










Servicios Personalizados
Revista

Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkIntersecciones en antropología
versión On-line ISSN 1850-373X
Intersecciones antropol. vol.11 no.1 Olavarría ene./jun. 2010
ARTÍCULO
Consumo de roedores en el sitio Villa la Punta, agro-alfarero temprano de la región Chaco-Santiagueña
Luis Manuel del Papa, Luciano José María De Santis y José Togo
Luis M. del Papa. CONICET, Cátedra de Anatomía Comparada, Facultad de Ciencias Naturales y Museo, Universidad Nacional de La PLata. Calle 64 entre diag. 113 y 120 s/n, La Plata, Buenos Aires, Argentina. E-mail: loesdelpapa@hotmail.com
Luciano J. M. De Santis. Cátedra de Anatomía Comparada, Facultad de Ciencias Naturales y Museo, Universidad Nacional de La PLata. Calle 64 entre diag. 113 y 120 s/n, La Plata, Buenos Aires, Argentina. E-mail: desantis@fcnym.unlp.edu.ar
José Togo. Facultad de Humanidades, Ciencias Sociales y de la Salud (UNSE). Avenida Belgrano (s) Nº 2180, Santiago del Estero, Argentina. E-mail: togofami@arnet.com.ar
Recibido 3 de Junio 2009.
Aceptado 7 de Julio 2009
RESUMEN
En este trabajo se analizan los restos óseos de roedores caviomorfos provenientes del Sitio Villa la Punta, departamento de Choya, ubicado al sudoeste de la provincia de Santiago del Estero. A partir del análisis de las modificaciones óseas, representación de partes anatómicas y el rango etario, se presentan las evidencias de consumo y manejo de los roedores caviomorfos por las poblaciones que se asentaron en dicho lugar en la etapa agroalfarera temprana, perteneciente a los grupos portadores de la cerámica denominada Las Mercedes. A partir de los resultados obtenidos se propone el consumo antrópico de roedores Dolichotinae y el ingreso al sitio por cuestiones naturales de los especimenes de Ctenomys sp. Los datos no permitieron hacer una distinción entre los agentes acumuladores de los restos de Caviinae. Dichos resultados constituyen las primeras evidencias presentadas del consumo de Dolichotinae, y un aporte fundamental para la zona de estudio.
Palabras clave: Dolichotinae; Caviinae; Ctenomys; Agroalfarero temprano; Evidencias antrópicas.
ABSTRACT
Human Consumption Of Rodents At Villa La Punta Site During The Early Agricultural-Potter Period Of The Chaco-Santiagueña Region. Caviomorph bone remains from the Villa La Punta site, Choya Department, southwest Santiago del Estero Province are analyzed in the current study. From the analysis of bone modification, represented body parts and age range, we present evidence of caviomorph rodent consumption and use by the populations that settled at the site in the early stages of an agricultural-potter economy, corresponding to the Las Mercedes pottery-bearing group. We propose that the Dolichotinae rodent remains are the result of human consumption while the Ctenomys sp. specimens entered the site due to the action of natural agents. The data did not allow a distinction among the agents that accumulated Caviinae remains to be made. Such results are the first evidence of Dolichotinae consumption and a fundamental contribution to the zone under study.
Keywords: Dolichotinae; Caviinae; Ctenomys; Early agricultural-potter; Antropic evidence.
INTRODUCCIÓN
En este trabajo se realiza el análisis de los restos óseos de roedores caviomorfos provenientes del sitio Villa la Punta, departamento de Choya, al sudoeste de la provincia de Santiago del Estero, en el extremo sur de las estribaciones de la serranía de Guasayán.Los materiales fueron rescatados por el grupo de investigación en el sitio denominado Villa La Punta "Guayacán" (Togo 2004), donde se comprobó la presencia de escasos fragmentos de alfarería en superficie, pertenecientes mayoritariamente al tipo cerámico Averías; paralelamente en uno de los cortes producidos por las lluvias se han recolectado fragmentos cerámicos Las Mercedes y restos faunísticos.
El recorte analítico del estudio de los roedores caviomorfos se debe a que los mismos representan los taxones más abundantes en la muestra y su interpretación como resultado del consumo humano es generalmente difícil (Bond et al. 1981; Stahl 1996). Identificar aquellos restos que ingresaron por acción antrópica o por agentes naturales resulta necesario para poder inferir el rol que tuvieron dichos taxones en la subsistencia humana.
Las investigaciones previas referentes a las evidencias del uso antrópico de roedores caviomorfos de la Argentina, se centran en los Caviinae (Acosta y Pafundi 2005; Brunazzo 1999; Quintana 2005; Quintana y Mazanti 2001; Quintana et al. 2002; Santiago 2004; Valverde 2001), Myocastor coypus (Brunazzo 1999; Ecosteguy 2007; Ecosteguy y Lanza 2007; González de Bonaveri 1997; Quintana y Mazanti 2001; Santiago 2004; Santini 2008), Lagostomus maximus (Quintana y Mazanti 2001; Valverde 2001), e Hydrochaeris hydrochaeris (Feuillet Terzaghi 2002; Santiago 2004).
Para Dolichotis patagonum (mara o liebre patagónica) no se presentan evidencias del consumo humano en los sitios arqueológicos en que fueron recuperados, aunque es planteada dicha actividad para el sitio Fortín Necochea (Crivelli et al. 1997). Dolichotis salinicola (conejo de los palos) está representado en Cerro El Dique de la provincia de Salta, pero en proporción escasa (dos por ciento del total) (Raffino et al. 1977) y para cuyos restos no se establecen las evidencias de consumo.
Para la región Chaco-santiagueña, si bien algunos autores hacen referencia al hallazgo en sitios arqueológicos de restos de D. patagonum, D. salinicola, Microcavia australis, Galea musteloides y Ctenomys sp. entre otros (e.g., Cione et al. 1979; Kraglevich y Rusconi 1931; Lorandi y Lovera 1972), en dichos trabajos no se realiza una distinción clara sobre si los mismos fueron producto del consumo humano; en algunos casos se plantea de modo general que los mamíferos recuperados del registro fueron consumidos, evidenciado por la fragmentación de los huesos producto del aprovechamiento de la médula ósea o para la confección de instrumentos (Cione et al. 1979).
Con respecto a los antecedentes que dan cuenta de la acumulación antrópica de roedores caviomorfos, en los distintos trabajos se emplearon diversos indicadores sobre la base de las posibilidades brindadas por los materiales de estudio, del registro arqueológico y de los controles tafonómicos. Entre ellos podemos mencionar las evidencias dejadas por los filos de instrumentos líticos (Ecosteguy 2007; Quintana 2005; Quintana y Mazanti 2001; Quintana et al. 2002; Santiago 2004; Santini 2008; Valverde 2001); las mismas representarían el tipo de evidencias de mayor "fortaleza" para las interpretaciones junto con la identificación de fracturas intencionales y la producción de instrumentos (Mazzanti y Valverde 2001; Santini 2008). Evidencias de menor poder inferencial, debido a la ambigüedad que se genera por procesos que alteran la distribución, la asociación y la representación de los restos, serían el quemado de los huesos (Acosta y Pafundi 2005; Brunazzo 1999; Santiago 2004), abundancia taxonómica y selección de partes esqueletarias (Acosta y Pafundi 2005; González de Bonaveri 1997; Santini 2008), selección de tamaños y edades de los individuos (Acosta y Pafundi 2005; Brunazzo 1999; Santini 2008).
En este trabajo se dan a conocer las evidencias de consumo y manejo de los roedores de la subfamilia Dolichotinae a partir de múltiples evidencias. Esto nos puede ayudar a interpretar ciertos aspectos de la vida de los pobladores del lugar durante la etapa agroalfarera temprana, portadores de la cerámica denominada Las Mercedes.
Sitio Villa la Punta: caracterización del área de estudio
Villa la Punta se encuentra en el departamento de Choya, al sudoeste de la provincia de Santiago del Estero y a 93 km de la capital de dicha provincia. La población se encuentra en el extremo sur de las estribaciones de la serranía de Guasayán (Figura 1). Dichas sierras raramente exceden los 300 m de altura sobre el nivel del mar (Ledesma 1961).
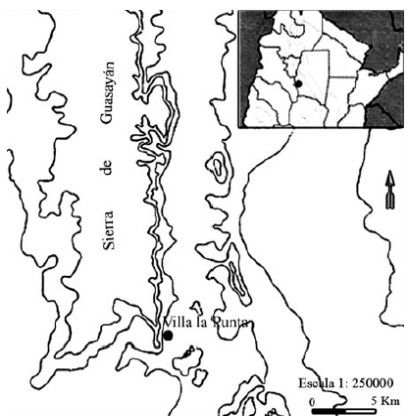
Figura 1. Ubicación del sitio Villa la Punta.
Fitogeográficamente pertenece al Dominio Chaqueño, Distrito Occidental; éste es un bioma en mosaico caracterizado por la presencia dominante de una cobertura arbórea (Aspidosperma quebracho-blanco, Schinopsis lorentzii, Ziziphus mistol, Caesalpinia paraguariensis, Prosopis sp., etc.), salpicado o acompañado de pastizales y otras hierbas no graminoideas (Cabrera 1971). En cuanto a la fauna, su distribución estaría encuadrada en la Subregión Andino-Patagónica, Dominio Central o subandino, Distrito Subandino (Ringuelet 1961).
Se integra por su localización en latitud y longitud, al espacio subtropical semiárido y continental. Las temperaturas promedio para la zona de estudio tienen valores de 20º C, con precipitaciones (se concentran en el período estival) de 450 mm anuales y capacidad de evapotranspiración potencial mayor, lo que determina una gran deficiencia hídrica (Ledesma 1979).
El asentamiento está ubicado al este de las estribaciones de la serranía, sitio a cielo abierto con pendientes suaves y múltiples cañadones, algunos de ellos con fuentes de agua permanente, lo que permitió la subsistencia a los grupos humanos desde época relativamente temprana (Togo 2004). La extensión del sitio se desconoce por no presentar materiales en superficie ni elevaciones artificiales o paredes de piedra; la evidencia se encuentra en las paredes de algunas cárcavas y cañadones.
Los materiales cerámicos son de procedencia "Las Mercedes"; cuyas características representativas son la cerámica de coloración gris-negra, lisa o decorada, en este último caso puede ser en positivo o en negativo (Togo 2004). Además de la cerámica cocida en atmósfera reductora, existen también las cocidas en atmósfera oxidante (Togo 2004). Junto a la cerámica típicamente de Las Mercedes aparece un conjunto pintado designado con el nombre de "Cortaderas"; esta cerámica tiene grandes similitudes, tanto formal como decorativamente, al estilo Alumbrera Tricolor como modificación del Condorhuasi Policromo de la zona de Ambato, provincia de Catamarca (Gómez 1966; Serrano 1958; Togo 2004). En el sitio de estudio se presentan variantes con respecto a otros, ya que se encuentra poco material grabado y pintado (Togo 2004).
En cuanto a la cronología de las Mercedes, en este último tiempo se han realizado varios fechados absolutos (Togo 2007) lo que nos permite compararla con otras entidades que poseen características similares y de edad conocida, por esta razón se considera que pueden corresponder al Período Temprano y con seguridad al Período Medio de la región Valliserrana (Togo 2007). Las Mercedes tendría matrices provenientes tanto de las culturas como Tafi-Candelaria por una parte y Cienaga-Alamito-Condorhuasi por la otra, o raíces originarias comunes para todos ellos (Togo 2007). En la actualidad, estamos analizando la posibilidad de que podría tratarse de una adaptación local de Candelaria. Por los fechados que se disponen, esta cultura se habría desarrollado entre el 350 y el 1100/1200 cal. DC.
La cronología del sitio corresponde a los fechados más antiguos para esta entidad cultural en la provincia, con edades radiocarbónicas de 1550 ± 60 AP (LP-1443; carbón vegetal) y 1580 ± 60 AP (LP-1438; carbón vegetal) (Togo 2007). La diferencia entre los fechados hace presuponer que la ocupación no fue prolongada, ya que los materiales analizados son muy homogéneos desde la base hasta los niveles superiores (Togo 2004).
Además de los restos de alfarería y arqueofaunísticos se rescataron desechos de talla, cuentas de collar y una placa de bronce.
MATERIALES Y MÉTODOS
Se escogió para la excavación la zona que se encontraba entre dos cañadones, por ser la zona libre con mayores posibilidades de obtención de materiales, en que se procedió a la apertura de dos cuadrículas, la Nº I de 2 x 1,66 m de lados (dimensión establecida por la disponibilidad entre ambos cañadones) y la N° II de 2 x 2 m de lado. Los materiales fueron recuperados de niveles artificiales de 20 cm; y se utilizó zaranda con un tamaño de malla de 5 mm (Togo 2004). Se determinó que los materiales se encontraban concentrados en un estrato promedio de 40 cm, entre los 20 y los 60 cm de profundidad, con abundantes restos de huesos quemados, carbones y fragmentos de cerámica (por lo menos en una de las cuadrículas) (Togo 2004).
Se analizaron 2612 restos (óseos, dientes, valvas y cáscaras de huevos) de los que se han podido determinar 16 taxones con distintos niveles de identificación (del Papa 2008); la determinación anatómica y taxonómica se realizó mediante material de comparación depositado en las colecciones de los Departamentos Científicos Zoología Vertebrados, Paleontología Vertebrados y Cátedra de Anatomía Comparada de la Facultad de Ciencias Naturales y Museo de La Plata. Se cuantificaron los materiales teniendo en cuenta las siguientes medidas de abundancia taxonómica y de partes esqueletarias: NISP, MNI, MNE y MAU% (Mengoni Goñalons 1999)
En la muestra faunística general y de los roedores en particular, se analizaron los aspectos tafonómicos tales como los estadios de meteorización (Beherensmeyer 1978); el estado de fragmentación de la muestra a través del porcentaje de elementos completos para cada nivel (Lyman 1994); como así también, la presencia de modificaciones por parte de depredadores como los carnívoros (e.g., Binford 1981; Blumenschine et al. 1996; Mondini 1995; Montalvo et al. 2007) y las aves Strigiformes (Andrews 1990; Fernández Jalvo y Andrews 1992); las producidas por roedores, ya sea, por la construcción de cuevas como por el roído de los huesos (Boceck 1986; Politis y Madrid 1988), y las ocasionadas por la acción de raíces (Lyman 1994).
Una de las vías de análisis para la interpretación de los restos es a través del estudio de la representación de las distintas partes del esqueleto, lo que nos puede informar si hubo una selección antrópica y descarte de las partes no utilizadas; así como el posible transporte en estado completo del esqueleto al sitio. Por otra parte, se estimó el perfil etario de la sub-muestra con el fin de inferir si la misma se corresponde con una preferencia en cuanto a la elección de las presas y la probable técnica de caza (ver Acosta y Pafundi 2005; Jones 2006). Para el caso de estudio, se calculó el porcentaje de especímenes fusionados y sin fusionar para la epífisis proximal del húmero y la distal del fémur (Bellocq y Kravetz 1983; Pearson et al. 1968), considerando de edad adulto a aquellos especimenes que completaron la fusión y juveniles a aquellos que no presentan dicha característica.
Se identificaron las modificaciones de la superficie ósea por factores antrópicos a nivel macroscópico, con lupa de mano de 15X y en los casos en que fue necesario se utilizó lupa binocular. Dichas modificaciones nos pueden informar del procesamiento de las presas, así como de su consumo (no sólo como alimento, sino también como materia prima); entre ellas se consideraron las marcas de corte (Blumenschine et al. 1996; Mengoni Goñalons 1999; Shipman 1981a), infiriendo la probable actividad que las produjo siguiendo los criterios de Binford (1981); atributos asociados a la fractura intencional (Blumenschine y Selvaggio 1988; Bunn 1989; Pickering y Egeland 2006) y el quemado de los huesos (Shipman et al. 1984; Stiner et al. 1995).
Es importante destacar que la acción térmica no suele representar el producto de la cocción de los alimentos, sino que también se puede dar por la acción de descartar "basura" por cuestiones de higiene e impedir la acción de carroñeros, o como combustible, e incluso pueden estar producidos por fuegos naturales (Cain 2005; Costamagno et al. 2005; De Nigris 2004; Kent 1993). Para poder distinguir entre estas acciones, se tuvieron en cuenta una serie de atributos como el color, textura y uniformidad entre otros.
RESULTADOS
Se ha podido determinar una especie de Aves (Rhea americana) y seis de Mammalia (Tolypeutes matacus, Chaetophractus vellerosus, Microcavia australis, Galea musteloides, Dolichotis patagonum y Dolichotis salinicola), y en los demás casos se han asignado a la categoría de género Spixia, Bulimulus, Quelonoidis, Tupinambis, Lama, Dolichotis y Ctenomys, Subfamilia Caviinae, Familia Muridae, Suborden Caviomorpha, Orden Artiodactyla y Rodentia, y Clase Gasteropoda, Aves y Mammalia (Tabla 1).
Tabla 1. Abundancia taxonómica

De los especímenes identificados, se observa que los roedores caviomorfos son los más abundantes (Figura 2a). En este trabajo se realiza el análisis de esta categoría, representada por Ctenomys sp., G. musteloides, M. australis, D. patagonum y D. salinicola, donde la predominancia de roedores Dolichotiinae es evidente (Figura 2b); durante el desarrollo de este trabajo se hará referencia a dicho recorte analítico como sub-muestra de estudio (recorte de la muestra general).
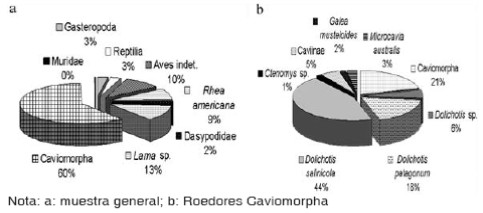
Figura 2. Porcentaje de abundancia de los taxones (NISP%).
Tafonomía
El estado de conservación de la muestra general es bueno; tomando como punto de comparación los estadios de meteorización (Beherensmeyer 1978), se observa que sólo en el nivel 2 de la cuadrícula I (VP I 2) se presenta una alta proporción (29,83%) de especímenes con un estadio medio (valor 3), siendo en el resto de los niveles de ambas cuadrículas, escaso para estos valores y nulo para mayores a éstos.
Se analizó el estado de fragmentación de la muestra, para ello se calculó el porcentaje de elementos completos para cada nivel (Lyman 1994) (Figura 3)

Figura 3. Porcentaje de elementos completos por nivel.
De esto se observa que los niveles mas afectados por la fragmentación son los inferiores (I 4 y II 2, II 3), tal vez debido a factores diagenéticos, evidenciado por la gran cantidad de especímenes que remontan entre sí (Lyman 1994), lo que nos puede indicar que el enterramiento de los materiales fue rápido (Shipman 1981b).
Los especímenes de los roedores caviomorfos presentan porcentajes similares de completitud con valores medios, tanto para los Dolichotinae (entre 41,23% y 42,21%) y Caviinae (56%). Los restos de Ctenomys sp. presentan una mayor proporción de elementos completos (85%). La mayoría de los elementos que se encuentran fracturados, presentan bordes angulosos y ásperos; esto nos informa que la mayoría de los mismos se produjeron por procesos postdepositacionales (Andrews 1990), tales como el pisoteo y el peso de los sedimentos. En un apartado posterior se analizan las fracturas de origen antrópico.
Con respecto a la alteración del registro por causas naturales, se evidencia una escasa acción de carnívoros (0,3%) y roedores (0,2%) sobre los restos óseos. No se observan marcas producto de la acción de ácidos gástricos atribuibles a depredadores (Strigiformes, Falconiformes, Carnivora) (e.g., Andrews 1990; Fernández Jalvo y Andrews 1992).
Las marcas de raíces se presentan entre un 7% y 15% en los niveles 1 y 2 y menor a 3% para el tercer nivel. No se descarta la acción de roedores como producto de la construcción de cuevas, que a pesar de no haber sido reconocidas en la excavación, las mismas se observan en las inmediaciones.
Los restos de roedores del género Ctenomys se encontraron en la cuadrícula I nivel 3, donde sólo se logró identificar seis elementos, los cuales se encontraban enteros en su mayoría: entre ellos, el cráneo (bula timpánica, hemimandíbula), escápula, húmero, y pelvis porción izquierda y derecha. En dichos especímenes no se evidencia la acción de corrosión por digestión (Andrews 1990; Fernández Jalvo y Andrews 1992), ni atributos asociados a la acumulación antrópica.
Representación de partes anatómicas
Con respecto a D. patagonum, se observa que la representación de partes anatómicas se caracteriza por la presencia de la mayoría de los elementos del esqueleto, con buena presencia de la columna vertebral (atlas, vértebras lumbares y sacras), los miembros (húmeros, radios, fémures), el cráneo (calculado por los fragmentos de maxilares, temporales y hemimandíbulas), hemimandíbulas y pelvis; siendo escasos aquellos elementos del autopodio (astrágalos, tarsales, carpales, metapodios y falanges) vértebras torácicas y cervicales en general (a excepción del atlas) (Figura 4).
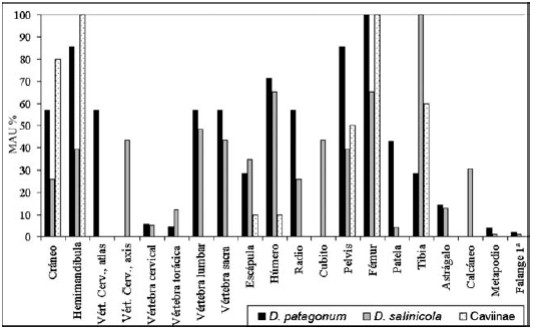
Figura 4. Representación de partes anatómicas (MAU%).
Para D. salinicola, también están presentes la mayor parte de los elementos del esqueleto, principalmente tibias, húmeros y fémures, con escasa representación de las distintas porciones del autopodio, vértebras torácicas y cervicales en general (excepto axis), y con buena representación para el resto del esqueleto, como el cráneo (principalmente timpánicos y hemimandíbulas), escápulas, radios, cúbitos, etc. (Figura 4).
Con respecto a los roedores de la Subfamilia Caviinae, tanto M. australis como G. musteloides están representados principalmente por el cráneo (maxilares y hemimandíbulas). Por la difícil diferenciación entre estas especies por los elementos post-craneanos, el cálculo de la representación de partes se realizó para roedores Caviinae, incluyendo los elementos de estas especies (Figura 4).
Se puede observar que están representados los elementos del cráneo (maxilares, hemimandíbulas) y miembros. La ausencia de algunos elementos se puede deber, tanto a un sesgo en el reconocimiento de los mismos por la fragmentación de los especímenes, como a la problemática de la determinación de partes esqueletarias poco diagnósticas como las vértebras.
Otra posible explicación a la ausencia de algunos elementos, podría ser la destrucción diferencial. También se considera que el quemado (ver apartado de acción térmica) de los huesos pudo generar la destrucción de partes del esqueleto (Lanata 1988); aunque por la escasa representación de estos roedores, no se descarta que el quemado de algunos elementos se deba a la asociación circunstancial por la muerte natural de los individuos y una posterior alteración espacial del registro.
Rango etario
Se analizó el rango etario para D. patagonum, D. salinicola, y Caviinae, sin embargo es sólo estimativo, ya que no se cuenta con un estudio sobre el desarrollo de los individuos de estas especies a nivel óseo. Para el caso de estudio, se calculó el porcentaje de especímenes fusionados y sin fusionar para la epífisis proximal del húmero y la distal del fémur (Bellocq y Kravetz 1983; Pearson et al. 1968), ya que dichos centros de osificación se fusionan de manera más tardía.
La estimación de la edad por la erupción dentaria se vio dificultada debido a que la mayoría de las mandíbulas y maxilares habían perdido los molares en el traslado y manejo de los materiales por parte de los investigadores o por estar fragmentados.
Con respecto a los Dolichotinae, a pesar que los especímenes que representan mejor el desarrollo ontogenético de los roedores son escasos, se observa que prácticamente están en similares proporciones (Tabla 2).
Tabla 2. Porcentaje de especimenes fusionados (F) y sin fusionar (SF).

Para los Caviinae hay que tener en cuenta la escasez de restos que pueden sobredimensionar estos cálculos, resultando en un 88,88% de especímenes juveniles (siete elementos de fémur sin fusionar correspondientes a siete individuos; y uno fusionado); en el caso del húmero, en la muestra sólo se encuentra un caso, el cual representa la edad juvenil.
Marcas de corte
En la sub-muestra de estudio se presentan marcas de corte en dos especímenes de fémur (porción posterior de cóndilo lateral de epífisis distal y cuello de epífisis proximal) de D. patagonum (Figura 5a). Se infiere la actividad de desarticulación para las mismas (Binford 1981), dada la ubicación de las marcas, la disposición transversal al eje del elemento, ser relativamente cortas (menores a cinco milímetros) y relativamente profundas. Si bien el trabajo de Binford hace referencia a mamíferos de mayor porte, lo que hace difícil la comparación con estos taxones (Quintana 2005), creemos que en este caso sería de utilidad para explicar las escasas marcas de corte representadas en la muestra.
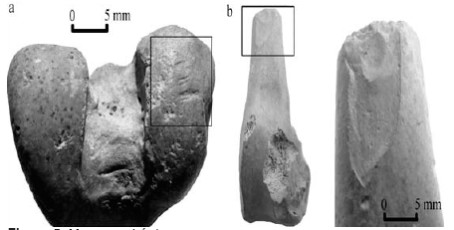
Figura 5. Marcas antrópicas.
Fracturas
Se observa que solo en los especímenes de D. patagonum se produjeron en estado fresco del hueso, algunos con atributos asociados a la fractura. Si bien estos restos son escasos (cuatro con fractura fresca), se procedió a realizar el porcentaje de especímenes fracturados en relación a los huesos largos susceptibles de obtener médula ósea de forma directa, descartando así el cráneo, vértebras, huesos del autopodio, cinturas y cúbito. El valor del índice de huesos fracturados es de 14,28%, de los cuales tres restos presentan atributos asociados a la fractura (negativos de lascado) (Tabla 3).
Tabla 3. Especímenes de D. patagonum con fracturas.
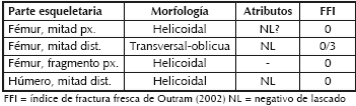
La mitad distal de un fémur presenta negativos de lascado y pequeños retoques en la superficie de fractura (Figura 5b), lo que podría estar representando algún momento de la cadena operativa, previo a la terminación de la confección de instrumentos.
Acción térmica
Se observa en la muestra general, que en los niveles VP I 4 y VP II 3 hay una gran cantidad de especímenes quemados (alrededor del 40%), manteniéndose para el resto de los niveles entre un 10% y un 20%. El porcentaje de todo el conjunto es de un 19,71%.
En la muestra general predominan los fragmentos indeterminados o categorías más abarcativas (como es de esperar por la acción del quemado de los huesos que genera la fragmentación de los mismos) (Lanata 1988; Whyte 2001).
De la sub-muestra en estudio, el 12,37% de los especímenes de D. patagonum y el 16,59% de D. salinicola presentan modificaciones por acción térmica. Por otro lado, los restos de Caviinae están representados por el 3,92% (un espécimen de G. musteloides y uno de M. australis).
Hacer inferencias respecto de la actividad que produjo el quemado de los huesos puede ser más complicado en el estudio de los roedores, ya que, en vez de asar partes de las carcasas (como en los casos de las presas de mayor tamaño), los roedores pudieron ser expuestos a las brasas enteros y con piel (al rescoldo) como se desprende de los estudios etnográficos sobre Tobas y Mocovíes del Chaco Occidental (Arenas 2003); por lo que la observación de algún patrón para la cocción de los alimentos se dificulta (al asarlos con piel, no es de esperar que los huesos presenten algún atributo del proceso de cocción). A su vez, los grupos humanos que habitaron el sitio estaban equipados con la tecnología necesaria para producir la cocción de los alimentos por hervido (restos de alfarería utilitaria).
Del análisis de la muestra general se observa una gran proporción de elementos quemados que presentan características generales de haber sido sometidos a un lapso prolongado de tiempo al fuego, donde predominan los restos con una coloración negra, y negra a blanquecina, que se distribuye en la mayor parte o todo el espécimen; algunos con superficies craqueladas (mayor grado de alteración). También se observan restos fragmentados que presentan una coloración similar tanto en la superficie interna como externa (posible quemado después del procesamiento de los elementos).
De estas características se desprende que la acción del quemado por descarte primó en todo el conjunto; esto se da para los Dolichotinae, donde están representados la mayoría de los elementos del esqueleto, como para los Caviomorpha. No se observa lo mismo en los restos de Caviinae dada la escasa alteración de los elementos por la acción térmica.
DISCUSIÓN Y COMENTARIOS FINALES
La integridad de la muestra es en general buena, con escasa acción de carnívoros y roedores como agentes formadores del registro y modificadores de los materiales que lo componen. En el caso de las cuevas producto de mamíferos cavadores como los roedores o dasipódidos, si bien no se identificaron en la excavación, no se descarta la presencia de las mismas y la probable distorsión del contexto arqueológico.
De los estadios de meteorización (en general baja alteración, excepto en el nivel I, 2 con mayor proporción de valores medios), como la fragmentación por diagénesis (evidenciado por el remontaje de especímenes óseos), nos indicaría que el enterramiento de los materiales fue relativamente rápido (Beherensmeyer 1978; Lyman 1994; Shipman 1981b).
Con respecto a las evidencias del consumo humano de los roedores, consideramos que la representación de partes de los Dolichotinae, se corresponde con una incorporación de los individuos en forma completa. La ausencia de elementos del autopodio podría corresponderse con el procesamiento de la piel de estos animales y el descarte de dichos elementos en la zona de producción o al acarreo de esas partes junto con la piel, ya que dicha acción se realiza por las zonas de menor cantidad de músculos, y desde donde se puede aprovechar mejor el cuero de los mismos (huesos del autopodio, extremo distal de tibia, de radio y cúbito, el cráneo y la mandíbula) (Binford 1981; Quintana 2005); sin embargo no hay evidencias de tal acción (huellas de corte en los elementos mencionados).
La ausencia de la mayoría de las vértebras cervicales y torácicas tendría que estar relacionada con otros factores, tal vez de destrucción diferencial por otros agentes tafonómicos, o de procesamiento.
Del análisis del rango etario y de la eto-ecología de las especies de Dolichotinae, se intenta explicar la posible forma de captura. La alta proporción de individuos juveniles nos podría indicar la obtención de mayor cantidad de individuos por evento de caza (más de 22 parejas suelen depositar sus juveniles en un pozo comunal; Campos et al. 2001; Redford y Eisenberg 1992), sin embargo dado que el número de restos que nos permiten hacer tal diferenciación (epífisis proximal del húmero y distal de fémur) son escasos, y al no existir un estudio detallado sobre el desarrollo etario de los restos óseos de estas especies, dicha inferencia no es conclusiva hasta tanto no se produzca una aproximación mas certera en estas cuestiones.
Dada la presencia de individuos adultos en similar proporción a los juveniles de ésta Subfamilia (los adultos forman parejas monógamas que evitan el contacto con otras; Redford y Eisenberg 1992); es posible que también se de una selección y búsqueda de individuos de tamaño mas grande (mayor cantidad de productos obtenidos por individuo), lo que acrecienta los costos de captura.
Otra manera de obtener éste recurso podría ser el uso de trampas; esto aumentaría la posibilidad de obtener individuos dispersos o de una menor disponibilidad (individuos adultos), con una disminución de los costos de captura y de búsqueda, lo que podría explicar la preponderancia del mismo con relación a aquellos de tamaño corporal más grande y de mayor rendimiento (e.g., Camelidae y Rheidae; Tabla 1).
Sin embargo cuando hablamos de masa corporal o de proteínas susceptibles de ser consumidas, el peso total estimado por el valor máximo para D. patagonum (12 kg; Baldi 2007) y D. salinicola (2,7 kg; Redford y Eisenberg 1992) de los individuos representados, sería de 83 kg; siendo menor al peso de un guanaco adulto de bajas latitudes (alrededor de 95 kg; Elkin 1996), los cuales están representados por tres individuos (pero con menor cantidad de partes esqueletarias).
Se considera que las marcas de corte, producto de la actividad de desarticulación de las presas de D. patagonum, fueron probablemente consecuencia del procesamiento para la elaboración de alimentos; y para dicha actividad fue necesario dividir la presa en unidades más pequeñas, quizás por el uso de recipientes para su cocción (hervido en ollas de alfarería u otro contenedor), o la obtención de materia prima para la confección de instrumentos (Figura 5b), así también por el reparto de bienes entre los integrantes de una familia o grupo de relaciones sociales mas cercano.
En cuanto a las fracturas, si bien el índice de especímenes fracturados no es bajo para D. patagonum (14,28%), hay que tener en cuenta que el canal medular de estos huesos no es demasiado grande como para inferir el consumo sistemático de médula ósea. Aunque no se descarta esto último (principalmente por el hervido de los huesos), también se considera que pueden ser el resultado de la formatización de instrumentos (Figura 5b).
Los roedores Dolichotinae están bien representados con atributos producto de la acción térmica a lo largo de la secuencia. Si bien no se pudo determinar la acción que los produjo por la gran representación de todos los elementos con este atributo, se considera que la acción de descarte en estructuras de combustión primó en todo el conjunto.
En el sitio Villa la Punta se pudo comprobar con mayor nivel de fortaleza en las inferencias realizadas, que los roedores Dolichotinae fueron utilizados por los humanos.
En los restos de D. patagonum se observan marcas de corte y atributos asociados a la fractura intencional, el quemado de los huesos con un buen porcentaje de los mismos, y la representación de partes y el rango etario concuerda con lo esperado en cuanto a una selección de presas y de su traslado en forma entera al lugar de consumo.
Para D. salinicola, se presentan atributos de acción térmica con un porcentaje algo mayor que D. patagonum, donde la asociación secundaria con estructuras de combustión quedaría descartada (tanto por la proporción de especímenes quemados, como por el grado de alteración de los mismos). Al igual que en el caso anterior, tanto la representación de partes y el rango etario representarían una selección por parte de los humanos.
En contraposición, se advierte que los restos de Ctenomys ingresaron al sitio por causas naturales, posiblemente por la muerte de un individuo entrampado en su madriguera (estos roedores están adaptados a la vida fosorial y pasan la mayoría de su vida bajo la superficie del suelo; Redford y Eisenberg 1992), si bien no se hallaron evidencias de cuevas en la cuadrícula; esto se infiere por los escasos restos recuperados (elementos enteros principalmente) en la cuadrícula I, nivel 3, que podrían pertenecer al mismo individuo. También podría representar los restos de presas de Strigiformes, especialmente aquellas que su acción no representa grandes modificaciones de la superficie ósea, ya que no se hallaron evidencias de la acción de corrosión gástrica (e.g., Tyto alba y Asio flammeus) (Andrews 1990; Gómez 2007).
Por último y más complicado, es explicar la presencia de los roedores Caviinae (G. musteloides y M. australis), ya que si bien son más abundantes que los restos recuperados de Ctenomys, la representación de los mismos no sería muy elevada; teniendo en cuenta que por su tamaño y por ser especies de hábitos gregarios diurnos (Redford y Eisenberg 1992; Rood 1972; Tognelli et al. 2001), se esperaría una mayor tasa de captura por evento de caza, y por lo tanto, una mayor cantidad de individuos en el registro.
El perfil etario de los Caviinae tampoco cumple con lo esperado para este taxón suponiendo una acumulación antrópica (se esperaría una mayor selección de individuos adultos, que aportarían mayor cantidad de nutrientes) (Acosta y Pafundi 2005). Este perfil podría ser esperado para una acumulación de aves Strigiformes, las que seleccionan los individuos juveniles de estas especies (selección dada por el tamaño de las presas que consumen) (Acosta y Pafundi 2005; Pardiñas 1999; Quintana et al. 2002). Este taxón, junto a Ctenomys, podría representar la acción de estos depredadores en la muestra. De ser así, estaríamos ante la presencia de un palimpsesto generado por la acción de diversos agentes, tanto antrópicos como de aves depredadoras, posiblemente Tyto alba y Asio flammeus dado que no se observan modificaciones de la superficie ósea (acción de ácidos gástricos).
El único atributo asociado a la acción antrópica para los Caviinae sería el quemado de los huesos, aunque en esta subfamilia los mismos son escasos (dos especímenes) y podría estar más relacionada con una asociación secundaria con estructuras de combustión o al tipo de cocción (al rescoldo). Si bien las evidencias sobre el consumo de éstos, por parte de los grupos humanos que habitaron el sitio de estudio no son conclusivas, no se descarta dicha posibilidad.
La evidencia aportada por otros investigadores sobre el uso antrópico de roedores Caviinae (Acosta y Pafundi 2005; Brunazzo 1999; Quintana 2005; Quintana y Mazanti 2001; Quintana et al. 2002; Santiago 2004; Valverde 2001), si bien no presentan las mismas características que las observadas en este trabajo, nos invita a desarrollar este interrogante para la región de estudio a partir de la adición de nuevas muestras para el análisis en el futuro.
El consumo de recursos de menor rendimiento ha sido propuesto para los primeros momentos de colonización de nuevos ambientes, constituyendo una estrategia ligada a procesos iniciales de adaptación, como es el caso del sitio Piuquenes en la cordillera andina de Chile central (Labarca 2005), donde se evidencia el consumo sistemático de Lagidium viscacia durante el lapso Pleistoceno-Holoceno.
Para el Holoceno Tardío, diversos autores consideran que se desarrolló una estrategia de intensificación de los recursos, donde se produce un aprovechamiento más acentuado de los animales pequeños, de menor rendimiento y una mayor incorporación de especies a la subsistencia, debido a la mayor cantidad de pobladores con menor movilidad para este período (e.g., González de Bonaveri 1997; Martínez y Gutiérrez 2004; Neme 2007; Quintana y Mazzanti 2001; Quintana et al. 2002).
En la región de estudio, la preponderancia de recursos pequeños se puede deber a una intensificación de los mismos por el incremento del sedentarismo, dado por la transición a la producción de alimentos (Neme y Gil 2008), del contexto agroalfarero temprano (grupos con complemento de agricultura; Togo 2005) al que se adscriben los materiales hallados. Esto estaría representando una estrategia de reducción del riesgo dado por la deficiencia hídrica de la zona y sus consecuencias para la predictibilidad de la agricultura. Sin embargo, para una mayor comprensión de lo antes expuesto, faltaría incorporar al estudio, el rol que tuvieron en la subsistencia el resto de las especies presentes en el sitio (e.g., Lama sp., Quelonoidis sp., Tupinambis sp., R. americana, aves indeterminadas, T. matacus y C. vellerosus).
Agradecimientos
A Amelia Barreiro por la traducción del resumen al inglés. A Mariano Santini, Germán Moreira y Carlos Quintana por la lectura de versiones preliminares del manuscrito; y a dos evaluadores anónimos, cuyos comentarios ayudaron a mejorar la producción del mismo. Lo aquí expresado es responsabilidad de los autores. Este trabajo forma parte del proyecto de beca doctoral del CONICET otorgada al primer autor.
REFERENCIAS CITADAS
1. Acosta, A. y L. Pafundi 2005 Zooarqueología y tafonomía de Cavia aperea en el humedal del Paraná inferior. Intersecciones en Antropología 6: 59-74. [ Links ]
2. Andrews, P. 1990 Owls, caves and fossils. University of Chicago Press, Chicago. [ Links ]
3. Arenas, P.2003 Etnografía y Alimentación Entre los Tobas Ñachilamoleek y Wichi-Lhukutas del Chaco Central (Argentina). Pastor Arenas editor, Buenos Aires. [ Links ]
4. Baldi, R.2007 Breeding success of the endemic mara Dolichotis patagonum in relation to habitat selection: Conservation implications. Journal of Arid Environments 68: 9-19. [ Links ]
5. Behrensmeyer, A. F. 1978 Taphonomic and ecologic information from bone weathering. Paleobiology 4: 150-162. [ Links ]
6. Bellocq, M. I. y F. O. Kravetz 1983 Identificación de especies, sexo y edad relativa a partir de restos óseos de roedores de la provincia de Buenos Aires, Argentina. Historia Natural 3 (10): 101-112. [ Links ]
7. Binford, L. R. 1981 Bones: Ancient Men and Modern Myths. Academic Press, Nueva York. [ Links ]
8. Blumenschine, R. J., C. W. Marean y S. D. Capaldo 1996 Blind test of inter-analyst correspondence and accuracy in the identification of cut marks, percussion marks, and carnivore tooth marks on bone surfaces. Journal of Archaeological Science 23: 493-507. [ Links ]
9. Blumenschine, R. J. y M. M. Selvaggio 1988 Percussion marks on bone surfaces as a new diagnostic of hominid behavior. Nature 333: 763-765. [ Links ]
10. Bocek, B. 1986 Rodent ecology and burrowing behavior: Predicted effects on archaeological site formation. American Antiquity 51 (3): 589-603. [ Links ]
11. Bond, M., S. E. Caviglia y L. Borrero 1981 Paleoetnozoología del Alero de los Sauces (Neuquén, Argentina); con especial referencia a la problemática presentada por los roedores en sitios patagónicos. Trabajos de Prehistoria 1: 95-111. [ Links ]
12 Brunazzo, G. A.1999 Investigaciones arqueológicas en el sitio La Norma (Partido de Berisso, Provincia de Buenos Aires, Argentina). Actas del XII Congreso Nacional de Arqueología Argentina, Tomo III: 101-106. La Plata. [ Links ]
13. Bunn, H. T. 1989 Diagnosing Plio-Pleistocene hominid activity with bone fracture evidence. En Bone Modification, editado por R. Bonnichsen y M. H. Sorg, pp. 299-315. Center for the Study of the First Americans, University of Maine, Orono. [ Links ]
14. Cabrera, A. L. 1971 Fitogeografía de la República Argentina. Boletín de la Sociedad Argentina de Botánica XIV (1-2), Buenos Aires. [ Links ]
15. Cain, C. R. 2005 Using Burned animal bone to look at Middle Stone Age occupation and behavior. Journal of Archaeological Science 32: 873-884. [ Links ]
16. Campos, C. M., M. F. Tognelli y R. A. Ojeda 2001 Dolichotis patagonum. Mammalian Species 652: 1-5. [ Links ]
17. Cione, A., A. M. Lorandi y E. P. Tonni 1979 Patrón de Subsistencia y Adaptación Ecológica en la Aldea Prehispánica "El Veinte", Santiago del Estero. Relaciones de la Sociedad Argentina de Antropología XIII: 102-116. [ Links ]
18. Costamagno, S., I. Théry-Parisot, J. P. Brugal y R. Guibert 2005 Taphonomic consequences of the use of bones as fuel: experimental data and archaeological consequences. En Biosphere to Lithosphere. New Studies in Vertebrate Taphonomy, editado por T. O'Connor, pp. 51-62, 9th ICAZ, Durham, 2002. Oxbow Books, Oxford. [ Links ]
19. Crivelli Montero, E. A., E. Eugenio, U. F. J. Pardiñas y M. J. Silveira 1997 Archaeological investigation in the plains of the Province of Buenos Aires, Llanura Interserrana Bonaerense. Quaternary of South America and Antartic Peninsula 10: 167- 209. [ Links ]
20. De Nigris, M. E. 2004 El consumo en grupos cazadores recolectores. Un ejemplo zooarqueológico de Patagonia meridional. Sociedad Argentina de Antropología, Buenos Aires. [ Links ]
21. del Papa, L. M. 2008 Informe final de Beca de Iniciación. Secretaria de Ciencia y Técnica, Universidad Nacional de La Plata, La Plata. MS. [ Links ]
22. Ecosteguy, P. D. 2007 Estudios sobre restos de Myocastor coypus en sitios de la cuenca inferior del río Salado. En Arqueología en las Pampas, editado por C. Bayón, A. Pupio, M. I. González, N. Flegenheimer y M. Frère, Tomo I, pp. 471-487. Sociedad Argentina de Antropología, Buenos Aires. [ Links ]
23. Ecosteguy, P. D. y M. Lanza 2007 Análisis de huellas en restos arqueofaunísticos de Myocastor coypus. Resúmenes ampliados del XVI Congreso Nacional de Arqueología Argentina: 167-171. Jujuy. [ Links ]
24. Elkin, D. C.1996 Arqueozoología de Quebrada Seca 3: Indicadores de Subsistencia Humana Temprana en la Puna Meridional Argentina. Tesis Doctoral inédita. Facultad de Filosofía y Letras, Universidad de Buenos Aires. Buenos Aires. [ Links ]
25. Fernández-Jalvo, Y. y P. Andrews 1992 Small mammal taphonomy of Gran Dolina, Atapuerca (Burgos), Spain. Journal of Archaeological Science 19:407-428. [ Links ]
26. Feuillet Terzaghi, M. R. 2002 El registro arqueofaunístico de vertebrados del sitio Playa Mansa I (Arroyo Seco, Depto. Rosario, provincia de Santa Fe). Aportes y perspectivas. Tesis de Licenciatura inédita. Facultad de Humanidades y Artes, Universidad Nacional de Rosario. Rosario. [ Links ]
27. Gómez, G. N. 2007 Predators categorization based on taphonomic análisis of micromammal bones: a comparison to proposed models. En Taphonomy and Zooarchaeology in Argentina, editado por M. A. Gutiérrez, L. Miotti, G. Barrientos, G. Mengoni Goñalons y M. Salemme, pp. 89- 103. BAR International Series S1601. Archaeopress, Oxford. [ Links ]
28. Gómez, R.1966 La Cultura de las Mercedes. Contribución a su Estudio. Edición privada, Santiago del Estero. [ Links ]
29. González de Bonaveri, M. I.1997 Potsherds, "coypo" teeth, and fish bones: Hunter-gatherer-fishers in the Río Salado (Pampa Region, Argentina). Quaternary of South America and Antartic Peninsula 10: 255-278. [ Links ]
30. Jones, E. L. 2006 Prey choice, mass collecting and the wild European rabbit (Oryctolagus cuniculus). Journal of Anthropological Archaeology 25: 275-289. [ Links ]
31. Kent, S. 1993 Variability in Faunal Assemblages: the Influence of Hunting Skill, Sharing, Dogs and Mode of Cooking on Faunal Remains at a Sedentary Kalahari Community. Journal of Anthropological Archaeology 12: 323-383. [ Links ]
32. Kraglievich, L. y C. Rusconi1931 Restos de vertebrados vivientes y extinguidos hallados por los Señores E. R. Wagner y hermano en túmulos precolombinos de Santiago del Estero. Physis 10: 229-241. [ Links ]
33. Labarca, R. 2005 Las vizcachas de Piuquenes: Evidencia de explotación de roedores durante la transición Pleistoceno-Holoceno en la cordillera andina de Chile central. Revista Werken 6: 63-80. [ Links ]
34. Lanata, J. L. 1988 ¿Huesos quemados, huesos contados? Los datos de un experimento. En De Procesos, Contextos y otros Huesos, editado por N. R. Ratto y A. F. Haber, pp. 75-90, Facultad de Filosofía y Letras, UBA, Buenos Aires. [ Links ]
35. Ledesma, N. R.1979 La verdad sobre el clima de Santiago del Estero. Cuaderno de Cultura 10 (17), Municipalidad de Santiago del Estero, Santiago del Estero. [ Links ]
36. Ledesma, R. 1961 Maquijata. Instituto de Lingüística, Folklore y Arqueología, Facultad de Filosofía y Letras, Universidad Nacional de Tucumán, Santiago del Estero. [ Links ]
37. Lorandi, A. M. y D. M. Lovera1972 Economía y Patrón de Asentamiento en la Provincia de Santiago del Estero. Relaciones de la Sociedad Argentina de Antropología VI: 173-191. [ Links ]
38. Lyman, R. L. 1994 Vertebrate Taphonomy. Cambridge University Press, Cambridge. [ Links ]
39. Martínez, G. y M. A. Gutiérrez 2004 Tendencias en la explotación humana de la fauna durante el Pleistoceno final y Holoceno en la Región Pampeana (Argentina). En Zooarchaeology of South America, editado por G. L. Mengoni Goñalons, pp. 81-98. BAR International Series, Oxford. [ Links ]
40. Mazzanti, D. L. y F. Valverde 2001 Artefactos sobre Hueso, Asta y Valva. En Cueva Tixi: Cazadores y Recolectores de las Sierras de Tandilla Oriental. 1 Geología, Paleontología y Zooarqueología, editado por D. L. Mazzanti y C. A. Quintana, pp. 157-180. Laboratorio de Arqueología. Universidad Nacional de Mar del Plata. Publicación Especial 1, Mar del Plata. [ Links ]
41. Mengoni Goñalons, G. L. 1999 Cazadores de guanacos de la estepa patagónica. Sociedad Argentina de Antropología, Buenos Aires. [ Links ]
42. Mondini, M. 1995 Artiodactyl prey transport by foxes in Puna rock shelters. Current Anthropology 36: 520-524. [ Links ]
43. Montalvo, C. I., M. E. M. Pessino y V. H. González 2007 Taphonomic análisis of remains of mammals eaten by pumas (Puma concolor, Carnivora, Felidae) in central Argentina. Journal of Archaeological Science 34: 2151-2160. [ Links ]
44. Neme, G.2007 Cazadores-recolectores de altura en los Andes meridionales. BAR International Series 1591. Archaeopress, Oxford. [ Links ]
45. Neme, G. A. y A. F. Gil 2008 Faunal Exploitation and Agricultural Transitions in the South American Agricultural Limit. International Journal of Osteoarchaeology 18 (3): 293-306. [ Links ]
46. Outram, A. K. 2002 Bone fracture and within-bone nutrients: an experimentally based method for investigating levels of marrow extraction. En Consuming Passions and Patterns of Consumption, editado por P. Miracle y N. Milner, pp. 51-62. McDonald Institute for Archaeological Research, Cambridge. [ Links ]
47. Pardiñas, U. F. J. 1999 Tafonomía de microvertebrados en yacimientos arqueológicos de Patagonia. Arqueología 9: 265-308. [ Links ]
48. Pearson, O. P., N. Binsztein, L. Boiry, C. Busch, M. Di Pace, G. Gallopin, P. Penchaszadeh y M. Piantanida 1968 Estructura social, distribución espacial y composición de edades de una población de tuco-tucos (Ctenomys talarum). Investigaciones Zoológicas Chilenas 13: 47-80. [ Links ]
49. Pickering, T. R y C. P. Egeland 2006 Experimental patterns of hammerstone percussion damage on bones: implications for inferences of carcass processing by humans. Journal of Archaeological Science 33: 459-469. [ Links ]
50. Politis, G. y P. Madrid 1988 Un hueso duro de roer: Análisis preliminar de la tafonomía del sitio Laguna Tres Reyes (Partido de Adolfo González Chaves. Provincia de Buenos Aires). En De procesos, contextos y otros huesos, editado por N. R. Ratto y A. F. Haber, pp. 29-44, Facultad de Filosofía y Letras, UBA, Buenos Aires. [ Links ]
51. Quintana, C. A. 2005 Despiece de microroedores en el Holoceno Tardío de las Sierras de Tandilla (Argentina). Archaeofauna 217: 227-241. [ Links ]
52. Quintana, C. A. y D. L. Mazzanti 2001 Selección y Aprovechamiento de Recursos Faunísticos. En Cueva Tixi: Cazadores y Recolectores de las Sierras de Tandilla Oriental. 1 Geología, Paleontología y Zooarqueología, editado por D. L. Mazzanti y C. A. Quintana, pp. 181-209. Laboratorio de Arqueología. Universidad Nacional de Mar del Plata. Publicación Especial 1, Mar del Plata. [ Links ]
53. Quintana, C. A., F. Valverde y D. L. Mazzanti 2002 Roedores y Lagartos Como Emergentes De La Diversificación De La Subsistencia Durante El Holoceno Tardío En Sierras De La Región Pampeana Argentina. Latin American Antiquity 13 (4): 455-473. [ Links ]
54. Raffino, R. A., E. P. Tonni y A. L. Cione 1977 Recursos alimentarios y economía en la región de la Quebrada del Toro, provincia de Salta, Argentina. Relaciones de la Sociedad Argentina de Antropología XI: 9-30. [ Links ]
55. Redford, K. J. y J. F. Eisenberg 1992 Mammals of the Neotropics. The southern Cone. Vol. 2. University of Chicago Press, Chicago. [ Links ]
56. Ringuelet, R. A. 1961 Rasgos fundamentales de la zoogeografía de la Argentina. Physis 22: 151-170. [ Links ]
57. Rood, J. P.1972 Ecological and behavioral comparisons of three genera of Argentine cavies. En Animal Behavior Monographs 5, editado por J. M. Cullen y C. C. Beer, pp. 1-83. Newark, Nueva York. [ Links ]
58. Santiago, F. C. 2004 Los roedores en el "menú" de los habitantes de Cerro Aguará (provincia de Santa Fe): su análisis arqueofaunístico. Intersecciones en Antropología 5: 3-18. [ Links ]
59. Santini, M.2008 Aprovechamiento de Myocastor coypus en sitios del Chaco Húmedo. Póster presentado en el I Congreso Nacional de Zooarqueología Argentina. Malargüe, Mendoza. MS. [ Links ]
60. Serrano, A.1958 Manual de la Cerámica Indígena. Edición Assandri, Córdoba. [ Links ]
61. Shipman, P. 1981a Aplications of scanning electron microscopy to taphonomic problems. Annals of the New York Academy of Sciences 276: 357-386. [ Links ]
62. Shipman, P. 1981b Life history of a fossil: an introduction to taphonomy and paleoecology. Harvard University Press, Cambridge. [ Links ]
63. Shipman, P., G. F. Foster y M. Schoeninger 1984 Burnt bones and teeth: an experimental study of colour, morphology, crystal structure and shrinkage. Journal of Archaeological Science 11: 307-325. [ Links ]
64. Stahl, P. W.1996 The recovery and Interpretation of Microvertebrate Bone Assemblages from Archaeological Contexts. Journal of Archaeological Method and Theory 3 (1): 31-75. [ Links ]
65. Stiner, M. C., S. L. Kuhn, S. Weiner y O. Bar-Yosef 1995 Differential burning, recrystalization, and fragmentation of archaeological bone. Journal of Archaeological Science 22: 223-237. [ Links ]
66. Tognelli, M. F., C. M. Campos y R. A. Ojeda 2001 Microcavia australis. Mammalian Species 648: 1-4. [ Links ]
67. Togo, J. 2004 Arqueología Santiagueña: Estado actual del Conocimiento y Evolución de un Sector de la Cuenca del Río Dulce. Tesis Doctoral inédita, Facultad de Ciencias Naturales y Museo, Universidad Nacional de La Plata. La Plata. [ Links ]
68. Togo, J. 2005 Asentamientos humanos y aprovechamiento de los recursos en Santiago del Estero. En Santiago del Estero. Una mirada ambiental, compilado por A. Giannuzzo y M. Ludueña, pp. 91-119. Facultad de Ciencias Forestales, Universidad Nacional de Santiago del Estero, Santiago del Estero. [ Links ]
69. Togo, J. 2007 Las Mercedes: los primeros fechados radiocarbónicos. Indoamerica, Nueva Serie Científica 1 (1): 51-79. [ Links ]
70. Valverde, F.2001 Huellas y Marcas sobre Hueso. En Cueva Tixi: Cazadores y Recolectores de las Sierras de Tandilla Oriental. 1 Geología, Paleontología y Zooarqueología, editado por D. L. Mazzanti y C. A. Quintana, pp. 137-155. Laboratorio de Arqueología. Universidad Nacional de Mar del Plata. Publicación Especial 1, Mar del Plata [ Links ]
71. Whyte, T. R. 2001 Distinguishing Remains of Human Cremations from Burned Animal Bones. Journal of Field Archaeology 8 (3-4): 437-448. [ Links ]

