










Servicios Personalizados
Revista

Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkIntersecciones en antropología
versión On-line ISSN 1850-373X
Intersecciones antropol. vol.12 no.1 Olavarría ene./jul. 2011
ARTÍCULO
Aprovechamiento de Myocastor coypus (Rodentia, Caviomorpha) en sitios del Chaco Húmedo argentino durante el Holoceno tardío
Mariano Santini
Mariano Santini. Consejo Nacional de Investigaciones Científicas y Técnicas (CONICET). Facultad de Ciencias Naturales y Museo. Departamento de Antropología. Museo de La Plata. Paseo del Bosque S/N. La Plata (1900). E-mail: marianosantini@yahoo.com.ar
Recibido 16 de junio 2010.
Aceptado 31 de agosto 2010
RESUMEN
De los mamíferos presentes en los sitios arqueológicos del Chaco Húmedo durante el Holoceno tardío, Myocastor coypus constituyó un recurso importante en la subsistencia de los grupos humanos que habitaron la zona de esteros, cañadas y selvas de ribera. El objetivo de este trabajo es presentar los resultados obtenidos del estudio de los materiales arqueofaunísticos correspondientes a M. coypus provenientes de los sitios El Cachapé Potrero V y Sotelo 1 (departamento Primero de Mayo, provincia del Chaco). Cronológicamente, estos sitios se sitúan entre los 1200 y 1690 años AP. Los materiales analizados ascienden a un NISP de 7868 y 4412 y un MNI de 251 y 259, para Potrero V y Sotelo 1, respectivamente, y representan el 84% y 77,1% del total de especímenes recuperados. Además del análisis cuantitativo se determinaron los perfiles etarios y se analizaron las modificaciones óseas, antrópicas y naturales, que permitieron identificar los agentes tafonómicos que intervinieron en la formación de los conjuntos muestreados. Los estudios mostraron que se realizó un aprovechamiento integral del coypo -al cual se lo utilizó tanto como fuente de alimento como de materia prima para la confección de instrumentos óseos-; y que, probablemente, también se haya aprovechado su cuero.
Palabras clave: Zooarqueología; Chaco Húmedo; Myocastor coypus.
ABSTRACT
Myocastor coypus (Rodentia, Caviomorpha) procurement at sites in the argentinean Chaco Humedo during the late Holecene Of the mammals from archaeological sites in Chaco Province, Myocastor coypus constituted an important subsistence resource for the human groups that inhabited the area of Esteros, Cañadas and Selvas de Ribera in the Late Holocene. The aim of this paper is to present the results of materials studies relating to M. coypus from the archaeological sites of El Cachapé Potrero 5 and Sotelo 1 (Primero de Mayo Department, Chaco Province). Chronologically, these sites are located between 1200 and 1690 years BP. The NISP was 7,868 and 4,412 and the MNI 251 and 259 for Potrero V and Sotelo I, respectively, accounting for 84% and 77.1% of all specimens recovered from the two archaeological sites. In addition to quantitative analysis, age profiles and osseous, anthropogenic, and natural modifications were analyzed, identifying the various taphonomic agents that influenced the formation of the samples. M. coypus was of great importance to the subsistence of human groups that inhabited the region. Such groups took full advantage of the rodent, using them not only as a food resource but also as a raw material for the manufacture of bone tools and probably also for their hides.
Keywords: Zooarchaeology; Humid Chaco; Myocastor coypus.
INTRODUCCIÓN
En los últimos años se ha comenzado a considerar que los roedores jugaron un rol importante en la subsistencia de los grupos prehispánicos de la Argentina. Numerosos autores (Martínez y Gutiérrez 2004; Quintana et al. 2002, entre otros) han planteado, para el Holoceno tardío, que el uso de recursos variados, sobre todo de aquellos de gran disponibilidad anual, de menor tamaño y rápida recuperación, formó parte de una estrategia que implicó una ampliación de la dieta. En este sentido, diversos autores (Acosta 2005; González de Bonaveri 1997, González 2005; Loponte y Acosta 2008; Santiago 2004) resaltan la importancia, en general, de los roedores en los conjuntos zooarqueológicos de la cuenca Paraná-del Plata. Asimismo, se ha registrado el consumo de roedores en sitios de las provincias de Santa Fe y Buenos Aires, y se realizaron investigaciones sobre el aprovechamiento de Cavia aperea (Acosta 2005; Brunazzo 1999), Hydrochaeris hydrochaeris (Feuillet Terzaghi 2002; Santiago 2004), Galea tixiensis (Quintana 2005, entre otros) y Lagostomus maximus (Valverde 2001). Así, la presencia de M. coypus (coipo o falsa nutria) es significativa en contextos arqueológicos vinculados a sistemas fluviales y lacustres, tanto de la Depresión del Salado, del delta del Paraná, como del Paraná santafesino (Acosta 2005; Escosteguy 2007; González de Bonaveri 2005), ya que se trata de un recurso característico de los humedales.
En forma reciente, las investigaciones arqueológicas en el sector ribereño Paraguay-Paraná, en la provincia del Chaco, han comenzado a esclarecer el panorama de los sistemas sociales del pasado en esta región (Calandra et al. 2000; Lamenza et al. 2006; Salceda et al. 2000; Santini y Plischuk 2006). En tal sentido, el objetivo de este trabajo consiste en analizar, en su calidad de recurso, la significación de M. coypus a partir del estudio de los conjuntos arqueofaunísticos de los sitios El Cachapé Potrero V y Sotelo I, de la provincia del Chaco.
La observación de huellas y marcas nos ha permitido investigar tanto las estrategias económicas, evaluando su aprovechamiento, como los distintos procesos tafonómicos que afectaron a los restos. Estos estudios contribuirán a ampliar el conocimiento de las sociedades cazadoras-recolectoras regionales, más aún cuando son escasos los trabajos zooarqueológicos en el área. La información aportada permitirá contribuir al conocimiento general de las estrategias de explotación de la fauna y el uso del espacio durante el Holoceno tardío.
Fisiografía de la subregión de Esteros, Cañadas y Selvas de Ribera
El área donde se localizan los sitios arqueológicos pertenece a la Provincia Fitogeográfica del Chaco, Distrito Oriental (Figura 1), que se ubica en el Sector Ribereño Paraguay-Paraná del Gran Chaco Meridional (Braunstein et al. 2002), específicamente en lo que se conoce como subregión de Esteros, Cañadas y Selvas de Ribera (Morello 1968) o subregión del Chaco de Bosques y Cañadas, que incluye toda la superficie comprendida entre los ríos autóctonos de la provincia del Chaco, tributarios del Paraguay-Paraná dentro del Chaco Húmedo (Burkart et al. 1999). El modelo de vegetación es el más heterogéneo del Chaco; hay varios tipos de bosques, por un lado, y pastizales y pajonales, por el otro. Las pequeñas diferencias de pendiente del terreno determinan distintas condiciones de permanencia de agua en el suelo, que actúan como modeladoras de las comunidades vegetales de la región. Como consecuencia, el paisaje se presenta como un mosaico de franjas de tierras altas con bosques en galería que acompañan a los cursos de agua, alternados con interfluvios bajos, donde se constituyen esteros, cañadas y bañados, con fisonomías de pastizal, pajonal y sabana. En términos zoogeográficos, el área de estudio se encuentra en el Distrito Chaqueño del Dominio Subtropical de la Subregión Guayano-Brasileña (Ringuelet 1961). La gran variedad de ambientes mencionados en el Chaco Húmedo (bosques, esteros, bañados, sabanas, pastizales, lagos y ríos) permite en la región la presencia de una notable diversidad y cantidad de fauna silvestre. Entre los vertebrados se encuentran más de 150 especies de peces, 40 de anfibios, 50 de reptiles, 350 de aves y 70 de mamíferos. La estructura de los recursos faunísticos de la región se caracteriza por tener una gran cantidad de especies gregarias de hábitos acuáticos, vinculadas a cuerpos de agua permanente.
Figura 1. Mapa del área de estudio.
Características ecoetológicas de Myocastor coypus
M. coypus es una especie de amplia distribución en la Argentina. Se encuentra estrechamente vinculado a cuerpos acuáticos, y se caracteriza por tener conductas territoriales y hábitos alimenticios crepusculares y nocturnos. Las hembras pueden tener hasta tres alumbramientos por año de hasta siete crías por vez, y se reproducen a lo largo de todo el ciclo anual (Colantoni 1993). El peso promedio de los adultos es de 6 kg, pudiendo llegar hasta los 10 kg. La fracción consumible directa de este roedor es aproximadamente el 48% de su peso vivo (Bianchi y Dabove 1984). Poseen hábitos gregarios: conforman grupos de entre 2 y 13 individuos adultos con sus crías. Sólo los machos jóvenes son solitarios. Pueden migrar largas distancias en épocas de grandes crecientes y/o en épocas de seca (Ferrari 1983). La densidad poblacional de este roedor es altamente sensible a la temperatura del agua, a la disponibilidad de alimento y a la cobertura vegetacional existente, y llega a una densidad mayor a 130 individuos por km2. En este sentido, la densidad está relacionada con los cuerpos de agua permanentes, pero también con la variación de los pulsos potámicos.
Los sitios arqueológicos
El Cachapé Potrero V (PV)
El sitio se halla sobre un albardón fluvial de unos 600 m2 ubicado a 80 m del curso de agua secundario Quia, a 150 m de un bañado temporario alimentado estacionalmente por las lluvias, a 10 km del río Guaycurú y a 15 km del río Selva de Oro, los dos ríos más importantes de la región de estudio en el departamento 1ro. de Mayo (26° 53' 47,2" LS y 59° 00' 49" LO). Está localizado a 81 km al noroeste de la ciudad de Resistencia, a 40 km de la costa del Paraguay y a 30 km del río Bermejo. La antigüedad del depósito fue estimada en 920 y 1200 años AP, sobre la base de dos fechados radiocarbónicos obtenidos en valvas de Pomacea canaliculata (920 ± 60 años AP; LP-1699; d13C= -8 ± 2‰) y restos óseos de M. coypus (1200 ± 60 años AP; LP-1730; d13C= -20 ± 2‰). Las excavaciones cubrieron una superficie de 16 m2 mediante cuadrículas de 2 x 2 m. El sedimento fue removido a través de niveles artificiales de 10 cm y tamizado en seco en zarandas con mallas metálicas de 5 mm. El Horizonte A presenta una potencia de 0,65 m, luego se inicia el Horizonte C. La mayor concentración de materiales arqueológicos se encuentra entre los 0,40 y 0,57 m de profundidad (considerando el nivel 0 al punto más alto del albardón).
Sotelo I (SI)
El sitio arqueológico Sotelo I se encuentra localizado a los 26º 47'31.9" LS y 59º 03'31.9" LO, sobre un albardón fluvial de 350 m2 ubicado a 70 m del riacho Malá y rodeado de bañados temporarios, distante a 11,7 km al norte de El Cachapé Potrero V. El sitio fue hallado durante una prospección llevada a cabo en el año 2005, en que se observó parte del material expuesto a la vera del camino vecinal que conduce desde la localidad de La Eduvigis al paraje Campo Antonioli. La excavación fue realizada en marzo del 2007 y cubrió una superficie de 24 m2, mediante cuadrículas de 2 x 2 m. Para realizarla, se utilizó la misma metodología de excavación que en PV. La antigüedad del depósito fue estimada en 1690 ± 90 años AP (LP-1917; d13C= -20 ± 2‰), sobre un fechado radiocarbónico realizado a partir de una muestra ósea de M. coypus proveniente del nivel 4 (0,40-0,50 m de profundidad). La columna estratigráfica, la potencia arqueológica y las mayores densidades de materiales exhiben características similares a las de PV. El Horizonte A presenta una potencia de 0,70 m, luego se inicia el Horizonte C. La mayor concentración de materiales arqueológicos se encuentra entre los 0,40 y 0,64 m de profundidad.
METODOLOGÍA
Las muestras analizadas fueron determinadas taxonómica y anatómicamente mediante el uso de material comparativo de la colección perteneciente a la División Zoología Vertebrados del Museo de La Plata. El análisis cuantitativo se efectuó a partir de los índices de abundancia taxonómica y de partes esqueletarias. Para conocer la abundancia de M. coypus se calculó el número de especímenes óseos (NISP) (Payne 1975) y el número mínimo de individuos (MNI). Para determinar la frecuencia relativa de cada elemento se calculó el número mínimo de elementos (MNE) y luego el número mínimo de unidades anatómicas (MAU y MAU%) (Binford 1984).
Se realizó una estimación del perfil etario de ambas muestras a partir de la secuencia de erupción dentaria de la mandíbula propuesta por Rusconi 1930 (en Acosta 2005). Se consideró como rasgo diagnóstico el grado de erupción de los molares 1, 2 y 3, en todos aquellos especímenes en los que fue posible determinar dicha característica.
Para evaluar la selección antrópica de M. coypus en la dieta de los cazadores-recolectores que habitaron el Chaco Húmedo, como también los procesos tafonómicos a los que sus restos óseos se vieron sujetos, se evaluaron las marcas y huellas presentes en ellos. Los principales indicadores para valorar el procesamiento fueron las huellas de corte, las quebraduras intencionales y la producción de instrumentos óseos. La identificación de huellas y marcas en el conjunto óseo fue llevada a cabo siguiendo los criterios dados por Binford (1981) y Mengoni Goñalons (1988, 1999). En este trabajo se ha utilizado el modelo de Quintana (Quintana 2005; Quintana et al. 2002) para roedores caviomorfos, basado en la morfología, distribución y localización anatómica de las huellas, como marco de referencia para evaluar el contexto de producción y las posibles actividades implicadas (cuereo, desarticulación y descarne). Las modificaciones óseas, tanto naturales como culturales, fueron relevadas mediante el uso de lupa binocular de 10x. Para estimar el grado de meteorización se tuvieron en cuenta los estadios propuestos por Beherensmeyer (1978).
RESULTADOS
Las muestras arqueofaunísticas están conformadas por 9363 especímenes óseos identificados en PV, y 5722 en SI. En la Tabla 1 se muestran las especies y clases mejor representadas en los conjuntos. La diversidad taxonómica presente en los sitios es alta, conformada en PV por 14 especies de mamíferos, 11 de peces, tres de reptiles y, por lo menos, seis de aves; mientras que en SI se encuentran diez especies de mamíferos, ocho de peces, tres de reptiles y, al menos, cinco especies de aves (estas últimas aún en estudio para su determinación taxonómica).
El total de huesos recuperados que fueron identificados como pertenecientes al taxón M. coypus es de 7868 para PV y de 4412 para SI, que representan el 84% y el 77,1% del total de especímenes recuperados en los sitios, con un MNI determinado de 251 y 259, respectivamente. Éste se obtuvo a partir de fémures en PV y de mandíbulas en SI, los que representan el 100% del MAU en los conjuntos estudiados (Tabla 2).
Sobre la base de las mandíbulas se realizó una aproximación a las clases de edad representadas en las muestras. Los porcentajes obtenidos, sobre el total de casos analizados en PV (n= 442) y Sotelo I (n= 503), indican un claro predominio de individuos con el M³ totalmente erupcionado o en erupción (estadios 5 y 6), que corresponden a 63% y 81%, respectivamente. Le siguen los individuos con M3 comenzando a erupcio nar (estadio 4), con 20% y 11,5%, luego aquellos con M2 desarrollado (estadio 3), con 13,5% y 6%, y, por último, los que presentan el M2 sin erupcionar (estadio 2), con 2,7% y 1,1% (Figura 2). Así, aunque están representados individuos de todos los rangos etarios, existe una clara selección hacia individuos adultos. El perfil etario en SI presenta una tendencia más marcada hacia la captura y consumo de individuos adultos respecto del sitio PV, mientras que la proporción de individuos considerados juveniles (7%) es sensiblemente menor. En ambos sitios, más del 82% de las mandíbulas presentan el M³ totalmente erupcionado o en proceso de erupción (estadios 4 al 6), y representan individuos con pesos estimados entre los 5 y 9 kg. Esto marcaría una biomasa estimada en, por lo menos, 1500 kg, lo cual constituye más del 60% de la biomasa total incorporada para cada sitio (Santini 2009).
En la Figura 3 se puede observar, a través del MAU%, la presencia de todos los elementos que componen el esqueleto completo. La representación anatómica muestra los valores más altos para las hemimandíbulas y los huesos superiores del miembro posterior (fémures y tibias), valores intermedios para los elementos del miembro anterior y valores muy bajos para los elementos anatómicos del esqueleto axial (vértebras y costillas) y del autopodio (astrágalos, calcáneos, metapodios y falanges).
Tabla 1. Valores de NISP, NISP% y MNI de los sitios PV y SI.
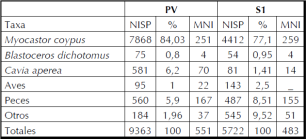
Tabla2. Representación anatómica (NISP, MNE, MAU y MAU%) de M. coypus en los sitios PV y SI.
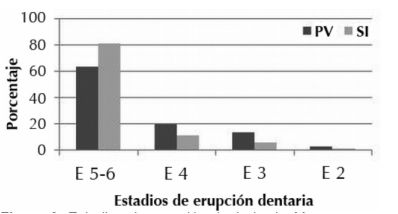
Figura 2. Estadios de erupción dentaria de M. coypus en ambos sitios.
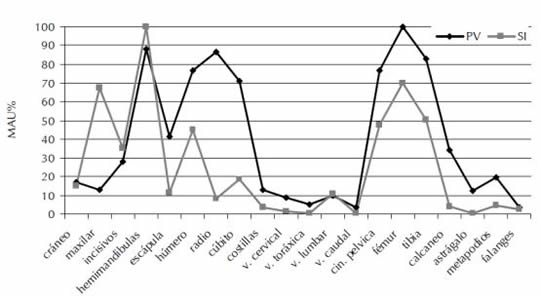
Figura 3. Representación anatómica (NISP, MNE, MAU y MAU%) de M. coypus en los sitios PV y SI.
Modificaciones óseas
Naturales
Dentro de los procesos tafonómicos analizados se observa que, en cuanto a la integridad general de la muestra, el proceso con mayor incidencia ha sido la depositación química, principalmente de óxido de manganeso, que afecta a más del 20% de la muestra (1667 especímenes en PV y 918 en SI). Así, una alta proporción de los huesos presenta una tonalidad que va desde un color pardo amarillento hasta el negro. Estos procesos caracterizan a los suelos de la región bajo estudio.
Por su parte, las marcas de raíces se presentan en un 8,2% de los restos (n= 660) en PV y 6,32% (n= 279) en SI. Los sitios se encuentran emplazados en albardones con presencia de árboles y, en menor medida, plantas rastreras cuyas raíces han sido un agente tafonómico de importancia para la integridad del registro, no tanto por la disolución química sino, fundamentalmente, por la dispersión y redistribución de los materiales en la matriz sedimentaria. En general, hasta los 0,15 m de profundidad se encuentran numerosas raicillas con abundantes ramificaciones, típicas de gramíneas y plantas pequeñas; luego aparecen las raíces de los árboles, generalmente compuestas por una o dos de gran tamaño que se prolongan hasta los 0,40 m, aproximadamente, en una amplia extensión (hasta 3 m de longitud), lo cual afecta tanto la distribución horizontal como vertical de los restos arqueológicos.
Con respecto al grado de meteorización, en ambos conjuntos óseos se observa una clara tendencia hacia los estadios 1 y 2 (sensu Behrensmeyer 1978), con porcentajes superiores al 93%. Esto sugiere un enterramiento relativamente rápido, concordante con las características geomórficas del ambiente. Los elementos que presentan grados más avanzados son, en su mayoría, aquellos ubicados en los niveles estratigráficos inferiores. Asimismo, los suelos de ambos albardones tienen un pH neutro, hecho que, junto con la protección física y química proporcionada por los bolsones de valvas de molusco en los cuales los restos óseos se encuentran entremezclados, habría permitido la supervivencia de los especímenes y su buen estado de conservación.
En cuanto a las modificaciones de los huesos producto de la actividad de roedores, se observan valores muy bajos para todos los taxa presentes en los conjuntos de ambos sitios (PV n= 63 y SI n= 25). En el caso de M. coypus, presentan marcas de roído 18 huesos en PV y 24 en SI; mientras que no se registraron modificaciones producidas por la actividad de carnívoros ni trazas de digestión como resultado de la regurgitación de aves en huesos de este taxón.
Además de la perturbación por acción de las raíces, la integridad del sitio Sotelo I ha sido modificada por causas de origen antrópico como consecuencia de la construcción de un camino vecinal y la remoción de materiales para realizar la instalación de una cerca perimetral.
De origen antrópico
La totalidad de las huellas identificadas corresponden a incisiones o cortes, que se presentan en un porcentaje significativamente bajo de restos respecto del total de especímenes identificados. De hecho, los restos con huellas de corte tienen una frecuencia relativa de 0,41% (n= 49) y 1,38% (n= 63) en PV y SI, respectivamente.
A pesar de la escasa cantidad de huellas de corte que se han registrado, se visualiza una tendencia en los dos conjuntos analizados. En este sentido, vemos que en PV, el 29% de éstas se localizan en las mandíbulas, seguidas por 23% en pelvis, 20% en fémures, 18% en tibias y 10% en húmeros, mientras que en SI, el mayor porcentaje corresponde a las unidades anatómicas fémur (25%), tibia (21%), mandíbula (19%), pelvis (16%) y húmero (13%).
En el caso de las hemimandíbulas, los cortes son en su mayoría longitudinales y están localizados en las ramas mandibulares (n= 3 y n= 2), en los procesos coronoides (n= 4 y n= 4), angular (n= 3 y n= 3), condilar (n= 2 y n= 1) o en dos procesos al mismo tiempo (n= 2 y n= 2). Estas características sugieren que los cortes pudieron haberse producido como consecuencia del proceso de desollamiento, al momento de separar la piel del resto del cuerpo, por la desarticulación de la mandíbula del cráneo y por la remoción de los músculos que unen la piel con el hueso. Según Acosta (2005), este hecho puede estar asociado a la modalidad o técnica utilizada para la extracción del cuero.
En los fémures, los cortes se ubican en la parte anterior del hueso, en las zonas media (40% y 31%, PV y SI, respectivamente) y distal de la diáfisis (60% y 56%). Para SI, el resto se encuentra en el área proximal (12,5%). No se registra corte alguno en las epífisis. Las huellas observadas, mayormente cortas, oblicuas o transversales, pudieron ser el resultado del corte en el extremo distal de la mayor masa de carne del muslo.
En las tibias, la distribución de las huellas sigue un patrón similar al observado para los fémures; la mayoría fueron efectuadas sobre la porción media (55,5% y 46%, PV y SI, respectivamente) y distal del hueso (44,5% y 38,5%), unas pocas en el área proximal de la diáfisis sólo en SI (15%), y ninguna en las epífisis. Las numerosas incisiones sobre el sector medio de la diáfisis serían consecuencia del corte de los tendones.
Aunque no se encontraron huellas en las epífisis, las registradas en los sectores más extremos de las diáfisis del fémur (proximales), de la tibia (proximales) y del húmero (distales), podrían deberse al desmembramiento primario y secundario. Las huellas en los acetábulos y en las ramas pélvicas próximas a ellos podrían vincularse a la desarticulación del miembro posterior (Binford 1981).
Un dato llamativo, compartido por ambos sitios, es la presencia recurrente de fracturas en fémures (PV: 70% y SI: 83%, del total) y, en menor medida, en tibias (PV: 45,5% y SI: 55,5%) (Figuras 4 y 5). Aunque no se han registrado atributos asociados a dichas fracturas, tales como negativos de lascado y puntos de impacto, se plantea que la fractura intencional de dichos elementos se realizó con el propósito de extraer la escasa médula ósea para aprovechar en su totalidad la poca grasa que tiene esta especie (Tulley et al. 2000).
Asimismo, para evaluar el grado de fragmentación de las muestras se utilizó la razón NISP/MNE. Los valores obtenidos fueron de 1,1 para PV y de 1,26 para SI, que sugieren una fragmentación baja. Sin embargo, si aplicamos esta razón solamente a los especímenes identificados como pertenecientes al cráneo, se observa un incremento significativo, con valores de 10,05 y 10,15 respectivamente. Esta alta fragmentación puede estar relacionada no sólo con la obtención de la masa cerebral, sino también con los posibles métodos de captura.
La cantidad de restos óseos con señales visibles de termoalteración es nula para el sitio PV, mientras que en SI sólo se identificaron tres fémures, dos tibias, dos húmeros y dos hemimandíbulas en estado de quemado avanzado. Esta escasa representación de huesos quemados puede estar relacionada con las técnicas de cocción de los alimentos, aunque no se descarta que la presencia de un alto porcentaje de especímenes con precipitación de óxido de manganeso pudiera haber producido un sesgo en su determinación.

Figura 4. Ejemplos de fracturas en fémur de M. coypus.

Figura 5. Detalle de superficie de fractura en fémur de M. coypus.
Instrumentos óseos
Como fuera anteriormente mencionado, y en coincidencia con lo presentado por Quintana et al. (2002) para otros taxones, se considera que M. coypus presentó un aporte global a la subsistencia. Dentro de este aporte se encuentra la producción de instrumentos realizados sobre hueso. La materia prima ósea y la madera han cumplido un importante rol para los grupos históricos de esta región (Arenas 2003; Métraux 1946; entre otros), y esto parece mantenerse en los registros arqueológicos prehispánicos, ante la ausencia de material lítico.
En ambos sitios se recuperaron instrumentos realizados sobre tibias de M. coypus; siete de ellos proceden de PV, y tres de SI. En todos los casos, se observan estrías longitudinales producto de la abrasión, y exhiben también pulido y lustre. Las piezas muestran fractura transversal en su porción proximal y su parte activa en el extremo distal se presenta en forma de punta roma y punzante (Figuras 6 y 7). Para sitios de la llanura aluvial del Paraná medio (norte de la provincia de Santa Fe), se planteó la posible existencia de instrumentos realizados sobre tibias de M. coypus (Pérez Jimeno 2006; Santiago 2004); sin embargo, hasta el presente no se han encontrado evidencias directas de tales instrumentos.
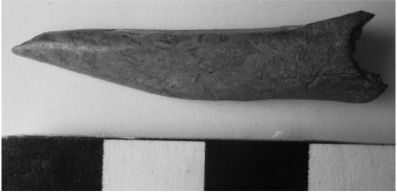
Figura 6. Ejemplo de instrumento realizado sobre tibia de M. coypus, sitio PV.

Figura 7. Ejemplo de instrumento confeccionado sobre tibia de M. coypus, sitio SI.
DISCUSIÓN Y CONSIDERACIONES FINALES
El aprovechamiento de M. coypus es uno de los principales rasgos que caracteriza a la explotación de recursos en estos dos sitios ubicados al norte de la confluencia de los ríos Paraguay-Paraná, no sólo por su alta abundancia relativa en los conjuntos arqueofaunísticos, sino también por su utilización integral por parte de los grupos humanos. El uso e importancia que tuvo como recurso multifuncional M. coypus para los grupos humanos que habitaron la región puede ser corroborado en diferentes crónicas. Existen referencias etnohistóricas respecto de su alta densidad y el aprovechamiento tanto de la carne como de sus cueros. Dobrizhoffer (1967) comenta, durante su estadía con los abipones,
Las que entre los Españoles se llaman nutrias y entre los Abipones Nichigehé abundan tanto en todos los ríos y lagos como entre nosotros las ranas en las charcas. Los salvajes las aprovechan en diversas maneras. Ellos comen la carne de las nutrias. Las mujeres quitan las patas de los cueros y los estiran bien cuadrados con palitos de madera para secarlos sobre el suelo [...] Con estos pellejos que curten solo a mano, componen unas mantas con tanto arte que aún el más perspicaz no descubre en ellas ni costuras ni comisuras. Con estas mantas que son completamente cuadradas cual sábana o manteles, se cubren los Abipones día, y noche contra el riguroso viento sur [...] (Dobrizhoffer 1967).
Considerando su alta predictibilidad, abundancia y fácil captura, habría constituido uno de los recursos con mayor tasa de retorno de la zona. Las características de este roedor, conjuntamente con la ubicación estratégica de los sitios en el paisaje, posibilitan plantear la hipótesis de una captura realizada en los cursos de agua y los bañados localizados en las inmediaciones de los sitios. Como fuera mencionado, se trata del ambiente predilecto de M. coypus, y es donde se concentran mayormente sus poblaciones, de modo tal que se reducen las distancias de obtención y, por lo tanto, los costos de búsqueda, captura y transporte.
Contrastando los datos de abundancia anatómica obtenidos con la información etnohistórica, es muy probable que se obtuvieran varios individuos por evento de caza (captura en masa),
Los Abipones salen generalmente a cazar nutrias cuando durante una sequía de largos años se secan casi por completo también los ríos y los lagos que entonces pueden vadearse de a pie. Ellos envían adelante sus galgos y en un solo día matan algunos cientos de nutrias [...] (Dobrizhoffer 1967).
A partir de los perfiles de edad llevados a cabo, se observa que la presión de captura se dio fundamentalmente sobre ejemplares adultos, los que proveen tanto mayor cantidad de carne como tamaño de pieles. Un aspecto que se debe tener en cuenta es la tecnología utilizada para la captura de las presas, ya que ésta influirá en la estructura etaria del conjunto óseo, tal como fuera planteado a partir de investigaciones etnoarqueológicas realizadas en África (Lupo y Schmitt 2005). La presencia de impronta de cordelería en la decoración de la cerámica en los sitios en cuestión permite inferir, de manera indirecta, la existencia de redes, las cuales podrían haber sido utilizadas para la confección de trampas para la captura de este roedor. Esta técnica habría permitido capturar indiscriminadamente grandes cantidades de individuos, lo cual podría explicar, además, la presencia de individuos juveniles en las muestras. Otro método posiblemente utilizado, descripto en las crónicas (Dobrizhoffer 1967), es el simple uso del garrote, con el cual se obtenían grandes cantidades de individuos por evento de caza, y que podría explicar, en parte, la alta fragmentación que presentan los cráneos.
En relación con las diferencias encontradas respecto de las frecuencias anatómicas, otros autores (e.g., Loponte 2008) plantean que la escasa representación de falanges podría deberse al descarte offsite durante el secado de las pieles, y que la baja presencia de costillas y vértebras se debería a su destrucción in situ por procesos tafonómicos y/o durante el procesamiento del esqueleto. De esta manera, si los grupos que habitaron el Chaco Húmedo hicieron uso de las pieles, tal como se desprende de las crónicas etnohistóricas (Dobrizhoffer 1967), podrían haber procesado y secado los cueros fuera del área de actividad doméstica y descartar los metapodios y las falanges en ese momento. Otra explicación posible podría ser que estos elementos hayan sido transportados conjuntamente con las pieles, considerando que éstas hayan sido bienes de intercambio, tal como se refleja en los relatos etnohistóricos,
[...] El 8 de marzo quise intentar un nuevo viaje hacia la zona de los indios tobas, que vivían entonces del otro lado del Paraná. Acompañe a algunos comerciantes que iban a cambiar pieles de qiya o de grandes ratas acuáticas [...] y se dirigió a la aldea a anunciar nuestra llegada e invitar a llevarnos las pieles de que se disponía. Un cuarto de hora después, vinieron treinta indios -hombres, mujeres y niños- pasando todos a nado el pantano, con sus vestidos ligeros y sus mercancías en la cabeza (D'Orbigny 1999 [1839]: 334).
Cabe aclarar que no se han observado marcas de origen antrópico en los elementos del autopodio recuperados en los conjuntos, las cuales están habitualmente asociadas a la técnica del cuereo en "bolsa" (Escosteguy, comunicación personal 2008).
Como se mencionó anteriormente, la fractura intencional de huesos largos de M. coypus no ha sido registrada en otros conjuntos zooarqueológicos en Argentina, aunque sí se cuenta con información a nivel etnográfico que refiere a conductas similares entre los Aché en Paraguay con especies de tamaño similar (Jones 1983: 103). Para el Chaco argentino, tanto en poblaciones wichí como toba, Arenas (2003: 161) observó, aunque en taxones de mayor tamaño, que luego de cocinadas las presas se guardaban los huesos enteros durante varios días y, ante la falta de otros alimentos, se cocinaba una sopa con los huesos partidos. Las fracturas registradas en los fémures y tibias de M. coypus no presentan las características típicas de aquellas producidas en estado fresco, por lo que se infiere que fueron hechas luego de algún procesamiento que hubiera modificado la humedad del hueso. Uno de los posibles procesos que pudo intervenir en este conjunto es el hervido previo a la fractura de los restos con el propósito de obtener la médula ósea.
Existen varios motivos para sostener que puede tratarse de fracturas de origen antrópico e intencionales, lo cual permitiría descartar la acción de procesos o agentes tafonómicos: 1- una altísima proporción de los fémures presentan fractura transversal u oblicua; 2- ninguna otra unidad anatómica presenta fracturas en porcentajes significativos, a excepción de las tibias (el otro elemento con cierta cantidad de médula ósea aprovechable). De hecho, la totalidad de las falanges (n= 537 y n= 394) y de los metapodios (n= 987 y n= 234) se encuentran enteros, y un porcentaje muy bajo (menos del 5%) de los restantes huesos largos presentan algún tipo de fractura en su zona medial; 3- no se observan grietas en los huesos producidas por desecación; y, por último, 4- es muy baja la probabilidad de que algún agente posdepositacional hubiera afectado sólo a las dos unidades anatómicas que contienen cantidad aprovechable de médula. Por otro lado, en todos los niveles de ambos sitios se recuperó gran cantidad de restos cerámicos, por lo que el hervido pudo haber sido el método culinario utilizado para la preparación de las presas para su consumo; a esto se suma la casi ausencia de elementos óseos con señales de termoalteración, lo que permitiría descartar la utilización de la técnica del asado para la preparación del alimento. Además, numerosos autores (Gifford-González 1993; Lupo y Schmitt 1997) han planteado que la cocción de los alimentos por medio del hervido constituye un método eficiente para maximizar la extracción del tejido adiposo, porque permite aprovechar la grasa de las especies explotadas y remover con mayor facilidad los tejidos blandos del hueso.
La mayoría de las huellas de corte se encuentran localizadas en la mandíbula, lo que permite suponer proponer actividades de desollamiento del animal, hecho ya evidenciado por otros autores para distintas especies de roedores (Acosta 2005; Quintana et al. 2002). La distribución de las huellas de corte en fémures, pelvis y húmeros caracteriza la desarticulación de los miembros del esqueleto axial. De todos modos, debe señalarse que la baja presencia de huellas de corte identificadas es lo esperable en especies de reducido tamaño, como es el caso de M. coypus, que permiten el manipuleo y procesamiento del esqueleto sin necesidad de utilizar tecnología. Por otra parte, no hay que olvidar que no siempre quedan trazas de la utilización de instrumentos cortantes en los huesos, aunque en la práctica estos hayan sido utilizados.
Respecto de la estacionalidad, es esperable que M. coypus estuviera presente a lo largo de todo el año. En el ámbito chaqueño, región de grandes fluctuaciones fluviales entre la estación seca y la húmeda, la mayor presión de caza debe haberse dado durante o inmediatamente después de la estación de lluvias, la cual se extiende entre los meses de octubre a marzo. Es en esta época cuando proliferan los bañados y esteros, y los cursos de agua recuperan sus caudales, lo que posibilita una mayor densidad de individuos por kilómetro cuadrado. Otro indicador de estacionalidad -que permite ubicar la mayor presión de captura en estos meses- es la presencia de grandes cantidades de valvas de moluscos del gasterópodo Pomacea canaliculata chaquensis, que presenta sus picos de actividad y mayor abundancia durante los meses lluviosos, y que se entierra para hibernar durante los meses de invierno.
Los grupos que habitaron los sitios Potrero V y Sotelo I durante el Holoceno tardío presentan numerosas recurrencias en su modo de vida, particularmente en cuanto a la tecnología cerámica y ósea, sus formas de asentamiento (Balbarrey et al. 2007), y la estrategia de subsistencia. La explotación y modo de procesar M. coypus constituye otro aspecto que permite vincular entre sí a las bandas cazadoras-recolectoras tardías de la región de esteros, cañadas y selvas de ribera del Chaco Húmedo. M. coypus representó un gran aporte para la subsistencia en general, ya que presenta numerosas ventajas en relación con otros recursos existentes en el ambiente, tales como su alta densidad poblacional, predictibilidad, y su fácil captura sin la necesidad de contar con una tecnología especializada. Así, este roedor habría proporcionado altas tasas de retorno con un costo de obtención bajo. Los sitios PV y SI representarían un momento de optimización en la explotación de M. coypus, con un aprovechamiento integral de todos los subproductos derivados, que habría significado no sólo la utilización de su carne y grasa como fuente de alimento, sino también sus restos como materia prima para la confección de instrumentos, y, posiblemente, su cuero. Lo expuesto permite concluir que este recurso ha sido de suma importancia en la subsistencia de los grupos cazadores-recolectores-pescadores que habitaron el área.
Agradecimientos
Este proyecto se concretó a través de una beca otorgada por el Consejo Nacional de Investigaciones Científicas y Técnicas (CONICET). Quisiera agradecer a los señores Eduardo Bolaños y Pablo Sotelo, por permitir gentilmente la realización de los trabajos de campo en sus respectivos establecimientos. Asimismo, extiendo este agradecimiento a los evaluadores, por las sugerencias realizadas.
REFERENCIAS CITADAS
1. Acosta, A. 2005 Zooarqueología de cazadores-recolectores del extremo nororiental de la provincia de Buenos Aires (humedal del río Paraná inferior, Región Pampeana, Argentina). Tesis Doctoral inédita. Facultad de Ciencias Naturales y Museo, Universidad Nacional de La Plata. La Plata. [ Links ]
2. Arenas, P.2003 Etnografía y alimentación entre los Toba-Ñachilamolek y Wichí-Lhuku'tas del Chaco Central (Argentina). Buenos Aires.
3. Balbarrey G., C. De Feo, G. Lamenza, M. Santini y H. A. Calandra 2007 Espacio Social y Territorialidad de Sociedades Prehispánicas del Chaco Argentino. Actas del XXVII Encuentro de Geohistoria Regional. Facultad de Filosofía, Universidad Nacional de Asunción, Asunción. Publicado en CD formato libro.
4. Behrensmeyer, A. K. 1978 Taphonomic and ecologic information from bone weathering. Paleobiology 4: 150-162.
5. Bianchi, M. A. y O. Dabove1984 Elaboración de carne de nutria. Utilización de la carne de nutria sudamericana en la elaboración de productos industriales para consumo humano. Noticieteca 14 (84): 222-224.
6. Binford, L. R. 1981 Bones: Ancient Men and Modern Myths. Academic Press, Nueva York.
7. Binford, L. R. 1984 Faunal Remains from Klasies River Mouth. Academic Press, Orlando.
8. Braunstein, J. A., S. A. Salceda, H. A. Calandra, M. G. Méndez y S. O. Ferrarini 2002 Historia de los chaqueños. Buscando en la "papelera de reciclaje" de la antropología sudamericana. Acta Americana, Journal of the Swedish Americanist Society 10 (1): 59-88.
9. Brunazzo, G. A.1999 Investigaciones arqueológicas en el sitio La Norma (Partido de Berisso, Provincia de Buenos Aires, Argentina). Actas del XII Congreso Nacional de Arqueología Argentina 3: 101-106. La Plata.
10. Burkart, R., N. Bárbaro, R. Sánchez y D. Gómez 1999 Eco-regiones de la Argentina. Administración de Parques Nacionales, Programa Desarrollo Institucional Ambiental, Buenos Aires.
11. Calandra, H. A., M. G. Méndez, S. A. Salceda, G. Lamenza, M. Lanciotti, L. Del Papa y N. Duhalde 2000 Estudio preliminar de los restos cerámicos hallados en el sitio arqueológico "El Cachapé" (Chaco, Argentina). Actas del XX Encuentro de Geohistoria Regional I: 157-171. Resistencia.
12. Colantoni, L. O.1993 Ecología poblacional de la nutria (Myocastor coypus) en la provincia de Buenos Aires. Fauna y Flora Silvestres 1: 1-24.
13. Dobrizhoffer, M.1967 Historia de los abipones. Tomo 1. Universidad Nacional del Noroeste, Facultad de Humanidades, Departamento de Historia, Resistencia.
14. D'Orbigny, A.1999 [1839] Viaje a la América Meridional I. Emecé, Buenos Aires.
15. Escosteguy, P. D.2007 Estudio sobre restos de Myocastor coypus en sitios de la cuenca inferior del río Salado. En Arqueología en Las Pampas, editado por C. Bayón, A. Pupio, M. I. González, N. Flegenheimer y M. Frère, Tomo 1, pp. 475-487. Sociedad Argentina de Antropología, Buenos Aires.
16. Ferrari, M. 1983 El coipo. Fauna Argentina 20. Centro Editor de América Latina, Buenos Aires.
17. Feuillet Terzaghi, M. R.2002 El registro arqueofaunístico de vertebrados del sitio Playa Mansa I (Arroyo Seco, Depto. Rosario, provincial de Santa Fe). Aportes y perspectivas. Tesis de Licenciatura inédita, Facultad de Humanidades y Artes, Universidad Nacional de Rosario, Rosario
18. Gifford-Gonzalez, D.1993 Gaps in the zooarchaeological analyses of butchery: is gender an issue. En: Etnhoarchaeological and Experimental Contributions to the Interpretation of Faunal Remains, editado por J. Hudson, pp. 181-199. Occasional Paper 21, Center for the Archeological Investigations, Southern Illinois University, Carbondale.
19. González, M. I. 2005 Arqueología de alfareros, cazadores y pescadores pampeanos. Sociedad Argentina de Antropología, Buenos Aires.
20. González de Bonaveri, M. I.1997 Potsherds, "coypo" teeth, and fish bones: Hunter-gatherer-fishers in the Río Salado (Pampa Region, Argentina). En Quaternary of South America and Antartic Peninsula, 10: 255-278.
21. Jones, K. T.1983 Forager archaeology: The Aché of Eastern Paraguay. En Carnivores, Human Scavengers and Predators: A Questions of Bone Technology, editado por G. M. LeMoine y A. S. MacEachern, pp. 171-191. The Archaeological Association of the University of Calgary, Calgary.
22. Lamenza, G., G. Balbarrey, B. Aguirre y H. A. Calandra 2006 Complejidad e interacción de sociedades prehispánicas ribereñas del Gran Chaco argentino. Actas del XXV Encuentro de Geohistoria Regional: 285-292. Resistencia, Chaco.
23. Loponte, D. 2008 Arqueología del humedal del Paraná inferior (bajíos ribereños meridionales). Asociación Amigos del Instituto Nacional de Antropología y Pensamiento Latinoamericano, Buenos Aires.
24. Loponte, D. y A. Acosta 2008 El registro arqueológico del tramo final de la cuenca del Plata. En Entre la tierra y el agua. Arqueología de humedales de Sudamérica, compilado por D. Loponte y A. Acosta, pp. 125-164. Asociación Amigos del Instituto Nacional de Antropología y Pensamiento Latinoamericano (AINA), Buenos Aires.
25. Lupo, K. D. y D. N. Schmitt 1997 Experiments in bone boiling: nutritional returns and archaeological reflections. Anthopozoologica 25-26: 137-144.
26. Lupo, K. D. y D. N. Schmitt 2005 Small prey hunting technology and zooarchaeological measures of taxonomic diversity and abundance: ethnoarchaeological evidence from Central African forest foragers. Journal of Anthropological Archaeology 24: 335-353.
27. Martínez, G. y M. A. Gutiérrez 2004 Tendencias en la explotación humana de la fauna durante el Pleistoceno final y Holoceno en la Región Pampeana (Argentina). En Zooarchaeology of South America, editado por G. L. Mengoni Goñalons, pp. 81-98. BAR International Series. 1298. Archaeopress, Oxford.
28. Mengoni Goñalons, G. L. 1988 Análisis de materiales faunísticos de sitios arqueológicos. Xama 1: 71-120.
29. Mengoni Goñalons, G. L. 1999 Cazadores de guanacos de la estepa patagónica. Sociedad Argentina de Antropología, Buenos Aires.
30. Métraux, A.1946 Ethnography of the Chaco. En Handbook of South American Indians, editado por J. Steward. Bulletin of the Bureau of American Ethnology 135 (I): 197-370. Washington, D.C.
31. Morello, J. 1968 La vegetación de la República Argentina. Las grandes unidades de vegetación y ambiente del Chaco argentino. Instituto Nacional de Tecnología Agropecuaria (Serie Fitogeográfica N°10), Buenos Aires.
32. Payne, S. 1975 Partial recovery an sample bias. En Archaeozoological Studies, editado por A. T. Clason, pp. 7-17. Ámsterdam.
33. Quintana, C. A.2005 Despiece de micro roedores en el Holoceno Tardío de las Sierras de Tandilla (Argentina). Archaeofauna 217: 227-241.
34. Quintana, C. A., F. Valverde y D. L. Mazzanti 2002 Roedores y lagartos como emergentes de la diversificación de la subsistencia durante el Holoceno tardío en sierras de la Región Pampeana Argentina. Latin American Antiquity 13 (4): 455-473.
35. Ringuelet, R. A. 1961 Rasgos fundamentales de la zoogeografía de la Argentina. Physis 22 (63): 151-170.
36. Salceda, S. A., M. G. Méndez, H. A. Calandra, M. Santini, M. A. Giovanetti y G. Couso 2000 Análisis preliminar de los restos faunísticos del Sitio El Cachapé (Chaco, Argentina). Actas del XX Encuentro de Geohistoria Regional II: 795-806. Resistencia, Chaco.
37. Santiago, F. C. 2004 Los roedores en el "menú" de los habitantes de Cerro Aguará (provincia de Santa Fe): su análisis arqueofaunístico. Intersecciones en Antropología 5: 3-18.
38. Santini, M.2009 Prehistoria de la región meridional del Gran Chaco: aportes del análisis de restos faunísticos en la reconstrucción de las estrategias adaptativas de los grupos aborígenes durante el Holoceno tardío. Tesis inédita. Facultad de Ciencias Naturales y Museo, Universidad Nacional de La Plata, La Plata
39. Santini, M. y M. Plischuk 2006 Sector Ribereño Paraguay-Paraná: análisis de los conjuntos de artefactos óseos provenientes de dos sitios arqueológicos. Actas del XXVI Encuentro de Geohistoria Regional, pp. 491-495. Resistencia, Chaco.
40. Tulley R. T., F. M. Malekian, J. C. Rood, M. B. Lamb, C. M. Champagne, S. M. Redmann, R. Patrick, N. Kinler y C. T. Raby 2000 Analysis of the nutritional content of Myocastor coypus. Journal of Food Composition and Analysis 13 (2): 117-125.
41. Valverde, F. 2001 Huellas y marcas sobre hueso. En Cueva Tixi: cazadores y recolectores de las sierras de Tandilla Oriental. 1 Geología, Paleontología y Zooarqueología, editado por D. L. Mazzanti y C. A. Quintana, pp. 137-155. Publicación Especial 1 del Laboratorio de Arqueología. Universidad Nacional de Mar del Plata, Mar del Plata.

