










Services on Demand
Journal

Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO
Related links
-
 Similars in
SciELO
Similars in
SciELO
Share

 Permalink
PermalinkIntersecciones en antropología
On-line version ISSN 1850-373X
Intersecciones antropol. vol.14 no.2 Olavarría July./Dec. 2013
ARTÍCULOS
Análisis zooarqueológico del sitio Paso Otero 4 (Partido de Necochea, provincia de Buenos Aires, Argentina). Aportes a la discusión de modelos de subsistencia de la región pampeana
María Clara Álvarez, Ana Paula Alcaráz, María A. Gutiérrez y Gustavo Martínez
María Clara Álvarez. Consejo Nacional de Investigaciones Científicas y Técnicas (CONICET)-Investigaciones Arqueológicas y Paleontológicas del Cuaternario Pampeano (INCUAPA). Facultad de Ciencias Sociales, Universidad Nacional del Centro de la Provincia de Buenos Aires (UNICEN). Av. Del Valle 5737 (B7400JWI). Olavarría, Buenos Aires, Argentina. E-mail: malvarez@soc.unicen.edu.ar
Ana Paula Alcaráz. CONICET-INCUAPA. Facultad de Ciencias Sociales, UNICEN. Av. Del Valle 5737 (B7400JWI). Olavarría, Buenos Aires, Argentina. E-mail: aalcaraz@soc.unicen.edu.ar
María A. Gutiérrez. CONICET-INCUAPA. Facultad de Ciencias Sociales, UNICEN. Av. Del Valle 5737 (B7400JWI). Olavarría, Buenos Aires, Argentina. E-mail: mgutierr@soc.unicen.edu.ar
Gustavo Martínez. CONICET-INCUAPA. Facultad de Ciencias Sociales, UNICEN. Av. Del Valle 5737 (B7400JWI). Olavarría, Buenos Aires, Argentina. E-mail: gmartine@soc.unicen.edu.ar
Recibido 28 de marzo 2012.
Aceptado 27 de agosto 2012
Resumen
El sitio Paso Otero 4 se localiza en el partido de Necochea, en la región pampeana. Los fechados radiocarbónicos sobre materia orgánica de sedimento ubican las ocupaciones humanas entre ca. 8900 y 4600 años AP. El objetivo de este trabajo es presentar las características del conjunto faunístico de Paso Otero 4, con el fin de discutir las principales tendencias de la subsistencia de los cazadores-recolectores que ocuparon el sitio durante el Holoceno temprano y medio y contribuir con nuevos datos a la discusión de los modelos de explotación faunística propuestos para la región pampeana. Se integran y discuten los resultados de los estudios zooarqueológicos y tafonómicos, los cuales fueron evaluados en conjunto con las dinámicas ambientales inferidas para el sitio. Las diferentes líneas de evidencia indican que éste habría funcionado como un campamento en el cual se llevaron a cabo actividades vinculadas a las últimas etapas de producción artefactual y manutención de instrumentos líticos, así como al procesamiento y consumo de una importante cantidad de taxones; entre ellos carnívoros, aves pequeñas y grandes, artiodáctilos, roedores grandes y armadillos pequeños y grandes. En términos comparativos, en las ocupaciones más tempranas (ca. 8900 a 7700 años AP; Niveles Inferiores) se registró una mayor diversidad faunística y se infirió una estrategia de subsistencia más diversificada. Por su parte, en las ocupaciones más tardías (ca. 7700 a 4600 años AP; Niveles Superiores) la diversidad de especies es un poco más baja que en momentos previos y la estrategia de subsistencia estuvo orientada hacia la obtención de artiodáctilos.
Palabras clave: Estudios zooarqueológicos y tafonómicos; Cazadores-recolectores; Holoceno temprano y medio; Modelos de subsistencia de la región pampeana.
Abstract
Zooarchaeological analysis of Paso Otero 4 site, Buenos aires province, Argentina. Constributions to the discussion of the pampean region subsistence models. Paso Otero 4 is located in the Pampean region of Argentina, within the Interserrana area. Human occupations are chronologically placed between ca. 8900 and 4600 years BP. The objective of this paper is to present the main characteristics of the faunal record of the site through the zooarchaeological and taphonomical analyses. In addition, huntergatherers subsistence trends for this time period are discussed. In the light of these new data, faunal exploitation models proposed for the Pampean region are evaluated. The results of different lines of evidence suggest that activities such as the final artefact production stages and maintenance of lithic instruments, as well as the processing and consumption of an important number of taxa such as carnivores, small and large birds, artiodactyls, large rodents, and small and large armadillos, were carried out at the site. In comparative terms, during the earlier occupations (ca. 8900 to 7700 years BP; Lower Levels) a diversified subsistence strategy is recorded, while during the later occupations (ca. 7700 to 4600 years BP; Upper Levels) the diversity of species is lower and the subsistence strategy is oriented to artiodactyl procurement.
Keywords: Zooarchaeological and taphonomical studies; Hunter-gatherers; Early and Middle Holocene; Subsistence models of the Pampean region.
INTRODUCCIÓN
Los estudios zooarqueológicos comenzaron a ocupar un rol protagónico y creciente en la arqueología de la región pampeana en los últimos 30 años. Como consecuencia de este desarrollo, se han publicado varias síntesis sobre modelos de subsistencia de los cazadores-recolectores pampeanos durante el Pleistoceno tardío y el Holoceno (Politis y Salemme 1990; Miotti y Salemme 1999; Quintana y Mazzanti 2001; Martínez y Gutiérrez 2004; Gutiérrez y Martínez 2008). El hallazgo y posterior excavación de nuevos sitios arqueológicos en la región durante los últimos años han brindado novedosos conjuntos faunísticos, cuyas cronologías los ubican en lapsos escasamente representados en el registro arqueológico, como el Holoceno medio (Bayón et al. 2010; Bonomo y León 2010; Massigoge 2011; entre otros). Estos nuevos datos han permitido complementar, y en algunos casos modificar, los modelos mencionados. A estos contextos se suma la evidencia del sitio Paso Otero 4, motivo de esta contribución. Paso Otero 4 (PO4) es un sitio a cielo abierto, localizado en la cuenca media del río Quequén Grande (región pampeana, provincia de Buenos Aires, Argentina) (Figura 1). Este sitio está ubicado en la localidad Paso Otero (38° 12' 41.79" S; 59° 06' 33.95" O), en la cual se registró un total de 12 sitios arqueológicos y depósitos naturales que abarcan cronológicamente ca. 10.500 años (Johnson et al. 1998, 2012; Martínez 2001, 2006; Gutiérrez et al. 2010, 2011; Martínez y Gutiérrez 2011a).
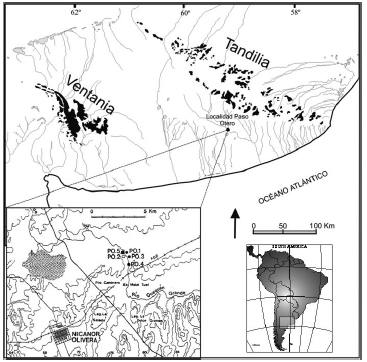
Figura 1. Ubicación del sitio Paso Otero 4.
El sitio PO4 fue sistemáticamente excavado en distintas temporadas de campo entre los años 2008 y 2010, en las cuales se recuperó una gran cantidad y diversidad de materiales arqueológicos en una superficie de 14 m2. El depósito arqueológico, asignado al Holoceno temprano y medio, alcanzó una profundidad de 6,15 m desde el nivel 0 (5,15 m desde la superficie). Los materiales se registraron en un paquete continuo, distribuidos a través de ca. 2,50 m, en sedimentos asignados al Miembro Río Salado de la Formación Luján, denominados en el sitio como Unidad 2 (Gutiérrez et al. 2010, 2011). En PO4 se recuperó una gran variedad de materiales zooarqueológicos y se determinó un total de 28.938 especímenes. Este número incluye gasterópodos marinos, cáscaras de huevo, placas de armadillos y unidades esqueletarias correspondientes a una gran diversidad de especies. El conjunto lítico está conformado por ca. 500 artefactos (Barros 2012).
Entre las categorías artefactuales predominan ampliamente los desechos de talla, seguidos por los instrumentos y los ecofactos. En relación con las rocas presentes, se destaca la ortocuarcita, con valores cercanos al ca. 95%. Respecto de los instrumentos, los más representados son las lascas con rastros y los artefactos de formatización sumaria (Barros et al. 2011; Barros 2012). El objetivo de este trabajo es dar a conocer las características del conjunto faunístico de PO4, con el fin de discutir las principales tendencias de la subsistencia de los cazadores-recolectores que ocuparon el sitio durante el Holoceno temprano y medio y contribuir con nuevos datos a la discusión de los modelos de explotación faunística propuestos para la región pampeana.
ESTRATIGRAFÍA Y CRONOLOGÍA
La secuencia estratigráfica definida para el sitio es la descripta para los valles fluviales de la provincia de Buenos Aires (Fidalgo et al. 1973; Zárate et al. 2000), y se presenta de manera esquemática en la Figura 2. En la base de la secuencia se reconoce la parte superior del Miembro Guerrero de la Formación Luján, denominada localmente Unidad 1. Por encima se registra el Suelo Puesto Callejón Viejo, el cual se observa hacia el perfil Norte de la excavación, pero en el sector del sitio no presenta continuidad. En discordancia erosiva con aquel, se encuentran los sedimentos del Miembro Río Salado de la Formación Luján, denominados localmente Unidad 2. Sobre esta última se registró un evento pedogenético vinculado al Suelo Puesto Berrondo. Finalmente, la secuencia se completa con sedimentos fluviales y eólicos que indican eventos periódicos de inundación correspondientes a la Unidad 3 y el suelo actual (Gutiérrez et al. 2010, 2011). Diez superficies de estabilización del paisaje (horizontes "A" de suelo enterrado) fueron identificadas a lo largo de esta secuencia (ver debajo), la cual, en líneas generales, es la que ha sido reconocida para el curso medio del río Quequén Grande (Johnson et al. 1998, 2012; Zárate et al. 1998, 2000; Martínez 2002- 2004). Dado que el registro arqueológico se localizó exclusivamente en la Unidad 2, en este trabajo sólo se informan las características geológicas básicas de esta unidad, así como sus dinámicas depositacionales. En este sentido, los sedimentos de esta unidad habrían sido depositados por procesos lacustres muy calmos y de baja energía, ocurridos en ambientes caracterizados por lagunas interconectadas a través del paisaje, en ambientes pantanosos (ver Zárate et al. 2000 por ambientes similares en otros sitios del curso medio de la cuenca).
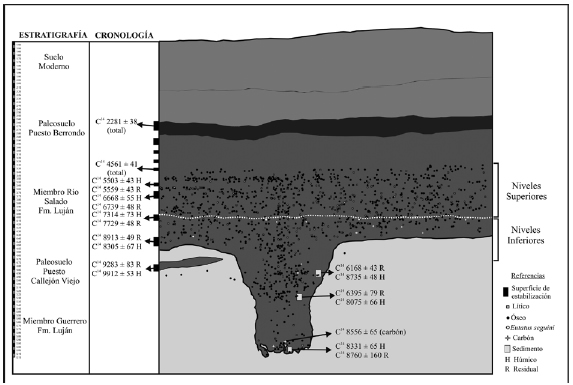
Figura 2. Perfil del sitio con la procedencia de los fechados expresados en años radiocarbónicos, la distribución de los materiales en planta y la división entre los Niveles Inferiores y los Niveles Superiores.
Hacia la base del depósito arqueológico se registró la presencia de un rasgo destacado. Se trata de una estructura semicircular, en forma de U, rellena con un sedimento marrón oscuro (Figuras 2 y 3). Dentro del rasgo, los restos arqueológicos recuperados fueron escasos y estaban concentrados principalmente en su parte superior. Sobre la base de distintas líneas de evidencia se discutió el origen y la funcionalidad de dicho rasgo. Se tuvo en cuenta su morfología y sus dimensiones, la relación entre la cronología obtenida de la secuencia estratigráfica y aquella proveniente del rasgo, el contenido y la distribución del material en su interior, así como parámetros químicos del sedimento de relleno. Esta información se interpretó considerando las características de cuevas y galerías producidas por animales cavadores y las de estructuras de cavado antrópico, como pozos de almacenamiento y de procesamiento, cisternas, basureros ceremoniales o depósitos rituales, pozos de almacenamiento de tecnofacturas, etc. De este análisis surgieron dos conclusiones: por un lado, se descartó el origen natural del rasgo y, por el otro, se sugirió que la hipótesis más parsimoniosa que explica su origen es la excavación de un pozo de agua (Gutiérrez y Martínez 2010; Martínez y Gutiérrez 2011b). Tres fechados radiocarbónicos de materia orgánica de sedimentos y uno sobre carbón sitúan la cronología del rasgo entre ca. 8700 y 8100 años AP.

Figura 3. Vista del perfil estratigráfico del sitio y del pozo de agua en la base.
Debido a la escasa preservación de colágeno, situación común a todos los sitios de la localidad (ver discusión en Gutiérrez et al. 2001), los distintos intentos por datar huesos a través de AMS no arrojaron resultados (Gutiérrez et al. 2011). La cronología de PO4 fue obtenida a partir de la datación de la materia orgánica de los suelos enterrados, al igual que en los demás sitios excavados en la cuenca (ver síntesis en Johnson et al. 2012). Los fechados presentados en este trabajo se expresan en años radiocarbónicos AP. De las diez superficies de estabilización reconocidas, siete han sido datadas (fracciones totales, húmicas y residuales; NSF Arizona AMS Laboratory), y arrojaron edades de ca. 9900 (Suelo Puesto Callejón Viejo), ca. 8900, ca. 7700, ca. 6700, ca. 5500, ca. 4600 y ca. 2200 años AP (Suelo Puesto Berrondo)1.
Las ocupaciones humanas están datadas entre ca. 8900 y ca. 4600 años AP (Figura 2). Los materiales arqueológicos se presentan de manera ininterrumpida entre los ca. 3,65 y 6,15 m y tienen, en general, una distribución homogénea. No obstante, operativamente se estableció una separación en Niveles Inferiores (NI) y Niveles Superiores (NS), sobre la base del registro de fauna extinguida en los primeros. Los NI (ca. 8900 a 7700 años AP) se definen desde la base del depósito arqueológico hasta ca. 4 m, incluyendo los restos provenientes del rasgo. Los NS (ca. 7700 a 4600 años AP) se extienden desde aquí hasta la parte medio/cuspidal del Miembro Río Salado de la Formación Luján (Figura 3). Dicha separación coincide también con cambios paleoambientales evidenciados por el registro de polen, diatomeas y fitolitos (Gutiérrez et al. 2011). Las ocupaciones humanas del Holoceno temprano habrían tenido lugar bajo climas regionalmente cálidos, pero interrumpidos por pulsos secos. Por otro lado, hacia el Holoceno medio el clima se habría tornado frío y árido, principalmente hacia el final de éste (Gutiérrez et al. 2011).
MATERIALES Y METODOLOGÍA
La muestra analizada comprendió el número total de especímenes óseos del sitio (N= 28.938), que fueron determinados a distinto nivel taxonómico y anatómico a través del uso de colecciones de referencia alojadas en la Facultad de Ciencias Sociales (Olavarría, UNICEN). Para la totalidad de los taxones determinados se obtuvieron las medidas de: abundancia taxonómica y anatómica, número de especímenes óseos identificados por taxón (NISP) y número mínimo de individuos (MNI) (Grayson 1984; Klein y Cruz-Uribe 1984; entre otros). Posteriormente, se construyeron perfiles esqueletales para Lama guanicoe (guanaco). Para esto se utilizó, además del NISP y el MNI, el número mínimo de elementos (MNE), el número mínimo de unidades anatómicas identificadas por especie (MAU) y esta misma medida estandarizada (MAU%) (Binford 1978; Klein y Cruz-Uribe 1984; entre otros).
Las siguientes variables tafonómicas fueron utilizadas para los taxones >1 kg, con el objetivo de evaluar la integridad de la muestra, así como de determinar la incidencia de agentes que modifican la superficie cortical de los huesos o producen su desplazamiento espacial: meteorización, marcas de roedores, marcas de carnívoros, marcas de raíces, manganeso, pisoteo, tipo de fractura y abrasión sedimentaria (Behrensmeyer 1978; Haynes 1980; Binford 1981; Shipman 1981a; Grayson 1984; Johnson 1985; Olsen y Shipman 1988; Lyman 1994; Gutiérrez y Kaufmann 2007). Estas variables fueron registradas para una submuestra conformada por 1389 especímenes, la cual incluye todos los restos óseos con excepción de los dientes, los huesos indeterminados, las placas de armadillos y los restos de gasterópodos. Con el objetivo de identificar indicadores de procesamiento en el conjunto óseo, se consideraron las siguientes variables: alteración térmica (David 1990; Benett 1999), tipo de fractura (Johnson 1985) y huellas de corte (Binford 1981; Shipman 1981b). Además, los perfiles esqueletales de Lama guanicoe fueron correlacionados con el Índice de Utilidad de Carne de Guanaco (Borrero 1990), a través de una correlación de Rho de Spearman.
Se utilizó el Índice de Artiodáctilos (IA) para evaluar la orientación al procuramiento de estos en la dieta (Broughton 1994). El IA fue calculado utilizando el NISP, tal como es propuesto por Lyman (2008). Dicho índice se calculó dividiendo la suma del NISP total de artiodáctilos por la suma de este número y el NISP total de los armadillos y roedores que presentaban evidencias de procesamiento antrópico. De forma complementaria se aplicó el Índice de Shannon-Wiener, utilizando el programa estadístico PAST version 2.08. Éste arroja un valor de diversidad a partir de la obtención de logaritmos de la proporción de especímenes y fue calculado sólo para aquellos taxones que presentaban evidencias de procesamiento. Finalmente, para evaluar la riqueza taxonómica se aplicó el Ntaxa, considerando sólo las especies con evidencias de procesamiento. Éste refiere al número de taxones identificados, los cuales son sumados, considerando los niveles taxonómicos de identificación alcanzados, con el objetivo de no sobredimensionar esta medida (Grayson 1991).
En el caso del conjunto de microvertebrados (<1 kg, roedores, anuros y ofidios, principalmente), se identificaron evidencias de digestión teniendo en cuenta, para el caso de los roedores, las categorías propuestas por Andrews (1990) y Fernández-Jalvo y Andrews (1992) para molares, incisivos y poscráneo. Para las restantes especies se determinó la presencia de pulido, redondeo y hoyuelos gástricos. En el caso de la meteorización, se emplearon los estadíos establecidos por Andrews (1990) para mamíferos <5 kg. Por último, para evaluar la intensidad de la acción de las raíces y del manganeso, fueron elaboradas las siguientes categorías, considerando el porcentaje del hueso afectado: 1 (0-25%), 2 (26-50%), 3 (51-75%) y 4 (76-100%).
RESULTADOS
Composición del conjunto faunístico
Considerando la diversidad de especies de la muestra analizada (N= 28.938, Tabla 1), 13.741 especímenes corresponden a fragmentos de cáscara de huevo, asignados a la Familia Rheidae, aunque probablemente correspondan a Rhea americana (ñandú), dada la distribución de las especies de Rheidae durante el Holoceno temprano y medio en el área (Tambussi y Acosta Hospitaleche 2002). Dos de los fragmentos de cáscara de huevo están decorados y dos poseen perforaciones antrópicas (Figura 4), mientras que ca. 18% están térmicamente alterados. De la muestra total, 10.768 especímenes corresponden a fragmentos indeterminados (<4 cm), de los cuales ca. 9% presenta alteración térmica. Por otro lado, se identificaron 2378 placas óseas, entre las que se encuentran representadas en porcentajes muy similares las de los armadillos Chaetophractus villosus (peludo) y Zaedyus pichiy (piche). En ambos casos, ca. 4% de estos restos están térmicamente alterados. Finalmente, se recuperaron 124 especímenes de gasterópodos marinos y uno de un bivalvo (Glycimeris longior). Los primeros están muy fragmentados, lo que imposibilita su determinación en niveles taxonómicos más específicos. Los datos para la totalidad de la muestra considerando el NISP, NISP%, MNI y las evidencias de procesamiento son presentados en la Tabla 1.
Tabla 1. Tabla taxonómica del sitio PO4.
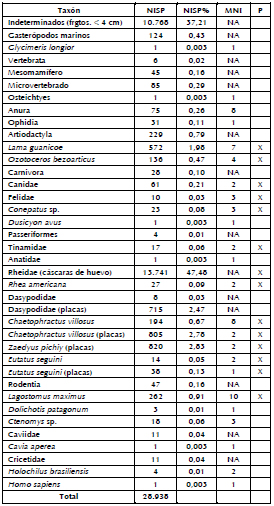
Referencias: NISP= número mínimo de especímenes; MNI= número mínimo de individuos; NA= no aplica; P= procesamiento; X= taxones procesados.

Figura 4. Fragmentos de cáscara de huevo con perforaciones antrópicas (abajo) y decoración (arriba).
Teniendo en cuenta el número de especímenes, la especie más representada es Lama guanicoe (guanaco), seguida por Lagostomus maximus (vizcacha), Chaetophractus villosus (peludo), Ozotoceros bezoarticus (venado de las pampas), carnívoros (entre ellos Dusicyon avus, cánido extinguido), Conepatus sp. (zorrino), Rhea americana (ñandú), Tinamidae, Anatidae, Eutatus seguini (armadillo extinguido) y Dolichotis patagonum (mara). También se identificó una segunda falange de Homo sapiens, recuperada con el resto de los materiales arqueológicos (Tabla 1). En el caso de los microvertebrados, la mayor frecuencia corresponde a anuros, siendo éste el Orden mejor representado. También predomina el Orden Rodentia, correspondiente a especímenes de roedores que no pudieron ser asignados a niveles taxonómicos más específicos, seguido por Ctenomys sp. y las Familias Caviidae y Cricetidae, con igual frecuencia. Asimismo, se pudieron determinar restos a nivel de especie, como es el caso de Holochilus brasiliensis y Cavia aperea, aunque su frecuencia es baja en relación con el conjunto total. Los reptiles se encuentran representados exclusivamente por restos de ofidios, y en su totalidad corresponden a vértebras. Por último, se recuperó un espécimen correspondiente a Osteichtyes (Tabla 1).
La diversidad faunística fue recalculada teniendo en cuenta la separación entre los NI y los NS. En los NS se concentra la mayor cantidad de especímenes de gasterópodos marinos (ca. 94%), así como de los restos de Conepatus sp. (ca. 91%) y de Lama guanicoe (ca. 77%). Las especies Ozotoceros bezoarticus y Lagostomus maximus, así como las Familias Canidae y Tinamidae presentan frecuencias similares en ambas unidades. Por otro lado, en los NI hay una mayor representación de Rhea americana (ca. 74%), Felidae (90%) y Chaetophractus villosus (ca. 60%). El NISP de Dolichotis patagonum no fue suficiente para evaluar las diferencias entre ambos niveles (uno en los NS y dos en los NI). Finalmente, Eutatus seguini y Dusicyon avus sólo están representados en los NI, mientras que Homo sapiens, Glycimeris longior y Anatidae sólo están presentes en los NS. En el caso de los microvertebrados, los NS concentran la mayor cantidad de los restos correspondientes a ofidios (ca. 87%) y Ctenomys sp. (ca. 72%). Por el contrario, en los NI, los anuros (ca. 75%) dominan ampliamente la muestra. Los especímenes atribuibles al Orden Rodentia presentan frecuencias similares en ambas unidades. En relación con las Familias Caviidae y Cricetidae, si bien cuentan con una mayor representación en los NS, dicha diferencia no es significativa debido a los bajos valores de NISP para cada una de éstas. Lo mismo ocurre en el caso de Holochilus brasiliensis, cuyo NISP no es suficiente para evaluar diferencias entre ambas unidades (tres en los NS y uno en los NI). Por último, Cavia aperea y Osteichtyes sólo están representados en los NS (Tabla 2).
Tabla 2. Diferencias en la composición faunística para los NS y los NI.
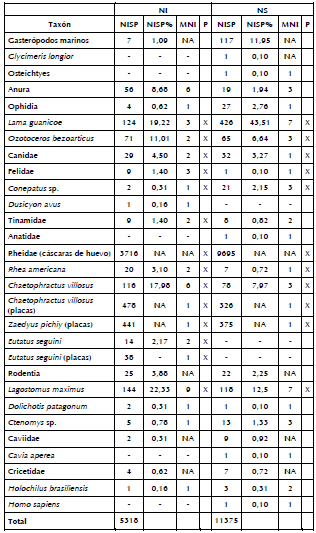
Referencias: NISP= número mínimo de especímenes; MNI= número mínimo de individuos; NA= no aplica; P= procesamiento; X= taxones procesados.
Análisis de las variables taxonómicas
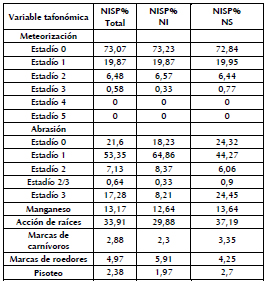
Como ya fue especificado en la metodología, para el análisis de las variables tafonómicas se consideró una muestra de 1389 especímenes para los taxones >1 kg (Tabla 3). Los restos faunísticos se caracterizan por una elevada presencia de especímenes con abrasión sedimentaria, principalmente en el estadío 1 y, en menor proporción, en los estadíos 2 y 3. El perfil de meteorización muestra una baja frecuencia de este agente, y predominan ampliamente los porcentajes de restos en el estadío 0, seguido por el 1 y, en menor medida, el 2 y el 3, en tanto que no se identificaron restos óseos asignables a los estadíos 4 y 5. La acción de raíces afectó considerablemente la superficie cortical de una importante cantidad de huesos. El manganeso fue identificado en menor porcentaje, pero de todas formas estaba bien representado en la muestra. Las marcas de carnívoros, roedores y pisoteo son escasas. En líneas generales, no hubo diferencias significativas entre los dos conjuntos, con la excepción de la acción de raíces, que es mayor en los NS, y la abrasión sedimentaria, la cual fue ligeramente superior en los NI (Tabla 3).
Tabla 3. Variables tafonómicas para taxones de porte mediano y grande (expresadas como NISP%) para el conjunto total y para los NS y NI.
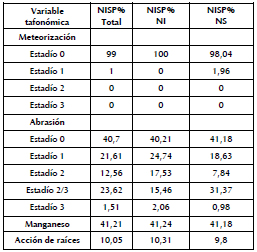
En el caso de los microvertebrados, se consideraron para el análisis de las variables tafonómicas aquellos restos que pudieron ser determinados a nivel anatómico y taxonómico. De esta manera, se analizó un total de 199 especímenes. Al igual que ocurre con los restos faunísticos de las especies de mayor tamaño, gran parte del conjunto óseo se caracteriza por la presencia de abrasión sedimentaria. Además, una gran cantidad de especímenes presenta tinción producida por manganeso, aunque la mayor parte de los restos óseos muestra un bajo porcentaje de su superficie afectada. Las raíces se observaron en baja proporción en el conjunto, y la meteorización sólo fue registrada en unos pocos especímenes de los NS (Tabla 4).
Tabla 4. Variables tafonómicas para microvertebrados (expresadas como NISP%) para el conjunto total y para los NS y NI.
Las modificaciones producidas por digestión fueron escasas (n= 15), por lo cual no se detallan en la Tabla 4, sino que se describen las tendencias generales. En el caso de los roedores, para molares e incisivos esta traza se presenta en baja frecuencia: ca. 10% corresponde a la categoría ligera, ca. 4,5% a la moderada y ca. 3% a la fuerte. Para el esqueleto poscraneal se observaron tendencias similares, correspondiendo ca. 8% a la categoría ligera y ca. 4% a la moderada. En las restantes especies de microvertebrados no se identificaron evidencias que sustenten la acción de predadores sobre estos. Finalmente, no se registraron diferencias significativas entre los NS y los NI para ninguna de las variables tafonómicas consideradas.
Medidas de abundancia y riqueza taxonómica y perfil de partes esqueletarias de Lama guanicoe
El Ntaxa fue de 11 para los NI y de 9 para los NS. El Índice de Artiodáctilos resultó en un valor de 0,73 para los NS, lo cual indica una estrategia más orientada a su explotación en comparación con los NI, para los cuales fue de 0,42, lo que muestra una dieta más orientada hacia los animales de menor porte. El Índice de Shannon-Wiener fue de 1,6 para los NS y de 1,9 para los NI, lo cual indica un poco más de heterogeneidad para la parte inferior del depósito.
El perfil de partes esqueletarias de Lama guanicoe fue construido para ambos niveles por separado. A continuación, se presentan los resultados de estos en las Tablas 5 y 6.
Tabla 5. Perfil de partes esqueletarias de Lama guanicoe para los NI.
Referencias: Indet= lateralidad no determinada; Px= proximal; D= diáfisis; Dt= distal.
Tabla 6. Perfil de partes esqueletarias de Lama guanicoe para los NS.

Referencias: Indet.= lateralidad no determinada; Px= proximal; D= diáfisis; Dt= distal.
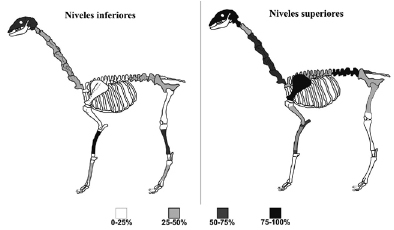
Como se observa en la Figura 5, en general hay una predominancia del esqueleto axial por sobre el apendicular, así como una baja representación de las costillas. Los valores más altos de MAU% para los NS son para las vértebras lumbares, seguidas por el cráneo con la vértebra atlas, la escápula, el sacro y las vértebras cervicales. Los miembros delanteros y traseros, así como el resto de las vértebras, tienen una representación media. Por último, los metapodios, huesos del autopodio y costillas se destacan por su baja representación (Tabla 5, Figura 5). Para los NI, la diáfisis del radiocúbito es la porción ósea más representada, seguida por el cráneo y la diáfisis de la tibia. El MAU% para los huesos largos es variable, dado que presenta valores altos, medios y bajos. Finalmente, los metapodios y las vértebras están escasamente representados, al igual que las costillas (Tabla 6, Figura 5).
Figura 5. MAU% para los restos óseos de Lama guanicoe en los NS y los NI.
Los perfiles de partes esqueletarias fueron correlacionados con el Índice de Utilidad Económica de Carne calculado por Borrero (1990). Para este análisis se incluyeron todos los restos óseos de Lama guanicoe, incluyendo los instrumentos óseos manufacturados sobre esta especie. Para los NI la correlación fue positiva y significativa (p= 0,040; Rs= 0,390) en tanto que para los NS resultó positiva y no significativa (p= 0,114; Rs= 0,305).
Evidencias de procesamiento
Los taxones que presentan evidencias de procesamiento antrópico son Canidae, Felidae, Rhea americana, Lama guanicoe, Tinamidae, Zaedyus pichiy, Ozotoceros bezoarticus, Lagostomus maximus, Conepatus sp., Chaetophractus villosus y Eutatus seguini. La información correspondiente al tipo de procesamiento registrado es resumida en la Tabla 7, considerando el NISP para cada uno de los taxones de los NI y los NS. Estos datos no se discuten en este trabajo en profundidad, ya que esto excedería su objetivo. Sin embargo, se describe esta información para Eutatus seguini, dado que son escasos los sitios en los cuales se ha identificado su aprovechamiento. En cuanto al procesamiento antrópico en la mayoría de los taxones, las tendencias entre los NS y los NI son similares al ser comparadas en términos de porcentajes.
Tabla 7. Taxones con evidencias de procesamiento expresadas en NISP para los NI y los NS.
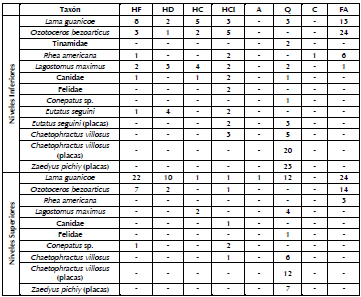
Referencias: HF= huellas de fileteo, HD= huellas de desarticulación, HC= huellas de corte, HCI= huellas de corte indeterminadas, A= ahumado, Q= quemado, C= calcinado, FA= fractura antrópica.
Se determinaron cinco instrumentos óseos, cuatro de ellos formatizados sobre huesos largos de Lama guanicoe y uno sobre un hueso de Artiodactyla (probablemente perteneciente también a Lama guanicoe). De estos, cuatro provienen de los NI, mientras que sólo uno fue recuperado de los NS. La funcionalidad de estos artefactos fue discutida por Álvarez (2012) y está aún en proceso de análisis (Figura 6).
Figura 6. Instrumentos óseos del sitio PO4 manufacturados sobre huesos largos de Lama guanicoe y Artiodactyla.
En los NI del sitio se identificaron 14 elementos óseos pertenecientes al armadillo extinguido Eutatus seguini. Estos son las hemipelvis, el astrágalo, el radio, los cúbitos, el húmero, un metacarpo, una vértebra caudal, una vértebra lumbar, la primera y segunda falange y dos tibias derechas, a través de las cuales se determinó un número mínimo de dos individuos (Figura 7a). De los 14 elementos, siete presentan huellas de corte. También se identificaron 38 placas óseas de este armadillo, tres de ellas quemadas y dos con huellas de corte en su lado ventral (Figura 7b). Estos restos, en conjunto, indican actividades de desarticulación, separación de la coraza, fileteo y consumo.

Figura 7. a) Elementos óseos de Eutatus seguini del sitio PO4; b) Placas de Eutatus seguini con huellas de corte.
En el caso de los microvertebrados, es necesario considerar que el procesamiento antrópico de estas especies puede no dejar trazas en los restos óseos, dado que su reducido tamaño permite su manipulación y procesamiento sin la necesidad de utilizar una tecnología específica (Stahl 1996; Kligmann et al. 1999). Más allá de esto, ninguno de los restos faunísticos de esta categoría presenta evidencias de explotación humana (i.e., huellas de corte, alteración térmica, fracturas antrópicas).
Huesos provenientes del rasgo
Los huesos provenientes del rasgo interpretado como un pozo de agua, incluidos en los NI, fueron también considerados como un conjunto separado. El objetivo de esta división fue evaluar si estos apoyaban o no la funcionalidad propuesta. Esta submuestra consistió en 281 especímenes, 211 correspondientes a taxones >1 kg y 70 a microvertebrados. Para los primeros, en general, las tendencias tafonómicas fueron las mismas que en el resto del conjunto óseo, con la excepción de un aumento en la frecuencia de especímenes con abrasión (sólo ca. 10% en el estadío 0). Por otro lado, dos tipos de modificaciones fueron identificadas exclusivamente en los especímenes provenientes del rasgo. Éstas son la presencia de una textura porosa en los huesos, acompañada por abrasión intensa (ca. 15%), así como de manchas negras (diferentes de las de manganeso), en forma de concreción sobre la superficie ósea de algunos huesos (ca. 9%). Ambas variables sólo fueron registradas en huesos localizados a una profundidad mayor a los 5 m y probablemente estén relacionadas a la presencia de agua. A la misma profundidad, pero fuera del rasgo, los restos óseos recuperados fueron una falange de la Familia Macrauchenidae, huesos dérmicos de megamamíferos correspondientes a Tardigrada (n= 4) y unos pocos elementos de Lama guanicoe (n= 4) y Dolichotis patagonum (n= 5). Estos especímenes (N= 14) estaban en sedimentos de la Formación Luján y no se encontraban asociados a los materiales arqueológicos. Si bien presentaban una abrasión avanzada, no mostraron evidencias de manganeso, cambios de coloración ni una textura diferente.
En cuanto al conjunto de microvertebrados, se destaca la alta frecuencia de restos de anuros (ca. 71,5%) en relación con los demás taxones presentes. A excepción del Orden Rodentia (ca. 17%), representado únicamente por incisivos aislados, el resto de los taxones muestra porcentajes menores al 6%. En cuanto a las variables tafonómicas, se observan tendencias similares a las del conjunto óseo total.
DISCUSIÓN
Tafonomía y procesos de formación del sitio
El análisis de las variables tafonómicas indicó la presencia de abrasión y manganeso, ambas relacionadas con un ambiente anegado o con presencia fluctuante de agua (López González et al. 2006), lo cual coincide con los resultados de los estudios geoarqueológicos antes mencionados. En el depósito arqueológico (ca. 8900 a 4600 años AP), las dinámicas ambientales estuvieron vinculadas a bordes de lagunas y encharcamientos. El agua habría actuado con posterioridad al procesamiento y transporte de las partes esqueletarias por parte de los grupos humanos, actividades que se habrían desarrollado sobre superficies de estabilización (Horizontes "A" de suelo). La acción hídrica habría ingresado al lugar, aunque con una baja energía, generando los perfiles de abrasión registrados y a la vez protegiendo a los restos óseos de una meteorización significativa. Estas fluctuaciones en los niveles del agua habrían llevado a la reexposición de algunos de los huesos, produciendo una meteorización más avanzada en determinados elementos. Posteriormente, se habría dado el entierro de estos, incorporando los materiales arqueológicos, así como otros de origen natural, representados principalmente por restos de microvertebrados. En el caso de estos últimos, la presencia de trazas leves de digestión sobre un escaso número de restos indicaría la participación de un predador de acción destructiva ligera (i.e., Tyto alba). Esto a su vez podría explicar la identificación de elementos atribuibles a taxones frecuentes en la dieta de estos predadores, como son los ofidios y los anfibios. Sin embargo, no se descarta que algunas especies hayan ingresado al sitio por otras causas, en particular aquellas vinculadas a hábitos fosoriales (i.e., Ctenomys sp.). Es probable que la dinámica ambiental mencionada, en conjunto con distintos agentes y procesos tafonómicos, se haya repetido a lo largo de toda la secuencia, siempre ligada a la acción hídrica de baja energía, en alternancia con eventos pedogenéticos.
En relación con los huesos provenientes del rasgo interpretado como un pozo de agua, estos escasos elementos presentan evidencias de agentes y procesos similares a las del resto del conjunto, pero habrían experimentado alteraciones diagenéticas más severas que afectaron su preservación, al modificar su textura haciéndola más porosa. Esto, en conjunto con la presencia de tinción y manganeso, apoya la idea de que dichos elementos fueron afectados por la presencia de agua, debido a las fluctuaciones de la napa freática una vez enterrados. Los elementos de microvertebrados provenientes de este rasgo, en particular los anuros, no muestran evidencias de digestión y su presencia podría estar vinculada al ingreso al pozo en momentos en que éste se encontraba abierto y con agua en su base. En este sentido, está representada casi la totalidad de los restos óseos de los anuros, con excepción de los craneales. Los aspectos mencionados indicarían que la presencia de estos especímenes estaría vinculada a aspectos etoecológicos de estos animales (i.e., hibernación y reproducción), los cuales habrían habitado el rasgo, habiendo quedado finalmente entrampados en él.
Tendencias en la subsistencia
Los aspectos referidos a presencia, consumo de presas y caracterización de la subsistencia sólo pueden ser discutidos en bloques temporales amplios. Este hecho se ve agravado por la ausencia en el contexto de fechas taxón debido a los problemas diagenéticos que afectaron a las unidades anatómicas (i.e., escasa o nula preservación del colágeno). Por otro lado, el análisis de los conjuntos líticos no ha permitido hasta el momento identificar diferentes tendencias tecnológicas en relación con la división propuesta entre los NI y los NS (Barros 2012).
En los NI se observó una mayor diversidad faunística y se infirió una estrategia de subsistencia más diversificada en relación con los NS. En estos últimos, la diversidad de especies es un poco más baja que en los NI y la estrategia de subsistencia estuvo más orientada hacia la obtención de artiodáctilos. En líneas generales, el registro faunístico indica que la disponibilidad de especies habría sido similar a lo largo del tiempo. Las excepciones están relacionadas con procesos de extinción diferencial. Eutatus seguini ha sido registrado previamente en otros sitios de la región pampeana para la transición Pleistoceno-Holoceno (i.e., Cerro La China y Cueva Tixi; Flegenheimer 1986-1987; Mazzanti 1997). En el caso del sitio PO4, la posición contextual y estratigráfica de esta especie permite ubicarla temporalmente en el Holoceno temprano (entre ca. 8900 y 7700 años AP), lo que confirma su supervivencia hacia avanzado dicho período (Gutiérrez et al. 2010, 2011), como también ha sido propuesto para el sitio Laguna El Doce (Cornaglia Fernández y Ávila 2011). Además, una fecha taxón proveniente de Arroyo Seco 2 sustenta esta cronología para el área (D. Rafuse comunicación personal, 2012). En el caso de la especie Dusicyon avus, se extinguió en la región en tiempos posteriores a la ocupación del sitio, durante el Holoceno tardío (Prevosti et al. 2011). En este punto, es pertinente mencionar que Dusicyon avus no fue incluido en los análisis de diversidad e IA, dado que no presentaba evidencias de procesamiento. Por otro lado, los valores de diversidad calculados tanto con los especímenes de Eutatus seguini como sin considerarlos son más altos para NI.
En este sentido, los cambios en la amplitud de la dieta serían independientes de la disponibilidad de especies. A su vez, la correlación entre el perfil de partes esqueletarias de Lama guanicoe y el Índice de Utilidad de Carne para este taxón resultó no significativa para los NS. Esto indica que, si bien habría habido una selección de las partes de este ungulado, no estuvo tan marcadamente orientada a las partes de alta utilidad como en los NI. Una contribución importante respecto del espectro de especies consumidas es la que aporta el sitio al Holoceno temprano (ca. 10.000-6500 años AP). El conjunto faunístico de PO4 se suma a la evidencia obtenida de sitios como El Guanaco, Arroyo Seco 2, La Moderna, Campo Laborde, La Olla sitio 1 y 2, Cueva El Abra y Barrio Las Dunas (Bayón y Politis 1996; Politis y Gutiérrez 1998; Politis y Madrid 2001; Mazzanti 2003; Politis et al. 2003, 2004; Politis y Messineo 2008; Flegenheimer et al. 2010; Bayón et al. 2012). Al registro de especies explotadas propuesto para el lapso Pleistoceno tardío-Holoceno temprano (Gutiérrez y Martínez 2008: tabla 2) se suman cánidos, félidos, tinámidos y mefítidos. Teniendo en cuenta el modelo propuesto por estos autores, el registro de Paso Otero 4 apoya la propuesta de una economía regional generalizada para el Pleistoceno tardío- Holoceno temprano, pero esta escala temporal de análisis de los conjuntos arqueofaunísticos no sería la adecuada. En este sentido, dados los datos brindados por nuevos sitios como PO4, la propuesta de este bloque temporal para caracterizar la subsistencia sería demasiado amplia y podría homogeneizar contextos faunísticos, escondiendo variabilidad en lapsos críticos como la transición Pleistoceno-Holoceno. El ejemplo más cercano es el del sitio PO5 (10.450- 10.200 años AP), localizado a 1,2 km de PO4, en el cual la fauna representada está compuesta casi exclusivamente por especies extinguidas (Martínez et al. 2013). Ambos contextos faunísticos, PO4 y PO5, difieren notablemente entre sí. En síntesis, aunque las economías para el lapso Pleistoceno tardío-Holoceno puedan ser clasificadas como generalizadas, a la luz de la evidencia de los nuevos sitios sería más adecuado manejar dos bloques temporales: Pleistoceno tardío (ca. 12.500-10.000 años AP) y Holoceno temprano (ca. 10.000-6500 años AP).
Para el Holoceno medio (6500-3500 años AP) Martínez y Gutiérrez (2004) propusieron una economía regional especializada. Sin embargo, el registro de nuevos sitios excavados recientemente ha permitido ampliar y modificar el conocimiento de este período. Las evidencias provistas por Alfar, el sitio 1 de la localidad Paso Mayor, Las Brusquillas 4 y el sector 2 del sitio 1 de la localidad El Guanaco (Bayón et al. 2010; Bonomo y León 2010; Massigoge 2011; Frontini 2012) han permitido identificar para el Holoceno medio una diversidad taxonómica mucho mayor a la conocida previamente. En el caso de PO4, es necesario considerar que los NS incluyen también algunos niveles correspondientes al Holoceno temprano. Es decir que, si bien las tendencias generales corresponden al Holoceno medio, no se descarta que los cambios en las estrategias de los grupos humanos puedan quedar enmascarados por las decisiones metodológicas tomadas por los analistas. En líneas generales, el registro de los NS indica que, si bien la subsistencia está más orientada a los artiodáctilos que en momentos previos, se observa simultáneamente una importante diversidad faunística, a la vez que gran parte de las presas son de tamaño pequeño. Tanto los datos generados para este sitio como aquellos derivados de las investigaciones recientes de los contextos antes mencionados para este lapso sugieren que la subsistencia no habría tenido necesariamente un énfasis en la especialización sobre algún recurso y que existiría cierta diversificación. Este replanteo del modelo probablemente se deba a la inclusión de nuevos sitios con cronologías poco frecuentes para el área (i.e., ca. 5900-5700; Paso Mayor y Alfar), localizados en áreas costeras y en bordes de sistemas fluviales importantes.
CONCLUSIONES
La secuencia estratigráfica registrada para PO4 y las dataciones obtenidas de sedimentos provenientes de ella muestran una remarcable coherencia interna. Estos aspectos del sitio, sumados a las diferencias en la presencia de algunos taxones, permitieron aislar a través de la secuencia dos conjuntos faunísticos que fueron referidos a los NI y a los NS. A pesar de este esfuerzo analítico, es claro que el sitio presenta bajos grados de resolución e integridad (ver discusión en Bailey 2007). Es por esto que, si bien en el sitio se identificaron diversas actividades, no resulta posible estimar una funcionalidad específica para éste, como por ejemplo, si corresponde a un campamento residencial o logístico. Más allá de esto, el conjunto óseo del sitio PO4 se caracteriza por una gran diversidad faunística y de esta manera contribuye con nueva evidencia arqueológica para la discusión de diferentes aspectos: supervivencia de la fauna extinguida, estrategias de subsistencia y nuevos datos para el Holoceno medio, un período escasamente representado en la región pampeana.
Agradecimientos
Este trabajo se desarrolló en el marco del INCUAPA-CONICET (Facultad de Ciencias Sociales, Universidad Nacional del Centro de la Provincia de Buenos Aires) y fue financiado por National Geographic Society (Archaeology and ancient environment in the Pampean region of Argentina; Grant #7181- 01) y Agencia Nacional de Promoción Científica y Tecnológica (Arqueología y tafonomía actualística en la llanura interserrana y sectores periserranos adyacentes; PICT 08-814). Los autores agradecen a La Dulce Seguros de Granizo por su apoyo durante los trabajos de campo 2008-2010. Agradecemos a Cristian Kaufmann y Daniel Rafuse por la asistencia técnica. Finalmente, cuatro evaluadores contribuyeron a mejorar sustancialmente este trabajo a través de valiosos comentarios y sugerencias.
1 Los números de identificación del laboratorio y valores de ä13C fueron publicados en un trabajo previo (Gutiérrez et al. 2011) para los fechados de la columna estratigráfica, con excepción de una nueva muestra datada en 6668 ± 55 (H) y 6739 ± 48 (R), ä13C= -19,2 para ambas fracciones (AA87930). Los resultados de dataciones obtenidas para el rasgo son presentados en esta publicación y la información referida a estos es: AA87935: 6168 ± 43 (R) y 8735 ± 48 (H), ä13C= -18,2 y -19,5, respectivamente; AA87936: 6395 ± 79 (R) y 8075 ± 66 (H); ä13C= -18,1 y -19,2, respectivamente; AA87934: 8556 ± 65, ä13C=-26,1; AA87937: 8331 ± 65 (H) y 8760 ± 160 (R), ä13C=-20,2 y -21, respectivamente.
REFERENCIAS CITADAS
1. Álvarez, M. C. 2012 Análisis zooarqueológicos en el Sudeste de la región pampeana. Patrones de subsistencia durante el Holoceno tardío. Tesis Doctoral inédita. Facultad de Ciencias Sociales, Universidad Nacional del Centro de la Provincia de Buenos Aires, Buenos Aires. [ Links ]
2. Andrews, P. 1990 Owls, Caves and Fossils. The University of Chicago Press, Londres. [ Links ]
3. Bailey, G. 2007 Time perspectives, palimpsests and the archaeology of time. Journal of Anthropological Archaeology 26: 198-223. [ Links ]
4. Barros, M. P. 2012 La circulación y el modo de explotación de las rocas en el sitio Paso Otero 4 durante el Holoceno temprano y medio (partido de Necochea, provincia de Buenos Aires, Argentina). En Peuplement et modalités d'occupation de l'Amérique du sud: l'apport de la technologie lithique / Povoamento e modalidades de ocupação humana na América do Sul: a contribuição da tecnologia lítica, editado por E. Boëda, M. Farias y A. Lourdeau. @rchéo-éditions.com, Francia. En prensa. [ Links ]
5. Barros, M. P., G. Martínez y M. A. Gutiérrez 2011 Análisis de la tecnología lítica del sitio Paso Otero 4 (partido de Necochea, provincia de Buenos Aires, Argentina). El uso de las rocas durante el Holoceno temprano y medio en la sub-región Pampa Húmeda. Trabajo presentado en el XVI Congresso UISSP, Universidad Federal de Santa Catarina, Florianópolis, Brasil. MS. [ Links ]
6. Bayón, C. y G. Politis 1996 Estado actual de las investigaciones en el sitio Monte Hermoso 1, provincia de Buenos Aires. Arqueología 6: 83-116. [ Links ]
7. Bayón, C., A. Pupio, R. Frontini, R. Vecchi y C. Scabuzzo 2010 Localidad arqueológica Paso Mayor: nuevos estudios 40 años después. Intersecciones en Antropología 11: 115-128. [ Links ]
8. Bayón, C., R. Frontini y R. Vecchi 2012 Middle Holocene settlements on coastal dunes, southwest Buenos Aires Province, Argentina. Quaternary International 256: 54-61. [ Links ]
9. Behrensmeyer, A. K. 1978 Taphonomic and Ecologic Information from Bone Weathering. Paleobiology 4: 150-162. [ Links ]
10. Bennett, J. 1999 Thermal Alteration of Buried Bone. Journal of Archaeological Science 26: 1-8. [ Links ]
11. Binford, L. 1978 Nunamiut Ethnoarchaeology. Academic Press, Nueva York. [ Links ]
12. Binford, L. 1981 Bones: Ancient Men and Modern Myths. Academic Press, Nueva York. [ Links ]
13. Bonomo, M. y D. C. León 2010 Un contexto arqueológico en posición estratigráfica en los médanos litorales. El sitioAlfar (Pdo. de General Pueyrredón, Pcia. De Buenos Aires). En Mamul Mapü: pasado y presente desde la arqueología pampeana, tomo 2, editado por M. Berón, L. Luna, M. Bonomo, C. Montalvo, C. Aranda y M. Carrera Aizpitarte, pp. 29-45. Libros del Espinillo, Ayacucho. [ Links ]
14. Borrero, L. 1990 Fuego-Patagonia Bone assemblage and the Problem of Comunal Guanaco Hunting. En Hunters of the Recent Past, editado por L. Davis y B. Reeves, pp. 373-399. One World Archaeology. Unwin Hyman, Londres. [ Links ]
15. Broughton, J. 1994 Late Holocene resource intensification in the Sacramento Valley, California: The vertebrate evidence. Journal of Archaeological Science 21: 501-514. [ Links ]
16. Cornaglia Fernández, J. y D. Ávila 2011 Presencia de Eutatus seguini (mammalia, dasypodidae) y Dusicyon avus (mammalia, canidae) en el sitio arqueológico Laguna El Doce (departamento General López, provincia de Santa Fe). Revista del Museo de La Plata. Arqueología 12 (86): 67. [ Links ]
17. David, B. 1990 How was this Bone Burnt? En Problem Solving in Taphonomy: Archaeological and Paleontological Studies from Europe, Africa and Oceania, vol. 2, editado por S. Solomon, I. Davidson y D. Watson, pp. 65-79. Tempus, The University of Quuensland, Queensland. [ Links ]
18. Fernández-Jalvo, Y. y P. Andrews 1992 Small Mammal Taphonomy of Gran Dolina, Atapuerca (Burgos), Spain. Journal of Archaeological Science 19: 407-428. [ Links ]
19. Fidalgo, F., F. O. de Francesco y U. Colado 1973 Geología superficial en las hojas Castelli, J.M. Cobo y Monasterio (Pcia. De Buenos Aires). Actas del V Congreso de Geología Argentina: 27-39. [ Links ]
20. Flegenheimer, N. 1986-1987 Excavaciones en el sitio 3 de la Localidad Cerro La China (Provincia de Buenos Aires). Relaciones de la Sociedad Argentina de Antropología XVII: 7-28. [ Links ]
21. Flegenheimer, N., C. Bayón, C. Scabuzzo, N. Mazzia, R. Vecchi, C. Weitzel, R. Frontini y M. Colombo 2010 Early-Holocene Human Skeletal Remains from the Argentinean Pampas. Current Research in the Pleistocene 27: 10-12. [ Links ]
22. Frontini, R. 2012 El aprovechamiento de animales en valles fluviales y lagunas del sur bonaerense durante el Holoceno. Tesis Doctoral inédita. Facultad de Filosofía y Letras, Universidad de Buenos Aires, Buenos Aires. [ Links ]
23. Grayson, D. K. 1984 Quantitative Zooarchaeology: Topics in the Analysis of Archaeological Faunas. Academic Press, Orlando. [ Links ]
24. Grayson, D. K. 1991 Alpine faunas from the White Mountains, California: adaptative change in the Late Prehistoric Great Basin. Journal of Archaeological Science 18: 438-506. [ Links ]
25. Gutiérrez, M. A. y C. Kaufmann 2007 Criteria for the Identification of Formation Processes in Guanaco (Lama guanicoe) Bone Assemblages in Fluvial-Lacustrine Environnments. Journal of Taphonomy 5 (4): 151-176. [ Links ]
26. Gutiérrez, M. A. y G. Martínez 2008 Trends in the faunal human exploitation during the Late Pleistocene and Early Holocene in the Pampean region (Argentina). Quaternary International 191: 53-68. [ Links ]
27. Gutiérrez, M. A. y G. Martínez 2010 Origen, funcionalidad y cronología de un rasgo arqueológico registrado en sedimentos de la Formación Luján: sitio Paso Otero 4 (curso medio del río Quequén Grande, Pdo. de Necochea). Trabajo presentado en el XVII Congreso Nacional de Arqueología Argentina, Mendoza. MS. [ Links ]
28. Gutiérrez, M. A., G. Martínez y C. Nielsen-Marsh 2001 Alteración Diagenética y Preservación Diferencial de los Conjuntos Óseos de la Localidad Arqueológica Paso Otero (Provincia de Buenos Aires, Argentina). Estudios Geológicos 56: 291-299. [ Links ]
29. Gutiérrez, M. A., G. Martínez, H. Luchsinger, M. C. Álvarez y M. P. Barros 2010 Investigaciones arqueológicas y geoarqueológicas preliminares en el sitio Paso Otero 4 (Partido de Necochea). En Mamul Mapü: pasado y presente desde la arqueología pampeana, editado por M. Berón, L. Luna, M. Bonomo, C. Montalvo, C. Aranda y M. Carrera Aizpitarte, pp. 69-84. Libros del Espinillo, Ayacucho. [ Links ]
30. Gutiérrez, M. A., G. Martínez, H. Luchsinger, S. Grill, A. Zucol, G. Hassan, M. P. Barros, C. Kaufmann y M. C. Álvarez 2011 Paleoenvironments in the Paso Otero locality during late Pleistocene-Holocene (pampean region, Argentina): an interdisciplinary approach. Quaternary International 245: 37-47. [ Links ]
31. Haynes, G. 1980 Evidence of Carnivore Gnawing on Pleistocene and Recent Mammalian Bones. Paleobiology 6: 341-351. [ Links ]
32. Johnson, E. 1985 Current Developments in Bone Technology. Advances in Archaeological Method and Theory 8: 157-235. [ Links ]
33. Johnson, E., G. Politis, G. Martínez, W. T. Hartwell, M. A. Gutiérrez y H. Haas 1998 The radiocarbon chronology of Paso Otero 1 in the Pampean Region of Argentina. Quaternary of South America and Antarctic Peninsula 11: 15-25. [ Links ]
34. Johnson, E., V. T. Holliday, G. Martínez, M. A. Gutiérrez y G. Politis 2012 Geochronology and Landscape Development in the Middle Río Quequén Grande at the Paso Otero Locality, Pampa Interserrana, Argentina. Geoarchaeology: An International Journal 27: 300-323. [ Links ]
35. Klein, R. y K. Cruz-Uribe 1984 The Analyses of Animal Bones from Archaeological Sites. The University of Chicago Press, Chicago. [ Links ]
36. Kligmann, D. M., C. Sesé y J. Barbadillo 1999 Análisis tafonómico de la fauna de microvertebrados del Alero 12 (Puna Meridional Catamarqueña Argentina) y sus implicancias para el comportamiento humano. Arqueología 9: 9-48. [ Links ]
37. López-González, F., A. Grandal-d`Ánglade y J. R. Vidal- Romaní 2006 Deciphering bone depositional sequences in caves through the study of manganese coatings. Journal of Archaeological Science 33: 707-717. [ Links ]
38. Lyman, R. 1994 Vertebrate Taphonomy. Cambridge University Press, Cambridge. [ Links ]
39. Lyman, R. 2008 Quantitative Paleozoology. Cambridge University Press, Cambridge. [ Links ]
40. Martínez, G. 2001 "Fish-tail" projectile points and megamammals in the Pampean region of Argentina. Antiquity 75 (289): 523-528. [ Links ]
41. Martínez, G. 2002-2004 Superficies de estabilización del paisaje (horizontes "A" de suelos enterrados) y el registro arqueológico de la localidad Paso Otero (Río Quequén Grande, Pdo. de Necochea). Arqueología 12: 179-199. [ Links ]
42. Martínez, G. 2006 Arqueología del curso medio del río Quequén Grande: estado actual y aportes a la arqueología de la región pampeana. Relaciones de la Sociedad Argentina de Antropología XXXI: 249-275. [ Links ]
43. Martínez, G. y M. A. Gutiérrez 2004 Tendencias en la explotación humana de la fauna durante el Pleistoceno Final y Holoceno en la Región Pampeana (Argentina). En Zooarchaeology of South America, editado por G. Mengoni Goñalons, pp. 81-98. BAR International Series 1298. Archaeopress, Oxford. [ Links ]
44. Martínez, G. y M. A. Gutiérrez 2011a Paso Otero 5: A summary of the interdisciplinary lines of evidence for reconstructing early human occupation and paleoenvironment in the Pampean region, Argentina. En Peuplements et Préhistoire de l'Amérique, editado por D. Vialou, pp. 271-286. Muséum National d'Histoire Naturelle, Département de Préhistoire, París. [ Links ]
45. Martínez, G. y M. A. Gutiérrez 2011b An Early Holocene water well in the Pampas Plains (Pampean Region, Argentina)? Resúmenes del 76th Annual Meeting de la Society for American Archaeology: 209. Sacramento, California. [ Links ]
46. Martínez, G., M. A. Gutiérrez y E. P. Tonni 2013 Paleoenvironments and faunal extinctions: the analysis of the archaeological assemblages at the Paso Otero locality (Argentina) during the Late Pleistoceneearly Holocene. Quaternary International 299: 53-63. [ Links ]
47. Massigoge, A. 2011 Nuevas evidencias arqueológicas del Holoceno medio y tardío del área Interserrana: localidad arqueológica Las Brusquillas (partido de San Cayetano, provincia de Buenos Aires). Resúmenes del VI Congreso de Arqueología de la Región Pampeana Argentina: 117. Facultad de Ciencias Naturales y Museo, Universidad Nacional de La Plata, La Plata. [ Links ]
48. Mazzanti, D. 1997 Excavaciones arqueológicas en el sitio Cueva Tixi, Buenos Aires, Argentina. Latin American Antiquity 8 (1): 55-62. [ Links ]
49. Mazzanti, D. 2003 Human settlements in caves and rockshelters during the Pleistocene-Holocene transition in the Eastern Tandilia Range, Pampean Region, Argentina. En Where the South Winds Blow. Ancient Evidences for Paleo South Americans, editado por L. Miotti, M. Salemme y N. Flegenheimer, pp. 57-61. Center for the Studies of the First Americans (CSFA) and Texas A&M University Press, College Station, Texas. [ Links ]
50. Miotti, L. y M. Salemme 1999 Biodiversity, taxonomic richness and specialists generalists during Late Pleistocene/Early Holocene times in Pampa and Patagonia (Argentina, Southern South America). Quaternary International 53-54: 53-68. [ Links ]
51. Olsen, S. y P. Shipman 1988 Surface Modification on Bone: Trampling Versus Butchery. Journal of Archaeological Science 15: 535-553. [ Links ]
52. Politis, G. y M. A. Gutiérrez 1998 Gliptodontes y cazadores-recolectores de la región pampeana (Argentina). Latin American Antiquity 9 (2): 111-134. [ Links ]
53. Politis, G. y P. Madrid 2001 Arqueología pampeana: estado actual y perspectivas. En Historia Argentina Prehispánica, Tomo II, editado por E. Berberian y A. Nielsen, pp. 737-814. Brujas, Córdoba. [ Links ]
54. Politis, G. y P. Messineo 2008 The Campo Laborde site: new evidence for the Holocene survival of Pleistocene megafauna in the Argentine Pampas. Quaternary International 191: 98-114. [ Links ]
55. Politis, G. y M. Salemme 1990 Pre-hispanic mammal exploitation and hunting strategies in the eastern Pampas sub-region of Argentina. En Hunters of the Recent Past, editado por L. Davis y B. Reeves, pp. 353-372. Unwin Hyman, Londres. [ Links ]
56. Politis, G., M. Bonomo y L. Prates 2003 Territorio y movilidad entre la costa atlántica y el interior de la región Pampeana. Estudios Ibero- Americanos XXIX (1): 11-35. [ Links ]
57. Politis, G., P. Messineo y C. Kaufmann 2004 El poblamiento temprano de las llanuras pampeanas de Argentina y Uruguay. Complutum 15: 207-224. [ Links ]
58. Prevosti, F. J., F. Santiago, L. Prates y M. C. Salemme 2011 Constraining the time of extinction of the South American fox Dusicyon avus (Carnivora, Canidae) during the late Holocene. Quaternary International 245 (2): 209-217. [ Links ]
59. Quintana, C. A. y D. L. Mazzanti 2001 Selección y Aprovechamiento de Recursos Faunísticos. En Cueva Tixi: Cazadores y recolectores de las Sierras de Tandilia Oriental, I Geología, Paleontología y Zooarqueología, editado por D. L. Mazzanti y C. A. Quintana, pp. 181-209. Publicación Especial 1. Laboratorio de Arqueología, Universidad Nacional de Mar del Plata, Mar del Plata. [ Links ]
60. Shipman, P. 1981a Life History of a Fossil. Harvard University Press, Cambridge. [ Links ]
61. Shipman, P. 1981b Applications of Scanning Electron Microscopy to Taphonomic Problems. Annals of the New York Academy of Sciences 276: 357-386. [ Links ]
62. Stahl, P. W. 1996 The recovery and Interpretation of Microvertebrate Bone Assemblages from Archaeological Contexts. Journal of Archaeological Method and Theory 3: 31-75. [ Links ]
63. Tambussi, C. P. y C. Acosta Hospitaleche 2002 Reidos (Aves) cuaternarios de Argentina: inferencias paleoambientales. Ameghiniana 39 (1): 95- 102. [ Links ]
64. Zárate, M., M. Espinosa y L. Ferrero 1998 Paleoenvironmental implications of the Holocene diatomite, Pampa Interserrana, Argentina. Quaternary of South America and Antarctic Peninsula 11: 135-152. [ Links ]
65. Zárate, M., R. Kemp, M. Espinosa y L. Ferrero 2000 Pedosedimentary and palaeoenvironmental significance of a Holocene alluvial sequence in the Southern Pampas, Argentina. The Holocene 10 (4): 481-488. [ Links ]

