










Servicios Personalizados
Revista

Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkIntersecciones en antropología
versión On-line ISSN 1850-373X
Intersecciones antropol. n.9 Olavarría ene./dic. 2008
Nuevas investigaciones sobre la ocupación prehispánica de la costa pampeana en el Holoceno tardío: el sitio arqueológico Claromecó 1 (partido de Tres Arroyos, provincia de Buenos Aires)
Mariano Bonomo, Diego Catriel Leon, Lucas Turnes y Eduardo Apolinaire
Mariano Bonomo. CONICET. Dpto. Científico de Arqueología, Facultad de Ciencia Naturales y Museo (UNLP). Paseo del Bosque S/Nº, 1900, La Plata, Buenos Aires, Argentina. E-mail: mbonomo@fcnym.unlp.edu.ar
Diego Catriel Leon. ANPCyT. INCUAPA, Facultad de Ciencias Sociales, UNCPBA. Avda. del Valle 5737, Olavarría, Buenos Aires, Argentina. E-mail: cleon@fcnym.unlp.edu.ar
Lucas Turnes. Dpto. Científico de Arqueología, Facultad de Ciencia Naturales y Museo, UNLP. Paseo del Bosque s/Nº, La Plata, Buenos Aires, Argentina. E-mail: mardelpela@hotmail.com
Eduardo Apolinaire. Dpto. Científico de Arqueología, Facultad de Ciencia Naturales y Museo, UNLP. Paseo del Bosque s/Nº, La Plata, Buenos Aires, Argentina. E-mail: eapolinaire@hotmail.com
Recibido 5 de Diciembre 2007.
Aceptado 12 de Abril 2008
RESUMEN
Los materiales arqueológicos disponibles para entender la ocupación holocénica del litoral marítimo pampeano están constituidos casi exclusivamente por artefactos líticos depositados en la superficie de la faja de médanos. En tal situación, el sitio Claromecó 1 (CL1) se destaca por su posición en estratigrafía y la preservación de restos óseos que permitió establecer su cronología. En este artículo se presentan los resultados del análisis de los materiales líticos, cerámicos y faunísticos allí recuperados. A partir de los distintos estudios realizados, se aborda la historia tafonómica, la estacionalidad, la explotación faunística, y la tecnología lítica y cerámica. Los resultados del análisis lítico indican que se han tallado mediante la técnica bipolar elevadas proporciones de rodados costeros. Estas características de los recursos líticos utilizados diferencian a este conjunto de la mayoría de los sitios de la llanura pampeana. Aun así, al igual que en los sitios del interior, los estudios faunísticos señalan que en CL1 se aprovecharon animales de origen continental. Por último, la información obtenida muestra que CL1 fue un campamento ocupado por cazadores-recolectores prehispánicos durante el Holoceno tardío.
Palabras clave: Cazadores-recolectores; Litoral marítimo; Holoceno tardío; Análisis lítico; Cerámico y arqueofaunístico.
ABSTRACT
New Research On The Prehispanic Occupation Of The Pampean Coast During The Late Holocene: Claromecó 1 Archaeological Site (Tres Arroyos, Buenos Aires Province.) . The archaeological materials available for understanding the Holocene human occupation of the Pampean coast consist almost exclusively of lithic artifacts deposited on the surface of dune strips. In this context Claromecó 1 (CL1) archaeological site stands out because of its stratigraphic position and the preservation of bone material, which allowed us to establish a chronology. In this paper, the results obtained from the lithic, ceramic and faunal analyses carried out at the site are presented. Lithic and pottery technology, taphonomical history, stationality, and faunal exploitaition were all examined using these different approaches. The results of the lithic analysis suggest that a high proportion of coastal pebbles were knapped using the bipolar technique. The characteristics of the lithic resources used at CL1 distinguish this assemblage from most of the sites on the Pampean plains. However, as with sites in the interior, the faunal research shows that in CL1 the origin of the exploited fauna was continental. Finally, the information obtained indicates that CL1 was a campsite occupied by pre- Hispanic hunter-gatherers during the late Holocene.
Keywords: Hunter-gatherers; Maritime littoral; Late Holocene; Lithic; Pottery and faunal analyses.
INTRODUCCIÓN
Las evidencias materiales de la ocupación prehispánica del litoral marítimo del área Interserrana bonaerense están constituidas por grandes cantidades de artefactos líticos depositados en la faja de médanos sin contextos estratigráficos ni cronológicos precisos. De acuerdo a los datos disponibles, estos sitios fueron caracterizados como talleres en los que la actividad principal fue la reducción bipolar de rodados costeros para la obtención de lascas destinadas a la manufactura de instrumentos (Bonomo 2005). Estos conjuntos costeros, generados por cazadores-recolectores del Holoceno medio y tardío, formaron parte de un sistema de asentamiento mayor que incluyó a la llanura y sistemas serranos del interior pampeano.
En el marco de las investigaciones llevadas a cabo en la costa atlántica desde 1999 (Bonomo 2005), se han prospectado y sondeado ambas márgenes del curso inferior del arroyo Claromecó y se ha relevado la cadena de médanos adyacente a este curso fluvial. Como producto de estas actividades se han detectado doce contextos con materiales arqueológicos, entre los que sobresale Claromecó 1 (CL1). Este sitio localizado en un ambiente transicional entre la faja de médanos y la llanura contigua se destaca por su posición en estratigrafía y la preservación de restos óseos que permitieron establecer su cronología absoluta.
En este artículo se presenta un fechado radiocarbónico obtenido para el sitio, los resultados del análisis tecnológico y morfológico de los artefactos líticos y la alfarería y del estudio taxonómico, anatómico y tafonómico del material faunístico allí recuperado. Se pone especial énfasis en un tipo de evidencia particular, las cáscaras de huevo de ñandú (Rehidae), para cuyo examen se proponen lineamentos analíticos específicos que permiten ampliar la información sobre el aprovechamiento de estas aves en el pasado.
SITIO ARQUEOLÓGICO CLAROMECÓ 1
La costa atlántica que se extiende al este y el oeste del arroyo Claromecó está caracterizada por plataformas rocosas discontinuas que interrumpen playas arenosas amplias afectadas por mareas de 2 m de amplitud media, dunas elevadas paralelas a la línea de ribera con vegetación psammófita y remanentes aislados de barrancas. La cuenca del arroyo Claromecó tiene una superficie de 2700 km2 (Isla y Espinosa 1998). Este curso desemboca en el océano atravesando la cadena de medanos litorales y exhibe un cauce con meandros encajonados producto de la erosión lateral de su caudal.
El sitio arqueológico Claromecó 1 (38º 50′21,8″ S; 60º 5′20,8″ O) está localizado sobre la margen izquierda del arroyo homónimo y a una distancia de 3 km de la costa atlántica (Figura 1). Se ubica contiguo al camino del Paseo de las Cascadas que bordea al arroyo Claromecó, en las inmediaciones de la cuarta cascada. A lo largo de este camino de tierra fueron hallados en superficie o semi-enterrados 136 artefactos líticos y restos faunísticos, parte de los cuales pudieron asignarse a guanaco (Lama guanicoe).

Figura 1. Imagen satelital con la ubicación del sitio Claromecó 1 y de los contextos arqueológicos mencionados en el texto.
En los años 2004 y 2005 se realizaron dos excavaciones en el sector adyacente al camino donde se detectó la mayor concentración de materiales arqueológicos en posición superficial. En el 2004 se excavaron 5 cuadrículas en el espacio comprendido entre el camino y el alambrado que delimita la chacra experimental integrada Barrow (INTA). En el 2005 este espacio fue eliminado por las máquinas que ampliaron el camino. Por tal motivo, hubo que plantear 16 nuevas cuadrículas dentro de la chacra experimental (Figura 2). Las dimensiones de las cuadrículas fueron de 1 x 1 m, totalizando una superficie abierta de 21 m2. Se excavó siguiendo niveles artificiales de 5 cm de espesor y el sedimento fue tamizado utilizando zarandas secas de 2 x 2 mm de malla en las cuadrículas 1 a 5 y de 1 x 3 mm para las cuadrículas 6 a 21.

Figura 2. Planta de la excavación de CL1 y perfiles con las unidades sedimentarias registradas.
En CL1 se recuperaron 423 artefactos líticos, 2 pigmentos minerales de color rojo, 87 tiestos cerámicos, 14 fragmentos de loza, 6 fragmentos de vidrio, 4494 restos faunísticos y 8 ecofactos (6 rodados costeros y 2 clastos de xilópalo sin modificación antrópica). Estos materiales se distribuyen principalmente en dos unidades sedimentarias, entre los 10 y 90 cm de profundidad desde el nivel 0 de la excavación ubicado en la superficie del terreno.
La unidad sedimentaria superior es un depósito eólico edafizado que se extiende desde el nivel 0 hasta los 50 o 65 cm de profundidad (dependiendo de la pendiente del terreno). El pH en esta unidad es de 8,08 y texturalmente es un sedimento franco-arenoso (Favier Dubois 2007). Contiene principalmente materiales posthispánicos (vidrio, loza y restos óseos de fauna doméstica -Bos taurus, Equs caballus y Ovis aries-). Los restos faunísticos que se agrupan en la parte superior de esta unidad no presentan meteorización ni otras modificaciones de importancia (i.e., sólo escaso registro de marcas de raíces). En esta unidad, aunque en menor medida, se presentan restos de fauna autóctona así como escasos materiales líticos y cerámicos restringidos a la base del depósito. En los especímenes faunísticos de la base se reconocieron modificaciones en las superficies óseas (grabados intensos por raíces, hoyos de disolución química, abrasión, termoalteración) similares a las registradas en la unidad subyacente, pero a diferencia de ésta, los hoyos de disolución son más acentuados.
La unidad sedimentaria subyacente es un paleosuelo erosionado que abarca desde los 50/65 a los 90 cm de profundidad. El pH registrado es de 8,16 y el sedimento es franco-arcillo-arenoso a francoarenosos (Favier Dubois 2007). En esta unidad, además, se hallaron gasterópodos dulceacuícolas y terrestres asociados a entornos acuáticos (Biomphalaria peregrina, Heleobia parchappii, Succinea meridionalis) (Simone 2006; Pamela Steffan, comunicación personal 2007). De acuerdo a las características de los gasterópodos y al análisis geoarqueológico se pudo determinar que el ambiente de depositación fue una planicie de inundación distal (Favier Dubois 2007; Pamela Steffan, comunicación personal 2007). En este paleosuelo se recuperó la gran mayoría del material prehispánico que es presentado en este artículo. Además, se registraron dos especímenes de fauna introducida en el techo de esta unidad (Figuras 3 y 4).
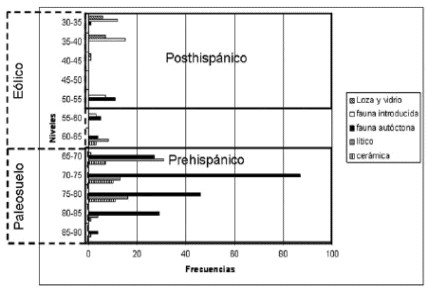
Figura 3. Distribución vertical de los materiales arqueológicos de CL1 (cuadrículas 6, 7, 14 y 15).
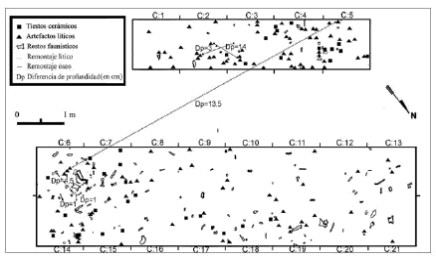
Figura 4. Distribución horizontal de los materiales arqueológicos registrados en el paleosuelo.
A los efectos de establecer la cronología de la ocupación del paleosuelo se efectuó un fechado ![]() por AMS en el Laboratory of Isotope Geochemistry, Department of Geosciences de Tucson, Arizona. Se procesó un desecho helicoidal sobre diáfisis de hueso largo de mamífero (pieza nº 5-4-4) que arrojó una edad de 800 ± 34 años AP y un δ C13 de -21,2 (AA- 64621). Esta datación calibrada en años calendarios con 2δ (95% de probabilidad) corresponde al lapso entre 1283 y 1165 DC. La fecha radiocarbónica obtenida es semejante a las de los eventos pedogenéticos registrados a 1,8 km de la desembocadura del arroyo Claromecó (840 AP; Isla y Espinosa 1998) y en el río Sauce Grande (995 AP; Rabassa 1989).
por AMS en el Laboratory of Isotope Geochemistry, Department of Geosciences de Tucson, Arizona. Se procesó un desecho helicoidal sobre diáfisis de hueso largo de mamífero (pieza nº 5-4-4) que arrojó una edad de 800 ± 34 años AP y un δ C13 de -21,2 (AA- 64621). Esta datación calibrada en años calendarios con 2δ (95% de probabilidad) corresponde al lapso entre 1283 y 1165 DC. La fecha radiocarbónica obtenida es semejante a las de los eventos pedogenéticos registrados a 1,8 km de la desembocadura del arroyo Claromecó (840 AP; Isla y Espinosa 1998) y en el río Sauce Grande (995 AP; Rabassa 1989).
ANÁLISIS DE LOS ARTEFACTOS LÍTICOS
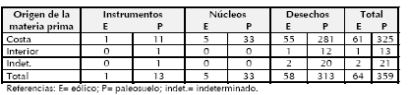
El estudio tecno-morfológico de los materiales líticos se efectuó siguiendo los lineamientos de Aschero (1975, 1983), considerando las modificaciones propuestas por Bonomo (2005) para el análisis de los productos bipolares. Con respecto a la muestra recuperada en el depósito eólico, predominan los desechos de talla y los núcleos bipolares en rodados costeros (Tabla 1). Además, fue hallada una base recta de una punta de proyectil apedunculada pequeña sobre basalto. La mayoría de estos materiales (75%) se encuentra en la base de la unidad y no presenta diferencias claras con los registrados en el paleosuelo subyacente, en cuanto a las proporciones de las diferentes materias primas, las categorías generales de artefactos, los tamaños y las técnicas de talla representadas. Por esta razón se considera probable que las muestras líticas de ambas unidadescorrespondan al mismo componente arqueológico. Sin embargo, dado que se han observado distinciones en el conjunto óseo (véase más adelante), no se incluirán las piezas halladas en el depósito eólico en el estudio de los materiales del paleosuelo.
Tabla 1. Categorías generales de artefactos líticos según la procedencia de las materias primas.
Los instrumentos líticos hallados en CL1 son escasos y la mayoría están fragmentados (n= 9). Las piezas enteras poseen tamaños pequeño y mediano pequeño y módulos longitud-anchura mediano normal y corto ancho. Entre los instrumentos no tallados, se recuperó un fragmento de artefacto de molienda pasivo simple elaborado sobre arenisca. El mismo muestra evidencias de alisado, como rastros macroscópicos de uso, en su superficie activa plana (Alejandra Matarrese, comunicación personal 2007).
Entre los tallados se destacan dos preformas de puntas y tres puntas de proyectil fragmentadas (bases o ápices con limbo) elaboradas a partir de lascas y de la formatización del cuerpo central de rodados de basalto (Figura 5). Las puntas de proyectil son triangulares pequeñas apedunculadas con bases rectas o cóncavas, manufacturadas mediante microrretoques y retoques extendidos. Los restantes instrumentos tallados son unifaciales y presentan un menor grado de modificación. Están constituidos por tres raspadores frontales cortos y perimetral, dos filos bisel asimétrico laterales largos, una raedera lateral y un artefacto de formatización sumaria de sílice, ftanita, basalto y ortocuarcita. Las formas-base son cuerpos centrales y lascas a las que se les aplicaron principalmente retoques y microrretoques marginales. A diferencia de las puntas bifaciales, que incluyeron en su secuencia de producción la elaboración de preformas y la formatización de sus caras, en estos instrumentos unifaciales se ha pasado directamente de la extracción de lascas al retoque de sus bordes para conformar un filo.

Figura 5. Puntas y preformas de basalto de CL1.
El registro de núcleos muestra la reducción de nódulos en el sitio. Los núcleos son todos bipolares y están elaborados sobre rodados costeros. Sin tomar en cuenta los núcleos fragmentados (n= 6), entre los que se registraron accidentes de talla paralelos u oblicuos al eje mayor de los rodados, se observa que en las piezas enteras predominan los tamaños mediano pequeño (51,8%) y mediano grande (33,3%) y los módulos longitud-anchura mediano alargado (40,7%) y mediano normal (33,3%). Si bien algunos rodados sólo poseen pequeños negativos de lascado en uno de los extremos, en la mayoría se efectuaron extracciones en los dos polos (58%) y en ambas caras de los nódulos (57,6%) evidenciando un grado de reducción importante.
Los desechos de talla están constituidos por lascas fracturadas con talón (41%) y sin talón (28,7%), desechos no clasificables (24,3%), lascas enteras (3,5%) y desechos indiferenciados (2,5%). La tendencia del conjunto lítico muestra que la reducción de las rocas estuvo orientada a la producción de lascas, aunque también han sido utilizados núcleos bipolares como formas-base de instrumentos. Las lascas enteras son principalmente de tamaño pequeño y módulos longitud-anchura mediano alargado y mediano normal. En concordancia con la alta frecuencia de rodados, están más representadas las lascas externas - secundarias (26,7%), primarias (25,3%), bipolares con corteza (17,6%) y de dorso natural (3,9%)- que las internas -de arista (12,6%), planas (6,1%), angulares (5,2%) y bipolares sin corteza 2,6%). En relación con esta abundancia de rodados y su reducción mediante la técnica bipolar, entre las lascas predominan aquellas con atributos vinculados con este procedimiento de talla, como los talones astillados (45%) y los bulbos indiferenciados (63,5%).
ANÁLISIS DE LA ALFARERÍA
La alfarería fue analizada teniendo en cuenta sus atributos morfológicos y tecnológicos macroscópicos siguiendo las propuestas metodológicas de Rye (1981), Rice (1987) y Orton et al. (1997). Debido a que gran parte de los tiestos del sitio (n= 87) tiene un tamaño menor a 1 cm2 y a que se observaron indicios de erosión en un 73,5% de los mismos, fue estudiada una muestra de 45 fragmentos. Los rangos de tamaño de los fragmentos de la muestra seleccionada corresponden a: 1,1-2 cm2 (57,5%), 2,1-3 cm2 (27,8%), 3,1-4 cm2 (12,9%) y 4,1-5 cm2 (1,8%). Un 26,6% presenta adherencias.
Las partes de las vasijas representadas son el cuerpo (84,4%) y el borde (15,6%). Las pastas son de tipo laminar y poseen un porcentaje menor de antiplástico con relación a la matriz. En los antiplásticos predomina el tamaño de grano medio (66,6%), seguido por el fino (24,4%), el grueso (2,3%) y el indeterminado (6,7%). Con respecto a la cocción, la más abundante es la reductora (53,3%), seguida por oxidante incompleta (31,2%), oxidante (8,8%) e indeterminada (6,7%). El acabado de la superficie con mayor frecuencia es el engobe (48,9%), seguido por el alisado (37,8%), raspado (8,8%) e indeterminado (4,5%). La técnica empleada para la elaboración fue el rodete, mientras que la decoración no fue muy frecuente, ya que sólo un tiesto muestra pintura roja (2,2%). Por último, la relación borde/labio está representada en el sitio por tres categorías (directa/plana, directa/redondeada y directa/puntiaguda) lo cual permitiría distinguir un número mínimo de tres vasijas abandonadas en CL1.
ANÁLISIS FAUNÍSTICO
Para el estudio de los restos faunísticos se siguieron las propuestas generales de diversos autores (Fisher 1995; Gutiérrez 2004; Lyman 1994; Mengoni Goñalons 1999). Además, se utilizaron diversos índices y herramientas analíticas particulares (Andrews 1990; Borrero 1990; De Nigris 2004; De Nigris y Mengoni Goñalons 2005; Elkin 1995; Frontini y Deschamps 2007; Giardina 2007; Gutiérrez y Kaufmann 2007; López Mazz y Estévez Escalera 2006; Mello Araujo y Marcelino 2003; Pardiñas 1999).
Aspectos taxonómicos, anatómicos y cuantitativos
Del total de materiales faunísticos de CL1 (n= 4494), se efectuó el análisis taxonómico y anatómico
de una muestra (n= 1157). Esta muestra está integrada por los restos óseos y malacológicos registrados en planta en todas las cuadrículas y aquellos recuperados por medio del cribado de los sedimentos en ocho cuadrículas (6, 7, 12, 13, 14, 15, 20 y 21). De los fragmentos óseos indeterminados (n= 735; 63,5%),
sólo el 26,8% (n= 197) son mayores a los 2 cm. Los valores de NISP, NME y NMI calculados para los restos identificados a algún nivel taxonómico (n= 422; 36,5%) se presentan en la Tabla 2.
Tabla 2. Taxa representados en el paleosuelo de CL1.

Si bien esta sección se centra en los materiales del paleosuelo, es necesario mencionar algunas características relevantes del contexto faunístico del depósito eólico. En este último predomina la fauna introducida sobre la autóctona. Así, una diferencia con el paleosuelo es la ausencia de moluscos marinos, tucu-tuco (Ctenomys sp.) y venado de las pampas (Ozotoceros bezoarticus) y la menor frecuencia de múridos (Muridae), armadillos (Dasipodidae), vizcacha (Lagostomus sp.) y guanaco. Por último, cabe destacarse el hallazgo en la base del depósito eólico de un fémur proximal izquierdo de una gallinácea (Cracidae), familia de aves que no se encuentra en la actualidad en el área (Narosky e Yzurieta 1987).
Restos de cáscaras de huevos de reidos
Los 751 fragmentos de cáscaras de huevos de reidos recuperados en el paleosuelo fueron analizados con el fin de estimar el número mínimo de huevos (NMH) ingresados al sitio1. Para ello se estudiaron colecciones de huevos actuales de Pterocnemia pennata (ñandú petiso) y Rhea americana (ñandú común). Estas colecciones fueron seleccionadas procurando que su procedencia fuese de diferentes regiones del país2 y privilegiando aquellas recolectadas en condiciones silvestres. En estas muestras se promedió el peso para ambas especies, luego, se calculó el peso de los fragmentos arqueológicos y a partir de estos datos se estimó el NMH. Es necesario especificar que parte de las cáscaras de huevo de CL1están termoalteradas, factor cuya incidencia en el conjunto se tuvo en cuenta, ya que provoca la pérdida de masa.
Con la finalidad de analizar las diferencias de peso, se compararon huevos enteros que estuvieran vacíos de P. pennata y R. americana. Para esto se utilizaron colecciones del Museo de La Plata (MLP; n= 4), del Museo de Historia Natural de San Rafael (MSR; n= 5), del Museo Argentino de Ciencias Naturales "Bernardino Rivadavia" (MBR; n= 13) y del Zoológico de La Plata (ZLP; n= 9). Sobre la base de esta muestra se calculó el peso medio de los huevos: 67,12 g para P. pennata y 75,62 g para R. americana (Tabla 3).
Tabla 3. Peso, diámetro y altura de cada huevo de Pteronemia pennata y Rhea americana.
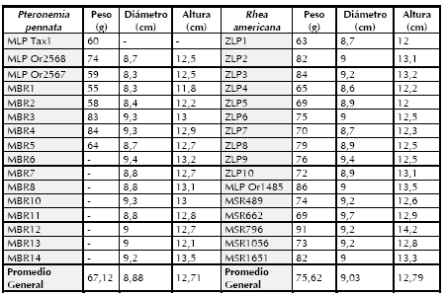
Con el objeto de evaluar si existían diferencias en los pesos de cada especie se aplicó el test de Student, obteniendo como resultado una diferencia no significativa (t= 0,53; p > 0,05). Esto concuerda con la ausencia de variaciones morfológicas significativas entre el diámetro y la altura para ambos taxa (Tabla 3). Al no existir diferencias entre los pesos de estas especies se decidió utilizar un peso promedio general de reidos de 71,37 g/huevo para comparar con la muestra arqueológica. Además, este peso promedio coincide con el obtenido para P. pennata por Fernández (1988- 1990).
Además del cálculo del peso, a partir del método de cuantificación de la densidad de poros de los huevos de reidos (Apolinaire y Turnes 2007), se pudo determinar que las cáscaras de huevo recuperadas en CL1 corresponden a R. americana. La totalidad de los fragmentos fueron pesados en una balanza digital Libra mod. 850, arrojando un valor de 137,94 g. Debido a que parte de la muestra (47,8%) presenta alteración térmica, se clasificaron los fragmentos dentro de las categorías señaladas por López Mazz y Estévez Escalera (2006). Estos autores realizaron un experimento en el que establecieron la relación entre temperatura, pérdida de masa y modificaciones en la superficie del huevo. Sobre la base de estos resultados se calculó la pérdida de masa por termoalteración en la muestra arqueológica, obteniendo un peso total corregido de 142,41 g (Tabla 4). Con este valor corregido y el peso obtenido para huevos de reidos (71,37 g) se pudo establecer que el NMH recuperados en el paleosuelo corresponde a dos huevos3. Sin embargo, debido a que en la unidad eólica también se registraron cáscaras de huevo sin termoalteración y al desplazamiento vertical que puede haber ocurrido con estos materiales (véase más adelante), se recalculó el NMH de los fragmentos termoalterados. Este valor de NMH recalculado es de un huevo.
Tabla 4. Categorías de termoalteración y peso de los fragmentos de cáscara de huevo de CL1

.Restos óseos
La especie más representada en el paleosuelo, tanto en cantidad de especímenes como de individuos, es guanaco (NISP= 72; NMI= 5). A partir de la secuencia de fusión ósea (Kent 1982) y de la comparación con un esqueleto de neonato (C. Kaufmann, comunicación personal 2007), se pudo establecer la presencia de un guanaco neonato, dos juveniles y otros dos probablemente adultos.
Para el estudio de los restos de esta especie, se aplicaron diferentes medidas de abundancia anatómica (NME, MAU y MAU%) para luego evaluar su representación esqueletaria por medio de distintos marcos de referencias (Tablas 5 y 6). El esqueleto axial se encuentra representado de manera dispar por el cráneo, mandíbula, vértebras cervicales y caudales, costillas y pelvis. En la Tabla 5 se observa que los MAU% más elevados corresponden a la mandíbula y la pelvis, mientras que el cráneo posee valores medios. Las costillas y vértebras cervicales son escasas y las vértebras toráxicas y lumbares están ausentes. Esta falta de elementos del esqueleto axial se puede deber a la pérdida de partes diagnósticas para su identificación. En este sentido, dentro de la categoría de mamíferos medianos (que podrían corresponder a guanaco), los especímenes más representados son las costillas (NISP=7). De todas maneras, si se compara el número de costillas registradas entre los mamíferos medianos con la frecuencia esperada de costillas de acuerdo al NMI de guanaco (NMI= 5) la frecuencia de este elemento sigue siendo baja (NMEe= 120)4.
Tabla 5. Abundancia de partes esqueletarias de guanaco.
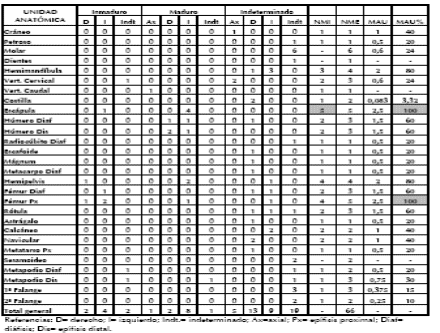
Tabla 6. Correlaciones establecidas para el MAU% de guanaco.

En relación con el esqueleto apendicular los valores más elevados se encuentran en las extremidades superiores, tanto delanteras como traseras. La escápula muestra el MAU% más alto y la diáfisis del húmero un valor medio. El cuarto trasero posee valores máximos de la porción proximal del fémur y medios de la diáfisis del mismo elemento. Los elementos de las extremidades medias, inferiores y del pie están subrepresentados o ausentes como es el caso de la tibia.
Con el fin de evaluar si el perfil esqueletario de guanaco en CL1 se debe a factores atricionales y/o antrópicos fue correlacionado, mediante el test rho de Spearman el MAU% con distintos índices: DMO (densidad mineral ósea), IC% (índice de carne) y DUI% (índice de secado) obtenidos para guanaco y Lama glama (Borrero 1990; De Nigris y Mengoni Goñalons 2005; Elkin 1995; Lyman 1994). Estas correlaciones se realizaron para el esqueleto completo, resultando todas no significativas (Tabla 6).
La ausencia de correlaciones de las unidades anatómicas de guanaco con los diferentes marcos de referencia sugiere, por un lado, que la supervivencia diferencial de los elementos óseos, mediados por la densidad, no sería el factor principal que determinaría el perfil esqueletario. Por otro lado, este perfil tampoco puede ser atribuido a decisiones de transporte diferencial de las distintas unidades anatómicas basadas exclusivamente en el rinde cárnico o en su potencial de secado. En el sitio se hallaron elementos con altos y bajos valores nutricionales, con una tendencia hacia los de mayores rindes. En este sentido, las regiones anatómicas estandarizadas (sensu De Nigris 2004) con mayores frecuencias en el sitio son aquellas ricas en grasa (extremidades superiores y cráneo) y carne (cinturas) susceptibles de ser c o n s u m i d a s inmediatamente (Figura 6).

Figura 6. MAU% y regiones anatómicas estandarizadas para guanaco.
Además de guanaco, en CL1 se recuperaron restos de moluscos, aves y diversos taxa de mamíferos (Tabla 2). En los párrafos siguientes es discutida la representación de partes esqueletarias de aquellos taxa considerados de importancia en la subsistencia de las sociedades prehispánicas o informativos sobre los procesos de formación de sitio.
Entre las aves, se identificó un fragmento de acetábulo de pelvis de ñandú, que sería la unidad anatómica con mayor cantidad de grasa y carne según los valores obtenidos para P. pennata (Giardina 2007). Respecto a los mamíferos, los escasos restos de múridos corresponden al esqueleto apendicular (NME= 3), mientras que de Ctenomys sp. sólo se registran elementos del cráneo (NME= 12). La representación esqueletaria general de estos micromamíferos (múridos y Ctenomys sp.) incluye elementos del esqueleto axial craneal así como húmero y tibia del esqueleto apendicular. Debido a los escasos elementos recuperados de estos taxa, sus perfiles anatómicos no marcan tendencias claras sobre el agente acumulador.
Las partes esqueletarias determinadas para los dasipódidos (Dasipodidae, Chaetophractus sp. y Chaetophractus villosus) corresponden al esqueleto postcraneal, tanto axial (NME= 3) como apendicular (NME= 4), así como a placas fijas y móviles del caparazón (NME= 64). En el caso de Lagostomus sp., se identificaron elementos craneales (NME= 6), del esqueleto apendicular posterior (NME= 9) y de la pelvis (NME= 2). Existen diferencias en la representación anatómica de estos dos grupos de mesomamíferos. En los dasipódidos están ausentes el cráneo y las cinturas y en las vizcachas faltan elementos del esqueleto axial postcraneal (con excepción de la pelvis) y del apendicular anterior. La inexistencia de marcos de referencia para dasipódidos y Lagostomus sp. no permite avanzar demasiado en la discusión del significado de sus perfiles esqueletarios. Aún así, la comparación realizada entre el MAU% de Lagostomus sp. y la densidad ósea promedio para lepóridos (Pavao y Stahl 1999) no mostraría que la frecuencia de partes esté asociada con este atributo (para un caso de aplicación similar véase Labarca 2005).
En el caso del venado de las pampas, está presente la pelvis (NME= 1) y algunos elementos del esqueleto apendicular posterior (NME= 5; tarsianos y metatarsianos). Las partes registradas en CL1 muestran valores densitométricos bajos a medios, si se los compara con los valores relativos calculados para Ovis aries (Symmons 2005). Los elementos presentes por tanto no serían producto de la preservación diferencial.
El hecho de que el perfil esqueletario de guanaco y, probablemente, los correspondientes a venado y vizcacha no estén mediados por la densidad mineral ósea, no descarta que algunos restos óseos estén subrepresentados debido a otros factores. Esta posibilidad no debe ser desestimada, sobre todo si se tiene en cuenta que el ambiente sedimentario corresponde a una planicie de inundación y que el agua puede haber transportado diferencialmente los huesos. Además, es necesario evaluar la influencia de otros agentes y procesos en la conformación del conjunto (i.e., erosión del paleosuelo y la consecuente destrucción de elementos, el accionar de raíces y animales fosoriales, etc.).
Efectos tafonómicos sobre los restos óseos
Para el estudio tafonómico de los materiales óseos la muestra ya mencionada fue dividida en dosconjuntos. En primer lugar, se analiza el conjunto compuesto por los especímenes óseos identificados a algún nivel taxonómico (n=149) y los restos óseos indeterminados mayores a los 2 cm (n= 197). En segundo lugar, se examina el conjunto constituido por los especímenes de edentados y roedores (n= 106). Las variables tafonómicas de estos mamíferos menores se presentan en una sección aparte, debido a que se quiere evaluar si los mismos se depositaron sincrónica o diacrónicamente a la ocupación prehispánica y si su ingreso al sitio fue natural o antrópico.
Evaluación de las variables tafonómicas
A continuación se presenta el análisis de las variables tafonómicas relevadas en los 346 especímenes óseos. La meteorización fue registrada en un 11% y se reparte en los siguientes estadios: 0 (n=4), 1 (n= 9), 2 (n= 8) y 3 (n= 17). El alto porcentaje (89%) de especímenes con estadios indeterminados se debe a sus reducidos tamaños o a que sus superficies estaban intensamente grabadas por raíces. La presencia del estadio de meteorización 3 está indicando que parte de los restos estuvieron expuestos por un tiempo prolongado antes de su sepultamiento.
Se registraron especímenes (4,8%; n= 17) con abrasión geológica, esto incluye pátinas brillosas sobre la superficie cortical (n= 8) y bordes redondeados y pulidos (n= 9). De estos especímenes, solo pudieron ser determinados dos de guanaco. Cabe aclarar que nueve de ellos están termoalterados y que el desarrollo de superficies brillosas o vítreas puede ser producto de la acción del fuego (Nicholson 1993). También se observó la depositación de carbonato de calcio en algunos ejemplares (4,1%; n= 14). Por su parte, las trazas generadas por carnívoros y roedores son muy escasas. Sólo se registraron dos especímenes de guanaco (0,6%) con marcas de carnívoros y uno indeterminado (0,3%) con marcas de roedores.
Las marcas de raíces fueron registradas en un 45,4% (n= 157) y los hoyos de disolución química en un 26,9% (n= 93). Cabe aclarar que cinco especímenes con hoyos de disolución se encuentran en un estadio 4 (5,4%) y los demás muestran estadios menores (sensu Gutiérrez 2004). De acuerdo a Montalvo (2002), la coloración más clara de las marcas de raíces indica que se formaron cuando el hueso estaba seco y bastante tiempo después de su entierro. En CL1 se observaron numerosas marcas de raíces con una coloración interna clara (amarillo pálido 2,5Y 8/3) y, en menor proporción, con una tonalidad similar al resto del espécimen (marrón oliva claro 2,5Y 5/4). Estos dos tipos de coloración interna concuerdan con lo esperable para restos sometidos a la acción de la vegetación del paleosuelo y, luego, de las plantas del sedimento eólico edafizado y el suelo actual.
La mayoría de los huesos se encontraba en estado seco al momento de ser fragmentados (Tabla 7). Sólo seis especímenes fueron fracturados en estado fresco (cuatro con marcas de impacto), de los cuales dos húmeros y un metapodio corresponden a guanaco y los otros tres a huesos largos de mamíferos. Las fracturas indeterminadas son aquellas en las que no fue posible observar atributos diagnósticos debido al intenso grabado por raíces de los especímenes (el 55,3% de los restos incluidos en esta categoría tienen más del 90% de su superficie grabada).
Tabla 7. Estado del hueso al momento de la fractura.

En el 16,8% de los especímenes (n= 58) se registraron alteraciones térmicas con diferentes grados: quemados (8,5%), carbonizados (67,1%) y calcinados (24,4%). Tres corresponden a elementos de guanaco (una vértebra cervical, una diáfisis de radio-cúbito y un sesamoideo). Además, se observaron cuatro especímenes que poseen una coloración negra en su interior y mantienen un tono marrón en su parte externa. Según Buikstra y Swegle (1989), los huesos en estado seco que son quemados suelen adquirir esta coloración. Sin embargo, procesos diagenéticos también tiñen los restos óseos con una tonalidad oscura (Marshall 1989; Shahack-Gross et al.1997).
Armadillos y roedores
Las variables relevadas sobre los 106 especímenes de micro (i.e., tuco-tuco) y mesomamífero (i.e., vizcacha y armadillos) son las mismas que para el primer conjunto. Los resultados se resumen en la Tabla 8, aunque es necesario efectuar una serie de especificaciones. En lo referido a la meteorización, los estadios (sensu Andrews 1990) registrados en micro y mesomamíferos fueron el 0 (n= 4), 1 (n= 1) y 3 (n= 1). La abrasión corresponde sólo a especímenes pulidos de vizcacha y peludo. El carbonato de calcio se depositó sobre los restos óseos pertenecientes a Ctenomys sp. Las marcas de raíces así como los hoyos de disolución química se identificaron tanto en micro como en mesomamíferos. Esta segunda variable presenta estadios semejantes al conjunto considerado en la sección previa.
Tabla 8. Representación de las variables tafonómicas.
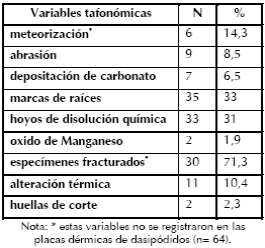
La proporción de especímenes fracturados es menor que en el conjunto anterior. Con respecto a la alteración térmica, se observaron placas dérmicas, huesos largos y del esqueleto axial de mesomamíferos (dasipódidos y vizcacha) con claras evidencias (calcinados o con una gradación de quemados a carbonizados). A esto se le agrega el registro de huellas de corte sobre una tibia y un cúbito de peludo que podrían ser el producto de la desarticulación del esqueleto (Figura 7).

Figura 7. Termoalteraciones y huellas de corte en especímenes de mesomamíferos (escala 2 cm).
De acuerdo al escaso número de especímenes de micromamíferos y a la ausencia de indicadores claros de explotación humana (huellas de corte, termoalteraciones, sobre representación de una clase etaria) se considera que el ingreso al sitio de la mayoría de estos roedores se debió a causas naturales etoecológicas o bien a predadores (Pardiñas 1999). La muerte natural de Ctenomys sp. (taxón más abundante en CL1) en el paleosuelo no parece posible debido a sus requerimientos ecológicos (Contreras 1984a). La ausencia de meteorización y la fragmentación de los elementos de los micromamíferos podrían estar indicando que los agentes responsables de su acumulación fueron predadores no humanos (Pardiñas 1999), aunque no se registraron egagrópilas o heces ni alteraciones por digestión.
Las trazas relevadas para los especímenes de peludo y vizcacha indican su explotación humana. Por su parte, la existencia de especímenes abradidos y con óxido de Manganeso se asocia con entornos saturados en agua (Dorn y Oberlander 1981). También, algunos individuos podrían haber muerto naturalmente en esta unidad. Sin embargo, los requerimientos ambientales de las vizcachas no concuerdan con un medio fluvial ni con este tipo de sedimento (Contreras 1984b) y trabajos actualísticos que analizaron los contenidos de cuevas de peludos no registraron casos de individuos muertos de esta especie (Abba et al. 2005; Frontini y Deschamps 2007).
Para finalizar se quiere mencionar la influencia que pudieron tener los animales de conductas fosoriales sobre el sitio. Durante la excavación se detectaron estructuras de cuevas, probablemente de dasipódidos, que alcanzaban el paleosuelo. No obstante, estos taxa que generan desplazamientos y mezclan materiales de distintas unidades, a partir de la distribución vertical de los materiales y de sus mayores frecuencias se pueden diferenciar las unidades originarias de depositación (Mello Araujo y Marcelino 2003).
DISCUSIÓN
Las trazas relevadas en los materiales arqueológicos, su distribución vertical y el contexto estratigráfico de CL1 permiten abordar la historia tafonómica y definir la existencia de dos componentes en el sitio. Los sedimentos y los gasterópodos del paleosuelo indican que la depositación de los materiales prehispánicos que caracterizan al componente inferior ocurrió en un sector distal de una planicie de inundación. A partir de los estadios de meteorización de los restos óseos se infiere que los materiales generados por la ocupación humana estuvieron expuestos en la superficie del terreno por cierto tiempo, durante el cual fue acumulada la mayoría de los micromamíferos por agentes naturales no claramente determinados. La presencia de tiestos cerámicos suavemente erosionados así como de escasos especímenes óseos levemente abradidos o con óxido de Manganeso señala que el agua participó en la formación del conjunto, aunque de manera limitada. En este sentido, la representación de partes esqueletarias de guanaco no se asocia de manera clara con un grupo de dispersión hídrica si se consideran las diferentes medidas propuestas por Gutiérrez y Kaufmann (2007).
Luego, se produjo una reducción del cuerpo de agua, evidenciada por la depositación de carbonato de calcio sobre algunos especímenes óseos, y el conjunto fue sepultado. Una vez enterrados los restos del componente inferior fueron alterados por la acción química y mecánica de las raíces. Con posterioridad, el techo del paleosuelo que contenía materiales prehispánicos fue erosionado (Favier Dubois 2007), reexponiendo parte del conjunto en la superficie. Este proceso erosivo perturbó solo parcialmente al componente prehispánico del paleosuelo, ya que las mayores frecuencias de hallazgos no se detectaron sobre la superficie erosiva (véase Mayer 2002).
Sobre esta superficie discordante se acumuló el sedimento eólico, edafizado en forma homogénea.5 En este momento se depositan los objetos posthispánicos tardíos (siglos XIX a XX) del componente superior. La vegetación asociada al desarrollo del suelo sobre el sedimento eólico afectó intensamente a los materiales del componente inferior del paleosuelo, fracturando a los elementos óseos y provocando probablemente la migración vertical de los ítems más pequeños. Como ya se mencionó, los armadillos también jugaron un rol importante en la conformación final del conjunto. Según Mello Araujo y Marcelino (2003) el desplazamiento de materiales por los armadillos no es selectivo por el tamaño y ocurre tanto hacia arriba cuando cavan sus cuevas como hacia abajo por gravedad o el colapso de las mismas. Esto, junto con la acción de las raíces, podría explicar el registro de fauna introducida en el paleosuelo y la asociación de materiales de factura indígena junto a objetos posthispánicos tardíos en la base de la unidad eólica.
Por tanto, la meteorización, las raíces y el accionar de los armadillos fracturaron el material prehispánico del componente inferior. En tanto que la erosión del paleosuelo, las raíces y los armadillos generaron la migración vertical y mezcla de los materiales de ambos componentes arqueológicos. Para finalizar, si bien se sucedieron estos marcados procesos postdepositacionales, el análisis realizado permite discernir los materiales derivados de la ocupación prehispánica y profundizar el estudio de las actividades llevadas a cabo en el sitio.
La fauna explotada por los ocupantes prehispánicos está constituida por animales de origen continental. El único taxa marino está representado por dos individuos de Zidona sp. (voluta), probablemente asociados a la recolección de exoesqueletos vacíos en el sector de playa (Bonomo 2007). El guanaco fue el recurso principal, aunque también se aprovecharon venado de las pampas, armadillos, vizcacha y ñandú. Las unidades seleccionadas del guanaco son aquellas ricas en carne, médula ósea y grasa (esqueleto apendicular superior, cinturas y cabeza). El análisis cuantitativo de los restos óseos apuntaría al consumo inmediato de estas unidades que son menos apropiadas para ser conservadas, mientras que los elementos con bajo valor nutricional (i.e., metapodios) o con alto rinde en carne (costillar y columna vertebral) se abandonaron, quizás luego de su consumo, en otro lugar.
El aprovechamiento inmediato de partes no susceptibles de ser conservadas también se condice con el hallazgo de una pelvis de ñandú. Su presencia en CL1 indicaría el transporte diferencial y consumo local de aquellas unidades anatómicas con elevado contenido de carne y grasa (Giardina 2007). Los animales de menor porte que fueron explotados (armadillos y vizcacha) pueden haber sido acarreados enteros al sitio, pero su perfil esqueletario no es del todo claro en este sentido. En el caso de los armadillos, las placas dérmicas termoalteradas en una sola de sus caras posiblemente estén mostrando que para su cocción se haya colocado al animal entero con el caparazón sobre el fuego, como fuera referido para distintos grupos etnográficos de Argentina (Arenas 2003: 426; Cox 2005: 259 [1863]; entre otros).
Teniendo en cuenta la estacionalidad de los recursos utilizados en CL1 se puede estimar el momento en que fueron abandonados los materiales del componente inferior. La puesta de huevos del ñandú comienza en primavera y se prolonga hasta enero (Fernández y Reboreda 1998). La parición de los guanacos ocurre desde diciembre hasta febrero (Larrieu et al. 1979; Raedeke 1978). El hallazgo de cáscaras de huevo de ñandú y de un neonato de guanaco señalaría que el sitio fue ocupado hacia finales de la primavera y comienzos del verano. El registro de Lama guanicoe y Lestodelphys sp. sugiere que las condiciones climáticas durante esta ocupación fueron áridas y más frías que en la actualidad (Tonni et al. 1999). Es llamativa la presencia de Cracidae, ya que la distribución actual de estas aves subtropicales llega al sur hasta la Depresión del Salado (Narosky e Yzurieta 1987). Si bien esto registra a estas gallináceas en un sector austral fuera de su área de distribución, la asociación con el componente prehispánico no es clara debido a que se recuperó en el depósito eólico.
Claromecó 1 tiene interesantes implicancias en relación a los contextos generados durante las ocupaciones humanas de la costa y del interior pampeano. La fauna de CL1 se asemeja a la de los sitios del interior del área Interserrana en donde se explotaron básicamente mamíferos terrestres (sobre todo guanaco). Esto también ocurre en sitios de las llanuras muy próximas a la costa, como Nutria Mansa 1 (a 3,5 km del litoral) y Quequén Salado 1 (a 11 km), donde los especímenes de mamíferos terrestres representan el 99,97% y 100%, respectivamente (Bonomo 2004, 2005; March et al. 2006).
La ubicación geográfica y la tecnología lítica utilizada hacen que CL1 posea características similares a los talleres de la faja de médanos del litoral marítimo. En los talleres costeros y en CL1 existe un predominio en la explotación de rodados respecto a las rocas serranas del interior. Al igual que en los talleres litorales (Bonomo 2005), la materia prima costera más abundante es el basalto, se registran cantidades sustanciales de piezas sobre rodados con corteza y los instrumentos son escasos. La talla bipolar también está presente, evidenciada por el hallazgo de abundantes núcleos y lascas con claros atributos asociados a esta técnica.
Una distinción con los sitios de la faja de médanos, donde gran parte de las rocas costeras poseen granos gruesos con calidades para la talla regulares o malas (32%) y bajas frecuencias relativas de calidades muy buenas (6%) (Bonomo 2004), es que los rodados reducidos en CL1 son de grano fino y muestran menores proporciones de calidades regulares o malas (6%) y mayores de muy buenas (22%). Estas particularidades implican que los nódulos costeros transportados sin previa modificación a CL1 han sido anteriormente seleccionados de acuerdo a sus aptitudes para la talla, ya que no pueden ser reemplazadas fácilmente como ocurre con los talleres donde los rodados están disponibles en sus inmediaciones.
Una selección similar se observó en Quequén Salado 1, sitio cercano y con una cronología de fines del Holoceno tardío (950-360 años AP) (Bonomo 2004; Madrid et al. 2002). En QS1 los rodados de basalto y sílice de buenas y muy buenas calidades para la talla también presentan porcentajes significativos (36%) y muestran un alto grado de aprovechamiento mediante la técnica de talla bipolar. Además en ambos sitios se recuperaron preformas y puntas de proyectil triangulares pequeñas de base recta o cóncava sobre materias primas costeras. A partir de la presencia de estas puntas pequeñas se infiere que ambos asentamientos habrían estado vinculados en parte con actividades de caza con arco y flecha.
En suma, los resultados radiocarbónicos obtenidos indican que la ocupación humana del sitio CL1 ocurrió a fines del Holoceno tardío, lo cual concuerda con el registro de puntas triangulares pequeñas y de alfarería en este contexto. Allí, se desarrollaron diversas actividades como la reducción de rodados, la manufactura de instrumentos líticos, el uso de pigmentos minerales, la utilización de cerámica con características tecnológicas homogéneas, el consumo de huevos de ñandú y de grandes y pequeños vertebrados. A esto se le agregan los siguientes indicadores propuestos por Sivertsen (1980) observados en CL1: 1) restos óseos dispersos, todos desarticulados y altamente fragmentados; 2) varios individuos de una especie representados por pocos elementos (i.e., 66 elementos que corresponden a cinco guanacos); 3) huesos con evidencias de procesamiento y consumo y 4) alta proporción de material lítico. Esta información permite proponer que CL1 fue un campamento que por la baja densidad de materiales recuperados sería transitorio, representando un solo evento de ocupación.
CONCLUSIONES
Claromecó 1 es un sitio cuyos materiales líticos proceden del litoral y las sierras de Tandilia. Un aspecto destacable en relación a la tecnología lítica es que se tallaron por medio de la técnica bipolar elevadas proporciones de rodados recolectados en el litoral atlántico. En cambio, las rocas del interior fueron menos utilizadas, lo que implicaría viajes menos frecuentes a las canteras serranas. Estas características en los recursos líticos explotados diferencian a este conjunto de la gran mayoría de los sitios de las llanuras y las sierras, donde los rodados costeros tuvieron un rol subsidiario, incluso en las cercanías de la costa. Aun así, al igual que en estos sitios del interior, los análisis faunísticos señalan que en CL1 se aprovecharon principalmente animales de origen continental. Su cronología fue establecida en 800 años AP, lo cual cobra relevancia debido a la escasez de dataciones para la costa pampeana. Por último, la información obtenida muestra que CL1 fue un campamento ocupado por cazadores-recolectores prehispánicos durante el Holoceno tardío.
Para finalizar, es interesante destacar la explotación de animales de porte medio en CL1. Esta mayor participación en la subsistencia de los taxa de menor tamaño fue identificada en diversos sitios tardíos del área Interserrana -Laguna del Trompa, La Toma-, de Tandilia -Cueva Tixi, Cueva El Abra-, de la Depresión del Salado -Localidad La Guillerma-, entre otros contextos (González 2005; Mazzanti y Quintana 2001; Quintana et al. 2003; Salemme 1987; Silveira 1991). Esto concuerda con el proceso de diversificación e intensificación en la explotación de la fauna planteado para las poblaciones humanas que ocuparon la región durante el Holoceno tardío (Martínez y Gutiérrez 2004). Esta transformación fue acompañada por un aumento demográfico, menor movilidad residencial, innovaciones tecnológicas, amplias redes de interacción social e intercambio entre grupos humanos de distintas zonas geográficas (Berón 2007; Bonomo 2005; González 2005; Mazzanti 2006; Politis et al. 2001). En este escenario, la explotación más intensa de recursos líticos de segunda categoría, como lo fueron los rodados a lo largo del desarrollo cultural pampeano, pueden representar un correlato tecnológico de este proceso de diversificación e intensificación.
Agradecimientos
Los trabajos de campo y laboratorio se financiaron con fondos de dos proyectos denominados "Una perspectiva suprarregional de la arqueología del sudeste de la región pampeana" (UNLP 11/N503) y "Arqueología de las poblaciones indígenas del sudeste de la región pampeana desde un abordaje suprarregional" (PIPCONICET 5424), dirigidos por Gustavo Politis. Además, se desea agradecer a María Gutiérrez y a los evaluadores del artículo (Mónica Berón y un revisor anónimo) por las sugerencias realizadas, a Pamela Steffan por el análisis de gasterópodos, a Cristian Favier Dubois por el estudio geoarqueológico, a Cecilia Deschamps, Germán Moreira y Estela Martín por las determinaciones de meso y micromamíferos y gasterópodos marinos. A Alejandra Matarrese por el análisis del instrumento lítico no tallado y a Irina Capdepont por la ayuda con la cerámica. A Miguel Giardina (MSR), Pablo Tubaro (MBR) y a la División de Ornitología (MLP) por su colaboración en el estudio de los huevos de reidos. Agradecemos especialmente a Mónica Berón, Luciano Prates y Agustina Massigoge por la lectura crítica del manuscrito e importantes comentarios. Los únicos responsables de los contenidos de este artículo son los autores.
1 Paralelamente a nuestros análisis, C. Quintana (comunicación personal 2007) ha desarrollado estudios similares para determinar el número mínimo de huevos de reidos (véase Quintana 2008 en este volumen de la revista).
2 Las muestras de huevos de Pterocnemia pennata provienen de la región patagónica (provincias de Chubut y Santa Cruz). En el caso de Rhea americana proceden de diferentes zonas del país (Chaco, Buenos Aires, La Pampa y Mendoza); solo en la colección del Zoológico de La Plata la puesta de los huevos aconteció en condiciones de cautiverio.
3 Es necesario aclarar que también se recuperó en el depósito eólico edafizado, a una profundidad de 60 cm y en una superficie de 1 m2, una concentración de fragmentos de huevo de Rhea americana sin evidencias de termoalteración. El peso de estos fragmentos (1118 g) indica como mínimo la presencia de 16 huevos. Este número concuerda con lo esperable para un nido que puede contener entre 8 y 56 huevos, aunque lo más común es que se acerque a los 20 o 30 huevos (Fernández y Reboreda 1998). Además, el depósito donde se recuperó esta concentración es similar al ambiente de médanos fijos donde actualmente nidifica esta especie (Bó y Darrieu 1991).
4 MNEe= número mínimo de elementos esperados.
5 La homogeneidad muestra que el sedimento eólico edafizado "podría corresponderse con el desarrollo de un espeso horizonte cumúlico, lo que implicaría un lento y continuado aporte de sedimentos eólicos sobre una superficie vegetada. Aunque quizás esta unidad superior se haya originado por el transporte eólico de sedimentos ya edafizados hacia el valle, derivados de la erosión de suelos arenosos en la localidad por el impacto de las actividades agropecuarias" (Favier Dubois 2007: 2).
REFERENCIAS CITADAS
Abba, A., D. Urdizar Sauthier y S. Vizcaino 2005 Distribution and use of burrows and tunnels of Chaetophractus villosus (Mammalia Xenarthra) in the eastern Argentinian pampas. Acta Theriologica 50 (1): 115-124. [ Links ]
Andrews, P. 1990 Owls, Caves and Fossils. Natural History Museum Publications, Londres. [ Links ]
Apolinaire, E. y L. Turnes 2007 Todos los huevos en la misma canasta?: Un método para la diferenciación interespecifica en Rehidos. Actas del XVI Congreso de Arqueología Argentina 2: 615-618. San Salvador de Jujuy. [ Links ]
Arenas, P. 2003 Etnografía y Alimentación entre los TobaÑachilamole#ek y Wichí-Lhuku´tas del Chaco Central (Argentina). Pastor Arenas, Buenos Aires. [ Links ]
Aschero, C. 1975 Ensayo para una clasificación morfológica de artefactos líticos. Informe presentado al CONICET. Copia disponible en el Laboratorio Nº 3, División Científica de Arqueología, Museo de La Plata (UNLP), La Plata. MS. [ Links ]
Aschero, C. 1983 Registro de códigos para atributos descriptivos aplicados a artefactos líticos. Informe presentado al CONICET. Copia disponible en el Laboratorio Nº 3, División Científica de Arqueología, Museo de La Plata (UNLP), La Plata. MS. [ Links ]
Berón, M. 2007 Circulación de bienes como indicador de interacción entre las poblaciones de la pampa occidental y sus vecinos. En Arqueología en las pampas, editado por C. Bayón, A. Pupio, M. I. González, N. Flegenheimer y M. Frére, vol. 1, pp. 345-364. Sociedad Argentina de Antropología, Buenos Aires. [ Links ]
Bó, N. y C. Darrieu 1991 Ornitosinosis de un sector de la zona de dunas costeras de la Provincia de Buenos Aires. En Situación ambiental de la Provincia de Buenos Aires. Recursos y rasgos naturales en la evaluación ambiental, vol. 1 (2), pp. 6-30, CIC, La Plata. [ Links ]
Bonomo, M. 2004 Ocupaciones humanas en el litoral marítimo pampeano. Un enfoque arqueológico. Tesis Doctoral inédita. Facultad de Ciencias Naturales y Museo (UNLP), La Plata. [ Links ]
Bonomo, M. 2005 Costeando las llanuras. Arqueología del litoral marítimo pampeano. Sociedad Argentina de Antropología, Buenos Aires. [ Links ]
Bonomo, M. 2007 El uso de los moluscos marinos por los cazadores-recolectores pampeanos. Chungara 39 (1): 87-102. [ Links ]
Borrero, L. A. 1990 Fuego-Patagonia bone assemblage and the problem of communal guanaco hunting. En Hunters of the Recent Past, editado por L. B. Davis y B. O. K. Reeves, pp. 373-399. Unwin Hyman, Londres. [ Links ]
Buikstra, J. y M. Swegle 1989 Bone Modification Due to Burning: Experimental Evidence. En Bone Modification, editado por R. Bonnichsen y M. H. Sorg, pp. 247-257. Center of the Study of the First Americans, University of Maine, Orono. [ Links ]
Contreras, J. 1984 Los tuco-tucos. En Fauna Argentina, vol. 4, editado por M. A. Palermo, pp. 1-32. Centro Editor de América Latina, Buenos Aires. [ Links ]
Contreras, J.1984 La vizcacha. En Fauna Argentina, vol. 3, editado por M. A. Palermo, pp. 1-32. Centro Editor de América Latina, Buenos Aires. [ Links ]
Cox, G. E. 2005 [1863] Viaje en las rejiones septentrionales de la Patagonia (1862-1863). Elefante Blanco, Buenos Aires. [ Links ]
De Nigris, M. 2004 El consumo en grupos cazadores recolectores: Un ejemplo zooarqueológico de patagonia meridional. Sociedad Argentina de Antropología, Buenos Aires. [ Links ]
De Nigris, M. y L. Mengoni Goñalons 2005 The guanaco as a source of meat and fat in the Southern Andes. En The Zooarchaeology of Fats, Oils, Milk and Dairying, editado por J. Mulville y A. Outram, pp. 160-166. Oxbow Books, Durham. [ Links ]
Dorn, R. y T. Oberlander 1981 Microbial origin of desert varnish. Science 213: 1245-1247 [ Links ]
Elkin, D. 1995 Volume Density of South American Camelid Skeletal Parts. International Journal of Osteoarchaeology 5: 29-37. [ Links ]
Favier Dubois, C. 2007 Sitio Claromecó 1. Breve informe geoarqueológico. Laboratorio Nº 3, División Científica de Arqueología, Museo de La Plata (UNLP), La Plata. [ Links ]
Fernández, J. 1988-1990 La Cueva de Haichol. Arqueología de los pinares cordilleranos del Neuquén. Anales de Antropología y Etnología 43-45 (I-III): 1-740. [ Links ]
Fernández, G. J. y J. C. Reboreda 1998 Effects of Clutch Size and Timing of Breeding on Reproductive Success of Greater Rheas. The Auk 115 (2): 340-348. [ Links ]
Fisher, W. 1995 Bone Modifications in Zooarchaeology. Journal of Archeological Method and Theory 2 (1): 7-68. [ Links ]
Frontini, R. y C. Deschamps 2007 La actividad de Chaetophractus villosus en sitios arqueológicos. El Guanaco como caso de estudio. En Arqueología en las pampas, editado por C. Bayón, A. Pupio, M. I. Gonzalez, N. Flegenheimer y M. Frére, Tomo 1, pp. 439-454. Sociedad Argentina de Antropología, Buenos Aires. [ Links ]
Giardina, M. 2007 Anatomía económica de Rehidae. Intersecciones en Antropología 7: 263-276. [ Links ]
González, M. I. 2005 Arqueología de alfareros, cazadores y pescadores pampeanos. Sociedad Argentina de Antropología, Buenos Aires. [ Links ]
Gutiérrez, M. A. 2004 Análisis Tafonómicos en el Area Interserrana (Pcia. de Buenos Aires).Tesis Doctoral inédita. Facultad de Ciencias Naturales y Museo (UNLP), La Plata. [ Links ]
Gutiérrez, M. A. y C. A. Kaufmann 2007 Criteria for the Identification of Formation Processes in Guanaco (Lama guanicoe) Bone Assemblages in Fluvial-Lacustrine Environments. Journal of Taphonomy 5 (4): 151-176. [ Links ]
Isla, F. y M. Espinosa 1998 Modelo sedimentario de colmatación de pequeños estuarios dominados por limo, Provincia de Buenos Aires. Actas de la VII Reunión de Sedimentología: 24-36. Salta. [ Links ]
Kent, J. 1982 The Domestication and Exploitation of the South American Camelids: Methods of Analysis and the Application to Circum-lacustrine Archaeological Sites in Bolivia and Peru. Tesis Doctoral inédita. Washington University, Saint Louis. [ Links ]
Labarca, R. 2005 Las vizcachas de Piuquenes: evidencias de explotación de roedores durante la transición Pleistoceno-Holoceno en la cordillera andina de Chile central. Werken 6: 63-80. [ Links ]
Larrieu, E., N. Oporto y R. Bigatti 1979 Avances en estudios reproductivos en guanacos de Río Negro (Argentina). Revista Argentina de Producción Animal 3 (2): 134-149. [ Links ]
López Mazz, J. M. y J. Estévez Escalera 2006 Reconstructing the ñandú (Rhea americana) eggs consumption process in the Prehistory (ca. 3100 B.P.) on the Atlantic coast of southeastern South America. MS. en archivo, Laboratorio Nº 3, División Científica de Arqueología, Museo de La Plata (UNLP), La Plata. [ Links ]
Lyman, R. L. 1994 Vertebrate Taphonomy. Cambridge University Press, Cambridge. [ Links ]
Madrid, P., G. Politis, R. March y M. Bonomo 2002 Arqueología microrregional en el sudeste de la Región Pampeana Argentina: el curso del río Quequén Salado. Relaciones de la Sociedad Argentina de Antropología 27: 327-355. [ Links ]
March, R. J., C. Rodríguez Loredo, P. Madrid, G. Politis, M. Bonomo, D. Joly, L. Laporte, S. Hinguant, D. Marguerie y C. Karlin 2006 Évolution et adaptation humaine autour de la Région des plaines et lacs du fleuve Quequén Salado. Pcia. de Buenos Aires (Argentine): premiers pas d´une archeologie regionale programme: la prospection. En Proceedings of the XIVth UISPP Congress, Préhistoire de l'Amérique/American Prehistory, editado por H. D. Yacobaccio, D. E. Olivera y S. Purin, pp. 211-231. BAR International Series1524. Archaeopress, Oxford. [ Links ]
Marshall, F. 1989 Bone Modification and "The Laws of Burial". En Bone Modification, editado por R. Bonnichsen y M. Sorg, pp. 7-24. Center of the Study of the First Americans, University of Maine, Orono. [ Links ]
Martínez, G. y M. Gutiérrez 2004 Tendencias en la explotación humana de la fauna durante el Pleistoceno final y Holoceno en la Región Pampeana (Argentina). En Zooarchaeology of South America, editado por G. L. Mengoni Goñalons, pp. 81-98. BAR Internacional Series 1298, Archaeopress, Oxford. [ Links ]
Mayer, J. 2002 Evaluating Natural Site Formation Processes in Eolian Dune Sands: A Case Study from the Krmpotich Folson Site, Killpecker Dunes, Wyoming. Journal of Archaeological Science 29: 1199-1211. [ Links ]
Mazzanti, D. L. 2006 La constitución de territorios sociales durante el Holoceno tardío. El caso de las sierras de Tandilla. Relaciones de la Sociedad Argentina de Antropología 31: 277-300. [ Links ]
Mazzanti, D. L. y C. A. Quintana (editores) 2001 Cueva Tixi: cazadores y recolectores de las sierras de Tandil oriental. 1 Geología, Paleontología y Zooarqueología. Publicación Especial 1. Laboratorio de Arqueología (UNMdP), Mar del Plata. [ Links ]
Mello Araujo, A. y J. Marcelino 2003 The Role of Armadillos in the Movement of Archaeological Materials: An Experimental Approach. Geoarchaeology: An International Journal 18 (4): 433-460. [ Links ]
Mengoni Goñalons, G. L. 1999 Cazadores de guanacos de la estepa patagónica. Sociedad Argentina de Antropología, Buenos Aires. [ Links ]
Montalvo, C. 2002 Root traces in fossil bones from Huayquerian (Late Miocene) faunal assemblage of Telén, La Pampa, Argentina. Acta Geológica Hispánica 37 (1): 37-42. [ Links ]
Narosky, T. y D. Yzurieta 1987 Guía para la identificación de las aves de Argentina y Uruguay. Asociación Ornitológica del Plata, Buenos Aires. [ Links ]
Nicholson, R. 1993 Morphological investigation of burnt animal bone and an evaluation of its utility in archaeology. Journal of Archaeological Science 20: 411-28. [ Links ]
Orton, C., P. Tyers y A. Vince 1997 La cerámica en arqueología. Crítica, Barcelona. [ Links ]
Pardiñas, U. F .J. 1999 Tafonomía de microvertebrados en yacimientos arqueológicos de Patagonia (Argentina). Arqueología 9: 265-340. [ Links ]
Pavao, B. y P. Stahl 1999 Structural density assays of leporid skeletal elements with implications for taphoomic, actualistic, and archaeological research. Journal of Achaeological Science 26: 53-66. [ Links ]
Politis, G., G. Martínez y M. Bonomo 2001 Alfarería temprana en sitios de cazadores-recolectores de la Región Pampeana (Argentina). Latin American Antiquity 12 (2): 167-181. [ Links ]
Quintana, C. 2008 Cálculo del número mínimo de individuos de huevos de ñandú. Intersecciones en Antropología 9. En prensa. [ Links ]
Quintana, C., F. Valverde y A. Albino 2003 Registro de fauna del sitio Cueva El Abra, Tandilla oriental, Provincia de Buenos Aires. Actas del XIII Congreso Nacional de Arqueología Argentina 3: 317-324. Córdoba. [ Links ]
Rabassa, J. 1989 Geología de los depósitos del Pleistoceno Superior y Holoceno en las cabeceras del Rió Sauce Grande, Provincia de Buenos Aires. Actas de las 1ª Jornadas Bonaerenses Geológicas: 765-790. Bahía Blanca. [ Links ]
Raedeke, K. J. 1978 El guanaco de Magallanes, Chile. Su distribución y biología. Publicación Técnica 4, Ministerio de Agricultura, Santiago de Chile. [ Links ]
Rice, P. M. 1987 Pottery Analysis. A Sourcebook. The University of Chicago Press, Chicago. [ Links ]
Rye, O. 1981 Pottery technology: principles and reconstruction. Taraxacum, Washington, D.C. [ Links ]
Salemme, M. 1987 Paleoetnozoología del sector bonaerense de la Región Pampeana con especial atención a los mamíferos. Tesis Doctoral inédita. Facultad de Ciencias Naturales y Museo (UNLP), La Plata. [ Links ]
Shahack-Gross, R., O. Bar-Yosef y S. Weiner 1997 Black-Coloured Bones in Hayonim Cave, Israel: Differentiating between Burning and Oxide Staining. Journal of Archeological Science 24: 439-446. [ Links ]
Silveira, M. 1991 Análisis de los restos óseos de las excavaciones de los sectores "H" y "J" del sitio Laguna del Trompa, Estancia La Herminia, Laprida, Pcia. de Buenos Aires. Boletín del Centro 3: 105-115. [ Links ]
Simone, L. R. 2006 Land and freshwater molluscs of Brazil. EGB, Fapesp, São Paulo. [ Links ]
Sivertsen, B. 1980 A Site Activity Model for Kill and Butchering Activities at Hunter- Gatherer Sites. Journal of Field Archaeology 7: 423-441. [ Links ]
Symmons, R. 2005 New density data for unfused and fused sheep bones, and a preliminary discussion on the modelling of taphonomic bias in archaeofaunal age profiles. Journal of Archaeological Science 32: 1691-1698. [ Links ]

