










Servicios Personalizados
Revista

Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkIntersecciones en antropología
versión On-line ISSN 1850-373X
Intersecciones antropol. n.9 Olavarría ene./dic. 2008
Análisis tafonómico de micromamíferos y mesomamíferos del sitio Laguna La Barrancosa 1 (Partido de Benito Juárez, provincia de Buenos Aires)
Gustavo N. Gómez y Pablo G. Messineo
Gustavo N. Gómez. INCUAPA, Departamento de Arqueología, Facultad de Ciencias Sociales, UNCPBA, Avda. del Valle 5737 (B7400JWI) Olavarría, Buenos Aires, Argentina. E-mail: ggomez@soc.unicen.edu.ar
Pablo G. Messineo. CONICET, INCUAPA, Departamento de Arqueología, Facultad de Ciencias Sociales, UNCPBA, Av. Del Valle 5737 (B7400JWI) Olavarría, Buenos Aires, Argentina. E-mail: pmessine@soc.unicen.edu.ar
Recibido 30 de Junio 2006.
Aceptado 17 de Febrero 2007
RESUMEN
En este trabajo se presentan los resultados obtenidos del análisis cuantitativo y tafonómico de las especies de micro y mesomamíferos recuperadas en el sitio Laguna La Barrancosa 1 (Partido de Benito Juárez, provincia de Buenos Aires). Distintos métodos analíticos (macroscópicos y microscópicos -MEB-) fueron empleados en dicho análisis. Los índices utilizados para comparar la representación del esqueleto postcraneal frente al craneal (pc/c y f+h/md+mx) indican que existe una mayor representación de este último, señalando que determinados agentes post-deposicionales han producido la pérdida de algunas partes esqueletarias. Estos agentes y los efectos de la corrosión producida por las raíces, así como las condiciones sedimentarias, han sido los principales modificadores de la superficie cortical de los restos óseos de micro y mesomamíferos. En el conjunto óseo analizado no se han detectado efectos producidos por la digestión de depredadores ni marcas de carnívoros, indicando que los restos de estas especies fosoriales no fueron incorporados por ninguna cadena trófica. Además, no se registraron evidencias de la acción antrópica sobre los restos óseos (e.g., marcas de corte, huesos quemados, etc.) y se plantea que los mismos corresponden a especies que habitaron y murieron in situ. Las cuevas registradas en el sitio y las marcas de roedores sobre algunos de los huesos de Lama guanicoe señalan que estas especies actuaron como agentes modificadores post-depositacionales del conjunto arqueológico.
Palabras clave: Micro y mesomamíferos; Análisis tafonómico; Diagénesis.
ABSTRACT
Taphonomic Analysis Of Micro And Mesomammal Bones From Laguna La Barrancosa 1 Archaeological Site (District Of Benito Juárez, Buenos Aires Province) . In this paper, results are presented from quantitative and taphonomic analyses of micro and mesomammal bones recovered from Laguna La Barrancosa 1 archaeological site. The analytical methods used included macroscopic and microscopic (SEM) techniques. The rates used (pc/c y f+h/md+mx) show that there is a greater representation of cranial elements than postcranial due to diagenetical agents, which were the principal cause of the loss of some skeletal parts. These diagenetical agents, including the corrosion produced by roots and sedimentological conditions, were the principal agents of the cortical surface damage of the micro and mesomammal bones. There were no traces of digestion or other predators' marks indicating that the micromammal species were not incorporated into the archaeological record by a trophic chain. Also, there was no evidence of anthropic action on the micro and mesomammal bones (e.g., cut marks, burned bones, etc.). The evidence indicates that these species lived and died in situ. The caves recorded at the site and rodents' marks on Lama guanicoe bones indicate that these species acted as post-depositional modifying agents of the bone assemblages.
Keywords: Micro and mesommamals; Taphonomic analyses; Diagenesis.
INTRODUCCIÓN.
El estudio de las características diagenéticas en la superficie cortical de los restos óseos de micro y mesomamíferos es una vía analítica fundamental si se quiere obtener información acerca de los procesos de formación de los sitios arqueológicos bajo estudio y de su medio ambiente depositacional y de fosilización. En este sentido, estos análisis son imprescindibles para confirmar la ausencia/presencia de agentes perturbadores antes y después del enterramiento de los materiales, los cuales pueden llegar a modificar las interpretaciones que se hagan de los distintos sitios.
Este trabajo tiene como objetivo el análisis cualitativo y cuantitativo de los restos óseos de las especies de meso y micromamíferos hallados en las excavaciones realizadas en el sitio arqueológico Laguna La Barrancosa 1 (Partido de Benito Juárez, provincia de Buenos Aires, Argentina). Estos análisis tratan de evaluar los procesos de formación de sitio ocasionados en parte por distintas especies de micro y mesomamíferos que actuaron como agentes naturales sobre el registro arqueológico (sensu Schiffer 1983). Por otra parte, el estudio tafonómico pretende determinar las condiciones por las cuales los distintos individuos han sido incorporados al sitio arqueológico y los procesos diagenéticos que han sufrido hasta el momento de su recuperación.
El estudio de las características diagenéticas (e.g., la compactación mecánica, abrasión, marcas de raíces, entre otros) en la superficie cortical de los huesos puede proveer información acerca de la historia tafonómica del sitio arqueológico y su medio ambiente depositacional (Andrews 1990; Behrensmeyer 1978; Denys et al. 1996, 1997; Montalvo 2002). En general, los huesos de micromamíferos pueden ser incorporados al sitio por la actividad de un depredador, por la acción humana o bien, por la depositación natural de los mismos (Andrews 1990; Fernández-Jalvo 1992).
Durante las décadas pasadas, el interés para determinar las causas de la incorporación de los restos óseos de micro y mesomamíferos en los sitios arqueológicos se ha incrementado y, por este motivo, se han desarrollado diversas técnicas y métodos analíticos para su análisis (Andrews 1990; Denys et al. 1987, 1997; Fernández Jalvo 1992). En Argentina, los estudios sobre micromamíferos han estado orientados a su asociación con determinados eventos climáticos ocurridos durante el Pleistoceno final y Holoceno (Pardiñas 1991; Tonni 1992; Vizcaíno et al. 1995; entre otros), pero escasamente se ha desarrollado una tafonomía relacionada a pequeños mamíferos (Acosta y Pafundi 2005; Gómez 2000, 2005; Pardiñas 1999, 2000, 2001; Quintana 2005). En años recientes, el registro arqueológico en la región Pampeana ha proporcionado una gran muestra de restos óseos de meso y micromamíferos de los cuales se puede obtener na importante información tafonómica (Politis y Madrid 1988; Quintana 2004, 2005; Quintana y Mazzanti 2001; entre otros). En algunos sitios, como por ejemplo Laguna Tres Reyes 1 (Partido de González Cháves), el análisis tafonómico ha permitido, por un lado, diferenciar la actividad de determinados roedores en la formación del registro arqueológico del sitio como agentes post depositacionales evidenciado por el alto porcentaje de marcas en los huesos de guanaco (Politis y Madrid 1988) y, por el otro, determinar que la depredación o carroñeo producido por otros animales (e.g., zorros) ha incorporado partes esqueletarias de mesomamíferos en el contexto arqueológico del sitio (Gutiérrez y Gómez 2007).
UBICACIÓN GEOGRÁFICA Y CARACTERÍSTICAS DEL SITIO
El sitio Laguna La Barrancosa 1 (LLB1) se halla ubicado a los 37° 19' 39'' de latitud Sur y a los 60° 06' 40'' de longitud Oeste (Figura 1). Las primeras prospecciones se llevaron a cabo entre Diciembre de 2000 y Febrero de 2001 y en esa ocasión se realizó un sondeo de 1 x 0,50 m en el sector donde se registraron restos óseos aflorando en la barranca. Entre Agosto de 2001 y Marzo de 2002 se excavaron 18 cuadrículas de 1 x 1 m, las cuales totalizan una superficie de aproximadamente 18 m2 (Messineo 2003, 2008; Messineo y Pal 2006).
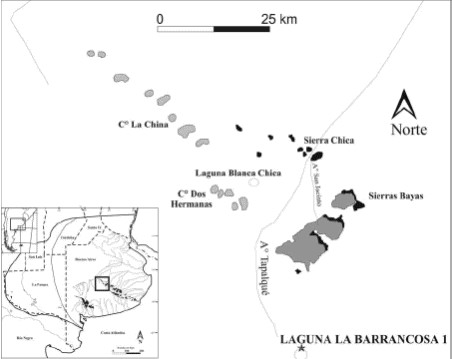
Figura 1: Ubicación geográfica del sitio Laguna La Barrancosa 1.
La excavación del sitio LLB1 se realizó siguiendo niveles artificiales de 5 cm a excepción de los niveles 2, 3 y 8 que fueron de 10 cm, debido a que los mismos contenían escasos materiales arqueológicos. Todos los materiales fueron registrados tridimensionalmente y se tamizó en agua todo el sedimento que se extrajo de la excavación, lo que permitió recuperar una gran cantidad de astillas óseas muy pequeñas, microdesechos líticos y huesos pertenecientes a micromamíferos y mesomamíferos (Messineo y Pal 2006; Pal 2006).
Durante las excavaciones realizadas se recuperaron restos óseos correspondiente a guanacos, materiales líticos de distintas materias primas y escasos fragmentos de cerámica. La distribución vertical de los materiales arqueológicos, tanto óseos como líticos, indican que la mayor densidad se encontraba en el nivel 6 (entre los 0,40 y 0,45 m de la superficie del terreno). Los niveles superiores e inferiores contenían pocos elementos arqueológicos y en algunas cuadrículas no se hallaron elementos culturales fuera de este nivel, evidenciando una escasa dispersión vertical de los materiales en el sitio (ver Pal 2006: Figura VI.1).
La distribución horizontal de los materiales (Figura 2) indica que existen diferentes patrones de distribución entre los materiales líticos y faunísticos, observándose áreas con mayor concentración de elementos faunísticos y áreas con mayor densidad de materiales líticos. Por otro lado, la distribución horizontal de los microdesechos líticos por materias primas (cuarcitas versus rocas silíceas) también muestra diferencias en su distribución (ver Pal 2006: Figuras VI.4 y VI.5).
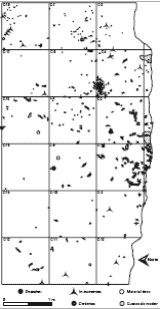
Figura 2: Plano de la excavación y distribución horizontal de los materiales arqueológicos (niveles 5, 6, 7 y 8 de excavación).
Los análisis desarrollados en el sitio LLB1 (faunísticos y tecnomorfológicos) estarían indicando que el mismo representaría un locus de actividades específicas en el cual se llevó a cabo el procesamiento secundario de guanaco. El alto porcentaje de fragmentación atribuido a la acción antrópica sugiere una intensiva explotación y aprovechamiento de las partes anatómicas que poseen un alto contenido de medula ósea. Con respecto a las materias primas líticas, se evidencia que la cuarcita (no local) ha sido la más utilizada en relación a otras rocas como por ejemplo la ftanita (local). Los instrumentos están representados por cuchillos y lascas con filos naturales con rastros complementarios que indicarían el empleo de los mismos en el procesamiento de los guanacos (Messineo 2003, 2008; Messineo y Pal 2006; Pal 2006).
Los análisis tafonómicos sobre las especies de mayor tamaño (guanaco) indican que las raíces (96,05%) y el deterioro químico (92,4%) son los principales agentes que modificaron la superficie cortical de los huesos. En la muestra analizada se evidenció que el 7,29% de los huesos presentaban huellas producidas por la acción de roedores y no se registraron marcas de carnívoros ni de pisoteo. Por su parte, la abrasión geológica sobre los elementos óseos sólo fue registrada en un 1,82% (Messineo 2003, 2008).
Con respecto a la distribución de los efectos provocados por la meteorización (sensu Behrensmeyer 1978) se observó que el 21,28% de los restos óseos presentaban un estadio 1 de meteorización, seguido en menores proporciones por los estadios 2 y 3 (8,51% y 3,04%, respectivamente) y un sólo elemento presentaba un estadio 4 de meteorización (0,3%). El resto de los elementos (66,87%) no presentaban meteorización (estadio 0) o ésta no pudo ser determinada debido a que la acción de las raíces y el deterioro químico afectaron la superficie cortical de los huesos (Messineo 2003, 2008).
GEOLOGÍA Y CRONOLOGÍA DEL SITIO LLB1
Los estudios estratigráficos, sedimentológicos, edafológicos y geomorfológicos han sido desarrollados por el Dr. Cristian Favier Dubois. En la margen norte de la laguna La Barrancosa (estratigrafía local) afloran sedimentos finos, arenosos, masivos de color castaño, de origen eólico, referibles a la Formación La Postrera. En ellos se registra la presencia de 3 ciclos principales de sedimentaciónpedogénesis, siendo el último el correspondiente al suelo actual. Estos tres ciclos pueden correlacionarse con los perfiles geológicos descriptos por Fidalgo et al. (1986) en la misma laguna.
La estratigrafía del sitio (Figura 3) refleja los tres ciclos principales desedimentación - pedogénesis observados en la estratigrafía local (0-0,40 m, 0,40-0,63 m y 0,63-+0,70 m). El último ciclo de pedogénesis (A1-A2-AC) corresponde al suelo actual, en parte afectado por actividades antrópicas (nivel 0-12 cm). El ciclo más antiguo (3BC) se halla erosionado, preservándose sólo el horizonte B o el BC y separado del ciclo intermedio por una discordancia de erosión. Este suelo evidenciaen algunos perfiles la estructura prismática y la presencia de cutanes típicas de un horizonte argílico (Bt), que indica un grado importante de desarrollo del mismo, correspondiendo su inicio probablemente al Holoceno temprano (Favier Dubois 2006).

Figura 3: Perfil estratigráfico del sitio Laguna La Barrancosa 1.
El ciclo intermedio de carácter cumúlico (2Acum), con mayor tasa de pedogénesis en su parte superior, es el que concentra la evidencia arqueológica. Un fechado de ![]() obtenido sobre una primera falange de Lama guanicoe dio una edad de 1676 ± 46 (AA-59507; hueso; δ13C= -20.214‰) que ubica cronológicamente al depósito en el Holoceno tardío (Messineo 2008; Messineo y Pal 2006).
obtenido sobre una primera falange de Lama guanicoe dio una edad de 1676 ± 46 (AA-59507; hueso; δ13C= -20.214‰) que ubica cronológicamente al depósito en el Holoceno tardío (Messineo 2008; Messineo y Pal 2006).
El análisis textural de las muestras sedimentarias del sitio concuerda con un sedimento de tipo loess que corresponde a la Formación La Postrera, siendo la textura franco arenoso y franco a franco arenoso, con una tendencia a la fracción arenosa en todo el perfil analizado (Tabla 1). El color de los sedimentos es homogéneo y en las muestras sedimentarias se halló carbonato pulverulento a partir del test de ácido acético (Favier Dubois 2006).
Tabla 1: Análisis sedimentológicos realizados al perfil estratigráfico del sitio LLB1.Número de muestras en relación con la Figura 3
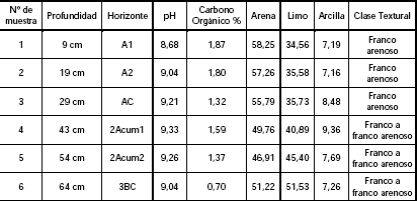
En las muestras se observa que los valores de Carbono orgánico son más elevados en el suelo actual (horizontes A1-A2) y en el ciclo intermedio de pedogénesis (2Acum1). Los porcentajes de arena diferencian, aunque no marcadamente, los tres ciclos de sedimentación-pedogénesis reconocidos. Los valores de pH indican condiciones fuertemente alcalinas a lo largo de toda la secuencia, pudiendo ser, al menos en parte, responsables del deterioro químico observado en la superficie cortical de los huesos recuperados en el sitio.
MÉTODOS
Para el análisis cuantitativo de los restos óseos de los micromamíferos, dada la dificultad de identificar a nivel de especie y de género una gran cantidad de los elementos óseos fracturados, se han considerado en conjunto los restos óseos de las diversas especies. Para calcular los distintos índices (e.g., abundancia relativa, etc.) se utilizó el MNE (Número Mínimo de Elementos) ya que permite una aproximación más exacta del posible número original de las partes anatómicas representadas. Dado el grado de fragmentación de los elementos óseos correspondientes a los micromamíferos se utilizó el MNE como un valor recompuesto para cada unidad anatómica,independientemente de la lateralidad (Binford 1984; Mengoni Goñalons 1999). Por su parte, para los incisivos que generalmente se encuentran muy fracturados se siguió el criterio propuesto por Klein y Cruz-Uribe (1984) según el cual los fragmentos son computados como fracciones. En el caso de incisivos los fragmentos son contados como un cuarto del elemento completo ya que de esta manera no se sobredimensiona a dicha unidad anatómica (ver Fernández-Jalvo 1992). El MNI (Número Mínimo de Individuos) fue calculado utilizando como base las distintas especies identificadas en el sitio y las unidades anatómicas más abundantes para cada una de las especies (e.g., dientes sueltos, hemimandíbulas, etc.).
Por último, la cuantificación de los elementos craneales fue realizada siguiendo la metodología empleada por Andrews (1990). En este caso, se contabilizan los especimenes óseos del cráneo en cuatro categorías debido a que estos huesos aparecer generalmente fracturados: cráneo entero, cráneo con zona cigomática completa, cráneo fracturado con arco cigomático y maxilar sin cigomático. Además, pueden aparecer fragmentos del cráneo sin ningún rastro de los maxilares o de la zona cigomática, por lo cual, los maxilares son cuantificados por separados de los fragmentos del cráneo.
Con el fin de determinar la representación anatómica y la abundancia relativa de las distintas partes esqueletarias de los micromamíferos se realizaron diferentes análisis cuantitativos. El índice de la abundancia relativa (Ri), tanto para cada una de las partes esqueletarias como para el esqueleto completo, fue calculado con la fórmula planteada por Andrews (1990):
donde: Ri: Abundancia relativa del elemento i; Ni: es el número del elemento i en la muestra; MNI: Número Mínimo de Individuos y Ei es el número del elemento i en el esqueleto de la presa.
Para el estimar las variaciones en la representación diferencial entre el esqueleto postcraneal y craneal se consideraron dos ecuaciones desarrolladas por Andrews (1990):
donde: pc: postcraneal; c: craneal.
Ambos índices miden el estado de preservación de los elementos tanto craneales como postcraneales, permitiendo realizar inferencias acerca de los procesos depositacionales y post-depositacionales que han sufrido los conjuntos óseos. Si ambas partes esqueletarias estuviesen representadas por igual el valor debe rondar el 100%.
En el cálculo de los distintos índices no se tuvo en cuenta a los marsupiales y armadillos por estar representados por escasos elementos y por tener fórmulas dentarias que no pueden ser aplicadas con el algoritmo pc/c de Andrews (1990). Dicho algoritmo fue planteado para micromamíferos con un premolar y dos molares por lo cual se lleva al valor de corrección a 16 para el cráneo (maxilares, mandíbulas y molares). En el caso de hallarse micromamíferos con un premolar y tres molares (e.g., algunos roedores caviomorfos) el valor de corrección de la fórmula 16 debería ser reemplazado con un valor de corrección de 20.
Por ejemplo, algunas especies de Ctenomys poseen un premolar y dos molares mientras que otras especies poseen un premolar y tres molares (un 3º molar de origen vestigial) que cuando se presentan aislados en la muestra son prácticamente indistinguibles. En la región Pampeana se encuentran presentes tanto Ctenomys talarum como Ctenomys australis. Esta última especie se halla distribuida sobre todo en la costa bonaerense y es la que posee un premolar y tres molares (Contreras y Reig 1965). Por su parte, Ctenomys talarum es la especie que tiene una mayor distribución hacia el interior de la provincia de Buenos Aires, prefiriendo zonas con una mayor vegetación y suelos firmes. Esta especie es la que aparece con un premolar y dos molares y la que tuvo una amplia distribución durante el Pleistoceno final y el Holoceno en la zona oriental de las Sierras de Tandilia (Contreras y Reig 1965; Quintana 2004).
En este sentido, cuando es posible diferenciar entre especies que poseen distinta fórmula dentaria debería utilizarse distintos valores de corrección para la fórmula de Andrews. Sin embargo, debido al grado de fragmentación que presentaban los materiales óseos de los caviomorfos del sitio fue imposible diferenciar entre las especies de Ctenomys que poseen distintas fórmulas dentarias (C. australis o C. talarum). Debido a esto, seha realizado el análisis de los micromamíferos en conjunto (tanto de cricétidos como de caviomorfos) y el valor de corrección utilizado para la fórmula pc/c fue de 16 (sensu Andrews 1990).
Por último, la pérdida de las partes distales de los elementos apendiculares fue estimada en este trabajo usando la siguiente ecuación (Andrews 1990):
Para el análisis tafonómico se consideraron las variables que varios autores han propuesto para el estudio de los micromamíferos y mesomamíferos presentes en los contextos arqueológicos (Andrews 1990; Andrews y Fernández-Jalvo 1997; Behrensmeyer 1978; Binford 1981; Fernández-Jalvo 1992; Gifford-González 1989; Gómez y Kaufmann 2007; Lyman 1994; Reitz y Wing 1999; Shipman 1981; Shipman y Rose 1983; Trolle-Lassen 1986; Zeiler 1987). En este trabajo, se han identificado sobre la superficie de los huesos las siguientes variables: fracturas, pisoteo, abrasión, manchas de óxido de manganeso, meteorización, marcas de raíces, corrosión producida por distintos agentes (depredadores, raíces y suelo), descamación y depresiones. La descripción de los distintos criterios para la identificación de estos caracteres se puede hallar en distintos artículos (Andrews 1990; Fernández-Jalvo 1992; Gómez 2000; entre otros).
El análisis tafonómico de los huesos de micro y mesomamíferos fue llevado a cabo usando la metodología desarrollada por Andrews (1990) y revisada por Fernández-Jalvo (1992). Esta metodología está basada en el análisis cualitativo y cuantitativo de los elementos esqueletarios de estas especies. También han sido considerados los valores de digestión, fracturación y representación anatómica obtenidos para depredadores sudamericanos actuales (ver Gómez 2000; Gómez y Kaufmann 2007).
La metodología de investigación también incluye el uso de imágenes realizadas por microscopio electrónico debarrido (MEB) de una muestra de los elementos óseos de micromamíferos provenientes de distintos niveles de excavación del sitio LLB1. El microscopio electrónico de barrido utilizado fue un FEI Quanta 200 con cámara ambiental que trabaja con tres niveles de vacío (bajo, alto y ambiental), con detectores electrónicos secundarios y retrodispersados para todos los niveles de vacío. Este microscopio tiene un sistema integrado de análisis Oxford Instruments Analytical - INCA con dos sistemas de detectores de rayos X. El microscopio utilizado se encuentra en el Museo Nacional de Ciencias Naturales, CSIC, Madrid, España.
RESULTADOS
Representación de especies y partes anatómicas
Como producto de las excavaciones y de los análisis cuantitativos realizados se obtuvo un NSP (número total de especimenes óseos identificables y no identificables) (sensu Grayson 1991) de 5772, de los cuales 818 (NISP: número de especimenes óseos identificables) han podido ser determinados, ya sea al nivel de clase, orden, familia o especie y, el resto (4954) corresponden a especimenes óseos no asignados a ningún taxón y se hallan representados por astillas de huesos muy pequeñas (Tabla 2).
Tabla 2: Cuantificación de las especies representadas en el sitio LLB1 (Tomado de Messineo 2008).

Con respecto a los especimenes óseos identificados (NISP total) se determinó que 656 correspondían a Artiodactyla de los cuales 353 pertenecían exclusivamente a guanaco (Lama guanicoe), en tanto que el resto (303) se determinó al nivel de género (Lama sp.). Además, en los niveles superiores ubicados, en el suelo actual, se hallaron, entre las especies de tamaño grande, dos ejemplares óseos no identificados posiblemente de fauna exótica (Tabla 2).
Por otro lado, se registró la presencia de taxones que pertenecían a mesomamíferos (n= 15) y micromamíferos (n= 144) (sensu Andrews 1990). Para los mesomamíferos la única especie determinada fue Chaetophractus villosus (peludo) y para los micromamíferos se identificaron Ctenomys sp. (tucotuco), Reithrodon sp. (ratón conejo), posiblemente Akodon (ratón pajizo), un Cricétido indeterminado y un marsupial. En el nivel 7 del sitio se halló una vértebra perteneciente a un pez que fue determinado como posible Basilichthys (pejerrey) y cuya introducción se debe a factores naturales y no fue tenida en cuenta en los análisis (Tabla 2).
Muchos de los restos óseos analizados se encuentran fracturados, por lo cual, sólo se los ha podido asignar como pertenecientes a roedores, micromamíferos o mesomamíferos no identificados. Por un lado, entre los micromamíferos identificados, se destacan los roedores de la familia Octodontidae, cuyo mayor porcentaje corresponde a Ctenomys sp. (29,6%). Dentro de la subfamilia Sigmodontinae, Reithrodon sp. representa un 12,6% y Akodon sp. el 3,1% del total. También se han detectado dos molares de un marsupial no identificado (1,3%) y un elemento correspondiente a un Cricétido no identificado (0,6%). Un 16,4% de los restos óseos han sido clasificados como especimenes pertenecientes a roedores indeterminados y un 27% a micromamíferos indeterminados (Tabla 2).
Por otro lado, los restos óseos de mesomamíferos son muy escasos y difíciles de identificar al nivel de especie debido al grado de fragmentación. La única especie determinada en el sitio corresponde a Chaetophractus villosus (placas dérmicas y una hemipelvis) con el 3,1%, el 1,3% fue asignado a restos óseos de armadillos no identificados (falanges y metápodos) y el 5% pertenece a mesomamíferos indeterminados (vértebras, astrágalos, falanges y metápodos) (Tabla 2).
Los elementos óseos mejor representados del conjunto de micromamíferos corresponden a los incisivos, maxilares y al esqueleto craneal. En el esqueleto postcraneal, los elementos óseos más numerosos son los fémures, seguidos por los húmeros y en menores porcentajes el resto del esqueleto (Tabla 3; Figura 4).
Tabla 3: MNE (Número Mínimo de Elementos) y abundancia relativa de las distintas partes esqueletarias de los micromamíferos hallados en el sitio LLB1.
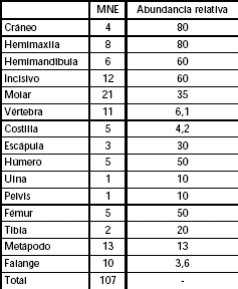
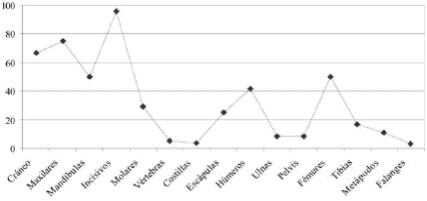
Figura 4: Abundancias relativas de las partes esqueletarias de los micromamíferos hallados en el sitio.
Los micromamíferos se hallan representados por un MNI= 5 (no incluidos los marsupiales y los armadillos). Este número está compuesto por dos individuos de Reithrodon (dos mandíbulas) y un individuo del resto de las especies identificadas (Ctenomys sp., Akodon sp. y Cricétido indeterminado). La baja abundancia relativa de los elementos óseos recuperados de micromamíferos en relación al MNI indican que existe una baja supervivencia (28,9%) de las partes esqueletarias de las especies halladas en el sitio. Por otra parte, la abundancia relativa de las distintas partes esqueletarias señala que los índices más altos pertenecen a elementos craneales, en tanto que, entre los elementos del esqueleto postcraneal, los mayores valores corresponden a húmeros y fémures (Tablas 3 y tabla 4; Figura 4).
Tabla 4: Valores absolutos y relativos de los índices y de las variables tafonómicas consideradas en el análisis de los micromamíferos (no se incluyeron los elementos óseos de los marsupiales y armadillos).
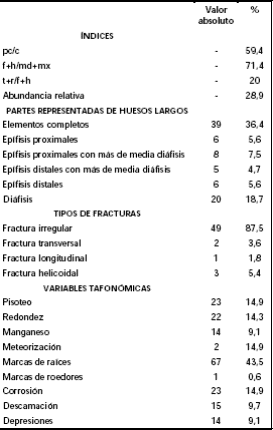
El índice pc/c obtenido fue de 59,4% lo que revela una mayor presencia del esqueleto craneal con relación al postcraneal. Por otra parte, el índice f+h/ md+mx, representa un 71,4%, lo que confirma la tendencia del índice anterior en el cual se observa una mayor presencia de los elementos craneales por sobre los postcraneales (Tabla 4). El resultado del índice t+r/ f+h (tibia+radio/fémur+húmero) es muy bajo (20%) mostrando que hay una importante pérdida de las partes distales del esqueleto apendicular de los micromamíferos.
La totalidad de los incisivos y molares se han hallado fuera de los alvéolos de las mandíbulas y maxilares, indicando que han existido procesos diagenéticos (e.g., abrasión y actividad de raíces) que han desmontado los elementos dentarios. El hallazgo completamente aislado de los dientes también se puede deber a la actividad excavadora y post-excavadora (e.g., cernido y limpieza de los materiales).
Análisis tafonómicos y diagenéticos
El 47,6% de los restos óseos recuperados se hallan representados por elementos craneales, mientras que el 52,3% corresponde al esqueleto postcraneal. Del total de los elementos postcraneales de los micromamíferos un 36,4% corresponde a huesos largos completos, el 42,1% está representado por distintas porciones óseas de los huesos largos (epífisis proximales, epífisis distales y diáfisis) y el 21,5% restante corresponde a otros elementos del esqueleto (e.g., falanges). Del total del material postcraneal fracturado (n= 55), se ha registrado un importante porcentaje de elementos con fracturas irregulares (87,5%), en tanto que para el resto se identificaron fracturas helicoidales (5,4%), transversales (3,6%) y longitudinales (1,8%) (Tabla 4).
De las variables tafonómicas consideradas (Tabla 4; Figura 5), no se han encontrado evidencias de digestión y las marcas producidas por las raíces se hallan representadas con el 43,5% (Figuras 6-9). Dichas marcas se registraron en los elementos óseos provenientes de los distintos niveles de excavación, estando los mayores porcentajes representados en los niveles 5 y 6. Los porcentajes de corrosión producidas por las raíces y/o el suelo (descartando la digestión por depredadores) y las marcas de pisoteo en los huesos de micromamíferos y mesomamíferos alcanzan un 14,9% cada una.
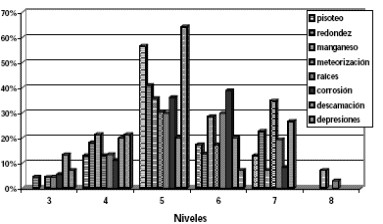
Figura 5: Representación de las variables tafonómicas por niveles artificiales de excavación.
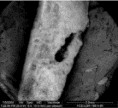
Figura 6: Imagen de un fragmento de diáfisis de micromamífero afectado por la acción de las raíces en la cual se observa una perforación (MEB).
Cabe destacar que el 14,9% de los huesos presentaban señales de meteorización. Los huesos con signos de meteorización se hallaban en todos los niveles de excavación excepto el 8 y los mayores porcentajes estaban representados en los niveles 5 y 7. Para determinar los distintos grados de meteorización se siguieron los cuatro estadios establecidos por Andrews (1990). El estadio 1 de meteorización es el más representado (n= 22) observándose una ligera fracturación paralela a la estructura fibrosa de los huesos y de la dentina (Figuras 7 y 8). Sólo un elemento alcanza el estadio 2 de meteorización (incisivo de Ctenomys sp.), en el cual se observa un agrietamiento y un astillamiento más extendido y una pérdida casi completa de la corona.
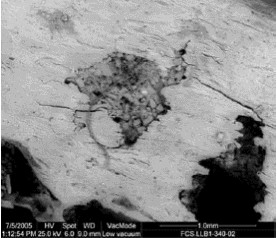
Figura 7: Superficie cortical de un hueso afectado por raíces y por la acción corrosiva de éstas que han llegado a producir perforaciones (MEB). La superficie se encuentra ondulada y una de las fracturas, no visible en la fotografía, está asociada a una raíz.

Figura 8: Incisivo de roedor con agrietamiento en la dentina posiblemente afectado por las raíces y por la acción química (alcalina) de los sedimentos (MEB). Dentina y esmalte resquebrajados principalmente en los bordes. No se observa modificación en el esmalte. El pH medido para los sedimentos del sitio varia entre 8,68 y 9,33 (Tabla 1).
La corrosión detectada en el conjunto óseo (14,9%) ya sea por raíces o de origen diagenético se distribuyen desde los niveles 3 al 7, siendo los mayores porcentajes en los niveles 5 y 6. En ciertos elementos pueden distinguirse claramente los surcos o perforaciones producidos por la acción de las raíces, mientras que en otros casos la corrosión superficial causada por procesos diagenéticos es leve y generalizada en todo el elemento.
Un 14,3% de los materiales se encuentra redondeados, sobre todo en los bordes de fractura y asociados, en muchos casos, a las marcas de abrasión. Tanto los bordes redondeados como la abrasión pudieron ocasionarse por procesos postdepositacionales, durante la formación de la laguna. Las marcas de abrasión en los restos óseos se han hallado entre los niveles 4 y 7, en tanto que los huesos con evidencias de redondeo poseen una distribución vertical entre los niveles 3 y 7. Tanto la abrasión como los bordes redondeados sobre la superficie cortical de los huesos se hallan representados en mayores porcentajes en el nivel 5 de excavación.
Otra de las variables observadas en los análisis tafonómicos y diagenéticos es la descamación, la cual se registró en el 9,7% de los restos óseos analizados y puede producirse debido a la acción de la meteorización. La distribución de elementos con descamación se halla entre los niveles 3 y 7, estando el mayor porcentaje en el nivel 7. Las dendritas de óxido de manganeso se encuentran en un 9,1% de los elementos y las mismas se concentran en el nivel 5, aunque tiene una distribución que va desde los niveles 4 al 8. La presencia de manchas de óxido de manganeso se puede deber en parte a que los elementos óseos han estado en ambientes de reducción de este mineral (e.g., ambientes saturados de agua, con poco oxígeno o sedimentos saturados de humedad). Al igual que el manganeso, las depresiones fueron registradas en el 9,1% de los restos óseos. Esta modificación sobre la superficie cortical de los huesos se manifiesta como consecuencia de la acción corrosiva de distintos agentes (e.g., las raíces) y, en algunos casos, la alcalinidad del suelo produce efectos similares a la meteorización, comenzando con una descamación y produciendo una superficie ondulada sin un agrietamiento previo. Los porcentajes de elementos con depresiones son muy pocos y su distribución se registra entre los nivel 3 y 6, con una mayor concentración en el nivel 5.
No se han encontrado marcas de carnívoros y sólo se registraron marcas de roedores sobre la superficie cortical de un hueso de micromamífero indeterminado (0,6%) procedente del nivel 6 de excavación. Por último, no se hallaron marcas de cortes ni huesos quemados sobre los elementos óseos de micro y mesomamíferos que indicasen el procesamiento de este tipo de especies por parte de los grupos cazadoresrecolectores que ocuparon el sitio.
El material de micromamíferos recuperado en el sitio arqueológico LLB1 se encuentra distribuido en todos los niveles de excavación, aunque su mayor densidad se registra en los niveles 5 y 6 (Figura 5). Estos niveles corresponden al techo del ciclo (2Acum1) en el cual se registra una mayor tasa de pedogénesis. El mayor porcentaje de las variables tafonómicas consideradas en el análisis de los materiales de micromamíferos se encuentran en el nivel 5 de excavación. Por el contrario, los escasos materiales óseos de mesomamíferos hallados, se han recuperado en los niveles superiores de excavación y en las cuadrículas que poseían galerías, no estando los mismos relacionados con el depósito arqueológico. En este sentido, los restos óseos de los micro y mesomamíferos corresponderían a una mezcla tafonómica (ver Fernández López 1986) de los restos óseos más recientes en niveles arqueológicos más antiguos producida por la etología de los propios animales cavadores.
DISCUSIÓN Y CONCLUSIONES
Los análisis cuantitativos señalan una escasa representación para las distintas especies de micromamíferos (NISP= 144) y mesomamíferos (NISP= 15). Los restos óseos de estas especies no son abundantes por lo que hay que considerar a los procesos diagenéticos que han ocurrido en el sitio luego de su formación como uno de los factores principales que han producido la pérdida de algunas partes anatómicas. A su vez, los distintos índices utilizados (pc/c y f+h/md+mx) indican que existe una mayor representación del esqueleto craneal sobre el postcraneal, señalando que algunos agentes diagenéticos han producido dicha pérdida.
Es claro que los restos óseos han sufrido distintos procesos post-depositacionales ya que es importante el porcentaje de elementos que registran marcas de raíces y fracturas irregulares asociadas a los distintos agentes. Las marcas de raíces y las evidencias de corrosión sobre los huesos presentaban una variada gama de intensidades (Figura 7). La corrosión observada pudo ser ocasionada por las raíces y acentuada por los ácidos húmicos que, junto a la actividad de hongos y bacterias, son los responsables directos de la disolución de los minerales de los huesos (e.g., fosfato y calcio).
Estos datos concuerdan con aquellos obtenidos de los análisis tafonómicos de las especies de mayor tamaño (guanaco) en los cuales se observa que la acción de las raíces fue el agente principal en la alteración de la superficie cortical de los huesos (Messineo 2003, 2008). La acción de las raíces estaría relacionada con la formación de un suelo de tipo cumúlico (horizonte 2Acum1) en el cual se hallaría una continua presencia de la cubierta vegetal.
El material hallado con signos de meteorización indica que un porcentaje de los huesos estuvo expuesto, quizás por un corto período de tiempo. Los huesos de micromamíferos con signos de meteorización apenas alcanzan un 10% y no sobrepasan el primer estadio (Andrews 1990). El material con manchas de óxido de manganeso (ca. 10%) indica su asociación con un ambiente húmedo, anaeróbico y este proceso diagenético generalmente se produce en sedimentos saturados de agua (e.g., suelo) donde el manganeso pueda oxidarse y quedar registrado en los huesos.
No hay evidencia de digestión por depredadores (e.g., zorros, aves, etc.) en los elementos esqueletarios de las especies de micromamíferos y mesomamíferos, ni tampoco existen evidencias firmes (e.g., marcas de carnívoros) de que estos pequeños mamíferos hayan sido depositados debido a la acción de algunos depredadores tales como los zorros o aves de presa nocturnas que habitan galerías (e.g., Athene cunicularia) u otros pequeños carnívoros como los zorrinos o hurones. Estos datos indican que los restos de estas especies fosoriales no fueron incorporados al sitio por ninguna cadena trófica.
Los estudios tafonómicos realizados sobre los huesos de guanaco indican que la acción de los roedores (marcas sobre la superficie cortical) se halla representada en un 7,29% de la muestra analizada, señalando que los mismos actuaron con posterioridad a la depositación del material arqueológicos (Messineo 2003, 2008). Además, se registraron en las excavaciones cuatro cuevas de roedores (bocas de túneles) pertenecientes por sus dimensiones a pequeños micromamíferos. Los análisis distribucionales desarrollados indican que los roedores no produjeron cambios significativos en la distribución vertical y horizontal de los materiales arqueológicos (ver discusión en Pal 2006).
El análisis tafonómico del material óseo de micro y mesomamíferos permite concluir que la mayor parte de las especies registradas habitaron el sitio, más probablemente luego del abandono por parte del grupo humano, ya que sobre estos materiales no se hallaron marcas de corte y huesos con evidencias de combustión que denoten una actividad antrópica. Las condiciones ambientales y sedimentológicas registradas en el sitio son propicias para que estas especies de roedores caviomorfos (e.g., Ctenomys) y de especies de mesomamíferos (e.g., armadillos) hayan habitado naturalmente en el sitio. En este sentido, los restos de micromamíferos y mesomamíferos hallados pertenecen a individuos que han vivido en el lugar, construyendo su red de galerías, las cuales poseen las características propias de las que realizan algunas especies de Ctenomys y, por lo tanto, estos restos corresponden a especies que han muerto in situ en momentos posteriores a la depositación de los materiales arqueológicos.
Agradecimientos
Queremos expresar nuestro agradecimiento al programa INCUAPA (Investigaciones Arqueológicas y Paleontológicas del Cuaternario Pampeano) de la Secretaría de Ciencia y Tecnología de la Universidad Nacional del Centro de la Provincia de Buenos Aires y su Director Dr. Gustavo Politis y Codirector, Lic. José L. Prado por apoyar nuestras investigaciones. Parte de este trabajo fue realizado en el marco de una estadía de investigación corta en el Museo Nacional de Ciencias Naturales de Madrid (España) a través de una beca otorgada por la Fundación Carolina a uno de los autores (G.N.G) y financiado por el PICT 04-12777 de la ANPCYT dirigido por el Dr. G. Politis. Agradecemos a la Dr. María Teresa Alberdi por sus comentarios y lectura crítica del manuscrito. Por último, deseamos agradecer a los dos evaluadores del trabajo.
REFERENCIAS CITADAS
Acosta, A. y L. Pafundi 2005 Zooarqueología y tafonomía de Cavia aperea en humedal del Paraná inferior. Intersecciones en Antropología 6: 59-74. [ Links ]
Andrews, P. 1990 Owls, Caves and Fossils. Natural History Museum Publications, Londres. [ Links ]
Andrews, P. y Y. Fernández-Jalvo 1997 Surface modifications of the Sima de los Huesos fossil humans. Journal of Human Evolution 33: 191-217. [ Links ]
Behrensmeyer, A. K. 1978 Taphonomic and ecological information from bone weathering. Paleobiology 4 (2): 150-162. [ Links ]
Binford, L. R. 1981 Bones: Ancient Men and Modern Myths. Academic Press, Nueva York. [ Links ]
Binford, L. R. 1984 Faunal Remains from Klasies River Mouth. Academic Press, Nueva York. [ Links ]
Contreras, J. y O. Reig 1965 Datos sobre la distribución del género Ctenomys (R. O.) en la zona costera de la provincia de Buenos Aires comprendida entre Necochea y Bahía Blanca. Phycis 25 (69): 169-186. [ Links ]
Denys, C., D. Geraards, J. J. Hublin y H. Tong 1987 Méthode d'étude taphonomique de microvertébrés. Application au site Pleistocène de Tighenif (Ternifine, Algérie). Archaeozoologia 1: 53-82. [ Links ]
Denys, C., C. T. Williams, Y. Dauphin, P. Andrews y Y. Fernández-Jalvo 1996 Diagenetical changes in Pleistocene small mammal bones from Olduvai Bed I. Palaeogeography, Palaeoecology, Palaeoeclimatology 126: 121-134. [ Links ]
Denys, C., P. Andrews, Y. Dauphin, T. Williams y Y. Fernández-Jalvo 1997 Towards a site classification: comparison of stratigraphic, taphonomic and diagenetic patterns and processes. Bulletin Societè Géologique 168 (6): 751- 757. [ Links ]
Favier Dubois, C. 2006 Informe Geológico y Sedimentológico del Sitio Laguna La Barrancosa 1. Departamento de Arqueología, Facultad de Ciencias Sociales, Universidad Nacional del Centro de la Provincia de Buenos Aires, Olavarría. MS. [ Links ]
Barrientos G. y F. Oliva 1997 Investigaciones arqueológicas en el sitio Gascón 1, Patido de Adolfo Alsina, Provincia de Buenos Aires Resúmenes del XII Congreso Nacional de Arqueología Argentina, p. 21 La Plata
Fernández-Jalvo, Y. 1992 Tafonomía de microvertebrados del complejo cárstico de Atapuerca (Burgos). Tesis Doctoral inédita. Departamento de Paleontología Facultad de Ciencias Geológicas, Universidad Complutense de Madrid, Madrid, España. [ Links ]
Fernández López, S. 1986 Sucesiones paleobiológicas y sucesiones registráticas. Revista Española de Paleontología 1: 29-45. [ Links ]
Fidalgo, F., R. O. Gentile y H. A. Correa 1986 Geología y geomorfología en la cuenca del Arroyo Tapalqué. Informe 30. Presentado a la Comisión de Investigaciones Científicas de la Provincia de Buenos Aires, La Plata. MS. [ Links ]
Gifford-González, D. P. 1989 Etnographic analogues for interpreting modified bones: some cases from East Africa. En Bone Modification, editado por R. Bonnichsen y M. H. Sorg, pp. 179-246. University of Maine, Center for the Study of the First Americans, Orono. [ Links ]
Gómez, G. N. 2000 Análisis tafonómico y paleoecológico de los micro y mesomamíferos del sitio arqueológico de Arroyo Seco 2 (Buenos Aires, Argentina) y su comparación con la fauna actual. Tesis Doctoral inédita. Departamento de Biología Animal I, Facultad de Ciencias Biológicas, Universidad Complutense de Madrid, Madrid, España. [ Links ]
Gómez, G. N. 2005 Analysis of bone modification of Bubo virginianus pellets from Argentina. Journal of Taphonomy 3 (1): 1-16. [ Links ]
Gómez, G. N. y C. A. Kaufmann 2007 Taphonomic analysis of Pseudalopex griseus (Gray, 1837) scat assemblages and their archaeological implications. Journal of Taphonomy 5 (2): 59-70. [ Links ]r
Grayson, D. K 1991 Alpine faunas from the White Mountains, California: adaptive change in the Late Prehistoric Great Basin? Journal of Archaeological Science 18: 483-506. [ Links ]
Gutiérrez, M. A. y G. N. Gómez 2007 Taphonomic analysis of micro y mesomammals from Tres Reyes 1 archaeological site (Pampean region, Argentina). En Taphonomy and Zooarchaeology in Argentina, editado por M. A. Gutiérrez, L. Miotti, G. Barrientos, G. Mengoni Goñalons y M. Salemme, pp. 105-119. BAR International Series 1601. Archaeopress, Oxford. [ Links ]
Klein, R. y C. Cruz-Uribe 1984 The Analysis of Animal Bones from Archaeological Sites. University of Chicago Press, Chicago. [ Links ]
Lyman, R. L. 1994 Vertebrate Taphonomy. Cambridge Manuals in Archaeology. Cambridge University Press, Cambridge. [ Links ]
Mengoni Goñalons, G. 1999 Cazadores de Guanacos de la Estepa Patagónica. Colección de Tesis Doctorales, Sociedad Argentina de Antropología, Buenos Aires. [ Links ]
Messineo, P. G. 2003 Análisis arqueofaunísticos en el sitio Laguna La Barrancosa 1 (Pdo. de Benito Juárez, Pcia. de Buenos Aires, Argentina). Archaeozoology: International Journal of Archaeozoology 12: 73-86. [ Links ]
Messineo, P. G. 2008 Investigaciones arqueológicas en la cuenca superior del Arroyo Tapalqué (Partidos de Olavarría y Benito Juárez, Provincia de Buenos Aires). Tesis Doctoral inédita. Facultad de Ciencias Naturales y Museo, Universidad Nacional de La Plata, La Plata. [ Links ]
Messineo, P. G. y N. Pal 2006 El Holoceno tardío en la cuenca superior del Arroyo Tapalqué (Partidos de Olavarría y Benito Juárez). En INCUAPA 10 Años. Perspectivas Contemporáneas en la Arqueología de las Regiones Pampeana y Norpatagónica, editado por G. G. Politis. Serie Monográfica Nº 6, INCUAPA (FACSOUNICEN), Olavarría. En prensa. [ Links ]
Montalvo, C. I. 2002 Root traces in fósil bones from tha Huayquerian (Late Miocene) faunal assemblege of Telén, La Pampa, Argentina. Acta Geológica Hispánica 37 (1): 37-42. [ Links ]
Pal, N. 2006 Aportes al estudio de la integridad del sitio Laguna La Barrancosa 1 (Partido de Benito Juárez, Provincia de Buenos Aires): Análisis de la distribución espacial, textural y tecnomorfológico de los microdesechos líticos. Tesis de Licenciatura inédita. Facultad de Ciencias Sociales, Universidad Nacional del Centro de la Provincia de Buenos Aires, Olavarría. [ Links ]
Pardiñas, U. F. 1991 Roedores, marsupiales y edentados en la secuencia de Fortín Necochea, General Lamadrid, Buenos Aires. Implicancias paleoambientales. Boletín del Centro 2: 139-153. [ Links ]
Pardiñas, U. F. 1999 Fossil murids: taxonomy, paleoecology, and paleoenvironments. En Quaternary of South America and Antarctic Peninsula 12, editado por J. Rabassa y M. Salemme, pp. 225-254. A. A. Balkema, Rotterdam, Bookfield. [ Links ]
Pardiñas, U. F. 2000 Tafonomía de microvertebrados en yacimientos arqueológicos de Patagonia (Argentina). Arqueología 9: 264-340. [ Links ]
Pardiñas, U. F. 2001 Condiciones áridas durante el Holoceno temprano en el sudoeste de la provincia de Buenos Aires (Argentina): vertebrados y tafonomía. Ameghiniana 38 (3): 227-236. [ Links ]
Politis, G. G. y P. E. Madrid 1988 Un hueso duro de roer: análisis preliminar de la tafonomía del sitio Laguna Tres Reyes 1 (Pdo. de Adolfo Gonzáles Cháves, provincia de Buenos Aires). En De Procesos, Contextos y Otros Huesos, editado por N. Ratto y A. Haber, pp. 29-44. Instituto de Ciencias Antropologías. Universidad Nacional de Buenos Aires, Buenos Aires. [ Links ]
Quintana, C. A. 2004 El registro de Ctenomys talarum durante el Pleistoceno tardío - Holoceno de las Sierras de Tandilia Oriental. Mastozoología Neotropical. Journal of Neotropical Mammalogy 11 (1): 45-53. [ Links ]
Quintana, C. A. 2005 Especies de microroedores en el Holoceno tardío las Sierras de Tandilia (Argentina). Archaeofauna 217: 227-241. [ Links ]
Quintana, C. A. y D. Mazzanti 2001 Selección y aprovechamiento de recursos faunísticos. En Cueva Taxi: Cazadores y Recolectores de las Sierras de Tandilia Oriental. Geología, Paleontología y Zooarqueología, editado por D. Mazzanti y C. A. Quintana, pp. 181-209. Laboratorio de Arqueología, Universidad Nacional del Mar del Plata, Publicación Especial I, Mar del Plata. [ Links ]
Reitz, E. J. y E. S. Wing 1999 Zooarchaeology. Cambridge Manuals in Archaeology, Cambridge University Press, Cambridge. [ Links ]
Schiffer, M. 1983 Toward the identification of formation processes. American Antiquity 48: 675-706. [ Links ]
Shipman, P. 1981 Life History of a Fossil. Harvard University Press, Cambridge. [ Links ]
Shipman, P. y J. J. Rose 1983 Evidence of butchery and hominid activities Torralba and Ambrona: An evaluation using microscopic techniques. Journal Archaeological Science 10: 465-474. [ Links ]
Tonni, E. P. 1992 Mamíferos y clima del Holoceno en la provincia de Buenos Aires. En El Holoceno en la Argentina 1, editado por M. Iriondo, pp. 64-78. CADINQUA, Buenos Aires. [ Links ]
Trolle-Lassen, T. 1986 Human exploitation of the Pine Marten [Martes martes (L.)] at the Late Mesolithic settlement of Tybrind Vig in Western Funen. En Nordic Late Quaternary Biology and Ecology, editado por L. K. Königsson, pp. 119-124. Striae, vol. 24, University of Aarhus, Dinamarca. [ Links ]
Vizcaíno, S. F., U. F. Pardiñas y M. S. Bargo 1995 Distribución de los armadillos (Mammalia Dasypodidae) en la región Pampeana (República Argentina) durante el Holoceno. Interpretación paleoambiental. Mastozoología Neotropical. Journal of Neotropical Mammalogy 2 (2): 149-166. [ Links ]
Zeiler, J. T. 1987 Exploitation of fur animals in Neolithic Swifterbant and Hazendonk (Central and Western Netherlands). Palaeohistoria 29: 245-261. [ Links ]

