










Servicios Personalizados
Revista

Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkIntersecciones en antropología
versión On-line ISSN 1850-373X
Intersecciones antropol. n.9 Olavarría ene./dic. 2008
Palinología arqueológica: su implicancia en el estudio del prehispánico tardío de las sierras de Córdoba (Argentina)
Matías E. Medina, Silvia C. Grill y M. Laura López
Matías E. Medina. Laboratorio de Prehistoria y Arqueología. Pabellón Argentina, Ala Sud, Ciudad Universitaria. 5000 Córdoba. E-mail: medina@comechingonia.com
Silvia C. Grill. Cátedra Geomorfología y Teledetección, Laboratorio de Palinología, Departamento de Geología, Universidad Nacional del Sur, San Juan 670. B8000ICN Bahía Blanca. E-mail: sgrill@criba.edu.ar
M. Laura López. FONCyT. Laboratorio de Prehistoria y Arqueología. Pabellón Argentina, Ala Sud, Ciudad Universitaria. 5000 Córdoba. E-mail: lauralopez@comechingonia.com
Recibido 20 de Mayo 2008.
Aceptado 23 de Junio 2008
RESUMEN
Cuatro sitios arqueológicos correspondientes al Periodo prehispánico tardío de las sierras de Córdoba (ca. 1000-300 AP): Los Algarrobos 1, Puesto La Esquina 1 (Pampa de Olaen), C.Pun.39 y Las Chacras 2 (valle de Punilla), fueron estudiados en base a su contenido palinológico. Los sitios Los Algarrobos 1 y Puesto La Esquina 1 registraron, como consecuencia de sesgos tafonómicos, insuficiente contenido polínico. En cambio, las asociaciones polínicas de C.Pun.39 y Las Chacras 2 permitieron inferir paleo-comunidades vegetales dominadas por Chenopodiaceae-Amaranthaceae y Brassicaceae (C.Pun. 39) y por el primer taxón (Las Chacras 2). Las dos familias vegetales tienen baja representación en la lluvia polínica actual. Se postula la posibilidad de que las significativas proporciones de Chenopodiaceae-Amaranthaceae podrían estar sugiriendo la presencia de cultivos prehispánicos de Chenopodium quinoa y Amaranthus caudatus en los alrededores de los sitios. Otras evidencias de macro y microrrestos ligadas a cultivos sustentan dicha hipótesis. Las importantes proporciones tanto de Chenopodiaceae-Amaranthaceae como de Brassicaceae registradas en C.Pun.39 son también analizadas como indicativas de sucesivos abandonos y reocupaciones de los sitios, coherentes con un patrón de alta movilidad residencial entre los grupos prehispánicos tardíos. Ambientes perturbados por actividad antrópica habrían contribuido al desarrollo de tales plantas.
Palabras clave: Palinología; Sitios Arqueológicos; Periodo prehispánico tardío.
ABSTRACT
Archaeological Palynology: Its Impact On The Study Of The Late Prehispanic Period Of The Sierras De Córdoba, Argentina. Four archaeological sites dating from the Late Prehispanic Period of the Sierras de Córdoba (ca. 1000-300 BP) are studied by means of palynological analysis. These sites are Los Algarrobos 1, Puesto La Esquina 1 (Pampa de Olaen), C.Pun.39 and Las Chacras 2 (Punilla valley). Los Algarrobos 1 and Puesto La Esquina 1 sites present scarce pollinic grains as a consequence of taphonomic bias. However, from the pollinic spectrum from C.Pun.39 and Las Chacras 2 sites it was possible to infer vegetational paleo-communities dominated by Chenopodiaceae- Amaranthaceae and Brassicaceae (at C.Pun.39 site) and by the former taxon (at Las Chacras 2 site). There is poor presence of both vegetable families in current pollinic samples. It is thought that the high proportions of Chenopodiaceae- Amaranthaseae might suggest the presence of pre-Hispanic Chenopodium quinoa and Amaranthus caudatus crops in the areas surrounding the sites. Further evidence, such as domestic plant remains and agricultural tools, supports this hypothesis. Nevertheless, the high proportions of Chenopodiaceae-Amaranthaceae and Brassicaceae found in C.Pun.39 would also indicate successive abandonment and reoccupation processes at the residential camps, probably seasonally. Environments affected by anthropic activity might have contributed to the growth of these plants.
Keywords: Palynology; Archaeological Sites; Late Prehispanic Period.
INTRODUCCIÓN
Hasta hace unos pocos años las estrategias de movilidad y subsistencia implementadas por los grupos agroalfareros que habitaron las sierras y llanuras de Córdoba durante el periodo prehispánico tardío (ca. 1000-300 AP) permanecían desconocidas. La incorporación de proyectos regionales permitió lograr avances significativos relacionados particularmente con el uso del paisaje y sus recursos (Berberián y Roldán 2001; Laguens 1999; Medina 2008; Pastor 2007; Pastor y Medina 2005; Recalde 2007; Rivero 2001).
El análisis de la información arqueológica actualmente disponible indica que las poblaciones tardías alternaban las estrategias agrícolas y predadoras de acuerdo a las condiciones variables del entorno adaptativo y la disponibilidad estacional de recursos silvestres (Medina 2008; Pastor 2007). Durante la estación productiva (primavera/verano), distintas unidades domésticas se agrupaban en los sectores deprimidos para cultivar maíz (Zea mays), zapallo (Cucurbita sp.) y porotos (Phaseolus sp.), así como para recolectar frutos chaqueños como la algarroba (Prosopis sp.) y el chañar (Geoffroea decorticans). El grupo coresidente, una vez finalizadas las tareas agrícolas y de recolección, se dispersaba hacia las porciones superiores de las pampas de altura para capturar artiodáctilos de alto rendimiento como el guanaco (Lama guanicoe) y el venado de las pampas (Ozotoceros bezoarticus), combinando momentos de baja y alta agregación. El desarrollo de un nicho económico amplio, de este modo, se acompañaba de cierto grado de movilidad residencial, momentos de dispersión/agregación de los grupos co-residenciales y de procesos estacionales de abandono de las prácticas productivas para abastecerse de recursos de caza y recolección.
En este trabajo la propuesta es evaluada sobre la base de información palinológica obtenida en cuatro sitios arqueológicos del sector central de las sierras de Córdoba: C.Pun.39, Las Chacras 2 (valle de Punilla), Los Algarrobos 1 y Puesto La Esquina 1 (Pampa de Olaen) (Figura 1). Los resultados, aún cuando su alcance es considerado preliminar, fueron analizados en términos de su significancia cultural, en el marco de las prácticas económicas y del uso del espacio por parte de las sociedades tardías. Específicamente, los espectros polínicos son utilizados para maximizar el potencial de la información arqueobotánica y discutir la presencia prehispánica de cultígenos con problemas de conservación diferencial en el registro arqueológico. En forma secundaria, se intenta inferir el patrón de movilidad y subsistencia a partir del estudio de su impacto sobre la vegetación local.
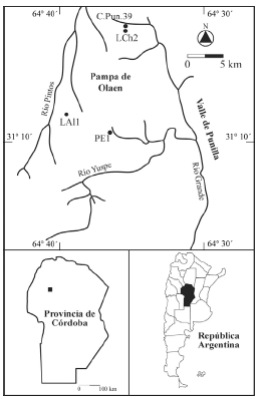
Figura 1. Ubicación geográfica de los sitios arqueológicos mencionados en este trabajo.
CARACTERÍSTICAS DEL ÁREA
Clima, vegetación
La provincia de Córdoba, ubicada dentro de la zona templada de Argentina, está dominada principalmente por dos centros de presión atmosférica: el Anticiclón Subtropical del Atlántico y el Ciclón Subtropical Continental del Noroeste. El primero de ellos confiere masas de aire cálido y húmedo a la porción sur y este de la provincia, mientras que el segundo brinda condiciones cálidas y secas al norte y oeste de la misma. En ambas áreas las precipitaciones se centran en el verano, aunque las características geomorfológicas heterogéneas (sierras, valles y bolsones) determinan variaciones en la cantidad de lluvia y en las temperaturas (Bridarolli y di Tada 1996).
Las temperaturas medias anuales varían entre 10º C y 18º C decreciendo de norte a sur y a medida que aumenta la altitud. La temperatura máxima media anual de 20º C circunda a la región montañosa, 14º C corresponde a las pampas de altura y 27º C alcanza en los salares y llanuras del noroeste de la provincia (Capitanelli 1979).
Las precipitaciones muestran la disminución de la humedad en sentido este-oeste, a excepción de las situaciones micro-climáticas que ocurren en la zona montañosa. Las mismas van desde más de 900 mm anuales en el este de la provincia y en los picos y pampas de altura, a menos de 400 mm en la región de los salares (al noroeste de la misma) (Capitanelli 1979)
Luti et al. (1979) clasifica a las comunidades vegetales de la provincia de Córdoba sobre la base de los dos ámbitos geográficos más importantes que las caracterizan: las llanuras y las sierras (Figura 2). Dentro de las planicies incluye al bosque chaqueño (oriental y occidental), los ambientes halófitos de las Salinas Grandes y Mar Chiquita, el espinal y la estepa pampeana. En las sierras, diferencia los pisos vegetacionales del bosque serrano, romerillal y pastizales de altura.
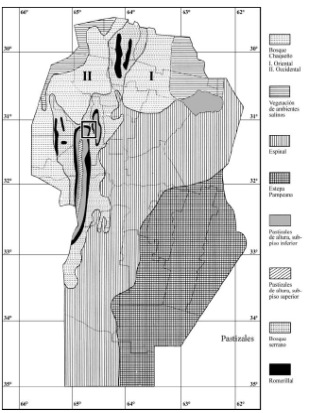
Figura 2. Vegetación de la provincia de Córdoba (modificado de Luti et al. 1979). El recuadro indica la localización del área de estudio.
El bosque chaqueño oriental cubre el bolsón ubicado entre las sierras del Norte y la laguna de Mar Chiquita, en el noreste de la provincia, siendo considerado la continuación meridional del bosque santiagueño. El estrato arbóreo está dominado por quebrachos (Aspidosperma quebracho-blanco; Aspidosperma quebracho-colorado), algarrobos (Prosopis nigra y Prosopis alba), talas (Celtis tala) y mistoles (Ziziphus mistol), mientras que en el sotobosque dominan las leguminosas Acacia sp., brea (Cercidium australis), así como la jarrilla (Larrea sp.). El estrato herbáceo se caracteriza por la presencia de gramíneas como Setaria argentina, Egragrostis lugens, Digitaria californica y Bromus uniliodes.
El bosque chaqueño occidental se desarrolla en el bolsón semi-árido que se extiende desde las Salinas Grandes hasta el límite noroeste del sector serrano. Su comunidad climax la componen: Bulnesia sp. (retama), Tabeuia nodosa (palo cruz), Larrea sp. (jarilla) y diversas mimosoideas del género Prosopis, mientas que el estrato herbáceo está dominado por las gramíneas Bouteloua sp. y Setaria sp., entre otras.
En las cercanías de las Salinas Grandes y la laguna de Mar Chiquita, el bosque chaqueño pierde abruptamente altura y densidad por la salinidad de los suelos. Predominan, en cambio, las plantas adaptadas a ambientes halófitos, fundamentalmente chenopodiáceas como el jume (Suaeda divaricata), el cachiyuyo (Atriplex argentina), verdolaga salada (Salicornia ambigua), jumecillo (Heterotachys olivaceus), entre otros.
La vegetación del Espinal, actualmente empobrecida por el desarrollo de prácticas agrícolas intensivas, ocupa las extensas planicies al este de las serranías. Está dominada por leguminosas mimosoideas, entre los que se encuentran Prosopis alba y Prosopis nigra y Acacia sp., acompañadas por Jodinia, Celtis y Aspidosperma (quebracho-blanco). A partir del paralelo 31º S, se funde con la estepa pampeana dando paulatinamente lugar a comunidades vegetales dominadas por gramíneas de los géneros Panicum, Stipa, Poa, Chloris y Baccharis.
El gradiente altitudinal del sector montañoso favoreció el establecimiento de una serie de pisos o cinturones de vegetación compuestos por especies de diversas familias y corrientes florísticas. El bosque serrano se desarrolla en los valles y piedemontes, con cotas que fluctúan entre los 500 y los 1100-1350 msnm, alcanzando mayor altitud en las laderas orientadas hacia el oeste. Los árboles están dominados por el molle de beber (Lithraea molleoides), el coco (Fagara coco) y el orco-quebracho (Schinopsis hankeana), alternándose con duraznos de las sierras (Kageneckia lanceolada) y manzanos del campo (Ruprechtia apetala), entre otros. El sotobosque se compone principalmente de Acacia cavens, mientras que el estrato herbáceo está representado por las bromeliáceas Dyckia floribunda y Deuterocohnia longipetala.
El piso de romerillar se extiende en forma continua entre los 1100 y los 1700 msnm, formando una amplia faja de transición entre el bosque serrano y el pastizal de altura. Dominan las compuestas arbustivas como el romerillo (Hetherothalamus alienus), romerito (Eupatorium buniifolium), Baccharis sp. y algunas cactáceas, que cubren las pendientes abruptas y de suelos rocosos.
Sobre terrenos relativamente planos localizados a partir de los 1100 msnm, se encuentra el piso denominado pastizal de altura, que Luti et al. (1979) subdivide en dos subpisos bien diferenciados. El subpiso inferior, que alcanza los 2000 msnm, está dominado por Stipa y Festuca. El subpiso superior posee, como respuesta al incremento orográfico de las precipitaciones, mayor riqueza de gramíneas, formando verdaderos prados de Alchemira pinnata, Deyeuxia hieronymii, Stipa sp., Bromus sp., Poa sp., etc. Los árboles están representados por el tabaquillo (Polylepis australis) y orco-molle (Maytenus boaria), que sólo crecen en las quebradas protegidas de los fuertes vientos que azotan la región.
UBICACIÓN GEOGRÁFICA Y CRONOLOGÍA DE LOS SITIOS
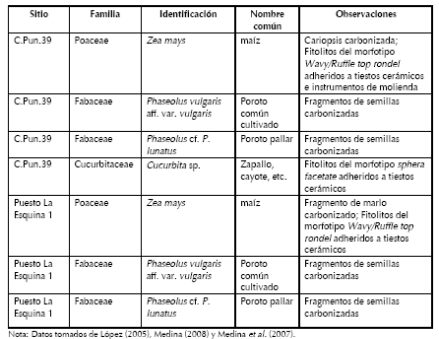
C.Pun.39 (31º 3´376 S, 64º 31´685 O, 1050 msnm, Departamento de Punilla) se localiza sobre ambas márgenes del arroyo Las Chacras, en la porción septentrional del valle de Punilla (Figura 1). La vegetación circundante es de bosque serrano, aunque empobrecido a causa de la altitud absoluta, con árboles aislados de Lithraea molleoides, Fagara coco, mimosoideas, Hetherothalamus alienus y pajonales de Stipa sp. En términos generales, comprende extensiones de 40 ha en donde la actividad agrícola moderna puso al descubierto una alta densidad y diversidad artefactual. Las tareas de campo implicaron la excavación de 16 m2, alcanzando profundidades máximas de 90 cm. La riqueza de los conjuntos recuperados, que incluyen macro y microrrestos de plantas cultivadas (Tabla 1), así como instrumentos vinculados al desmonte y roturado del terreno (azadas líticas), indica que en C.Pun.39 se realizaron actividades diversas propias de los espacios domésticos, incluyendo la producción de alimentos. Sobre muestras de carbón aproximadamente encolumnadas se obtuvieron tres dataciones radiocarbónicas cuyos resultados fueron 525 ± 36 AP (AA64819; Phaseolus sp.; δ13C= -24,6‰), 716 ± 39 AP (AA62339; carbón; δ13C= -24,17‰) y 854 ± 39 AP (AA62338; carbón; δ13C= -24,32‰), posicionando al sitio en diferentes momentos del periodo prehispánico.
Tabla 1. Taxones de plantas cultivadas recuperadas en C.Pun.39 y Puesto La Esquina 1.
Las Chacras 2 (LCh2, 31º 3´792 S, 64º 31´680 O, 1060 msnm) se localiza a 1000 m al sur de C.Pun.39, sobre la cabecera de una quebrada lateral (Figura 2). Comprende un reparo rocoso de 21 m2 cubiertos, en donde se excavaron 6 m2 y se alcanzaron profundidades máximas de 70 cm. Toda la secuencia del depósito está dominada por materiales cerámicos y líticos cuyas características tecnológicas respondenal periodo prehispánico tardío (ca. 1000-300 AP). Sin embargo, las dataciones efectuadas sobre restos óseos con huellas de corte -i.e. 154 ± 43 AP (AA64821;δ13C= -12,4‰), 466 ± 45 AP (AA68142; δ13C= -19,1‰), 560 ± 45 AP (AA68143; δ13C= -20,7‰) y 3819 ± 55 AP (AA64822; δ13C= -16,5‰)- y partículas de carbón dispersas -i.e. 126 ± 38 AP (AA68144; δ13C= -24,4‰)-, la ausencia de una estratigrafía arqueológica claramente definida, la identificación de especímenes de fauna europea, así como cuevas y abundantes excrementos de vizcacha (Lagostomus maximus), indican que el sitio tiene una alta dinámica postdepositacional y una baja resolución cronológica. Los sedimentos del sitio, de este modo, promedian la lluvia polínica de los últimos ca. 4000 años, situación que trajo aparejada dificultades al momento de incorporar los resultados a la discusión final.
Los Algarrobos 1 (LAl1, 31º 6´563 S, 64º 39´487 O, 1100 msnm, Departamento de Punilla) es un asentamiento a cielo abierto localizado en los límites occidentales de la Pampa de Olaen, sobre la margen derecha del arroyo homónimo (Figura 1). Aún cuando el sitio se emplaza en el pastizal de altura, es posible encontrar elementos arbóreos del bosque serrano, fundamentalmente Celtis tala y mimosoideas. Durante las tareas de campo sólo se excavaron 4 m2, alcanzándose profundidades máximas de 40 cm. Las actividades identificadas a partir del análisis de los conjuntos arqueológicos superficiales y estratigráficos confirmaron su utilización como sitio agrícola y residencial. Sobre muestras de carbón se obtuvo una datación de 949 ± 40 AP (AA64818; δ13C= -24,3‰).
Puesto La Esquina 1 (PE1, 31º 9´306 S, 64º 37´498 O, 1140 msnm; Departamento de Punilla) se localiza en los terrenos potencialmente agrícolas del fondo de una quebrada bien protegida de la Pampa de Olaen (Figura 1). La cobertura vegetal está dominada por Stipa sp., mientras que las laderas rocosas presentan a b u n d a n t e s H e t h e r o t h a l a m u s alienus. Las excavaciones implicaron la apertura de 15 m2 y profundidades máximas de 60 cm. El conjunto cerámico, óseo, lítico, arqueofaunístico y paleobotánico (Tabla 1), así como la presencia de una posible estructura de vivienda, indicaron la realización de actividades múltiples propias de los espacios residenciales, como la producción, almacenamiento, procesamiento, cocción y consumo de alimentos, manufactura, mantenimiento y descarte de artefactos, etc. Sobre muestras de carbón encolumnadas se obtuvieron dos dataciones radiocarbónicas cuyos resultados fueron 365 ± 38 AP (AA64816; δ13C= - 23,8‰) y 362 ± 43 AP (AA64815; δ13C= -10,2‰), posicionando al sitio en momentos previos a la conquista española e indicando la rápida formación del depósito.
MATERIALES Y MÉTODOS
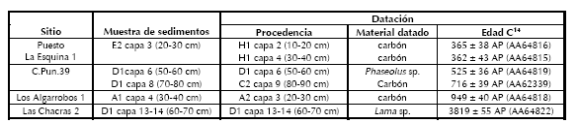
Las muestras de sedimentos fósiles fueron recolectadas respetando los niveles artificiales utilizados en la excavación y siguiendo el protocolo comúnmente utilizado para estudios palinológicos en arqueología (Fish 1994; Pearsall 2004; Piperno 2006). Ante la imposibilidad de analizar la totalidad de las mismas, el estudio se centró preliminarmente en aquellas que presentaban mayor grado de asociación con dataciones radiocarbónicas (Tabla 2). Para obtener análogos de la vegetación moderna y controlar posibles contaminaciones, se tomaron muestras de sedimentos superficiales en los sitios C.Pun.39 y LCh2 (Piperno 1995).1
Tabla 2. Dataciones radiocarbónicas de los sedimentos seleccionados para el estudio polínico.
La concentración polínica de los palinomorfos se analizó de acuerdo con la metodología propuesta por Heusser y Stock (1984). Se pesaron 25 g de cada muestra y antes de realizarse el tratamiento físicoquímico se agregaron 3 tabletas conteniendo 12.542 esporas de polen foráneo (Lycopodium sp.). Se contaron como mínimo 200 granos de polen, utilizándose un criterio similar al de área mínima propuesto por Bianchi y D´Antoni (1986). Las frecuencias relativas y absolutas (y las sumas totales de granos de polen en las muestras actuales) fueron graficadas en diagramas polínicos (Figuras 3 y 4). Las muestras LA11, C.Pun.39 capa 8 y PE1, al registrar sumas insuficientes de granos de polen, no fueron consideradas al momento de efectuar dichas estimaciones. Otros "microfósiles no polínicos", tales como zygnemataceas, esporas de hongos, de bryophytas, etc., se contaron fuera de las sumas polínicas (Tablas 3 y 4). Tanto los recuentos como las determinaciones se realizaron bajo luz transmitida normal en un microscopio Olympus BH2 Nº 100786. Para la asignación taxonómica se utilizó material de referencia de la Palinoteca del Laboratorio de Palinología (Universidad Nacional del Sur) y bibliografía especializada (Heusser 1971; Hooghiemstra 1984; Markgraf y D´Antoni 1978; Reille 1992; van Geel 1982- 1983; entre otros).

Figura 3. Diagrama polínico de las muestras fósiles.

Figura 4. Diagrama polínico de las muestras superficiales (sitios C.Pun.39 y Las Chacras 2).
Tabla 3. Microfósiles no polínicos registrados en los sitios arqueológicos. Los números expresan ejemplares hallados en cada muestra. (*) sensu van Geel (1982-1983) (**) (Hooghiemstra 1984).
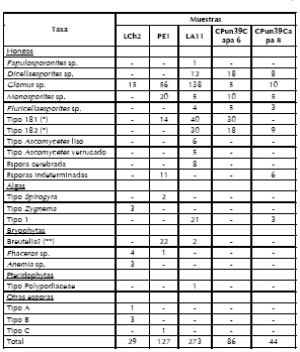
Tabla 4. Componentes no polínicos registrados en las muestras actuales de los sitios CPun.39 y LCh2 *sensu van Geel et al. (1982-1983) (**) Hooghiemstra (1984).
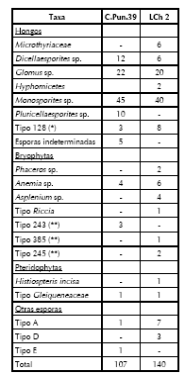
RESULTADOS
Muestras fósiles
El espectro polínico de Las Chacras 2 (edad radiocarbónica máxima: 3819 ± 55 AP) está dominado por Chenopodiaceae-Amaranthaceae (60%), Asteraceae: Tubuliflorae: Bidens sp. y Ambrosia sp., entre otras (16%), Liguliflorae: (1%), Poaceae (5,3%) y Brassicaceae (1%). Los elementos del monte arbustivo están representados por: Solanaceae (2,1%), Zygophyllaceae (Larrea divaricata: 2,1%), Rubiaceae (1,6%), Labiate (1,6%), Anacardiaceae (1%) y Ulmaceae (Celtis sp., entre otras) (1%). Con menos del 1% están presentes: Convolvulaceae, Malvaceae, Ephedraceae y Portulacaceae. El polen de plantas acuáticas está asociado a: Cyperaceae (1%), Potamogetonaceae (2,1%) y Myriophyllum sp. (<1%). Entre las esporas de hongos se documentaron: Dicellaesporites sp., Glomus sp., Monosporites lagenarius, Papulosporonites sp. y Pluricellaesporites sp., de zygnemataceas (Zygnema sp.), de bryophytas (Phaceros sp. y Anemia sp.) y esporas tipo A y B completan la asociación. La concentración polínica absoluta es: 784 granos/g de sedimento.
En Los Algarrobos 1 (edad: 949 ± 40 AP) se identificaron las siguientes familias vegetales: Apiaceae, Asteraceae, Brassicaceae, Poaceae, Chenopodiaceae- Amaranthaceae, Ephedraceae, Euphorbiaceae, Myricaceae, Oleaceae, Solanaceae, Zygophyllaceae, Fabaceae (Acacia sp.), asociados a granos de polen indeterminados así como a polen extra-regional de Juglandaceae, Myrtaceae y Podocarpaceae. Las esporas de hongos están presentes con: Corticiaceae, Dicellaesporites sp., Glomus sp., Monoporisporites sp., Pluricellaesporites sp., tipo 181 y tipo 182 (van Geel 1982-1983), tipo Ascomycetes liso y verrucado, esporas cerebradas y las esporas de algas (tipo 1), esporas de bryophytas (Breutelia? Hooghiemstra 1984) y tipo Polypodiaceae. La concentración polínica absoluta es de: 164 granos/g de sedimento.
C.Pun.39 capa 8 (edad: 716 ± 39 AP) presenta Brassicaceae, Caryophyllaceae, Chenopodiaceae- Amaranthaceae, Convolvulaceae, Malvaceae, Oleaceae, tipo Rosaceae, Fabaceae (Acacia sp.) y polen de plantas acuáticas Myriophyllum sp. y Thyphaceae/Sparganiaceae. Entre las esporas de hongos: Dicellaesporites sp., Glomus sp., Monosporites sp., Pluricellaesporites sp., tipo 182 (van Geel 1982-1983) y esporas indeterminadas y esporas tipo 1 de algas completan la asociación. La concentración polínica absoluta es: 98 granos/g de sedimento.
En la muestra de sedimentos de C.Pun.39 capa 6 (edad: 525 ± 36 AP) dos tipos polínicos dominan la asociación: Chenopodiaceae- Amaranthaceae (38%) y Brassicaceae (37%), secundariamente asociadas a Asteraceae (tubuliflorae) (7%), Poaceae (3%) y Malvaceae (1%). Dentro de los elementos del monte arbustivo, el mayor porcentaje le corresponde a Oxalidaceae (5,21%). Con porcentajes poco significativos e inferiores al 1%, están presentes las familias Caryophyllaceae, Scrophulariaceae, Solanaceae y Acacia sp. El polen extra-regional está representado por Podocarpaceae (1,42%), Myrtaceae (1%) y Nothofagaceae (<1%). El polen de plantas acuáticas Cyperaceae (1.42%) y Thyphaceae/Sparganiaceae (1%) y algunos granos indeterminados, completan la asociación. Las esporas de hongos están representadas por: Dicellaesporites sp., Glomus sp., Monoporisporites sp., Pluricellaesporites sp., esporas tipo 181 y 182 (van Geel 1982-1983). La concentración polínica absoluta es de: 445 granos/g de sedimento.
En Puesto La Esquina 1 (edad: 362 ± 43 AP, 365 ± 38 AP) se registra la presencia de Asteraceae, Poaceae, Chenopodiaceae-Amaranthaceae, Euphorbiaceae, Geraniaceae, Solanaceae, Zygophyllaceae y Fabaceae (Acacia sp.). Algunos granos de polen indeterminados y el polen extra-regional de Araucariaceae, Myrtaceae y Podocarpaceae, completan la asociación. Las esporas de hongos están presentes con: Glomus sp., Monoporisporites sp., tipo 181 (van Geel 1982-1983) y esporas indeterminadas. Algunos de ellos se presentan totalmente oxidados. Las esporas de zygnemataceas: Spirogyra sp., bryophytas: (Breutelia?) (Hooghiemstra 1984) y Phaceros sp. y esporas tipo C son registradas en esta asociación. La concentración polínica absoluta es de 180 granos/g de sedimento.
Muestras actuales
El espectro polínico superficial de C.Pun.39 esta dominado por tipos polínicos pertenecientes al sustrato herbáceo: Cyperaceae (27%), Poaceae (22.5%) y Asteraceae (21.5%). Siguen en importancia las Chenopodiaceae-Amaranthaceae con 17%, siendo poco significativo el porcentaje de Brassicaceae (<1%). Dentro del polen arbóreo, con porcentajes que no superan el 2%, se hallan entre otros: Schinus molle, Acacia sp., Celtis sp., Fagara coco. Entre las esporas de hongos, se identificaron Glomus sp., Monosporites sp., Dicellaesporites sp. y Multicellaesporites sp. Las esporas de bryophytas están representadas por: Phaceros sp., Anemia sp., tipo Riccia, tipo A y B. Las esporas de pteridophytas, escasamente registradas, están asociadas a Gleicheniaceae, tipo 128 y 243 (Hooghiemstra 1984).
La muestra actual de Las Chacras 2 es similar en los componentes del sustrato herbáceo a la de C.Pun.39: Poaceae (22,5%) y Asteraceae (20,5%). Siguen en importancia Chenopodiaceae-Amaranthaceae (12,5%), mientras que Brassicaceae presenta porcentajes muy bajos (1,5%). Pinnaceae (16,5%) domina el polen arbóreo, siendo el resto de los componentes inferiores al 2%. El polen de plantas acuáticas está representado por Cyperaceae (11,5%). Entre las esporas de bryophytas se identificaron: Anemia sp., tipo A y C, esporas tipo 245 (Hooghiemstra 1984) y algunas indeterminadas completan la asociación.
DISCUSIÓN
Los registros polínicos de diferentes regiones de Argentina señalan que la composición florística no varió significativamente durante el lapso Último Máximo Glacial/Holoceno (Barreda et al. 2007). Sin embargo, algunos conjuntos polínicos han sufrido variaciones en sus proporciones y composición indicando cambios latitudinales y longitudinales significativos a lo largo de los gradientes norte-sur y este-oeste (Barreda et al. 2007). El establecimiento de los patrones oceanográficos y atmosféricos que rigen las condiciones climáticas actuales a partir del Holoceno tardío (ca. 3000 AP) acentuó las similitudes entre las biotas actuales y las fósiles posteriores a esa cronología (Heusser 1989a, 1989b, 1990; Markgraf 1991).2
Estos argumentos denotan la importancia de los datos aportados por la "lluvia polínica actual" en el área donde se efectúan análisis palinológicos fósiles. En lo que hace específicamente al presente estudio, la comparación entre los espectros polínicos fósiles y actuales sugiere que los últimos no constituyen análogos de los primeros. La exigua representación en las muestras modernas de aquellas familias vegetales altamente conspicuas en los depósitos arqueológicos, como Chenopodiaceae-Amaranthaceae y Brassicaceae, justifican esta postura.
Estudios botánicos realizados en las últimas décadas señalan que las Chenopodiaceae Amaranthaceae no constituyen un componente importante del bosque serrano ni del pastizal de altura (Cabido et al. 1998; Kraus et al. 1999; Luti et al. 1979; Nuñez y Cantero 2000; Sayago 1969). En cambio, son dominantes en los ambientes halófitos que rodean a las Salinas Grandes (NO de la provincia) y la laguna Mar Chiquita (NE) (Luti et al. 1979), a más de 100 km de los sitios estudiados. A nivel palinológico, espectros polínicos con altos porcentajes de esta familia se asocian a ambientes naturales de estepa halófita (Grill y Guerstein 1995; Vilanova et al. 2006), situaciones climáticas subhúmedas/secas (Prieto 1996, 2000; entre otros), paisajes antrópicamente modificados (Bray 1995; Piperno y Pearsall 1998) y a la presencia arqueológica de cultivos andinos (Lupo et al. 2006).
Una situación similar ocurre con Brassicaceae, que no constituye un elemento importante dentro de las comunidades vegetales serranas actuales. Sí tiene mayor frecuencia en las llanuras orientales, en especial Lepidium sp., donde se desarrolla como maleza de los cultivos comerciales (Kraus et al. 1999; Nuñez y Cantero 2000). En relación a estudios palinológicos, el polen de esta familia constituye porciones significativas en espectros polínicos superficiales y subsuperficiales de la región pampeana con fuerte asociación a disturbios antrópicos o eólicos (Grill 1995; Grill et al. 2007; Prieto 1996; entre otros).
Algunos autores llegaron a caracterizar a Chenopodiaceae-Amaranthaceae como plantas "seguidoras de campamentos", ya que es frecuente observar parches de estos arbustos en los terrenos abiertos con suelos continuamente perturbados que rodean viviendas rurales y/o semi-urbanas (Bruno 2006; Kuznar 1993; Smith 1992). Esta denominación podría extenderse a Brassicaceae, cuyo polen suele estar asociado, en depósitos históricos y actuales, a una intensa actividad humana, junto a árboles y malezas tales como Polygonaceae, Pinus sp., Eucalyptus sp., Carduus sp., etc. (Grill 1997, 2003; Borromei 1995, 1998; Trivi de Mandri et al. 2006; entre otros).
Las frecuencias excepcionales de Chenopodiaceae-Amaranthaceae, asociadas a una datación de 525 + 36 AP en C.Pun.39 y a una baja resolución cronológica en LCh2, sugiere que plantas con polen similar a los cultígenos quínoa (Chenopodium quinoa) y amaranto (Amaranthus caudatus) crecían abundantemente en los alrededores de los sitios (Fish 1994; Lupo et al. 2006; McLauchlan 2003). Una serie de evidencias y argumentos permiten sostener la hipótesis de que probablemente provienen de chacras de cultivo y no de especies silvestres. Las mismas son:
- la mayoría de las plantas cultivadas no dispersan el polen a grandes distancias, probablemente como producto de los cambios genéticos que acompañaron a la domesticación, por lo que su presencia en sedimentos arqueológicos aporta información directa y confiable acerca de su cultivo dentro de un radio local limitado (Fish 1994: 62);
- distintos modelos de dispersión de polen verifican la alta relación entre un determinado espectro polínico con la vegetación circundante (Prentice 1985; Moseholm et al. 1987), siendo la participación de otras fuentes de emisión regionales y/o extraregionales poco significativa;
- las evidencias geomorfológicas, estratigráficas y pedológicas indican condiciones más cálidas y húmedas que las actuales para el segmento 1000-500 AP (Carignano 1999), desacreditando la posibilidad de que los altos porcentajes de Chenopodiaceae-Amaranthaceae y Brassicaceae sugieran mayor aridez;
- el registro superficial de azadas líticas en C.Pun.39, así como en PE1 y LAl1, permite sostener que los sitios residenciales también fueron importantes locus de producción agrícola;
- en C.Pun.39 capa 6 el polen de Chenopodiaceae-Amaranthaceae se encuentra en el mismo nivel estratigráfico que los macrorrestos carbonizados de un cotiledón de poroto común cultivado (Phaseolus vulgaris aff. var. vulgaris), datado por AMS en tiempos prehispánicos y un cariopsis de maíz;
- los restantes cultígenos arqueológicos documentados en C.Pun.39 (Medina et al. 2007) comúnmente se dan junto a Chenopodium sp., formando la típica "tétrada americana": maíz, zapallo, poroto y quínoa (Lagiglia 2001; Oliszewski 2004; Smith 1992);
- las chenopodiáceas y amarantáceas se desarrollaron como cultivos de importancia económica ca. 3500 AP en varias regiones del área andina, donde continúan siendo un componente significativo de la dieta (Bruno 2006);
- los granos de quínoa y amaranto son frecuentemente molidos en morteros y conanas (Oliszewski 2004), artefactos recuperados en superficie y en estratigrafía;
- tienen un alto valor nutritivo, sólo comparable con el maíz (Smith 1992);
- son plantas resistentes a las bajas temperaturas, heladas y sequías, lo que las convierte en cultígenos óptimos para enfrentar las frecuentes adversidades ambientales del sector serrano de Córdoba (Browman 1987; Bruno 2006);
- la quínoa, junto con el maíz, es frecuentemente mencionada en los documentos del siglo XVII. Por ejemplo, en un voluminoso juicio por tierras entre dos encomenderos de la zona centro-norte del valle de Punilla que data de 1639, los testigos declaran "antiguamente sembraban quinua los indios de Cosquin" (Archivo Histórico de la Provincia de Córdoba [AHPC], Escribanía I, Legajo 72, Expediente 2, f. 75v); "sembraban quinua y sapallos y mays" (f. 82v); "unos morteros que ycieron los dichos indios para moler quinua y mays" (f. 80v);
- censos florísticos sugieren que Chenopodium sp. y Amaranthus sp. no abundan actualmente en ninguno de los pisos florísticos de las sierras de Córdoba;
- no son componentes importantes en la "lluvia polínica actual" del área de estudio.
Los valores de Chenopodiaceae-Amaranthaceae y Brassicaceae, por otro lado, podrían estar indicando que C.Pun.39 fue abandonado y reocupado en reiteradas ocasiones, probablemente en forma estacional, creando un hábitat abierto continuamente perturbado por la actividad antrópica que favoreció al crecimiento de plantas colonizadoras de este tipo de ambientes (Bruno 2006; Fish 1994; Kuznar 1995; Pearsall 2004; Piperno 1995, 2006; Pohl et al. 1996; Smith 1992; Trivi de Mandri et al. 2006).3 Esto es coherente con un patrón de alta movilidad residencial entre los grupos prehispánicos tardíos, con abandono de los sitios residenciales y de las prácticas productivas en ciertos momentos del año a fin de aprovechar al máximo frutos chaqueños y productos de caza disponibles en distintos puntos del espacio serrano (Pastor 2007). La baja inversión en infraestructura agrícola, la ausencia de estructuras habitacionales imperecederas en superficie, la posible presencia de casas-pozo, los resultados de las dataciones y el restante paisaje arqueológico de las sierras de Córdoba, argumentan a favor de este aserto.
La ausencia de evidencias arqueológicas que soporte el cultivo prehispánico de tubérculos andinos como la papa (Solanum sp.) y la oca (Oxalis tuberosa), o aquellos propios de las tierras bajas como la batata (Ipomea batata), aún cuando son citados por la documentación etnohistórica, puede obedecer a ciertos problemas de preservación diferencial (Oliszewski 2004; Pearsall 1995, 2004; Perry 2004; Piperno 2006; Piperno y Pearsall 1998; Smith 2006). No se descarta que estén incluidos a una baja resolución taxonómica en los tipos polínicos Solanaceae, Oxalidaceae y Convolvulaceae, respectivamente. Al igual que para Chenopodiaceae-Amaranthaceae, el análisis de los microfósiles de almidón adheridos a tiestos cerámicos e instrumentos de molienda, cuyo estudio se encuentra en proceso, probablemente permita verificar o descartar la presencia de estos taxones (Oliszewski 2004; Perry 2004; Smith 2006).
Finalmente cabe acotar que la exigua cantidad de granos de polen registrada en los sitios C.Pun.39 (capa 8), PE1 y LAl1, las cuales impidieron la reconstrucción de las paleocomunidades vegetales, puede asociarse a factores tafonómicos. Evidencias de condiciones oxidantes (esporas de hongos y otros restos vegetales totalmente ennegrecidos) fueron registradas en C.Pun.39 (capa 8). En los sitios PE1 y LAl1, la importante proporción de esporas de hongos (ver Tabla 3) denota la mayor resistencia de las mismas que los granos de polen a factores tales como condiciones alternantes de humedad y sequía, alcalinidad del medio, etc. En consecuencia, la oxidación química (por exposición aérea de los sedimentos) habría sido la responsable de la destrucción de los palinomorfos en la capa 8 del sitio C.Pun.39 y la oxidación biológica (por hongos y bacterias) la responsable de la destrucción de los mismos en los sitios PE1 y LAl1.
CONCLUSIONES
El estudio palinológico realizado en C.Pun.39, LCh2, PE1 y LAl1 permitió reconstruir la vegetación desarrollada en dos de ellos: C.Pun.39 y LCh2, aunque en este último con una resolución que limitó el alcance de los resultados obtenidos. La falta de correspondencia entre la lluvia polínica actual y los espectros fósiles, habiéndose descartado la participación de fuentes emisoras no-locales y/o condiciones de mayor aridez, fue atribuida al impacto producido por la colonización del paisaje por grupos humanos que combinaban recursos agrícolas y silvestres junto con patrones de movilidad estacionales.
Las sociedades tardías, de esta manera, no sólo se adaptaron al ambiente serrano mediante prácticas económicas y de uso del espacio específicas, sino que también interactuaron con él modificándolo. Eso implicaría asumir que los grupos agroalfareros, aún con una tecnología sencilla y sin formar centros urbanos, contribuyeron a alterar las comunidades vegetales de fines del Holoceno. En consecuencia, los estudios que intenten comprender la dinámica evolutiva del ecosistema serrano no pueden dejar de lado a las poblaciones prehispánicas. Los depósitos arqueológicos bien datados, en este sentido, aportan datos significativos para el conocimiento del complejo escenario evolutivo y sus cambios a través del tiempo.
A lo largo de este trabajo, sin embargo, sólo se realiza una exploración preliminar, en gran parte a causa de que los estudios palinológicos aún son fragmentarios y se limitan a la porción final del Holoceno. Esto obliga a incrementar la base de datos mediante nuevas excavaciones e incorporar sitios aún no analizados desde esta perspectiva. Asimismo, es necesario introducir estudios polínicos off-site (Pearsall 1995) sobre columnas de sedimentos obtenidas en lagunas y pantanos, que permitan completar la información derivada de los sitios arqueológicos. De este modo se reforzarán o modificarán las implicancias aquí expuestas.
Agradecimientos
Queremos expresar nuestro agradecimiento al Dr. Eduardo Berberián, quién se encargó de dirigir las diferentes etapas de la realización de este trabajo. A Timothy Jull del NSF Arizona Facility (Grant EAR01- 15488), por su colaboración en la realización de los fechados. A la Dra. Mirta Quattrocchio por la lectura crítica del manuscrito. A Juliana Cornago por la colaboración en la traducción del abstract. En los trabajos de campo participaron Diego Rivero, Sebastián Pastor, Natalia Bidart, Bruno Catania, Catriel Greco, Sergio Clavero, Ezequiel Gilandenghi, Pablo Giorno, Jonathan Gómez, Paula Granda, Ana Liboreiro, Yesica Paredes, Danilo Setten y Mariana Silva.
1 Las muestras se tomaron de una cuadrícula de 1 m2 y entre 0-2 cm de profundidad (Piperno 2006).
2 No obstante, en los últimos 1000 años se registraron dos eventos climáticos importantes en el sector central de Argentina: la "Anomalía Climática Medieval" y la "Pequeña Edad de Hielo". El primero, datado entre el 1000 AP y la conquista, presentó características subhúmedas/templadas, con inviernos más moderados que los actuales, mayores precipitaciones y gran excedente hídrico (Carignano 1999). El segundo, contemporáneo con la instalación del sistema colonial español, se caracterizó por la alternancia de sequías extremas con lluvias torrenciales, provocando la reducción de lagunas y la reactivación de procesos erosivos (Cioccale 1999; Piovano et al. 2004).
3 El conjunto fósil de LCh2 puede estar reforzando estos argumentos. Sin embargo, sus problemas cronoestratigráficos requieren ser explorados en profundidad para lograr un mayor soporte empírico.
REFERENCIAS CITADAS
Barreda, V., M. Anzótegui, A. Prieto, P. Aceñolaza., M. Bianchi, A. Borromei, M. Brea, M. Caccavari., S. Garrala., S. Grill, G. Guerstein, A. Lutz, M. Mancini, G. Ottone, M. Quattrocchio, E. Romero, M. Zamaloa y A. Zucol 2007 Diversificación y cambios de las angiospermas durante el Neógeno en Argentina. En Ameghiniana 50- Aniversario Publicación Especial 11, editado por S. Archangelsky, T. Sánchez y E. P. Tonni, pp. 173-191. Asociación Paleontológica Argentina, Buenos Aires. [ Links ]
Berberián, E. y F. Roldán 2001 Arqueología de las Sierras Centrales. En Historia Argentina Prehispánica, editado por E. Berberián y A. Nielsen, pp. 635-691. Editorial Brujas, Córdoba. [ Links ]
Bianchi, M. y H. D'Antoni 1986 Depositación del polen actual en los alrededores de Sierra de los Padres (Prov. de Buenos Aires). Actas del VI Congreso Argentino de Paleontología y Bioestratigrafía, pp. 16-27. Mendoza. [ Links ]
Borromei, A. 1995 Palinología, estratigrafía y paleoambientes del Pleistoceno Tardío-Holoceno en el valle del río Sauce Grande, provincia de Buenos Aires, Argentina. Polen 7: 19-31. [ Links ]
Borromei, A. 1998 Vegetación y clima del Cuaternario tardío en el valle superior del Río Sauce Grande, provincia de Buenos Aires, Argentina. Polen 9: 5-15. [ Links ]
Bray, W. 1995 Searching for environmental stress: climatic and anthropogenic influence on the landscape of Colombia. En Archaeology in the Lowland American Tropics. Current analytical methods and recent applications, editado por P. Stahl, pp. 96-112. Cambridge University Press, Cambridge. [ Links ]
Bridarolli, M. y I. di Tada 1996 Algunos aspectos de la geografía física de la provincia de Córdoba. En Biodiversidad de la Provincia de Córdoba, editado por I. di Tada y E. Bucher, pp. 15-38. Universidad Nacional de Río Cuarto, Río Cuarto. [ Links ]
Browman, D. 1987 Agro-pastoral risk management in the Central Andes. Research in Economic Anthropology 8: 171-200. [ Links ]
Bruno, M. 2006 A morphological approach to documenting the domestication of Chenopodium in the Andes. En Documenting domestication. New genetic and archaeological paradigm, editado por M. Zeder, D. Bradley, E. Emshwiller y B. Smith, pp. 32-45. University of California Press, Berkeley. [ Links ]
Cabido, M., G. Funes, E. Pucheta, F. Vendramini y S. Díaz 1998 A chorological analysis of the mountains from Central Argentina. Is all what we call Sierra Chaco really Chaco? Contribution to the study of the flora and vegetation of the Chaco. Candollea 53: 321-331. [ Links ]
Capitanelli, R. 1979 Clima. En Geografía Física de la Provincia de Córdoba, editado por J. Vázquez, R. Miatello y M. Roque, pp. 45-138. Editorial Boldt, Buenos Aires. [ Links ]
Carignano, C. 1999 Late Pleistocene to recent climate change in Córdoba Province, Argentina: geomorphological evidence. Quaternary International 57/58: 117-134. [ Links ]
Cioccale, M. 1999 Climatic fluctuations in the Central Region of Argentina in the last 1000 years. Quaternary International 62: 35-47. [ Links ]
Fish, S. 1994 Archaeological palynology of garden and fields. En The Archaeology of garden and field, editado por N. Miller y K. Gleason, pp. 44-69. University of Pennsylvania Press, Philadelphia. [ Links ]
Grill, S. 1995 Análisis palinológico de un perfil cuaternario en la cuenca del arroyo Napostá Grande, localidad: García del Río, Provincia de Buenos Aires. Actas de las VI Jornadas Geológicas Bonaerenses 1: 99-107. Junín. [ Links ]
Grill, S.1997 Palinología de un perfil cuaternario en el valle del arroyo Napostá Grande, provincia de Buenos Aires, Argentina. Polen 8: 23-40. [ Links ]
Grill, S. 2003 Análisis palinológico de sedimentos cuaternarios en la cuenca inferior del río Quequén Salado, provincia de Buenos Aires, Argentina. Polen 12: 37-52. [ Links ]
Grill, S. y G. Guerstein 1995 Estudio palinológico de sedimentos superficiales en el estuario de Bahía Blanca, Buenos Aires (Argentina). Polen 7: 40-49. [ Links ]
Grill, S., A. Borromei, G. Martínez, M. Gutiérrez, M. Cornou y D. Olivera 2007 Palynofacial analysis in alkaline soils and paleoenvironmental implications: the Paso Otero 5 archaeological site (Necochea District, Buenos Aires province, Argentina). Journal of South American Earth Sciences 24: 34-47. [ Links ]
Heusser, C. 1971 Pollen and Spores of Chile. The University of Arizona Press, Tucson. [ Links ]
Heusser, C. 1989a Late Quaternary vegetational and climatic of southern Tierra del Fuego. Quaternary Research 31: 396-406. [ Links ]
Heusser, C. 1989b Polar Perspective of Late Quaternary - Quaternary Climatics in the Southern Hemisphere. Quaternary Research 32: 60-71. [ Links ]
Heusser, C. 1990 Late glacial and Holocene vegetation and climate subantartic South America. Review of Paleobotany and Palynology 65: 9-15. [ Links ]
Heusser, L. y C. Stock 1984 Preparation techniques for concentrating pollen from marine sediments and other sediments with low pollen density. Palynology 8: 225-227. [ Links ]
Hooghiemstra, H. 1984 Vegetational and climatic history of the High Plain of Bogotá, Colombia: a continuous record of the last 3,5 million years. J. Cramer, Alemania. [ Links ]
Kraus, T., C. Bianco y C. Nuñez (editores) 1999 Los ambientes naturales del Sur de la Provincia de Córdoba. Fundación de la Universidad Nacional de Río Cuarto, Río Cuarto. [ Links ]
Kuznar, L. 1993 Mutualism between Chenopodium, herd animals and herders in the South Central Andes. Mountain Research and Development 13 (3): 257-265. [ Links ]
Kuznar, L.1995 Awatimarka. The Ethnoarchaeology of an Andean Herding Community. Harcourt Brace College Publisher, Forth Worth. [ Links ]
Lagiglia, H. 2001 Los orígenes de la agricultura en la Argentina. En Historia Argentina Prehispánica, editado por E. Berberián y A. Nielsen, pp. 41-81. Editorial Brujas, Córdoba. [ Links ]
Laguens, A. 1999 Arqueología del contacto hispano-indígena. Un estudio de cambios y continuidades en las Sierras Centrales de Argentina. BAR International Series 801. Archaeopress, Oxford. [ Links ]
López, L. 2005 Los pobladores productores de alimentos de las sierras de Córdoba. Primeras evidencias arqueobotánicas de los sitios Tala Cañada 1 y C.Pun.39. La Zaranda de Ideas 1: 89-91. [ Links ]
Lupo, L., J. Kulemeyer, C. Aschero y A. Nielsen 2006 Evidencias palinológicas de intervención humana en el paisaje desde el precerámico al formativo de Puna y Quebrada de Humahuaca. Resúmenes del XIII Simposio Argentino de Paleobotánica y Palinología, p. 85. Bahía Blanca. [ Links ]
Luti, R., M. Bertrán de Solís, F. Galera, N. Muller de Ferreira, M. Berzal, M. Nores, M. Herrera y J. Barrera 1979 Vegetación. En Geografía Física de la Provincia de Córdoba, editado por J. Vazquez, R. Miatelo y M. Roque, pp. 297-368. Editorial Boldt, Buenos Aires. [ Links ]
Markgraf, V. 1991 Late Pleistocene environmental and climatic evolution in southern South America. Bamberger Geographische Schiriften Bd. 11: 271-278. [ Links ]
Markgraf, V. y H. D´Antoni 1978 Pollen Flora of Argentina. The University of Arizona Press, Tucson. [ Links ]
McLauchlan, K. 2003 Plant cultivation and forest clearence by prehistoric North Americans: pollen evidence from Port Ancient, Ohio, USA. The Holocene 13 (4): 557-566. [ Links ]
Medina, M. 2008 Diversificación económica y uso del espacio en el tardío prehispánico del Norte del valle de Punilla, Pampa de Olaen y Llanura Noroccidental (Córdoba, Argentina). Tesis Doctoral inédita, Facultad de Filosofía y Letras, Universidad de Buenos Aires, Buenos Aires. [ Links ]
Medina, M., L. López y E. Berberián 2007 Agricultura y recolección en el Tardío Prehispánico de las Sierras de Córdoba (Argentina): el registro arqueobotánico de C.Pun.39. Centro de Estudios Históricos "Profesor Carlos S.A. Segreti", Córdoba. MS. [ Links ]
Moseholm, L., R. Weeke y B. Peterson 1987 Forecast of pollen concentration of Poaceae (Grasses) in the air by Time Series Analysis. Pollen et Spores 29: 305-322. [ Links ]
Nuñez, C. y J. Cantero 2000 Las plantas medicinales del Sur de la Provincia de Córdoba. Universidad Nacional de Río Cuarto, Río Cuarto. [ Links ]
Oliszewski, N. 2004 Estado actual de las investigaciones arqueobotánicas en sociedades agroalfareras del área Valliserrana del Noroeste Argentino (0-600 d.C). Relaciones de la Sociedad Argentina de Antropología XXIX: 211-227. [ Links ]
Pastor, S. 2007 Arqueología del valle de Salsacate y pampas de altura adyacentes (Sierras Centrales de Argentina). Una aproximación a los procesos sociales del período prehispánico tardío (900-1573 d.C.). Tesis Doctoral inédita, Facultad de Ciencias Naturales y Museo, Universidad Nacional de La Plata, La Plata. [ Links ]
Pastor, S. y M. Medina 2005 El uso Prehispánico Tardío de los ambientes serranos de altura. Investigaciones Arqueológicas en la Pampa de Achala, San Luis y Olaen (Córdoba, Argentina). La Zaranda de Ideas 1: 39-54. [ Links ]
Pearsall, D. 1995 "Doing" paleoethnobotany in the Tropical Lowlands: adaptation and innovation in methodology. En Archaeology in the Lowland American Tropics. Current analytical methods and recent applications, editado por P. Stahl, pp. 113- 129. Cambridge University Press, Cambridge. [ Links ]
Pearsall, D. 2004 Plants and people in ancient Ecuador: the ethnobotany of the Jama River Valley. Case Studies in Archaeology. Jeffrey Quilter Series Editor, Belmont. [ Links ]
Perry, L. 2004 Starch analyses reveal the relationship between tool type and function: an example from the Orinoco valley of Venezuela. Journal of Archaeological Science 31: 1069-1081. [ Links ]
Piovano, E., D. Ariztegui, S. Bernasconi y J. McKenzie 2004 Stable isotopic record of hydrological changes in subtropical Laguna Mar Chiquita (Argentina) over the last 230 years. The Holocene 14 (4): 535-535. [ Links ]
Piperno, D. 1995 Plant microfossils and their application in the New World tropics. En Archaeology in the Lowland American Tropic. Current analytical methods and recent applications, editado por P. Stahl, pp. 130-153. Cambridge University Press, Cambridge. [ Links ]
Piperno, D. 2006 Phytoliths. A Comprehesive guide for archaeologist and paleoecologist. Altamira Press, Lanham. [ Links ]
Piperno, D. y D. Pearsall 1998 The origins of agriculture in the Lowland Neotropic. Academic Press, San Diego. [ Links ]
Pohl, M., K. Pope, J. Jones, J. Jacob, D. Piperno, S. de France, D. Lentz, J. Gifford, M. Danforth y K. Josserand 1996 Early agriculture in the Maya Lowlands. Latin American Antiquity 7 (4): 355-372. [ Links ]
Prentice, I. 1985 Pollen representation, source area and basin size: toward a unified theory of pollen analysis. Quaternary Research 23: 76-86. [ Links ]
Prieto, A. 1996 Late Quaternary vegetational and climatic changes in the pampa grassland of Argentina. Quaternary Research 45: 73-88. [ Links ]
Prieto, A.2000 Vegetational history of the Late glacial - Holocene transition in the grasslands of eastern Argentina. Palaeogeography, Palaeoclimatology, Palaeoclimatology 157: 167-188. [ Links ]
Recalde, A. 2007 Representaciones rupestres del período agroalfarero en el sector Oeste de la Provincia de Córdoba. Arqueología 14. En prensa. [ Links ]
Reille, M. 1992 Pollen et Spores d´Europe et d´Afrique du Nord. Laboratoir de Botanique Historique et Palynologie, Marseille. [ Links ]
Rivero, D. 2001 Movilidad logística y sitios de ocupación breve en comunidades formativas prehispánicas de las Sierras de Córdoba. Tesis de Licenciatura inédita, Facultad de Filosofía y Humanidades, Universidad Nacional de Córdoba, Córdoba. [ Links ]
Sayago, M. 1969 Estudio fitogeográfico del Norte de Córdoba. Boletín de la Academia Nacional de Ciencias de Córdoba XLVI (2-4). [ Links ]
Smith, B. 1992 Rivers of Change. Essays on early agriculture in Eastern North America. Smithsonian Institution Press, Washington. [ Links ]
Smith, B. 2006 Documenting domesticated plants in the archaeological record. En Documenting domestication. New genetic and archaeological paradigm, editado por M. Zeder, D. Bradley, E. Emshwiller y B. Smith, pp. 15-24. University of California Press, Berkeley. [ Links ]
Trivi de Mandri, M., L. Burry y H. D´Antoni 2006 Dispersión-depositación del polen actual en Tierra del Fuego, Argentina. Revista Mexicana de Biodiversidad 77: 89-95. [ Links ]
van Geel, B., D. P. Hallewas y J. P. Pals 1982-1983 A Late Holocene deposit under the Westfriese Zeedijk near Enkhuizen (Prov. of Noord-Holland, The Netherlands): palaeoecological and archaelogical aspects. Review of Palaeobotany and Palynology 38: 269-335. [ Links ]
Vilanova, I., A. Prieto y S. Stutz 2006 Historia de la vegetación en relación con la evolución geomorfológica de las llanuras costeras del Este de la Provincia de Buenos Aires durante el Holoceno. Ameghiniana 43: 147-159. [ Links ]

