










Servicios Personalizados
Revista

Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkIntersecciones en antropología
versión On-line ISSN 1850-373X
Intersecciones antropol. v.10 n.1 Olavarría ene./jun. 2009
ARTÍCULOS
Arqueofaunas, osteometría y evidencia artefactual en Pastos Grandes, Puna de Salta: secuencia de cambio a lo largo del Holoceno temprano, medio y tardío en el sitio Alero Cuevas
Gabriel E. J. López
Gabriel E. J. López. CONICET - Instituto de Arqueología, Facultad de Filosofía y Letras, Universidad de Buenos Aires. 25 de mayo 217, 3° piso. Buenos Aires. Argentina. E-mail: gabelope@yahoo.com
Recibido 17 de Julio 2008
Aceptado 09 de Octubre 2008
RESUMEN
Este trabajo presenta los análisis de diversidad taxonómica y osteometría del sitio Alero Cuevas, en Pastos Grandes, Puna de Salta, con el fin de analizar los cambios en el consumo de recursos faunísticos y en el nicho económico a lo largo del Holoceno. Las estrategias de caza primero, y pastoreo posteriormente, fueron predominantes en el nicho económico de los grupos humanos de Pastos Grandes. La osteometría es una línea de evidencia que sirve como punto de partida para el estudio de los cambios relacionados con la domesticación y/o introducción de camélidos domesticados. En este sentido, se considera relevante distinguir la domesticación de la consolidación de una estrategia pastoril. Este último proceso ocurrió a lo largo del Holoceno tardío, convirtiéndose en la estrategia predominante del nicho económico. De todas formas la caza siguió siendo importante como estrategia económica. Finalmente, de manera complementaria se consideraron otras evidencias arqueológicas, principalmente los artefactos líticos, que en asociación con la información proveniente de los conjuntos arqueofaunísticos, fueron relevantes para comprender los cambios ocurridos en el nicho económico.
Palabras clave: Nicho económico; Arqueofaunas; Osteometría; Artefactos; Pastos Grandes.
ABSTRACT
Archaeofaunal, Osteometrical And Artefactual Evidence In Pastos Grandes, Puna Of Salta: The Sequence Of Change Through The Early, Middle And Late Holocene In Alero Cuevas Site. This paper presents an analysis of the taxonomic and osteometric diversity of the Alero Cuevas site, Pastos Grandes, Puna of Salta, in order to analyze changes in the consumption of faunal resources and this economic niche throughout the Holocene. Hunting and subsequently pastoralism were the predominant strategies in the economic niche of the human groups of Pastos Grandes. The osteometríc data is a line of evidence that serves as a starting point for the study of the changes related to the domestication and/or introduction of domesticated camelids. In this sense, it is important to distinguish domestication from the consolidation of a pastoralist strategy. This latter process occurred throughout the Late Holocene, becoming the predominant strategy of the economic niche. Nevertheless, hunting continued as an important economic strategy. Finally, complementary archaeological evidence is considered, chiefly the lithic artifacts. The lithic technology presents significant information for understanding the changes that occurred in the economic niche.
Keywords: Economic niche; Archaeofaunal evidence; Osteometric data; Artifacts; Pastos Grandes.
INTRODUCCIÓN AL PROBLEMA DE INVESTIGACIÓN
El objetivo principal es estudiar los cambios en el nicho económico de Pastos Grandes, en la Puna de Salta, fundamentalmente en cuanto al consumo de recursos faunísticos y su relación con los cambios en otros aspectos del registro arqueológico. La cuenca de Pastos Grandes se encuentra a una altura promedio de 4000 msnm, y está conformada por las aguas que descienden de los nevados de Pastos Grandes a través de quebradas y vegas y desembocan en el salar del mismo nombre (Figura 1).
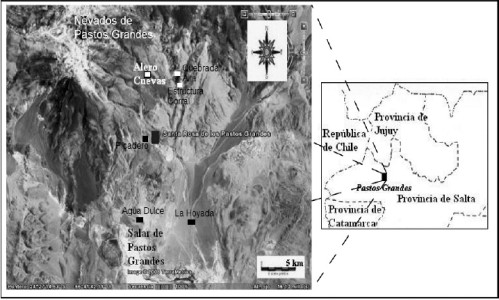
Figura 1. La cuenca de Pastos Grandes en el contexto regional y los sitios arqueológicos detectados.
El estudio del nicho económico de las poblaciones humanas en el pasado es fundamental para explicar la adaptación de las mismas al ambiente. El nicho económico hace referencia a las adaptaciones de un organismo a su entorno (Pianka 1982). En los organismos humanos implica la adaptación a partir de estrategias económicas particulares. La organización del nicho económico puede basarse en la existencia de una estrategia predominante, a partir de la cual se estructura la adaptación a un determinado entorno (Muscio 2004; Yacobaccio et al. 1997).
En la Puna en general, y en Pastos Grandes en particular, el consumo de recursos faunísticos fue fundamental para la adaptación humana (López 2008). Particularmente el consumo de los camélidos, los recursos faunísticos de más alto rendimiento en la Puna. Yacobaccio (2001a), plantea la existencia de un proceso de intensificación en el consumo de los camélidos en los Andes Centro Sur, el cual habría implicado un proceso local de domesticación. A lo largo del Holoceno, la caza y posteriormente el pastoreo fueron las estrategias predominantes del nicho económico de muchos grupos locales de la Puna (Yacobaccio et al. 1997). Sin embargo, la conformación de los nichos económicos puede haber implicado una alta variabilidad local, dado que en algunas áreas como Antofagasta de la Sierra, en la Puna de Catamarca o San Antonio de los Cobres, en la Puna de Salta, existen sectores aptos para la agricultura y la recolección de vegetales. De esta forma, Muscio (2004), propone que en la Quebrada de Matancillas, Puna de Salta, la agricultura de baja escala fue la estrategia predominante que determinó la organización del nicho económico durante el Período Temprano (ca. 2000 AP). Asimismo, Babot (2006), propone un proceso de intensificación en Antofagasta de la Sierra en el uso de artefactos de molienda y vegetales a partir de finales del Holoceno medio.
Por lo tanto, la organización del nicho económico en la Puna varía en la escala local y esto está condicionado especialmente por la disponibilidad de los ambientes aptos para determinadas estrategias económicas como la agricultura. En otras palabras, la variabilidad ambiental de la Puna explicaría las distintas estrategias económicas desarrolladas por los grupos locales, lo cual se desprende de un postulado básico de la ecología del comportamiento humano: la adaptación es una respuesta a la diversidad del entorno local (Boone y Smith 1998; Smith 1992).
Sin embargo, en líneas generales, la caza primero y el pastoreo después fueron estrategias esenciales para la adaptación humana en la Puna y la organización de los nichos económicos. A partir del establecimiento del pastoreo como estrategia económica, en distintas áreas se consolidaron nichos económicos de pastoreo-caza (Yacobaccio et al. 1997). De esta forma, en este trabajo se prestará especial atención al consumo de recursos faunísticos en el área de estudio, particularmente a partir de la secuencia del sitio Alero Cuevas, con una cronología que se extiende a lo largo del Holoceno temprano, medio y tardío. Esto tiene especial interés, dado que se espera que tanto la recolección como la agricultura a nivel local, no hayan sido estrategias económicas sustentables en el largo plazo en la organización del nicho económico de las poblaciones de Pastos Grandes, principalmente por la altura del área y distintos condicionantes ecológicos que impiden el normal crecimiento de una diversidad de cultivos. Las fuentes etnohistóricas y etnográficas consultadas así lo atestiguan (Abeledo 2006; Cerri 1993). Sin embargo, el consumo de vegetales producto de la interacción con grupos de otras áreas pudo haber sido una estrategia complementaria no predominante en el nicho económico, pero necesaria para la adaptación humana.
EL CONSUMO DE RECURSOS FAUNÍSTICOS EN LA PUNA
El consumo de recursos faunísticos en la Puna a lo largo del Holoceno se centró en los camélidos, tanto silvestres como domesticados (Cartajena et al. 2003; Yacobaccio et al. 1994). La domesticación produjo cambios morfológicos y genéticos que diferenciaron a los camélidos domesticados de sus parientes silvestres. Puede ser definido como el proceso de adaptación de una población de animales a los humanos y al ambiente de cautividad por cambios genéticos que ocurren en varias generaciones y eventos de desarrollo ambientalmente inducidos (Yacobaccio y Vilá 2002).La domesticación produce el aislamiento genético y cambia el fenotipo del segmento poblacional domesticado (Yacobaccio 2001b).En la actualidad, los camélidos domesticados están compuestos por la llama (Lama glama) y la alpaca (Lama pacos), y los camélidos silvestres por la vicuña (Vicugna vicugna) y el guanaco (Lama guanicoe). Sin embargo, se recuerda que la alpaca no habita la Puna Argentina y no hay evidencias de su presencia en épocas prehispánicas en la región (Olivera 1997). Por lo tanto, la llama habría sido el único camélido domesticado de la Puna Argentina. No es la intención profundizar aquí en la etología de los camélidos, simplemente se recuerda que estos recursos son predecibles y altamente disponibles en la Puna, mientras no sufran situaciones graves de estrés ambiental. Por lo tanto, a partir del modelo de amplitud de la dieta, cuyas variables clave son el retorno o rendimiento individual y la abundancia, se predice un alto consumo de estos recursos, dado que se combinan las dos variables necesarias para su inclusión en la dieta. En particular, se destaca que los rankings de recursos muestran a los camélidos en los primeros lugares, aunque con diferencias entre los silvestres y los domesticados (Muscio 2004). Por lo tanto, es necesario marcar características diferenciales entre la caza y el pastoreo como estrategias económicas para obtener estos recursos.
La caza implica la obtención directa de biomasa animal y el cazador considera a los animales como un alimento ya presente en el ambiente que explota. Por el contrario, el pastoreo tiene como objetivo la conversión de biomasa vegetal (pasturas) en biomasa animal para el consumo humano y para el pastor los animales son un capital que acumula y reproduce (Chang y Koster 1986). Los animales domesticados requieren el control humano de su reproducción, alimentación y organización del territorio, al contrario de los animales silvestres (López y Medina 2001). En consecuencia, el pastoreo suma costos de manejo con respecto a la caza pero anula los costos de búsqueda (López 2003). La llama tiene un valor extra por su capacidad para el transporte y la utilización de su fibra (Olivera 1997). En este sentido, fue propuesto un modelo de intercambio basado en las caravanas de llamas (Nuñez 1994; Nuñez y Dillehay 1979).
Por lo tanto, la caza y el pastoreo deben ser vistas como dos estrategias diferentes para la obtención de recursos de alto rendimiento con distintos costos. Las escalas temporales de largo plazo manejadas aquí permiten analizar los cambios en el consumo de recursos faunísticos y los cambios en las estrategias económicas a lo largo del tiempo. Como se señaló anteriormente, durante el Holoceno, en la Puna se evidencia un crecimiento en la representación de los camélidos en los conjuntos arqueofaunísticos. Esto se vincularía a distintas estrategias, tales como un aumento de la caza de estos recursos, el manejo de segmentos poblacionales por mecanismos como el protective herding en el marco del proceso de domesticación, y posteriormente la introducción de una estrategia pastoril (Yacobaccio 2001b).
Tomando como muestra 18 sitios de los Andes Centro Sur, Yacobaccio (2001a), expone la tendencia al incremento en el consumo de los camélidos a lo largo del Holoceno. En el Holoceno temprano se evidencia una mayor variabilidad en el consumo de los recursos faunísticos, incluyendo camélidos, roedores (principalmente vizcachas), y cérvidos en menor medida. Durante el Holoceno medio, se comienza a evidenciar el incremento del consumo de los camélidos en el promedio de los conjuntos relevados, mientras que esta tendencia se acentúa durante el Holoceno tardío (Yacobaccio 2001a).
La evidencia del sitio Alero Cuevas, correspondiente a las distintas etapas del Holoceno, representa nueva información que aporta a la discusión acerca de los cambios en el consumo de recursos faunísticos y las estrategias económicas.
EL SITIO ALERO CUEVAS: CONJUNTOS ARQUEOFAUNÍSTICOS Y DIVERSIDAD TAXONÓMICA
Los conjuntos arqueofaunísticos analizados provienen de las distintas capas fechadas en el sitio Alero Cuevas, ubicado en la cuenca de Pastos Grandes, Puna de Salta (López 2008). El sitio Alero Cuevas, se ubica en la quebrada del mismo nombre, en un talud de un cerro que rodea las mismas. El ancho es de ca. 20 m y presenta ca. 9 m de cobertura en la parte más profunda hacia la línea de goteo. Por lo tanto se trata de un alero con una amplia disponibilidad para el refugio humano, principalmente teniendo en cuenta la escasez de los mismos en el área.
La investigación en Pastos Grandes, y más particularmente en el sitio Alero Cuevas, se realizó en el marco de un proyecto de doctorado (López 2008).
Se trataba de un área con un vacío de información arqueológica, por lo que la metodología distribucional fue adecuada para realizar un acercamiento a las características del registro arqueológico en la escala areal (e.g., Belardi 1992; Borrero et al. 1992; Ebert 1992; Muscio 2004; Ratto 2003; entre otros). En este sentido, la excavación del sitio Alero Cuevas se inscribió dentro de esta metodología y se excavaron 4 cuadrículas de 1 x 1 m y un sondeo de 0,5 x 0,5 m. Esto se hizo con el fin de establecer las características estratigráficas, arqueológicas y cronológicas de las ocupaciones humanas. La excavación permitió registrar una larga secuencia estratigráfica con fechados en el Holoceno temprano, medio y tardío (López 2008).
Para el estudio de las arqueofaunas, las unidades básicas de análisis fueron los especímenes. Los conjuntos arqueofaunísticos se conformaron a partir de su correspondencia con cada una de las capas fechadas.
Los conjuntos analizados correspondieron a la capa F4, con fechados sin calibrar entre 9650 ± 100 AP y 8504 ± 52 AP, que la ubican en el Holoceno temprano (López 2008); a la capa F3, con fechados de 6510 ± 80 AP y 6506 ± 58 AP (Holoceno medio); a la capa F2, fechada en 4210 ± 70 AP entre finales del Holoceno medio y comienzos del Holoceno tardío; y por último a dos capas correspondientes al Holoceno tardío, en contextos donde se espera la consolidación del pastoreo como estrategia predominante. Estas son las capas C1, con un fechado de 2020 ± 60 AP y X, con un fechado de 643 ± 35 AP (Figura 2).
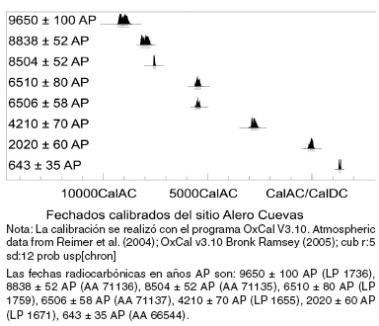
Figura 2. Fechados radiocarbónicos calibrados del sitio Alero Cuevas (tomado de López 2008).
Cada uno de los conjuntos fue analizado siguiendo la propuesta de Mengoni Goñalons (1999), incluyendo la identificación anatómica y taxonómica, la cuantificación a partir de distintas medidas, el control tafonómico de las muestras y la osteometría para conocer la diversidad de especies de camélidos (López 2008). Entre todos estos aspectos, este trabajo se centrará en el estudio de la diversidad taxonómica, considerando como límite máximo de identificabilidad el nivel de Familia, y en la determinación de especies de camélidos a partir de osteometría y evidencias contextuales. De todas formas, brevemente, es importante destacar que el control tafonómico de los conjuntos analizados indicó una tendencia a una alta integridad (Binford 1981; Lyman 1994). Las bajas o nulas correlaciones de la representación de partes esqueletarias con la densidad; la predominancia de estadios de meteorización bajos (principalmente entre 0 y 2); la baja frecuencia de marcas de carnívoros y otros agentes naturales (salvo los roedores en las capas superiores y sectores puntuales de la estratigrafía); y la presencia de marcas antrópicas en todos los conjuntos, relacionadas al descarne, la desarticulación y la extracción de médula, indican en conjunto que el agente humano fue el principal generador y modificador del registro analizado (para un estudio detallado ver López 2008). A su vez, existe una clara asociación contextual de los conjuntos arqueofaunísticos con el resto de la evidencia arqueológica en cada una de las capas.
Con respecto a la representación de partes esqueletarias, también brevemente, se señala que en todos los conjuntos, en mayor o menor medida, se evidencia una alta diversidad de unidades anatómicas, tanto del esqueleto apendicular como del esqueleto axial (para un estudio más detallado ver López 2008).
A nivel taxonómico, el conjunto de la capa F4 (ca. 9650 AP - 8500 AP) está compuesto por un total de 631 especímenes identificados al nivel de Familia. La cuantificación por NISP indicó la representación de 575 especímenes de camélidos y un porcentaje de 91,12%. El MNI indicó la presencia de 8 individuos. Por su parte se recuperaron 55 especímenes de chinchíllidos que representaron el 8,71%. El MNI de chinchíllidos permitió registrar 4 individuos, por lo que también a partir de esta medida se estableció un predominio de camélidos. Asimismo se registró un solo espécimen de cérvido (0,15% en NISP%).
El conjunto de la capa F3 (ca. 6500 AP) presentó 258 especímenes identificados al menos hasta el nivel de Familia, registrándose 243 asignables a camélidos (94,18% del NISP%). El MNI fue de 5 individuos. Los chinchíllidos fueron detectados en solo 14 especímenes (5,42%) y el MNI indicó la presencia de 1 individuo. También se recuperó un espécimen de zorro (Ducisyon sp.), que por el momento no puede descartarse ni su entrada por razones tafonómicas ni por cuestiones antrópicas. Igualmente lo tafonómico tiene más sustento si se tiene en cuenta que se trata de un solo espécimen y sin ninguna evidencia antrópica. En relación con las frecuencias de los taxones, es importante destacar el aumento de los camélidos con respecto al Holoceno Temprano y el decrecimiento de los chinchíllidos. Sin embargo, estas frecuencias, comparativamente, permiten señalar que los camélidos siempre ocuparon un lugar predominante en la dieta de las poblaciones del área.
El conjunto de la capa F2 (ca. 4200 AP) indicó una más baja diversidad taxonómica que los anteriores. En particular se evidenció una concentración aún más notable en el consumo de camélidos. El conjunto total de especímenes identificados al nivel de Familia comprendió 345. De ellos, 327 correspondieron a camélidos, que representaron el 94,78% del total NISP. El MNI permitió cuantificar 3 individuos. Los chinchíllidos continuaron la reducción de su frecuencia, con un porcentaje de 5,21% (18 especímenes). El MNI fue de 1 individuo. Por lo tanto se destaca que sólo se relevaron dos taxones al nivel de Familia.
El conjunto de la capa C1 (ca. 2000 AP), presentó un total de 639 especímenes al nivel de Familia. Al igual que en el conjunto de la capa F2, sólo se encontraron representados los camélidos y los chinchíllidos. Los camélidos fueron los taxones más ampliamente
consumidos, evidenciados en 605 especímenes (94,67%). El MNI fue de 4 individuos. Asimismo se recuperaron 34 especímenes asignables a chinchíllidos que contribuyeron en 5,32% del NISP. El MNI fue de 2 individuos. También se recuperaron fragmentos de cáscara de huevo de suri que no se consideraron en el NISP.
El conjunto de la capa X (ca. 640 AP) presentó la menor diversidad taxonómica de la secuencia. Asimismo fue el conjunto más reducido al nivel de Familia (N= 100), lo que podría deberse a la acotada extensión horizontal y vertical de esta capa. Los camélidos representaron el 100% del conjunto en NISP. El MNI fue de 4 individuos.
Por lo tanto, en relación con el objetivo de este trabajo, en primer lugar es importante señalar que los conjuntos arqueofaunísticos presentan una muy baja diversidad taxonómica. La predominancia de los especímenes de camélidos es notable en todos los conjuntos. Es importante señalar que la fragmentación de los conjuntos de camélidos no habría afectado las frecuencias registradas en NISP, tal como lo indica la predominancia de estos taxones sobre los chinchíllidos en MNI. Al respecto la razón entre el NISP y el MNE (índice de fragmentación) no se correlaciona con los valores de NISP% de cada uno de los conjuntos (r de Spearman -0,2 p 0,74).
Igualmente, más allá de esta mayoría de camélidos, se evidencian algunos cambios en las frecuencias de los taxones, especialmente entre el Holoceno temprano y el Holoceno medio en adelante. Hacia el Holoceno medio existe una mayor concentración en el consumo de los camélidos que en el Holoceno temprano y una disminución de la frecuencia de chinchíllidos. De todas maneras, los cambios porcentuales entre los dos conjuntos son bajos, por lo que no existe una clara tendencia hacia la intensificación de los camélidos en cuanto a la frecuencia representada de estos taxones (Figura 3).
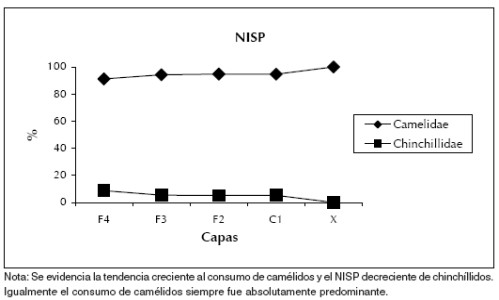
Figura 3. NISP de los camélidos y los chinchíllidos a lo largo del Holoceno en las capas fechadas del Alero Cuevas.
OSTEOMETRÍA
La osteometría es una de las técnicas que más se usó para discriminar entre especies de camélidos en la Puna. En general, se reconoce que los resultados obtenidos por esta técnica no pueden tomarse de forma aislada, ya que sólo ilustran una tendencia en la representación de especies, diferenciadas por el gradiente de tamaño de los camélidos. En este sentido el gradiente de tamaño en la Puna Argentina de mayor a menor es llamas-guanacos-vicuñas. Uno de los principales limitantes de esta técnica es la escasez de muestras de referencia tomadas como estándar actual, particularmente en el caso del guanaco del Noroeste Argentino. Esta especie es relevante debido a su posición intermedia en el rango de tamaño de los camélidos en los Andes Centro Sur. En este sentido vale aclarar que no se pueden tomar como estándar las medidas procedentes de guanacos de la Patagonia, debido a que son más grandes que los del Noroeste Argentino. Por lo tanto, considerando estas limitaciones, igualmente se prefirió comparar con la muestra de guanaco actual disponible (en este caso un ejemplar de los Valles Calchaquíes medido por D. Elkin y G. Mengoni Goñalons, comunicación personal 2008) y al menos realizar una aproximación tentativa a la diversidad de tamaños de los camélidos. Se reconoce que este valor tomado como estándar no puede representar a toda la especie y su variabilidad poblacional. Tampoco se conoce su variabilidad en tamaño a lo largo del Holoceno, lo cual es esperable en una escala cronológica de largo plazo en ambientes cambiantes (ver Mengoni Goñalons y Yacobaccio 2006). Igualmente, si bien se reconocen las limitaciones planteadas, las mediciones sobre especímenes arqueológicos y su comparación con valores actuales es relevante para mostrar una tendencia en la variación de tamaños en los conjuntos a lo largo del tiempo.
En general, se considera que es factible distinguir un grupo de tamaño pequeño (que corresponde a las vicuñas) de un grupo de tamaño más grande (en cuyo extremo se encuentran las llamas). Por este motivo los valores intermedios podrían interpretarse como correspondientes a guanacos, aunque lamentablemente no se cuenta con parámetros claros para determinar la variabilidad intrapoblacional de esta especie y hasta qué valor límite o rango de tamaño puede ser considerado dentro de una especie u otra (especialmente para la distinción entre llama y guanaco). Con estas advertencias, aquí se usará la técnica de Meadow (1987) basada en las distancias de logaritmos para mostrar patrones tentativos en la variabilidad de tamaños en las mediciones realizadas sobre falanges primeras y comparadas con el estándar de guanaco actual. Si bien se cuenta con otras mediciones estándar de llamas y vicuñas actuales, la comparación con la medición de guanaco parece más apropiada en términos de esta técnica teniendo en cuenta que presenta un carácter intermedio de tamaño. La técnica de Meadow (1987) de las distancias de logaritmos, se basa en la fórmula: log x - log z= (log /z).
La fórmula expresa las distancias logarítmicas entre la medida del espécimen arqueológico (x) y la medida estándar homóloga del camélido actual (z). Esto se hizo con la intención de mostrar más claramente las diferencias entre las distintas mediciones. De esta forma, a partir de un estándar actual (representado por la línea cero), se marcan tendencias en la representación de los tamaños de los huesos. Los valores negativos (ubicados hacia la izquierda del estándar actual de guanaco representado por la línea cero), se corresponderían con vicuñas, mientras que los valores mayores (ubicados hacia la derecha de la línea cero) podrían representar llamas o guanacos muy grandes. Si se tomaran valores de vicuñas como estándar, los valores de estos taxones se agruparían en torno a cero y los valores de guanaco y llama se correrían claramente hacia la derecha. Por el contrario si se tomaran valores estándar de llamas los valores de guanacos y especialmente de vicuñas se distanciarían claramente hacia la izquierda adquiriendo valores negativos altos.
Para el cálculo de las distancias de logaritmos se seleccionó una sola variable de medición de las falanges primeras, el ancho máximo de la cara articular proximal, dado que de esta forma se puede realizar una comparación precisa (ver Elkin 1996). Si bien existen varios huesos discriminantes que probablemente resultan útiles a nivel osteométrico, las falanges primeras suelen estar altamente representadas y bien preservadas en el registro arqueológico, por lo que fueron ampliamente usadas en el Noroeste Argentino (Elkin et al. 1991). En particular, se ha usado el ancho de la cara articular proximal, la cual resulta una medida relevante a nivel comparativo y regional. Las mediciones de la cara articular proximal de guanaco andino, vicuñas y llamas actuales medidas por Elkin, Mengoni Goñalons y Yacobaccio (ver Mengoni Goñalons y Yacobaccio 2006), son las que muestran menor variabilidad entre la falange delantera y trasera, por lo que a los fines de este trabajo se prefirió no diferenciar a las mismas. Esto se realizó con el objetivo de no perder información (mediciones) en varios casos en los que no se puede diferenciar claramente entre falange trasera y delantera.
Si bien a lo largo de la secuencia se midieron 62 especímenes de todas las capas y conjuntos del Alero Cuevas, en este trabajo la muestra se circunscribe a 22 especímenes de falanges primeras en las cuales se pudo tomar la medición del ancho de la cara articular proximal (para un detalle de los datos de las otras mediciones ver López 2008). De esta forma, si bien la muestra es más acotada resulta más clara para indicar tendencias a lo largo del Holoceno a nivel comparativo. Por el contrario, otras partes esqueletarias, más allá de las falanges, no se encontraron presentes reiteradamente en las distintas capas (y en condiciones de medición), por lo cual no fueron adecuadas para establecer tendencias en la variación temporal. Inclusive aquí no se consideraron otros 10 especímenes de falange primera por no presentar la posibilidad de medición de la cara articular proximal y en consecuencia no sirvieron para comparar entre todas las capas para observar tendencias en las variaciones de tamaño a lo largo del Holoceno. En este sentido, es posible determinar cambios en el tamaño de los huesos que podrían ser relacionados al proceso de domesticación de los camélidos.
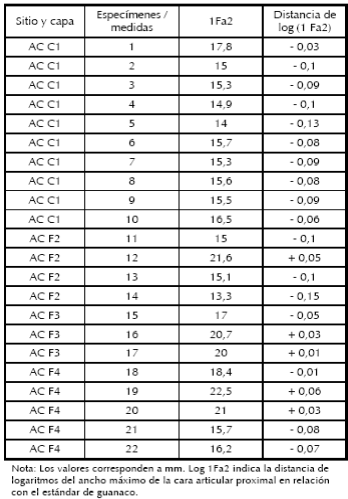
Para la selección de los especímenes medidos se siguieron los criterios de Elkin et al. (1991), en particular, tener madurez osteológica, buena preservación, no estar termoalterados y presentar los puntos de medición adecuados, lo cual acota la muestra. Igualmente, aunque no se consideraron en este trabajo, para todos los especímenes se tomaron las distintas mediciones posibles en cada uno, ya que son datos importantes que pueden servir para comparar con otros sitios (ver López 2008). En la Tabla 1 se observan las distintas mediciones realizadas sobre el ancho de la cara articular proximal de la falange primera.
Tabla 1. Osteometría del ancho máximo de la cara articular proximal de la falange 1, capas C1 (ca. 2000 AP), F2 (ca. 4200 AP), F3 (ca. 6500 AP) y F4 (ca. 9650-8500 AP) del Alero Cuevas.
Los análisis por osteometría permitieron detectar variaciones en el tamaño de los huesos de camélidos medidos. Estas variaciones se reflejan principalmente a través de valores negativos y positivos (a la izquierda y la derecha del estándar, respectivamente), que indicarían probablemente la presencia de distintas especies de camélidos. Particularmente importante resulta el hecho de poder distinguir entre especies domesticadas y silvestres, dado que implican estrategias económicas distintas (pastoreo y caza) (Izeta y Scattolin 2006).
En su mayoría se registran valores negativos, aún en la capa de 2000 AP, cuando se esperan valores más altos (Tabla 1, Figura 4). Justamente los valores negativos representan los tamaños más pequeños, correspondientes probablemente a vicuñas. De esta forma, la alta presencia de vicuñas en las mediciones efectuadas indica la importancia de la caza de camélidos silvestres aún en momentos donde el pastoreo se encuentra consolidado como estrategia económica de los grupos de la Puna. La discusión más interesante y problemática surge en relación con los valores positivos. En el conjunto de la capa F4 (Holoceno temprano), dos especímenes muestran valores positivos que podrían interpretarse como guanacos, uno de tamaño más grande que el otro (Figura 4). Esta interpretación se debe a que el guanaco presentaría rangos de tamaño superiores a los actuales, tal como se evidencia a partir de las mediciones de huesos en otros sitios de la Puna Argentina, provenientes de contextos fechados en el Holoceno temprano, lo que marcaría un decrecimiento en su tamaño a lo largo del Holoceno (Mengoni Goñalons y Yacobaccio 2006). En la capa F3 (Holoceno medio), los valores positivos también podrían interpretarse como guanacos, ya que se encuentran más cerca del estándar. Además ninguno de ellos presenta un valor tan alto como uno de los especímenes del Holoceno temprano, representativo de rangos de tamaño de guanaco más grandes, por lo que es probable que durante el Holoceno medio hayan comenzado a adquirir tamaños más cercanos al estándar actual.
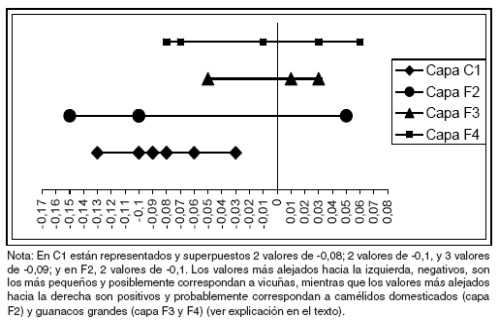
Figura 4. Gráfico de los valores estandarizados del ancho máximo de la cara articular proximal de la falange primera a partir de la distancia de logaritmos tomando un estándar de guanaco actual dado por la línea cero.
En la capa F2 (ca. 4200 AP), es sumamente interesante la división entre dos rangos de tamaño: uno pequeño correspondiente a vicuñas, y otro mucho más grande en comparación. Este último, presenta un tamaño bastante superior al estándar de guanaco actual (Figura 4), recuperado en un contexto arqueológico cuya cronología se vincula con los inicios de la domesticación de los camélidos. Por lo tanto, es probable su adscripción a un camélido domesticado y/o en proceso de domesticación. Igualmente se reconoce que se debe profundizar la investigación al respecto, dado que la muestra aún es pequeña, pero también se debe tener en cuenta que éste es un punto de partida para el estudio de los cambios ocurridos entre fines del Holoceno medio y comienzos del Holoceno tardío.
En la capa C1 (ca. 2000 AP) el ancho de la cara articular proximal de la falange primera, muestra valores probablemente correspondientes a vicuñas, aunque debe tenerse en cuenta que otras mediciones sobre este elemento óseo si bien ratifican esta tendencia aportan la presencia de especímenes mucho más grandes. Este es el caso de dos mediciones del largo máximo de la falange primera (1FA1), que claramente superan los estándares de vicuña y guanaco actual, y más aún el de las llamas actuales. Estas mediciones no se consideraron aquí porque correspondieron a especímenes en los que no pudo tomarse el ancho de la cara proximal debido a la fragmentación de algún sector necesario para esta medición. Estos especímenes corresponderían a camélidos domesticados, particularmente llamas de un rango de tamaño superior al actual. Esto mismo se observó en el sitio Matancillas cronológicamente comparable con la capa C1, ubicado también en la Puna de Salta, más precisamente en San Antonio de los Cobres (López 2003) con una economía cárnica basada en el pastoreo y la caza, aunque en el marco de un nicho donde la horticultura fue la estrategia predominante (Muscio 2004).
Por lo tanto, se puede sostener a partir de éste y otros indicadores contextuales el desarrollo de una estrategia pastoril consolidada hacia el 2000 AP, aunque probablemente el Alero Cuevas por su ubicación estratégica para la obtención de camélidos silvestres, principalmente vicuñas, refleje aún en contextos pastoriles la importancia económica de la caza. Con esto se desea señalar el rol del sitio Alero Cuevas como un punto estratégico para la caza a lo largo del Holoceno, lo cual se evidencia a partir de la osteometría. La importancia de la caza y su inserción en economías agropastoriles puneñas fue advertida por distintos autores, principalmente en aleros y cuevas (Escola 2002; Olivera y Elkin 1994; Yacobaccio et al. 1997).
En este sentido, aunque no se consideraron en este trabajo, en su mayoría, las mediciones efectuadas sobre la escápula, el metatarso, el húmero, el fémur, el radioulna, y la tibia indican valores correspondientes probablemente a vicuñas, aunque también hay evidencias relacionadas con camélidos domesticados (ver López 2008).
En síntesis, los análisis osteométricos permitieron mostrar una tendencia preliminar en la variación de tamaño del ancho de la cara articular proximal de la falange primera, que puede interpretarse a partir de la variación de especies de camélidos representadas en las distintas capas fechadas del Alero Cuevas. De todos modos, se debe seguir ampliando la muestra arqueológica y actual para avanzar en el análisis de la variabilidad intraespecífica e interespecífica.
EVIDENCIAS ARTEFACTUALES
Complementariamente con la información relevada a partir del análisis arqueofaunístico, es importante destacar brevemente la asociación contextual de evidencia artefactual en las distintas capas bajo estudio. En este sentido, desde el marco teórico de la ecología del comportamiento humano (Smith 1992), se priorizó el estudio de las estrategias tecnológicas, generales o particulares, para resolver problemas adaptativos (Nelson 1991), las cuales pueden variar desde un extremo compuesto por sistemas tecnológicos altamente costosos hasta aquellos de más bajo costo (Bousman 1993). Este último autor propone un continuum entre estrategias de maximización de recursos usando tecnologías costosas en un extremo y estrategias de minimización del tiempo compuestas de tecnologías poco costosas en el otro extremo.
Escola (2004) postula que la inversión de trabajo debe considerar distintas variables, la producción de la forma base, la estandarización en los soportes y el tipo de retoque en la formatización y regularización final de los instrumentos. En resumen, la combinación de los distintos atributos de los artefactos líticos (formatización, forma base, estandarización en el tamaño y materia prima) permite realizar un acercamiento a las estrategias tecnológicas vinculadas a maximizadores de recursos o minimizadores del tiempo (Bousman 1993). Esto es útil a los fines del marco teórico utilizado, y debe ser tomado como una aproximación cualitativa al grado de inversión energética. De este modo, las definiciones señaladas son lo suficientemente flexibles para dar cuenta de la variabilidad existente, dado que puede haber instrumentos costosos por su forma base pero no por su formatización o viceversa. En relación con la forma base, la diferenciación más inclusiva fue entre tecnología de lascas y tecnología de extracciones laminares. Esta última fue definida por la "obtención de formas alargadas de bordes laterales paralelos o subparalelos a partir de núcleos preparados con este objetivo y que presentan un patrón distintivo y recurrente en el registro lítico de los conjuntos del caso de estudio, con una relación entre el largo y el ancho del doble o más" (López 2008:159).
Entre las materias primas se diferenciaron locales (aquellas cuyas fuentes se ubican dentro de la cuenca de Pastos Grandes, en rangos no superiores a los 20 km de distancia) y no locales (ubicadas afuera de la cuenca de Pastos Grandes). Asimismo, dentro de las no locales, se consideraron intermedias las ubicadas en un rango no mayor de 50 km y lejanas las que superan este valor arbitrario, pero que refleja la distancia hacia otras áreas o cuencas como San Antonio de los Cobres o el salar de Pocitos-Quirón (López 2008). De todas formas, a los fines de reflejar los cambios notables en el uso de materias primas a lo largo de la secuencia del sitio Alero Cuevas, se sumaron todas las no locales, incluyendo las intermedias y las lejanas, las cuales se diferenciaron claramente de las locales (Figura 5).
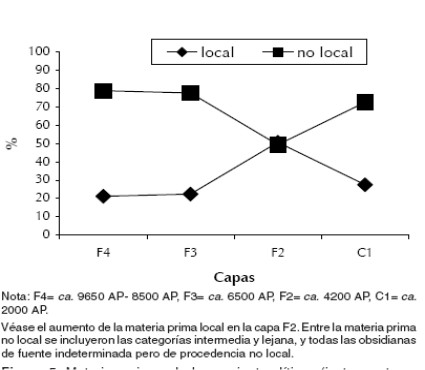
Figura 5. Materias primas de los conjuntos líticos (instrumentos y desechos de talla) en las capas del Alero Cuevas.
En la capa F4, correspondiente al Holoceno temprano, se estudió un conjunto lítico compuesto de 2098 desechos de talla y 45 instrumentos (Restifo 2007). Entre los desechos de talla predominaron las microlascas e hipermicrolascas en obsidiana no local, especialmente aquella proveniente de la fuente de Quirón distante más de 30 km del sitio Alero Cuevas. Por su parte, los instrumentos se caracterizaron en general por el bajo esfuerzo tecnológico, con bajos niveles de formatización, todos manufacturados sobre forma base de lascas. Dentro de ellos, se destacan las puntas triangulares apedunculadas por ser morfologías ampliamente expandidas en los Andes Centro Sur a lo largo del Holoceno temprano. De todas maneras, tampoco en las puntas se evidencian altos niveles de formatización (Restifo 2007). Tanto en los desechos de talla como en los instrumentos, la materia prima predominante fue no local. A nivel superficial en el área de estudio, los instrumentos que pueden ser indicadores de cronología relativa del Holoceno temprano son escasos y en general se hallaron en contextos aislados.
En la capa F3, del Holoceno medio, el descarte artefactual fue menor, registrándose una muestra de 70 especímenes entre desechos de talla e instrumentos. Esto se relacionaría a la conformación pedregosa de esta capa, quizás por procesos erosivos que generaron desprendimientos del techo del alero. De hecho, los fechados obtenidos en esta capa se superponen, lo que indicaría ocupaciones humanas discretas (Figura 2). La materia prima predominante fue no local, principalmente obsidiana de Quirón.
En la capa F2, fechada entre fines del Holoceno medio y comienzos del Holoceno tardío, el material asociado con el conjunto arqueofaunístico, corresponde a distintas clases de artefactos líticos, incluyendo desechos de talla, instrumentos y núcleos. Lo más llamativo resultó la recurrencia de una clase particular de artefactos a los que se denominó artefactos lanceolados unifaciales (López 2009), sobre forma base laminar y asociados en capa con un núcleo de tecnología de extracciones laminares. Una característica particular en casi todos los artefactos de esta clase es la presencia de retoque sectorizado en la cara ventral, generalmente en la parte proximal, posiblemente para reducir el espesor del bulbo y probablemente relacionado al enmangue. La asociación de estos artefactos y tecnología de extracciones laminares indica un cambio tecnológico notable no registrado en las otras capas. La proliferación de estos artefactos estandarizados en determinadas variables métricas y asociados con tecnología de extracciones laminares es recurrente a lo largo de la cuenca de Pastos Grandes y en áreas relativamente cercanas como San Antonio de los Cobres (Muscio 2004) y Salinas Grandes, en la Puna de Jujuy (Fernández Distel 1978). Asimismo, es notable la disminución de las materias primas no locales y el aumento de las materias primas locales en esta capa (Figura 5), y en las concentraciones de superficie relacionadas por cronología relativa con este conjunto como La Hoyada y Picadero. Los artefactos lanceolados unifaciales son interpretados como tecnología de procesamiento posiblemente relacionada a la necesidad de aumentar la eficiencia y/o minimizar el riesgo en un contexto de mayores presiones socioecológicas y posiblemente domesticación de camélidos, aunque no se descarta en algunos casos su uso como tecnología extractiva (Figura 6). Este contexto sería propicio para innovaciones tecnológicas con mayor inversión de energía pero eficientes para cumplir distintas tareas, aunque restan realizar análisis funcionales. Su recurrencia en el registro arqueológico indicaría mecanismos de transmisión cultural sesgada actuando sobre la variabilidad, lo cual es esperable en contextos de aumento del tamaño de los grupos y mayor complejidad tecnológica (Boyd y Richerson 1985). Asimismo, estos cambios fueron observados en el Norte de Chile para una cronología similar en sitios como Puripica 1, donde la proliferación de tecnología de procesamiento (cuchillos) asociada con variaciones en el tamaño de los huesos y un aumento de camélidos subadultos, fue interpretada en relación con el procesamiento intenso de estos recursos en un contexto de domesticación (Nuñez 1981).

Por lo tanto, es claro que entre fines del Holoceno medio y comienzos del Holoceno tardío se están generando procesos de cambio, evidenciados tanto en las arqueofaunas como en la tecnología lítica. En este contexto cronológico, se registró en el sitio Agua Dulce en el borde del salar de Pastos Grandes, un hallazgo bioarqueológico a cielo abierto, fechado en 3738 ± 46 AP (López y Miranda 2008). El mismo consiste de distintas partes esqueletarias de un individuo de sexo masculino asociadas a un artefacto lítico circular, plano y pulido en los bordes, que podría corresponder a tecnología ornamental, cuya frecuencia es muy alta en estos contextos cronológicos (Aschero y Yacobaccio 1999). Asimismo, la presencia de marcas antrópicas y un artefacto lítico incrustado en el radio derecho del individuo resulta un punto de partida para investigar hipótesis que den cuenta de competencia y conflictividad en este marco de mayores presiones socioecológicas (ver López 2008).o tanto, en Pastos Grandes, las estrategias predominantes del nicho económico se relacionaron al consumo de recursos faunísticos, especialmente los de más alto rendimiento, los camélidos. De esta forma, además de profundizar en los estudios arqueofaunísticos para dar cuenta de las estrategias económicas que desarrollaron los grupos humanos a lo largo del Holoceno en Pastos Grandes, también se incluirá en el estudio presentado aquí, la evidencia contextual relacionada, particularmente la tecnología lítica.
En la capa C1, fechada en ca. 2000 AP, las arqueofaunas se asocian con un conjunto lítico compuesto de desechos de talla e instrumentos. Entre estos últimos sólo las puntas de proyectil presentan mayor inversión de energía, todas con retoque bifacial extendido. Éstas se encuentran representadas en el 41,9% del conjunto de instrumentos. Este alto porcentaje de puntas de proyectil en conjunto con la evidencia proporcionada por la osteometría de un alto consumo de vicuñas, sustenta la importancia de la caza en contextos de pastoreo (ver Olivera y Elkin 1994; Yacobaccio et al. 1994). Por el contrario, la tecnología de procesamiento registra escasa inversión de energía, lo que muestra estrategias posiblemente distintas con respecto a la tecnología extractiva. La materia prima predominante vuelve a ser la no local, principalmente obsidiana de Quirón. En este contexto, donde se espera un nicho pastoril-cazador consolidado, en el marco de grupos con movilidad residencial reducida, las interacciones sociales habrían sido necesarias como forma de minimizar el riesgo (Escola 1996). Al respecto, la recuperación de cuentas de valva, una de ellas proveniente del Océano Pacífico, cañas macizas de tierras bajas, marlos de maíz posiblemente provenientes de valles mesotermales, entre otros hallazgos, indican el impacto de este proceso. En este sentido, Nuñez y Dillehay (1979) propusieron un modelo de interacción basado en el tráfico caravanero de llamas, aunque no se descartan otras formas de interacción. En cuanto a la cerámica, la misma presenta similitudes con otras de contextos puneños, particularmente por la tonalidad oscura y el pulido.
En este contexto, a nivel superficial se evidencia la proliferación de estructuras arquitectónicas en el sector de quebradas donde se encuentra el Alero Cuevas (particularmente en la Quebrada de Santa Rosa). Una de estas estructuras correspondiente al sitio Quebrada Alta fue fechada en 1488 ± 41 AP (López 2008). Por lo tanto, es probable que las mismas reflejen una mayor estabilidad residencial a lo largo del Holoceno tardío, particularmente en contextos donde el pastoreo se consolidó como la estrategia predominante. Entre las estructuras arquitectónicas, cabe destacar la presencia de varias asignables a corrales, una de las cuales, Estructura Corral, presenta un fechado de 859 ± 45 AP (López 2008). Este último fechado se relaciona también con el más tardío del sitio Alero Cuevas, de ca. 640 AP. Esta capa presenta conjuntos arqueológicos más reducidos, y entre el conjunto lítico se puede destacar la presencia de puntas de proyectil triangulares con pedúnculo y puntas lanceoladas chicas. También se registraron marlos de maíz, probablemente conseguidos a través de interacciones sociales.
DISCUSIÓN
Los análisis arqueofaunísticos permitieron detectar una baja diversidad taxonómica a lo largo de todas las capas. Asimismo se registró un consumo absolutamente mayoritario de camélidos por sobre el resto de los taxones, particularmente los chinchíllidos. Por lo tanto, no se evidencia un proceso de intensificación notable en el consumo de camélidos a lo largo del tiempo, pero sí una mayor concentración aún, a partir del crecimiento del NISP de estos recursos y el decrecimiento de los chinchíllidos. Esto es más notable desde el Holoceno medio (Figura 3), pero se destaca la importancia de los camélidos para el consumo humano desde los comienzos del Holoceno.
La osteometría permitió evidenciar variaciones en el tamaño de determinados especímenes óseos que fueron interpretados a partir del consumo de distintas especies de camélidos a lo largo del Holoceno. En el Holoceno temprano, la estrategia predominante en el nicho económico fue la caza de camélidos silvestres, y entre ellos tanto vicuñas como guanacos, lo que indica una diversificación a partir del consumo de recursos de muy alto retorno energético. En este sentido, se debe tener en cuenta que los guanacos del Holoceno temprano serían más grandes en tamaño que los actuales del Noroeste Argentino (Mengoni Goñalons y Yacobaccio 2006). Esto también quedó evidenciado por las mediciones osteométricas del sitio Alero Cuevas. Al respecto, la diversificación puede ser tomada como una estrategia de minimización del riesgo (Winterhalder et al. 1999), pero es importante destacar que en este caso se habría realizado sin perder la eficiencia en el consumo de recursos faunísticos (ingreso a la dieta de los de mayor retorno). Esta tendencia continuaría durante el Holoceno medio, ya que la osteometría evidenció el posible consumo de vicuñas y guanacos, aunque estos últimos de tamaño menor con respecto al Holoceno temprano.
Entre fines del Holoceno medio y comienzos del Holoceno tardío, continúa el consumo de vicuñas pero también se evidencian cambios osteométricos que podrían relacionarse con animales domesticados o en proceso de domesticación. En este sentido, es claro como un valor del ancho de la cara articular de la falange primera, aumenta en la capa correspondiente a esta cronología. De todas maneras no se puede sostener de forma concluyente la relación con la domesticación de camélidos, pero sí es un punto de partida importante para indagar acerca de este proceso. En esta capa también se evidenciaron cambios en el perfil etario, relacionados con el aumento de especímenes no fusionados y neonatos (López 2008), lo cual fue considerado por distintos autores como indicador de domesticación [e.g., Nuñez (1981) para el caso de Puripica1]. Más allá de estos indicadores de cambio en el conjunto arqueofaunístico en el sitio Alero Cuevas, las evidencias a nivel regional permiten plantear que en este contexto cronológico ocurrieron los cambios relacionados con la domesticación y/o introducción de camélidos domesticados (ver Yacobaccio 2001a, 2001b).
En relación con la evidencia contextual tanto del sitio Alero Cuevas como del área en general, se pueden señalar cambios importantes a nivel tecnológico concordantes con los cambios en los conjuntos arqueofaunísticos. La proliferación de una tecnología de extracciones laminares y su asociación como forma base de una clase particular denominada artefactos lanceolados unifaciales, tanto en capa como en superficie, indicarían a su vez el cambio de estrategia tecnológica, quizás en un contexto donde se privilegió el aumento de la eficiencia, en lo que Bousman (1993) denominó maximización de recursos, que implica tecnologías más costosas como la laminar (ver también Nelson 1991). Esto es esperable en contextos donde aumenta la demanda energética, ya sea de recursos silvestres o domesticados, particularmente a partir de mayores presiones dependientes de la densidad (Broughton 1999). Los cambios hacia la domesticación y/o introducción de camélidos domesticados serían una respuesta adaptativa a estas presiones. En un contexto de mayor segmentación ambiental en distintos parches de recursos por procesos generales de mayor aridez (Nuñez y Grosjean 1994), aumentarían los costos de dispersión espacial y en consecuencia la concentración de población en los mismos, disparando un aumento del tamaño de los grupos sociales y la competencia por los recursos. Por lo tanto, estas nuevas presiones socioecológicas podrían haber sido disparadores del cambio hacia nuevas estrategias tendientes a aumentar la eficiencia y/o minimizar el riesgo. La domesticación de camélidos y/o introducción de camélidos domesticados podría insertarse en este contexto. De todas formas la domesticación no implica la consolidación de una estrategia pastoril, el cual sería un proceso posterior, que requeriría una reorganización social en unidades domésticas más pequeñas (López 2007). Se desconoce cómo habría sido esta reestructuración social pero se considera que fue necesaria para la consolidación de un nuevo nicho económico.
En este contexto también aumenta el uso de materia prima local, lo cual fue interpretado como una evidencia de reducción de la movilidad residencial en la Puna de Catamarca (Hocsman 2002; Pintar 1995). La reducción de la movilidad habría sido necesaria para el proceso de domesticación de camélidos, por mecanismos como el protective herding (Yacobaccio 2001b). Por el momento, la evidencia de estructuras arquitectónicas agrupadas antes del 4000 AP en el Norte de Chile en sitios como Tulán 52 y Puripica 1 (Nuñez 1992) no se repite en la Puna Argentina, siendo un indicador relevante para el estudio de cambios en la movilidad residencial.
Posteriormente, durante el Holoceno tardío se habría consolidado un nicho económico pastoril, de lo cual no solamente existe evidencia osteométrica que permite señalar la presencia de camélidos domesticados sino también indicadores contextuales como corrales arqueológicos. La introducción de la cerámica en el área se habría producido en este marco. También existen evidencias de un aumento de las interacciones sociales interpretadas aquí como una forma de minimizar el riesgo en un contexto de reducción de la movilidad residencial, tal como se desprende de la proliferación de estructuras arquitectónicas.
Finalmente, se debe remarcar que la caza siempre jugó un rol fundamental en el nicho económico de los grupos humanos de Pastos Grandes, tal como lo muestra la evidencia osteométrica inclusive en un contexto de consolidación del pastoreo como estrategia económica predominante. A su vez, la alta representación de puntas de proyectil que tienden a una alta inversión de energía avala esta hipótesis. En este marco, la combinación de estas dos estrategias permitiría minimizar el riesgo consumiendo recursos de alto retorno.
Agradecimientos
Agradezco a todas las personas que me ayudaron en la realización de este trabajo, las cuales fueron mencionadas en la tesis de doctorado. Esta investigación fue posible por una beca de doctorado de CONICET.
REFERENCIAS CITADAS
1. Abeledo, S. 2006 Sistemas de cooperación entre pastores de Santa Rosa de los Pastos Grandes: modelos evolutivos de estrategias de cooperación, transmisión cultural y mecanismos de penalización. Trabajo presentado en las VII Jornadas de Jóvenes Investigadores en Ciencias Antropológicas. Instituto Nacional de Antropología y Pensamiento Latinoamericano, Buenos Aires. MS. [ Links ]
2. Aschero, C. y H. Yacobaccio 1999 20 años después: Inca Cueva reinterpretado. Cuadernos del Instituto Nacional de Antropología y Pensamiento Latinoamericano 18: 7-18. [ Links ]
3. Babot, M.2006 El papel de la molienda en la transición hacia la producción pastoril: un análisis desde la Puna meridional Argentina. Estudios Atacameños 32: 75-91. [ Links ]
4. Belardi, J.1992 De lo espacial a lo temporal: Explorando distribuciones de artefactos. Revista de Estudios Regionales 10: 35-67. [ Links ]
5. Binford, L. R.1981 Bones: Ancient Men and Modern Myths. Academic Press, Nueva York. [ Links ]
6. Boone, J. y E. Smith 1998 Is it evolution yet? A critique of evolutionary archaeology. Current Anthropology 39: 141-173. [ Links ]
7. Borrero, L., N. Franco, J. Lanata y J. Belardi 1992 Distribuciones arqueológicas y tafonómicas en la margen Norte del Lago Argentino (Santa Cruz, Argentina). Actas del XII Congreso Nacional de Arqueología Chilena, pp. 23-31. Boletín del Museo Regional de la Araucanía 4, Temuco. [ Links ]
8. Bousman, B.1993 Hunter gatherer adaptations, economic risk and tool design. Lithic Technology 18: 59- 86. [ Links ]
9. Boyd, R. y P. Richerson 1985 Culture and the Evolutionary Process. University of Chicago Press, Chicago. [ Links ]
10. Bronk Ramsey, C. 2005 OxCal progam v3.10. Página web http://www.rlaha.ox.ac.uk/O/oxcal.php. (22 de Mayo de 2007). [ Links ]
11. Broughton, J.1999 Resource depression and Intensification During the Late Holocene, San Francisco Bay. University of California Press, Berkeley, Los Ángeles. [ Links ]
12. Cartajena, I., L. Nuñez y M. Grosjean2003 Los camélidos en la vertiente occidental de la Puna de Atacama: una visión desde el arcaico temprano al formativo temprano. Trabajo presentado al III Taller de Zooarqueología de Camélidos. Grupo de Zooarqueología de Camélidos (ICAZ), Tilcara. MS. [ Links ]
13. Cerri, D.1993 [1903] El Territorio de los Andes, Reseña geográfica descriptiva por su primer gobernador. Universidad Nacional de Jujuy, San Salvador de Jujuy. [ Links ]
14. Chang, C. y H. Koster1986 Beyond bones: Toward and archaeology of pastoralism. Advances in Archaeological Method and Theory 9: 97-147. [ Links ]
15. Ebert, J.1992 Distributional Archaeology. University of New Mexico Press, Albuquerque. [ Links ]
16. Elkin, D.1996 Arqueozoología de Quebrada Seca 3: Indicadores de Subsistencia Humana Temprana en la Puna Meridional Argentina. Tesis doctoral inédita. Facultad de Filosofía y Letras, Universidad de Buenos Aires, Buenos Aires. [ Links ]
17. Elkin, D., C. M. Madero, G. L. Mengoni Goñalons, D. Olivera y H. D. Yacobaccio 1991 Avances en el estudio arqueológico de los camélidos en el noroeste argentino. Trabajo presentado en la VII Convención Internacional de Especialistas en Camélidos Sudamericanos. Jujuy. MS. [ Links ]
18. Escola, P.1996 Riesgo e incertidumbre en economías agropastoriles: Consideraciones teórico-metodológicas. Arqueología 6: 9-23. [ Links ]
19. Escola, P. 2002 Caza y pastoralismo: un reaseguro para la subsistencia. Relaciones de la Sociedad Argentina de Antropología XXVII: 233-245. [ Links ]
20. Escola, P. 2004 La expeditividad y el registro arqueológico. Chungara, Revista de Antropología Chilena vol. 36: 49-60. [ Links ]
21. Fernández Distel, A.1978 Nuevos hallazgos precerámicos en la región de Salinas Grandes, Puna de Jujuy Argentina. Revista del Instituto de Antropología VI: 15-62. [ Links ]
22. Hocsman, S. 2002 ¿Cazadores-recolectores complejos en la Puna meridional Argentina? Entrelazando evidencias del registro arqueológico de la microregión de Antofagasta de la Sierra Catamarca. Relaciones de la Sociedad Argentina de Antropología XXVII: 193-214. [ Links ]
23. Izeta, A. y M. C. Scattolin 2006 Zooarqueología de una unidad doméstica Formativa: El caso de Loma Alta (Catamarca, Argentina). Intersecciones en Antropología 7: 193-207. [ Links ]
24. López, G. 2003 Pastoreo y caza en el Temprano de la Puna de Salta: Datos osteométricos del sitio Matancillas 2. Intersecciones en Antropología 4: 17-27. [ Links ]
25. López, G. 2007 Aspectos sociales de la transición al pastoralismo en la Puna: una perspectiva evolutiva. En Producción y circulación prehispánicas de bienes en el sur andino, compilado por A. Nielsen, M. Rivolta, V. Seldes, M. Vázquez y P. Mercolli, pp. 83-103. Editorial Brujas, Córdoba. [ Links ]
26. López, G. 2008 Arqueología de Cazadores y Pastores en Tierras Altas: Ocupaciones humanas a lo largo del Holoceno en Pastos Grandes, Puna de Salta, Argentina. BAR S1854, South American Archaeology Series 4. John and Erica Hedges, Oxford. [ Links ]
27. López, G. 2009 The study of the archaeological record of Santa Rosa de los Pastos Grandes, Puna of Salta, Argentina, from an inclusive evolutionary perspective. En Theoretical and Methodological Issues in Evolutionary Archaeology. Toward an Unified Darwinian Paradigm , editado por H. Muscio y G. López, pp. 49-55. BAR International Series 1915, Archaeopress, Oxford. [ Links ]
28. López, G. y M. Medina 2001 Optimización y riesgo desde las arqueofaunas: su aplicación en el análisis de adaptaciones con estrategias de producción de alimentos de la Puna de Salta y del Sector central de Sierras Centrales. Trabajo presentado en el XIV Congreso Nacional de Arqueología Argentina. Universidad Nacional de Rosario, Rosario. MS. [ Links ]
29. López, G. y P. Miranda 2008 El "muerto" del salar: descripción de un hallazgo bioarqueológico a cielo abierto datado en ca. 3700 AP en el borde del salar de Pastos Grandes, Puna de Salta. Arqueología 14: 199-215. [ Links ]
30. Lyman, R.1994 Vertebrate Taphonomy. Cambridge University Press, Cambridge. [ Links ]
31. Meadow, R.1987 Techniques for comparing bone measurement data from small samples. Trabajo presentado a la Northeastern Faunal Analysis Conference. University of Connecticut, Connecticut. MS. [ Links ]
32. Mengoni Goñalons, G.1999 Cazadores de guanacos de la estepa patagónica. Sociedad Argentina de Antropología, Buenos Aires. [ Links ]
33. Mengoni Goñalons, G. y H. Yacobaccio 2006 The Domestication of South American Camelids. A View from the South-Central Andes. En Documenting domestication, New genetics and archaeological paradigms, editado por M. Zeder, D. Bradley, E. Emshwiller y B Smith, pp. 228-243. University of California Press, Berkeley. [ Links ]
34. Muscio, H. 2004 Dinámica Poblacional y Evolución Durante el Período Agroalfarero Temprano en el Valle de San Antonio de los Cobres, Puna de Salta, Argentina. Tesis doctoral inédita. Facultad de Filosofía y Letras, Universidad de Buenos Aires, Buenos Aires. [ Links ]
35. Nelson, M.1991 The study of technological organization. Journal of Archaeological Method and Theory 3: 57-100. [ Links ]
36. Nuñez, L.1981 Asentamientos de cazadores tardíos en la Puna de Atacama: hacia el sedentarismo. Chungara 8: 137-168. [ Links ]
37. Nuñez, L. 1992 Ocupación Arcaica en la Puna de Atacama: secuencia, movilidad y cambio. Prehistoria Sudamericana. Nuevas perspectivas, editado por B. Meggers, pp. 283-307. Washington, Taraxacum. [ Links ]
38. Nuñez, L.1994 Cruzando la Cordillera por el Norte: Señoríos, caravanas y alianzas. En La Cordillera de los Andes: Ruta de encuentros, pp. 9-19. Editorial Fundación Museo Chileno de Arte Precolombino, Santiago de Chile. [ Links ]
39. Nuñez, L. y T. Dillehay 1979 Movilidad giratoria: Armonía social y desarrollo en los Andes Meridionales. Universidad del Norte, Antofagasta. [ Links ]
40. Nuñez, L. y M. Grosjean 1994 Cambios ambientales pleistoceno-holocénicos: Ocupación humana y uso de recursos en la Puna de Atacama (Norte de Chile). Estudios Atacameños 11: 11-24. [ Links ]
41. Olivera, D.1997 La importancia del recurso Camelidae en la Puna de Atacama entre los 10.000 y los 500 años AP. Estudios Atacameños 14: 29-41. [ Links ]
42. Olivera, D. y D. Elkin 1994 De cazadores y Pastores: El proceso de domesticación en la Puna Meridional Argentina. Zooarqueología de Camélidos 1: 95-124. [ Links ]
43. Pianka, E.1982 Ecología evolutiva. Editorial Omega, Barcelona. [ Links ]
44. Pintar, E.1995 Los conjuntos líticos de los cazadores holocénicos en la Puna Salada. Arqueología 5: 9-23. [ Links ]
45. Ratto, N.2003 Estrategias de caza y propiedades del registro arqueológico en la Puna de Chaschuil (Dpto. de Tinogasta, Catamarca, Argentina). Tesis doctoral inédita. Facultad de Filosofía y Letras, Universidad de Buenos Aires, Buenos Aires. [ Links ]
46. Reimer, P. J., M. G. L Baillie, E. Bard, A. Bayliss, J. W. Beck, C. Bertrand, P. G. Blackwell, C. E. Buck, G. Burr, K. B. Cutler, P. E. Damon, R. L. Edwards, R. G. Fairbanks, M. Friedrich, T. P. Guilderson, A. G. Hogg, K. A. Hughen, B. Kromer, F. G. McCormac, S. Manning, C. Bronk Ramsey, R. W. Reimer, S. Remmele, J. R. Southon, M. Stuiver, S. Talamo, F. W. Taylor, J. van der Plicht, and C. E. Weyhenmeyer 2004 IntCal04 terrestrial radiocarbon age calibration, 0-26 cal Kyr BP. Radiocarbon 46: 1029-1058. [ Links ]
47. Restifo, F. 2007 Análisis de un conjunto lítico del Holoceno Temprano en el Alero Cuevas, Pastos Grandes, Puna de Salta. Actas del XVI Congreso Nacional de Arqueología Argentina 3, pp. 207-213- Universidad Nacional de Jujuy, San Salvador de Jujuy. [ Links ]
48. Smith, E. A.1992 Human Behavioral Ecology I. Evolutionary Anthropology 1(1): 20-25. [ Links ]
49. Winterhalder, B., F. Lu y B. Tucker1999 Risk-Sensitive Adaptive Tactics: Models and Evidence from Subsistence Studies in Biology and Anthropology. Journal of Archaeological Research 7(4): 301-348. [ Links ]
50. Yacobaccio, H. 2001a Cazadores complejos y domesticación de camélidos. En El Uso de los Camélidos A Través del Tiempo, editado por G. Mengoni Goñalons, D. Olivera y H. Yacobaccio, pp. 261-281. Ediciones del Tridente, Buenos Aires. [ Links ]
51. Yacobaccio, H. 2001b La domesticación de camélidos en el Noroeste Argentino. En Historia Argentina Prehispánica, editado por E. E. Berberián y A. E. Nielsen, Tomo 1, pp 7-40. Editorial Brujas, Córdoba. [ Links ]
52. Yacobaccio, H., D. Elkin, y D. Olivera 1994 ¿El fin de los sociedades cazadoras? El proceso de domesticación animal en los Andes centro-sur. En Arqueología de cazadores-recolectores. Límites, casos y aperturas, compilado por J. Lanata y L. A. Borrero, pp. 23-32. Arqueología Contemporánea 5, edición especial, Buenos Aires. [ Links ]
53. Yacobaccio, H. y B. Vilá 2002 Condiciones, mecanismos y consecuencias de la domesticación de los camélidos. Estudios Sociales del NOA 5(5): 4-27. [ Links ]
54. Yacobaccio, H., C. Madero, M. Malmierca y M. Reigadas 1997-1998 Caza, domesticación y pastoreo de camélidos en la Puna Argentina. Relaciones de la Sociedad Argentina de Antropología XXII-XXIII: 389-428. [ Links ]

