m2 excavados (el sondeo previamente realizado formó parte de lo que se denominó posteriormente cuadrícula 1).
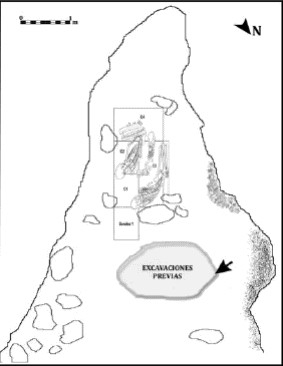
Figura 3. Esquema de la cueva OB1. Ubicación del sector excavado por el equipo y del área alterada por aficionados. Se señaliza donde se halló la epífisis distal de húmero humano.
En el sector del entierro se reconocieron tres de las once unidades estratigráficas identificadas en el sitio (Barberena et al. 2006). La unidad I está compuesta por un nivel de guano disgregado y por un nivel subyacente de guano muy compacto con estratificación tabular (0-38 cm). La unidad X (38-70/75 cm) representa el sedimento que apoya sobre los bloques de roca de caja que cubren al entierro. La unidad XI (70/75-90 cm) corresponde a la matriz del entierro que se encuentra en contacto con los basaltos de la roca de base (Figura 4). Se defiende que las unidades X y XI tienen una génesis antrópica, en asociación con la depositación del entierro (ver infra).
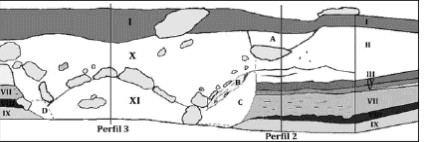
Figura 4. Unidades estratigráficas del sitio OB1. Ver detalles de las referencias de las unidades y los rasgos señalados en Barberena et al. (2006).
El sitio cuenta con seis fechados radiocarbónicos realizados sobre diferentes materiales (Tabla 1), que evidencian un uso redundante de la cueva durante el Holoceno tardío desde aproximadamente 4000 años AP en adelante.
Tabla 1. Fechados disponibles para el sitio Orejas de Burro 1.
CONTEXTO ESTRATIGRÁFICO Y MORTUORIO
En el sector más profundo de la cueva OB1 se inhumaron sincrónicamente cinco individuos hace ca. 3500 años, en una estructura dentro de un área excavada intencionalmente para tal fin. Los esqueletos fueron cubiertos directamente con grandes bloques de roca volcánica de la cueva (Figura 5), conformando una estructura de entierro semejante a las denominadas "chenque", las que se reconocen en el área desde el Holoceno tardío final hasta momentos históricos. A diferencia de este tipo de inhumaciones, usualmente construidas a cielo abierto y a nivel de la superficie, el caso que presentamos es subsuperficial y se emplaza dentro de una cueva. Un antecedente en lo formal de este tipo de entierros en cueva se encuentra en el sitio Baño Nuevo, en la región de Aisén, Patagonia chilena (Mena et al. 2003).

Figura 5. La excavación de la estructura de entierro. El individuo 2 todavía muestra un gran bloque sobre el tórax. Parte de los individuos 1 y 3 se encuentran debajo de rocas.
Dentro de las características contextuales del enterratorio se destaca la presencia de una superficie artificial de vegetales -que probablemente corresponden a gramíneas- depositada por sectores debajo de los individuos y que funcionó como superficie de apoyo. Los vegetales se encontraban visibles en algunas de las regiones donde apoyaban los cuerpos tales como los cráneos, las pelvis y los pies. Además, los esqueletos presentaban abundantes rastros de ocre, que también se podían distinguir en los vegetales, sedimentos y huesos de fauna asociados. Se entiende que, como parte del rito de inhumación, loscuerpos fueron tratados directamente con ocre o, eventualmente, envueltos o cubiertos con una superficie (i.e., cuero) previamente teñida. En el nivel del entierro no se hallaron restos de este material, aunque en la unidad X que cubría la estructura se recuperaron pequeños fragmentos de cuero que no presentaban tinción con ocre. El tratamiento mortuorio otorgado a los individuos es homogéneo. Una excepción puede relacionarse con la mayor presencia de ocre en el individuos 3 (> 50% de los elementos teñidos).
No se detectaron otras modificaciones antrópicas predepositacionales. Se puede mencionar el hallazgo de tres cuentas óseas pulidas y tres punzones confeccionados sobre huesos de aves, dos de los cuales se encuentran grabados con incisiones oblicuas y paralelas, recuperados en la unidad X (L'Heureux 2008: Figuras 7 y 13). Estos ornamentos e instrumentos no pueden ser interpretados como parte del ajuar fuenerario, ya que no se hallaban en asociación directa con los esqueletos, sino por sobre y entre los vértices que formaban las rocas que los cubrían.
Posteriormente, esta estructura fue cubierta con los sedimentos removidos al cavar la fosa, numerosos huesos de guanaco redepositados y rocas de procedencia endógena. Como producto de este proceso se creó una unidad estratigráfica artificial (unidad X), con estructura de tipo clasto sostén dado los numerosos contactos existentes entre los huesos, rocas y otros elementos depositados conjuntamente con la matriz. Esto se corresponde con los criterios postulados por Gargett (1989) para discriminar entre entierros intencionales e incidentales (para una posición contraria ver Pettitt 2002).
El análisis estratigráfico realizado en conjunto con los aspectos contextuales del entierro permite defender que se trata de un único evento de depositación de los cinco esqueletos y de la matriz sedimentaria que los contiene. A nivel estratigráfico, se localizó y se pudo seguir una clara discordancia erosiva presente en todos los límites de la estructura del entierro. Esta discordancia interrumpe una serie de unidades estratigráficas preexistentes, que fueron detectadas a ambos lados del entierro (Figura 4). A nivel contextual, dada la estrecha vinculación horizontal existente entre los esqueletos, eventos sucesivos de depositación habrían producido cierto desarreglo anatómico en el conjunto, que no fue verificado. Eventos muy cercanos en el tiempo, ocurridos por ejemplo en el lapso de semanas y en forma previa a una pérdida importante del tejido blando, podrían constituir una excepción. No obstante, se considera que esto es poco probable en función de los datos estratigráficos presentados con respecto a los límites externos de la estructura. A su vez, para ciertos fines y en una escala arqueológica de trabajo, esta situación también puede ser tratada como de 'sincronía'. En otras palabras, el conjunto mortuorio presenta una gran resolución temporal, aunque la migración vertical de materiales no asociados puede haber ocurrido. Esto puede aplicarse particularmente a restos óseos de guanaco y de puma hallados en la matriz del entierro -unidad XI-, algunos de los cuales podrían estar asociados al mismo y otros no.
Otro aspecto contextual se refiere al conjunto óseo recuperado por sobre el entierro, compuesto principalmente por restos de guanaco (entre otros ítems faunísticos), que se encuentra contenido en la unidad estratigráfica X. Hay distintos aspectos formacionales que deben ser considerados y que contribuyen a discutir su vinculación con el entierro. Como ha sido planteado (Barberena et al. 2006), en primer lugar debe evaluarse su tempo de formación: puede tratarse de un conjunto formado atricionalmente o en forma simultánea. En segundo lugar, hay que considerar la posición cronológica de los materiales que componen el conjunto: ¿Se corresponde con la del entierro o es posterior? Esto tiene implicaciones para el análisis de las prácticas mortuorias, dado que la primera posibilidad alimenta una situación de asociación conductual entre los esqueletos humanos y el conjunto faunístico depositado por sobre los mismos.
En relación con el primero de los puntos, hay un conjunto de propiedades estratigráficas relevantes para mencionar. Como se ha planteado, la unidad X está compuesta por abundantes restos óseos de fauna de diferentes taxa -y escasos artefactos líticos- que le confieren una estructura clasto sostén. Por esto, muestra abundantes contactos entre los huesos, la presencia de espacios vacíos, huesos entrampados entre las superficies de contacto de los bloques y distribución heterogénea del sedimento fino que conforma la matriz. Todo esto permite defender unadepositación antrópica simultánea y caótica del conjunto faunístico que conforma dicha unidad. En este sentido, el mismo se habría depositado en forma sincrónica.
Con respecto a la relación cronológica entre el conjunto faunístico de la unidad X y el entierro -unidad XI-, existe una estrecha asociación estratigráfica, ya que los huesos de fauna de la unidad X apoyan directamente sobre los bloques que cubren el entierro. Esto puede sugerir cercanía cronológica, aunque claramente no la demuestra. Se realizaron fechados radiocarbónicos sobre una tibia de guanaco procedente del conjunto óseo del relleno -ubicado por sobre el entierro a 67 cm de profundidad- y sobre el esqueleto 1 del entierro. Al comparar las edades 14c de la muestra de guanaco (3490 ± 75 años AP) y la del esqueleto humano (3565 ± 45 años AP), se observa que son indistinguibles a nivel estadístico y se ubican en un mismo rango temporal, aún considerando solamente un sigma. Esto marca una situación de penecontemporaneidad entre ambos conjuntos, ratificando las inferencias realizadas a partir de los datos estratigráficos (Barberena et al. 2006). Nuevamente, esto no demuestra la contemporaneidad de ambos conjuntos, aunque la respalda en forma importante.
Teniendo en cuenta lo expuesto, el conjunto faunístico se habría depositado en forma penecontemporánea con respecto al entierro. Es difícil alcanzar una resolución temporal mayor que permita defender que los restos de guanaco formaron parte del mismo evento de depositación de los restos humanos. No obstante, todos los datos evaluados son consistentes con las expectativas estratigráficas y cronológicas que se desprenden de dicha situación.
Para concluir, se debe señalar que, además de los cinco individuos presentes en la estructura de entierro, se recuperó un sexto individuo representado por tres huesos sin relación anatómica (dos vértebras lumbares y una décima costilla derecha con ocre) que corresponden a uno o más individuos adultos. Estos huesos formaban parte del depósito de relleno que cubría el entierro y no tienen relación alguna con los esqueletos inhumados en el mismo. Se infiere que estos huesos fueron incorporados a la unidad X cuando se rellenó la fosa con los sedimentos retirados al cavarla y con otros adyacentes que aportaron rastros de entierros previos inhumados en la cueva. A estos deben sumarse los dos esqueletos excavados anteriormente por Guerra de Fretes (1977) ya mencionados. En relación con estos últimos, al limpiar los perfiles del sector alterado por los aficionados se recuperó una epífisis distal de húmero de un individuo adulto (Figura 3). Probablemente, tal elemento corresponda a alguno de los dos esqueletos excavados por este grupo, aunque esto no puede ser confirmado.
INFORMACIÓN BIOARQUEOLÓGICA E ISOTÓPICA
Los individuos
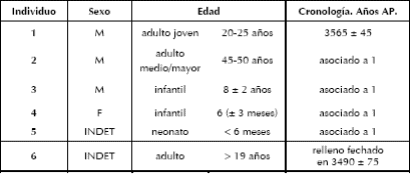
De los cinco individuos del enterratorio, dos son adultos de sexo masculino y los tres restantes son subadultos de sexo masculino (N° 3), femenino (N° 4) e indeterminado (N° 5) (Tabla 2). En los subadultos se pudo estimar el sexo de acuerdo con las variables cualitativas del ilion y la mandíbula (Schutkowski 1993; Weaber 1980), a excepción del individuo N° 5, que no presenta elementos óseos diagnósticos. La estimación del sexo en los adultos se realizó a partir del análisis de múltiples indicadores morfológicos en cráneo y pelvis (Buikstra y Ubelaker 1994). No resultó sencillo estimar este parámetro, ya que no siempre se registraron datos coherentes entre el cráneo y la pelvis de cada individuo, por lo que se decidió utilizar el promedio de los indicadores analizados. En la estimación de la edad de muerte de los individuos subadultos se tuvo en cuenta la secuencia de desarrollo y erupción dental (Ubelaker 1989). Para los individuos adultos se emplearon el estado de obliteración de las suturas craneales (Meindl y Lovejoy 1985) y los cambios en la morfología de la superficie auricular del ilion (Lovejoy et al. 1985); cabe aclarar que no se preservaron las sínfisis púbicas de ninguno de los individuos adultos. Dado lo pequeño que es el conjunto, la variabilidad etaria registrada resulta importante.
Tabla 2. Estimación sexual y etarea de los individuos de OB1.
Los esqueletos se hallaron depositados muy próximos entre sí y en un sector semi circular muy estrecho (ca. 1,75 m2). El individuo 1 se encontraba en posición decúbito lateral izquierdo, el cráneo en dirección este, los miembros superiores extendidos hacia la pelvis y los inferiores semiflexionados (Figura 6). Entre el húmero izquierdo y el tórax del individuo 1 se encontraron los restos de un individuo infantil (individuo 4) de alrededor de seis meses de vida (Tabla 2; Ubelaker 1989), en posición extendida decúbito dorsal con el esplacnocráneo apuntando al torso del individuo 1 (Figura 7). En la periferia de este individuo se encontraron otros elementos que en un principio se interpretaron como parte de la dispersión del esqueleto del individuo 4, pero que resultó representar otro infante, bastante más incompleto y disperso, del cual sólo puede decirse que es de menor edad que 4 (ya que no se recuperaron los gérmenes dentales para evaluar el estado de desarrollo de las coronas de los dientes deciduos). Este fue denominado individuo 5 y se decidió caracterizarlo como "neonato". A escasos 50 cm del cráneo de 1 y en dirección sur, se halló el individuo 2, depositado con una posición y orientación semejante al individuo 1, con los miembros superiores extendidos sobre la pelvis y los inferiores bien flexionados. Entre 1 y 2 se ubicaba un individuo juvenil de alrededor de 8 años, denominado individuo 3 (Tabla 2; Ubelaker 1989). Éste, en cambio, se encontraba depositado decúbito lateral derecho y con el cráneo apuntando hacia el oeste (Figura 6).
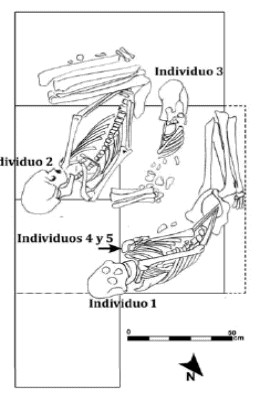
Figura 6. Planta de excavación. Localización, disposición y orientación de los cinco individuos recuperados.

Figura 7. Localización de los infantes. El individuo 4 apoya directamente sobre el húmero izquierdo del individuo 1. Próximo, se observa la hemimandíbula derecha del individuo 5.
Los esqueletos se encuentran alineados en un eje sur-este pero con diferentes orientaciones (1, 2 y 4 el cráneo orientado al este y 3 hacia el oeste). Los dos esqueletos recuperados por Guerra de Fretes (1977) tenían una disposición semejante, ya que se hallaban decúbito lateral, semiflexionados y alineados. Por otra parte, la orientación de los esqueletos recuperados por este grupo coincide con la del individuo 3 (cráneo hacia el oeste), mientras que es relativamente opuesta con respecto a los otros tres individuos.
Los esqueletos se hallaron en buen estado de preservación, aunque variable según los casos (ver más adelante). Igualmente, presentaron un grado de integridad diferencial. Los individuos 1 y 2 se encontraron completos y totalmente articulados. El individuo 3 se mantenía articulado y con una estrecha asociación anatómica, pero algo incompleto y con una preservación regular. El individuo 4 estaba en posición anatómica estrecha y en buen estado de conservación. El individuo 5 se encontraba representado sólo por diecisiete elementos (Tabla 3) con una preservación relativamente buena, aunque sin relación anatómica entre sí. Es importante mencionar que cuatro elementos del individuo 1 (falanges y falangetas) y tres elementos axiales de los infantiles (cuerpos de vértebras y fragmento de sacro) fueron hallados en un sector de cuevas o túneles de roedores localizados en el sondeo 1, próximo al área del entierro y colindante al esqueleto 1 (Franco et al. 2008).
Tabla 3. Estado de integridad y completitud de los esqueletos

Dada la alta integridad de los esqueletos, se pudo estimar la estatura de los individuos adultos a partir de la técnica del método anatómico (Fully 1956), la cual consiste en sumar las longitudes de todos los elementos óseos que conforman la estatura del individuo añadiéndole una corrección por los tejidos blandos y las edades de los mismos. De este modo se pudo reconstruir que el individuo 1 tenía en vida una estatura de aproximadamente 156,9 cm y el individuo 2 de 156,8 cm (M. Béguelin, comunicación personal 2007).
Indicadores de patologías
Se evaluó la presencia de patologías infecciosas, traumáticas y modificaciones degenerativas a nivel óseo y bucal (Buikstra y Ubelaker 1994; Clarke y Hirsch 1991; Hillson 1996; Larsen 1997; Mensforth 1991; Roger y Waldron 1995). En ninguno de los esqueletos se observaron evidencias de patologías infecciosas, traumas o violencia interpersonal a nivel esqueletal que pudieran asociarse con su causa de muerte. El único individuo que muestra una serie de modificaciones óseas relevantes tanto traumáticas como degenerativas es el 2, las que se relacionan con su avanzada edad y, posiblemente, con las exigentes actividades realizadas en vida asociadas con la caza y la recolección.
Este individuo mostraba reiteradas fracturas antiguas bilaterales localizadas en las últimas costillas del torax, algunas soldadas con desplazamiento (Figura 8). En las vértebras lumbares se observó una abundante formación de osteofitos alrededor de los bordes de los cuerpos vertebrales. Este crecimiento óseo en los márgenes de las articulaciones aparece comúnmente con el avance de la edad, y aunque son característicos de enfermedades articulares, como la osteoartrosis o e n f e r m e d a d e s degenerativas del disco intervertebral, su sola presencia no incrementa la prevalencia de dichas patologías. No obstante, este individuo también presentó una importante modificación del contorno del cuerpo vertebral, crecimiento óseo, eburnación y considerable porosidad en la superficie ósea en las vértebras cervicales, que hacen pensar en una enfermedad osteoartrósica activa (Figura 9). Aparte de estas modificaciones en la región cervical, en toda la columna se registra la reducción de la altura de los cuerpos vertebrales y un desvío de las espinas y los ejes de los cuerpos (séptima cervical y dorsales) hacia la izquierda, denotando una leve escoliosis.

Figura 8. Patologías traumáticas presentes en tórax del individuo 2: a) 10° costilla derecha con fractura soldada con desplazamiento; b) Callo óseo en el extremo distal de la 9° costilla izquierda.

Figura 9. Vértebras cervicales 3, 4 y 5 del individuo 2 (vista craneal y caudal). Se observan los contornos del cuerpo y las carillas modificados, osteofitos, eburnación y considerable porosidad.
A nivel bucal se registraron modificaciones fisiológicas y degenerativas asociadas con la edad y la dieta. Se observa un avanzado grado de desgaste dental considerando las edades de los individuos adultos, es decir, el tiempo de uso de los dientes anteriores (Smith 1984) y molares (Scott 1979; tabla 4). Dependiente de la elevada tasa de desgaste y del tipo de dieta consumida, no se registran caries. En cambio, sí se relevaron abscesos complejos y apicales en los individuos adultos, que serían producto de la infección pulpar debida a su exposición por el avanzado grado de desgaste. La consecuencia final de este proceso se evidencia en la elevada prevalencia de dientes perdidos antemortem registrada en el individuo 2 (Tabla 5), donde la mandíbula presenta casi la totalidad de los alvéolos reabsorbidos, con excepción de tres piezas que se hallan in situ y dos aisladas (Figura 10). Este caso puede relacionarse con el final de la vida funcional de las piezas dentales posteriores a partir de los 40 años de edad y de los dientes anteriores a partir de los 30 años en cazadores recolectores con elevada tasa de desgaste. El grado y velocidad de desgaste en los molares se debe principalmente al tipo de dieta y los procesos de abrasión y atrición involucrados en el acto de masticación, y en los dientes anteriores y premolares al mismo proceso combinado eventualmente con la utilización extramasticatoria de estas piezas dentales -como instrumento-.
Tabla 4. Medias del desgaste oclusal (Scott 1979; Smith 1984) de los individuos adultos.

Tabla 5. Modificaciones fisiológicas y patológicas de las cavidades bucales de los individuos adultos.

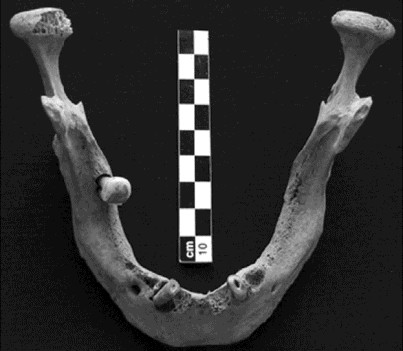
Figura 10. Mandíbula del individuo 2. Se aprecia la casi totalidad de pérdidas dentales antemortem, el desgaste completo de los caninos con exposición de la cavidad pulpar y la modificación del borde de la tabla alveolar (porosidad, abscesos complejos).
El Dr. Martín Fugassa (CONICET-UNMdP) ha realizado análisis paleoparasitológicos en muestras de sedimentos procedentes de la cavidad abdominal del individuo 2 y de otros sectores de la cueva para control. Los resultados de estos estudios apuntan a la presencia de parásitos en el esqueleto que pueden corresponderse con un parasitismo verdadero o infección patente de Trichuris sp., y con la presencia de parásitos zoonóticos (Capillaria sp.) en tránsito por el consumo de presas crudas parasitadas (guanaco) o por la percolación de unidades superiores -contaminación- (Fugassa y Barberena 2006).
Modificaciones tafonómicas
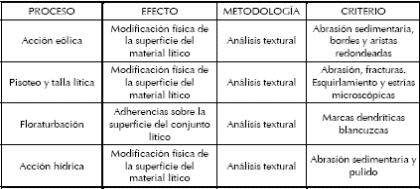
Se evaluaron diferentes modificaciones tafonómicas de naturaleza no antrópica en los elementos óseos humanos (Blumenschine et al. 1996, 2007; Domínguez-Rodrigo y Barba 2007; Lyman 1994, entre otros). Como es esperable para un entierro primario no diferido sin procesos posteriores de exposición, los huesos no presentan meteorización (Behrensmeyer 1978). En cambio, se registraron importantes frecuencias de otras modificaciones postdepositacionales tales como: daños mecánicos, precipitación de CaCo3 y manganeso, marcas de roedor y fragmentación postdepositacional.
El 65,67% de los huesos presenta tinción de manganeso, estando todos los esqueletos igualmente comprometidos (Tabla 6). Las superficies óseas se hallaron afectadas por una importante frecuencia de exfoliación cortical (56,38%) principalmente observable en los individuos subadultos, como también lo fueron las fracturas postdepositacionales (37,42%). Es llamativa la presencia de modificaciones mecánicas en las superficies corticales de los huesos (19,33%). Se considera que estos daños podrían haberse producido por el contacto de las superficies de los huesos con las aristas de las rocas que los cubrían, en forma combinada con la actividad de pisoteo en la superficie de la cueva (tanto por agentes antrópicos como naturales). Estos daños mecánicos se observan de forma variable, en su mayoria como 'pits' acompañados de estrías y fisuras radiales, y en ocasiones con hundimiento del tejido cortical (Figura 11). Se registraron, al igual que en los especímenes de fauna de la unidad X, rastros de CaCo3 en las superficies de los huesos acorde con la importante presencia de nódulos de carbonato en los sedimentos de las unidades VII, VI, V y II. La acción de roedores es relativamente baja (12,14%; Tabla 6), teniendo en cuenta la alta frecuencia de Ctenomys sp. intrusivos recuperados en la excavación (> 16300 especímenes; L'Heureux 2008) y en la superficie de la cueva.
Tabla 6. Modificaciones tafonómicas registradas en los elementos óseos humanos.
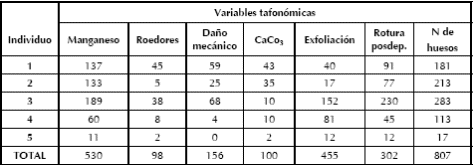

Figura 11. Fémur derecho del individuo 3. Se observa una serie de daños mecánicos sobre la diáfisis: perforaciones óseas con hundimiento del tejido compacto y fisuras radiales; pits irregulares más superficiales.
Isótopos estables y paleodieta
Se realizaron análisis de δ13C colágeno, δ15N y δ18O para cuatro esqueletos del sitio, uno de los cuales fue fechado en forma directa por 14c. El conjunto mortuorio de OB1 es un caso excepcional en el marco de los análisis isotópicos que hemos realizado hasta el momento ya que, como se ha mencionado, está compuesto por cinco individuos depositados simultáneamente. En forma independiente del tipo de vinculación social y/o biológica existente entre los mismos, esto sugiere que accedemos a la posibilidad
de evaluar la variación dietaria existente en algún nivel intra poblacional, definido en un sentido amplio y poco preciso. Análisis de ADN actualmente en marcha a cargo del Dr. Mauricio Moraga (Pontificia Universidad Católica de Chile) podrán contribuir con este tema. Para el conjunto total de muestras humanas analizado hasta el momento en el área -28 individuos- no puede defenderse una asociación de este tipo. El caso del sitio Cabo Vírgenes 17 representa un registro aparentemente diacrónico compuesto sólo por dos individuos adultos (L'Heureux et al. 2003). Otros casos procedentes de la costa y el interior del sector septentrional de Santa Cruz presentan una situación diferente, donde este tipo de variación es más fácilmente observable por la presencia de concentraciones mayores de individuos (Castro y Moreno 2000; Goñi y Barrientos 2004). Otro elemento de interés radica en la variación etaria que caracteriza a OB1, que representa un rango que va desde un individuo neonato a un adulto mayor. Este constituye el principal eje de variación que puede tratarse a partir de estos datos, ya que no se registró variación a nivel de sexo. Analizamos brevemente cada uno de estos cuatro casos (Tabla 7).
Tabla 7. Composición y resultados isotópicos del entierro múltiple de OB1.

Para el individuo 1 (adulto joven/masculino) se obtuvo un valor de δ13C colágeno de -18,1‰, que lo
ubica en el sector que se ha definido como umbral para la identificación del consumo de recursos marinos(Barberena 2008). Si se realiza una interpretación lineal del valor, implica un consumo de estos recursos en un nivel aproximado al 20% del total de la dieta, aunque dado que existen muestras de guanaco con valores de ca. -19 o -18‰, no se considera que este dato aislado pueda ser atribuido confiablemente al consumo de recursos marinos. Aquí es donde la información de δ15N se torna clave. El valor de 13,1‰ es elevado y concuerda con un consumo de alimentos del mar de la magnitud mencionada. Ante la aparente ausencia local de un enriquecimiento en los valores de δ15N en los herbívoros por aridez (Ambrose 1991, 2000; Dupras y Schwarcz 2001), un valor superior a 13 implicaría el consumo de recursos marinos en una proporción probablemente baja. El individuo 2 (adulto mayor/masculino) representa una variación dentro de la situación comentada para el individuo 1. El valor de δ13C colágeno se encuentra levemente empobrecido con respecto al umbral que se ha definido, lo cual sugeriría que no consumió recursos marinos. Sin embargo, el valor de δ15N es elevado (13,7‰ ), aún más que en el caso del individuo 1. Se enfatiza, entonces, que el valor de δ13C lo ubica en un terreno de probable consumo de recursos del mar en bajas proporciones y la información de δ15N lo respalda en forma independiente. El individuo 3 (8 ± 2 años) presenta valores consistentes que marcan que no hay consumo de recursos marinos: el valor de δ13C se ubica claramente empobrecido con respecto al umbral y es típico para las muestras terrestres en Patagonia meridional (Barberena 2002), mientras que el valor de δ15N es bajo y tampoco permite defender el consumo de recursos del mar. Esto es interesante dado que marca una diferencia con respecto a los individuos 1 y 2, que son adultos. Este posible eje etario de variación paleodietaria no puede ser desarrollado a partir de este único caso, aunque sí puede servir como base para una hipótesis contrastable a futuro: los recursos marinos son consumidos en forma sistemática a partirde una cierta edad mínima. El individuo 4 (6 ± 3 meses) es lactante, por lo que se encuentra un nivel trófico por sobre su madre (Dupras et al. 2001; Katzenberg et al. 1996). Esto permite descartar el consumo de recursos marinos como explicación para el valor elevado de δ15N. Otra observación que puede realizarse es que la madre de este individuo tendría valores de δ15N considerablemente más bajos que los dos individuos adultos de OB1 y presentaría valores isotópicos interpretables como de dieta 'terrestre'.
DISCUSIÓN Y CONCLUSIONES
Las evidencias recuperadas en el sitio OB1 contribuyen a dos temas generales de discusión que están siendo tratados en el Proyecto Magallania, dirigido por el Dr. Luis Borrero, en referencia con el Holoceno tardío en Patagonia meridional: a) la organización geográfica de las poblaciones humanas, en particular las pautas de interacción establecidas entre las costas marinas y el interior del continente, y b) las prácticas mortuorias.
Las evidencias isotópicas de este sitio marcan la existencia de variabilidad dietaria en una escala sincrónica y para dos casos puede defenderse el consumo relativamente usual de recursos marinos. Esto implicaría alguna forma de contacto con la costa del estrecho de Magallanes, ubicada 17 km hacia el sur, o con la costa atlántica, unos 60 km hacia el este (distancias tomadas en línea recta). Distintas evidencias llevan a sugerir que esta interacción se estableció principalmente hacia el sur, con la costa del Estrecho, que habría ejercido un atractivo más fuerte para las poblaciones humanas (Barberena 2008; Borrero y Barberena 2006). Dado que OB1 se localiza dentro de una franja espacial pericostera que se defiende como vinculada con el mar, las evidencias isotópicas que indican el consumo de recursos marinos son concordantes. Por otra parte, en este sitio también se han recuperado numerosos especímenes de fauna marina y costera, incluyendo diferentes especies de moluscos y de aves (L'Heureux 2008).
A nivel de las prácticas mortuorias, las evidencias presentadas marcan un registro relativamente anómalo o poco abundante en el registro de Patagonia meridional. El entierro estudiado constituye una estructura en la cual se inhumaron cinco inviduos de distintas edades en forma conjunta. El tratamiento mortuorio incluyó la excavación de un foso, la generación de una discontinua superficie artificial de apoyo con vegetales, el uso de ocre y el sepultamiento con bloques de roca de caja directamente sobre los cuerpos, por encima de los cuales se depositó parte de los sedimentos removidos. De este modo, se generó una unidad enriquecida en huesos de guanaco y otros materiales por sobre el entierro con una estructura clasto sostén asimilable a un 'relleno', cuya cronología no se discrimina de la de los esqueletos. Las dos unidades estratigráficas implicadas (X y XI) son interpretadas como artificiales (sensu Gargett 1989).
Los datos presentados por Guerra de Fretes (1977) para el sitio son limitados y poco precisos, aunque su integración con la información presentada aporta elementos interesantes. La autora informa que realizó una excavación en el sector central de la cueva y recuperó restos humanos correspondientes a dos individuos: "Los dos cuerpos se hallaban en posición de cúbito dorsal derecho y fetal, alineado uno al lado del otro con las cabezas orientadas hacia el oeste y las caras hacia el norte, de frente al exterior" (Guerra de Fretes 1977: 102). Aparentemente los mismos habrían sido encontrados por debajo de bloques de la roca de caja y sedimentos, aunque no puede realizarse una interpretación adecuada del contexto. A partir de nuestro trabajo resulta claro que ambos conjuntos no constituían una misma inhumación, dado que el pozo realizado por Guerra de Fretes se encuentra alejado del entierro excavado (Figura 3). A su vez, durante nuestra excavación se expusieron los límites de la estructura de entierro, lo que permite defender que la misma no se extendía, en forma continua al menos, hasta el sector modificado por Guerra de Fretes. Estos datos indican que el sitio OB1 fue utilizado en más de una ocasión para la depositación formal de restos humanos. La presencia de huesos humanos en el relleno por sobre la estructura de entierro también puede apuntar en este sentido. Sin embargo, el sitio no fue ocupado en forma exclusiva para la realización de actividades mortuorias, ya que se registraron diferentes tipos de actividades, que incluyen lentes de fogones, descarte de abundantes valvas marinas y formatización de instrumentos líticos (Franco et al. 2008).
Otro aspecto que interesa resaltar del análisis mortuorio es el carácter de depositación simultánea de los cinco esqueletos recuperados en la estructura. La gran mayoría de los restos humanos del CVPA y los espacios periféricos que lo rodean proceden de eventos inhumatorios individuales o, en ciertos casos, compuestos por dos individuos (Bird 1988; Guichón et al. 2001; L'Heureux et al. 2003; Martin 2006; Prieto 1993-1994). Hay sitios que presentan numerosos esqueletos humanos, pero para la mayoría de ellos no se puede defender una depositación simultánea, que en el caso de entierros primarios implicaría una coocurrencia en la muerte de varios individuos. Se ha señalado que en estos espacios de Patagonia meridional hay tres casos defendibles de inhumación de numerosos individuos en un único evento: Cerro Sota (Bird 1988), Cañadón Leona 5 (Bird 1988) y OB1, que constituyen un número total de 20 esqueletos (Barberena 2008; Barberena y Borrero 2007). Es particularmente llamativo que OB1 y Cerro Sota están fechados en torno a 3500 años AP., mientras que el contexto de Cañadón Leona sugiere que puede tener una cronología comparable (Hedges et al. 1992; Prieto et al. 1998). Por lo tanto, estos únicos casos de eventos de múltiples muertes asociadas se ubicarían en torno a dicha cronología.
En el marco de baja demografía esperable para estas sociedades de Patagonia meridional, los eventos de mortandad interceptados por estas muestras adquieren un importante significado poblacional. El análisis de las causas de muerte se torna en un objetivo central a partir de aquí, y las enfermedades o situaciones de violencia son las principales hipótesis a tener en cuenta. En el caso de OB1, las evidencias bioarqueológicas presentadas no permiten identificar causas específicas de muerte, aunque un estudio detallado en este sentido debe realizarse aún con los conjuntos de los otros sitios referidos (Cerro Sota y Cañadón Leona). De todos modos, se debe señalar que el 60% de los individuos presentes en OB1 (N° 2, 4 y 5) representan las edades más susceptibles de sufrir consecuencias mortales frente a distintos tipos de eventos externos (pe. fluctuaciones en el clima y/o disponibilidad de recursos).
Por último, se considera que las evidencias presentadas para el sitio OB1 contribuyen a integrar la información sobre prácticas mortuorias con el estudio de ciertos aspectos de la dinámica poblacional humana en Patagonia meridional.Un total de 752 especímenes (44% del total de especímenes identificados de guanaco y de astillas no identificadas) presenta algún tipo de marca o combinación de ellas distribuidas de la siguiente manera: 355 con marcas de origen exclusivamente natural (carnívoros y/o roedores), 123 con huellas sólo de procesamiento antrópico, 144 con marcas naturales y huellas de procesamiento antrópico y 110 con manchones de carbón y de ocre.
Agradecimientos
Este trabajo se desarrolló en el marco del Proyecto Magallania, dirigido por el Dr. Luis A. Borrero y subvencionado por las siguientes instituciones: CONICET (PIP 2390, PIP 5676), ANPCyT (PICT 04- 9498), National Geographic Society (Grant 7736-04) y la Facultad de Filosofía y Letras de la Universidad de Buenos Aires (UBACyT F133). Agradecemos a nuestros compañeros de equipo por su gran ayuda durante el trabajo de campo y laboratorio. A Aníbal Cufré y Antonio Zaldívar, el 'maestro', por la enorme ayuda brindada durante nuestra estadía en la estancia Monte Aymond. Al Dr. Esteban Perazzo y la Compañía de Tierras del Sud por su colaboración. Igualmente, queremos agradecer a Jorge Suby y Mauricio Massone por las modificaciones sugeridas, que sirvieron para mejorar nuestro trabajo.
1 También hay una mínima referencia sobre la presencia y aparente trabajo del sacerdote M. J. Molina en el sitio (Molina 1972), aunque sin detalles publicados.
REFERENCIAS CITADAS
Ambrose, S. H. 1991 Effects of diet, climate and physiology on nitrogen isotope abundances in terrestrial foodwebs. Journal of Archaeological Science 18 (3): 293-317. [ Links ]
Ambrose, S. H. 2000 Controlled Diet and Climate Experiments on Nitrogen Isotope Ratios of Rats. En Biogeochemical Approaches to Paleodietary Analysis, editado por S. H. Ambrose y M. A. Katzenberg, pp. 243-267. Advances in Archaeological and Museum Science 5.Kluwer Academics/Plenum Press, Nueva York. [ Links ]
Barberena, R. 2002 Los límites del mar. Isótopos estables en Patagonia meridional. Sociedad Argentina de Antropología, Buenos Aires. [ Links ]
Barberena, R. 2008 Arqueología y biogeografía humana en Patagonia meridional. Sociedad Argentina de Antropología, Buenos Aires. [ Links ]
Barberena, R. y L. A. Borrero 2007 Aspectos geográficos y temporales en el registro arqueológico del Holoceno tardío en Pali Aike. Trabajo presentado en el XVI Congreso Nacional de Arqueología Argentina, San Salvador de Jujuy. MS. [ Links ]
Barberena, R., A. Blasi y C. Castiñeira 2006 Geoarqueología en cuevas: el sitio Orejas de Burro 1 (Pali Aike, Argentina). Magallania 34 (1): 119-138. [ Links ]
Behrensmeyer, A. K.1978 Taphonomic and ecologic information from bone weathering. Paleobiology 4: 150-162. [ Links ]
Bird, J. 1988 Travels and archaeology in South Chile. Editado por J. Hyslop. University of Iowa Press, Iowa. [ Links ]
Blumenschine, R. J., C. W. Marean y S. D. Capaldo 1996 Blind test of inter-analyst correspondence and accuracy in the identification of cut marks, percussion marks, and carnivore tooth marks on bone surfaces. Journal of Archaeological Science 23: 493-507. [ Links ]
Blumenschine, R. J., K. A. Prassack, C. D. Kreger y M. C. Pante 2007 Carnivore tooth-marks, microbial bioerosion, and the invalidation of Dominguez-Rodrigo and Barba's (2006) test of Olduvai hominid scavenging behavior. Journal of Human Evolution 53: 420-426. [ Links ]
Borrero, L. A. y R. Barberena 2006 Hunter-Gatherer Home Ranges and Marine Resources. An Archaeological Case from Southern Patagonia. Current Anthropology 47 (5): 855-867. [ Links ]
Buikstra, J. y D. Ubelaker (editores) 1994 Standards for data collection from human skeletal remains. Arkansas Archaeological Survey Research Series 44, Nueva York. [ Links ]
Castro, A. S. y J. E. Moreno 2000 Noticia sobre enterratorios humanos en la costa norte de Santa Cruz, Patagonia argentina. Anales del Instituto de la Patagonia (Serie Ciencias Humanas) 28: 225-231. [ Links ]
Clarke, N. G. y R. S. Hirsch 1991 Tooth dislocation: The relationship with tooth wear and dental abscesses. American Journal of Physical Anthropology 85: 293-298. [ Links ]
Corbella, H. 1999 Dataciones radimétricas en Pali Aike, Patagonia austral. Actas del XIV Congreso Geológico Argentino II: 265-268. Salta. [ Links ]
Domínguez-Rodrigo, M. y R. Barba 2007 Five more arguments to invalidate the passive scavenging version of the carnivore-hominidcarnivore model: a reply to Blumenschine et al. (2007). Journal of Human Evolution 53: 427-433. [ Links ]
Dupras, T. L. y H. P. Schwarcz 2001 Strangers in a Strange Land: Stable Isotope Evidence for Human Migration in the Dakhleh Oasis, Egypt. Journal of Archaeological Science 28 (11): 1199-1208. [ Links ]
Dupras, T. L., H. P. Schwarcz y S. I. Fairgrieve 2001 Infant Feeding and Weaning Practices in Roman Egypt. American Journal of Physical Anthropology 115: 204-212. [ Links ]
Franco, N. V., G. L. L'Heureux, J. E. Charlin y R. Barberena 2008 El sitio Orejas de Burro 1 y la arqueología del campo volcánico Pali Aike. MS. [ Links ]
Fugassa, M. y R. Barberena 2006 Cuevas y zoonosis antiguas: paleopatología del sitio Orejas de Burro 1 (Santa Cruz, Argentina). Magallania 34 (2): 57-62. [ Links ]
Fully, G. 1956 Une nouvelle méthode de détermination de la taille. Annales de Médicine Légale 35: 266-273. [ Links ]
Gargett, R. H. 1989 Grave Shortcomings. The Evidence for Neanderthal Burial. Current Anthropology 30 (2): 157-190. [ Links ]
Goñi, R. A. y G. Barrientos 2004 Poblamiento tardío y movilidad en la cuenca del Lago Salitroso. En Contra viento y marea. Arqueología de Patagonia, compilado por M. T. Civalero, P. M. Fernández y A. G. Guráieb, pp. 313- 324. INAPL-SAA, Buenos Aires. [ Links ]
Guerra de Fretes, E. 1977 Informe preliminar del hallazgo de restos humanos en una de las cuevas del paraje denominado "Monte Aymond". Actas y Memorias del IV Congreso Nacional de Arqueología Argentina, Revista del Museo de Historia Natural de San Rafael, IV (1/4): 101-102. San Rafael, Mendoza. [ Links ]
Guichón, R. A., R. Barberena y L. A. Borrero 2001 ¿Dónde y cómo aparecen los restos óseos humanos en Patagonia Austral? Anales del Instituto de la Patagonia (Serie Ciencias Humanas) 29: 103-118. [ Links ]
Hedges, R. E. M., R. Housley, C. Bronk y G. Van Klinken 1992 Radiocarbon Dates from the Oxford AMS System: Archaeometry Datelist 15. Archaeometry 34: 337-357. [ Links ]
Hillson, S. 1996 Dental anthropology. Cambridge University Press, Cambridge. [ Links ]
Katzenberg, M. A., D. A. Herring y S. R. Saunders 1996 Weaning and Infant Mortality: Evaluating the Skeletal Evidence. Yearbook of Physical Anthropology 39: 177-199. [ Links ]
Larsen, C. S. 1997 Bioarchaeology: Interpreting behavior from the human skeleton. Cambridge University Press, Nueva York. [ Links ]
L'Heureux, G. L. 2008 La arqueofauna del Campo Volcánico Pali Aike. El sitio Orejas de Burro 1, Santa Cruz, Argentina. Magallania 36. En prensa. [ Links ]
L'Heureux, G. L, R. Guichón, R. Barberena y L. A. Borrero 2003 Durmiendo bajo el faro. Hallazgo de un entierro humano en Cabo Vírgenes (C.V.17), Provincia de Santa Cruz, República Argentina. Intersecciones en Antropología 4: 87-97. [ Links ]
Lovejoy, O., R. Meindl, T. Pryzbeck y R. Mensforth 1985 Chronological methamorphosis of the auricular surface of the ilium: A new method for the determination of adult skeletal age at death. American Journal of Physical Anthropology 68: 15- 28. [ Links ]
Lyman, R. L. 1994 Quantitative units and terminology in zooarchaeology. American Antiquity 59: 36-71. [ Links ]
Martin, F. M. 2006 Carnívoros y huesos humanos de Fuego- Patagonia. Aportes desde la tafonomía forense. Sociedad Argentina de Antropología, Buenos Aires. [ Links ]
Meindl, R. S. y C. O. Lovejoy 1985 Ectocranial suture closure: a revised method for the determination of skeletal age at death based on the lateral anterior sutures. American Journal of Physical Anthropology 68: 57-66. [ Links ]
Mena, F., O. B. Reyes, T. W. Stafford Jr. y J. Southon 2003 Early human remains from Baño Nuevo 1 cave, central Patagonian Andes, Chile. Quaternary International 109-110: 113-121. [ Links ]
Mensforth, R. 1991 Paleoepidemiology of porotic hyperostosis in the Libben and BT-5 skeletal populations. Kirtlandia 46: 1-47. [ Links ]
Molina, M. J. 1972 Nuevos aportes para el estudio del arte rupestre patagónico. Anales de la Universidad de la Patagonia San Juan Bosco 4 (1/1): 62-182. [ Links ]
Pettitt, P. B. 2002 The Neanderthal dead: exploring mortuary variability in Middle Palaeolithic Eurasia. Before Farming 4: 62-87. [ Links ]
Behrensmeyer, A. K.1991 Terrestrial Vertebrate Accumulations. En Taphonomy: Releasing the Data Locked in the Fossil Record, editado por P.A. Allison y D.E.G. Briggs, pp. 291-335, Plenum Press, New York. [ Links ]
Prieto, A. 1993-1994 Algunos datos en torno a los enterratorios de la región de Magallanes. Anales del Instituto de la Patagonia (Serie Ciencias Humanas) 22: 91-100. [ Links ]
Binford, L. 1978 Nunamiut Ethnoarchaeology. Academic Press, New York. [ Links ]
Prieto, A., F. Morello, R. Cárdenas y M. Christensen 1998 Cañadón Leona: a sesenta años de su descubrimiento. Anales del Instituto de la Patagonia (Serie Ciencias Humanas) 26: 83-105. [ Links ]
Rogers, J. y T. Waldrom 1995 A field guide to joint disease in archaeology. John Wiley & Sons, Chichester. [ Links ]
Schutkowski, H. 1993 Sex determination on infant and juvenile skeletons: I. Morphognostic features. American Journal of Physical Anthropology 90: 199-205. [ Links ]
Scott, E. C. 1979 Dental wear scoring technique. American Journal of Physical Anthropology 51: 213-218. [ Links ]
Smith, B. H. 1984 Patterns of molar wear in hunter-gatherers and agriculturalists. American Journal of Physical Anthropology 63: 39-56. [ Links ]
Ubelaker, D. 1989 Human Skeletal Remains. Segunda edición. Taraxacum Press, Washington D. C. [ Links ]
Weaber, D. S. 1980 Sex differences in the ilia of a know sex and age sample of fetal and infant skeletons. American Journal of Physical Anthropology 52: 191-195. [ Links ]

