










Serviços Personalizados
Journal

Artigo
 Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares em
SciELO
Similares em
SciELO
Compartilhar

 Permalink
PermalinkIntersecciones en antropología
versão On-line ISSN 1850-373X
Intersecciones antropol. vol.15 no.1 Olavarría jun. 2014
ARTÍCULOS
Consumo de camélidos durante el período de Desarrollos Regionales: una perspectiva local
Juan Maryañski
Juan Maryañski. Consejo Nacional de Investigaciones Científicas y Técnicas (CONICET). Instituto Nacional de Antropología y Pensamiento Latinoamericano (INAPL). 3 de Febrero 1378 (C1426BJN), Buenos Aires, Argentina. E-mail: juanmki@gmail.com
Recibido 11 de mayo 2012.
Aceptado 26 de diciembre 2012
RESUMEN
En este trabajo se presenta un análisis comparativo de conjuntos zooarqueológicos procedentes de distintos contextos funcionales contemporáneos del sitio Bilcapara (departamento de Potosí, Bolivia), un asentamiento residencial conglomerado del período de Desarrollos Regionales (900-1450 DC). Se observan contrastes en la representación taxonómica y anatómica de camélidos, relacionados con la circulación de recursos silvestres y las trayectorias posmatanza de algunas unidades anatómicas. Se argumenta que estos patrones dependen de las escalas de consumo y contextos de uso de la fauna involucrados en cada caso. Asimismo, se sostiene que los estudios zooarqueológicos a múltiples escalas, que contemplen la variación local en sitios internamente diferenciados a nivel funcional, son una vía complementaria para ampliar el conocimiento sobre las pautas económicas de las sociedades prehispánicas del Sur Andino.
Palabras clave: Zooarqueología de camélidos; Escalas de consumo; Economía del Período de Desarrollos Regionales.
ABSTRACT
Camelid consumption during regional development period: a local perspective. This paper presents a comparative analysis of zooarchaeological assemblages from different contemporary functional contexts of Bilcapara site (Potosí Department, Bolivia), a Regional Developments Period (900-1450 DC) aggregated residential settlement. Contrasts in taxonomic and anatomic camelid representation, related to wild resources flowing and post-slaughter trajectories of some anatomical units are observed. It is argued that this patterns depends on consuming scales and contexts of animal use. Also, it is argued that zooarchaeological analysis at multiple-scales, wich look at local variation in internaly differenciated sites, are a complementary way to broaden our knowledge about the economic patterns of South Andean prehispanic societies.
Keywords: Camelid zooarchaeology; Consumption scales; Regional Developments Period economy.
INTRODUCCIÓN
La ampliación de los estudios zooarqueológicos sistemáticos durante las últimas décadas ha contribuido a esclarecer el panorama en torno al uso de recursos faunísticos entre las sociedades que habitaron los valles y quebradas del surandinas durante el período Tardío o de Desarrollos Regionales (900-1450 DC, en adelante PDR). De esta manera, y aunque todavía permanecen importantes vacíos de información, fue posible trascender la noción vaga y general de "economía agropastoril" invocada en los primeros trabajos que se interrogaron sobre esta problemática, en pos de investigaciones empíricamente sustentadas (Madero 1993; Izeta 2008; Izeta et al. 2009b).
Estos estudios poseen particular interés dado que se insertan en la discusión de problemáticas de mayor alcance, relacionadas con los procesos de cambio social que, al margen de las diversas interpretaciones propuestas sobre su naturaleza, caracterizan los primeros siglos del segundo milenio DC y definen al PDR como segmento temporal específico. En este sentido, la mayoría de los esfuerzos se han orientado a evaluar principalmente: a) las articulaciones entre caza y pastoreo; b) la utilización de los animales desde la perspectiva de los múltiples recursos que ofrecieron; y c) las especificidades de estas prácticas en estas sociedades demográficamente importantes y de fuerte base agrícola -en contraste con la situación que caracterizó a las distintas porciones de la Puna- (Madero 1993; Izeta 2008; Izeta et al. 2009b; Mercolli 2011, entre otros). Estos temas han sido normalmente abordados a partir del análisis de conjuntos procedentes de asentamientos residenciales, a través de la evaluación de tendencias de largo plazo en el registro zooarqueológico y considerando perspectivas espaciales amplias -i.e., región o microrregión, independientemente de que la referencia empírica provenga de uno o varios sitios o depósitos por unidad temporal-.
Sin embargo, otras dimensiones de variabilidad en el registro zooarqueológico de los conjuntos tardíos de valles y quebradas han recibido menor atención. Dos características centrales de los asentamientos residenciales de esta época, sobre todo de aquellos posteriores al ca. 1250 DC, son la alta concentración edilicia y la importante heterogeneidad interna que exhiben desde el punto de vista arquitectónico y funcional, dada por la presencia de vías de circulación, complejos domésticos, espacios públicos o "plazas" y áreas de descarte definidas (Nielsen 2007; Raffino 2007). Esta heterogeneidad constituye una oportunidad inmejorable para estudiar el registro zooarqueológico a múltiples escalas, en la medida en que los conjuntos recuperados en estos espacios son producto del descarte resultante de las actividades en ellos realizadas (Mengoni Goñalons 2010). Dada esta situación, sostenemos que el análisis de la variabilidad arqueofaunística a escala local es una vía adecuada para enriquecer y/o complementar las hipótesis surgidas de estudios regionalmente orientados. Por escala local hacemos referencia al nivel del sitio arqueológico cuyo pautamiento arquitectónico y artefactos allí descartados indican recurrentemente su uso como locus residencial (ver Raffino 2007 para una síntesis casuística). El abordaje de la variabilidad en esta escala, entonces, depende de la definición de unidades espaciales intrasitio que involucren potencialmente distintos rangos de actividades.
En este marco, el propósito de este trabajo es dar cuenta del uso diferencial de camélidos, desde el punto de vista taxonómico y anatómico en contextos contemporáneos de diferente funcionalidad. Para esto se presentan los resultados de un análisis comparativo llevado a cabo sobre dos conjuntos recuperados en sendos basurales procedentes del sitio Bilcapara (departamento de Potosí, Bolivia). Las muestras proceden, respectivamente, de un complejo doméstico y de un contexto público. El sitio se ubica en la Cuenca Media del río Grande de San Juan (en adelante, CMRGSJ) y consiste en un asentamiento residencial conglomerado de "trazado concentrado" (sensu Raffino 2007) característico de la fase tardía -posterior al ca. 1250 DC (Nielsen 2007)- del PDR. A continuación se presenta en detalle la problemática planteada y el marco empleado para abordarla. Luego se caracterizan los contextos a analizar de Bilcapara y la región en que el sitio se inserta. Finalmente, se exponen los resultados alcanzados, y su posterior discusión.
ZOOARQUEOLOGÍA DE ASENTAMIENTOS TARDÍOS
Las investigaciones realizadas hasta el momento en sitios prehispánicos tardíos de valles y quebradas se han ocupado principalmente de las estrategias de explotación de animales y sus transformaciones en relación con los procesos de cambio social y deterioro climático observados con posterioridad al 1000 DC. Un elemento común del registro faunístico de este segmento temporal es el predominio de camélidos y, dentro de este taxón, la alta frecuencia de individuos adultos y la presencia variable de especies domésticas y silvestres (Madero 1993; Izeta 2008; Izeta et. al 2009b). El componente pastoril y cazador en la economía de estas sociedades fue articulado posiblemente a través de distintas estrategias de aprovechamiento del heterogéneo ambiente que caracteriza a los valles adyacentes al altiplano puneño.
Sobre la base de esta información se han discutido hipótesis en torno a la ocurrencia de procesos de diversificación, especialización, reorientación de las estrategias productivas y de aumento/detrimento del componente cazador. Independientemente de las múltiples hipótesis empleadas para discutir casos particulares, es destacable la variabilidad observada en el registro zooarqueológico disponible, más allá del recurrente predomino de camélidos en los conjuntos (Izeta 2008; Izeta et al. 2009b). Las causas de esta variabilidad pueden relacionarse con: a) el desigual volumen de información producido en las distintas regiones; b) la existencia de diferentes métodos para generar los datos primarios (i.e., osteometría, perfiles de edad, criterios de cuantificación); y c) la presencia de contrastes situacionales en el uso y explotación de recursos animales, las cuales se reflejarían en las características de los conjuntos. Mientras los primeros dos puntos aguardan la ampliación de las investigaciones regionales para ser resueltos, el último de ellos pone de relieve la necesidad de indagar el registro zooarqueológico a través de múltiples escalas y considerando la variabilidad presente en los distintos contextos de descarte involucrados.
El análisis de esta dimensión de variabilidad desde el punto de vista regional permitió documentar, para momentos prehispánicos tardíos, la existencia de lugares de extracción/producción de recursos faunísticos diferenciados de las localidades de residencia permanente (e.g., Madero 1992; Olivera y Grant 2009; Nielsen et al. 2010a). Menos atención recibió, sin embargo, el estudio de la variabilidad interconjunto a escala local, la cual depende en gran medida de la consideración de los diversos contextos de uso de la fauna (aunque ver entre otros Rodríguez Loredo 1997- 1998; Mercolli y Seldes 2007; Dantas 2010; Lema et al. 2011).
Un primer abordaje a esta problemática debe atender a las diferencias presentes en la organización interna de los asentamientos y su correlato en términos de prácticas de consumo y expectativas de descarte (Mengoni Goñalons 2010). En el marco del estudio de sociedades sedentarias y "políticamente complejas", este nivel de análisis es frecuentemente dirigido a evaluar el acceso diferencial de especies, grupos etarios o unidades anatómicas por parte de sectores sociales de distinta jerarquía (Miller y Burger 1995; Jackson y Scott 2003; DeFrance 2009). Sin embargo, esta línea de análisis difícilmente pueda resultar adecuada para el caso de muchos de los grupos que habitaron los valles de la cordillera oriental durante el PDR. La "orientación corporativa" que parece haber caracterizado a estas formaciones sociales implica la apropiación colectiva de recursos como uno de los ejes fundamentales de las estrategias políticas del período (Nielsen 2006). Esto no excluye eventuales diferencias en la distribución de recursos faunísticos entre unidades domésticas, pero advierte sobre la necesidad de evaluar otras causas que generan variación en la circulación y descarte de los mismos e inciden en el pautamiento espacial del registro zooarqueológico.
Un patrón recurrente en varios sitios residenciales tardíos es el contraste entre los espacios domésticos y aquellos espacios destinados a actividades públicas (i.e., "plazas", Nielsen 2006; Raffino 2007), los cuales presentan rasgos arquitectónicos y pautas de acumulación de materiales claramente diferenciadas. En este sentido, al margen del carácter de las actividades llevadas a cabo en uno y otro tipo de contexto, las mismas se distinguen por las escalas sociales de consumo involucradas en cada caso. Distintos autores han discutido los indicadores materiales de los eventos de consumo público o comunal, generalmente asociados a actividades ceremoniales, con el fin de evaluar las posibilidades de su reconocimiento arqueológico (Dietler y Hayden 2001; Miracle 2002). Si bien consideramos que la identificación de este tipo de prácticas debe atender a las situaciones culturalmente específicas de cada caso (Nielsen 2006), postulamos como hipótesis de trabajo que las escalas de consumo inciden en las decisiones sobre el tratamiento posmatanza de camélidos y, por lo tanto, en los patrones de ingreso y descarte de unidades anatómicas arqueológicamente observables en cada contexto (Miracle 2002). Desde un punto de vista comparativo es esperable entonces, en los espacios públicos, la acumulación de desechos óseos como producto del consumo inmediato de importantes volúmenes de alimento, con una consecuente menor selección antrópica en los perfiles anatómicos de camélidos. Por el contrario, las acumulaciones de desechos en espacios domésticos, hipotéticamente vinculadas al consumo familiar, serían el resultado de trayectorias más prolongadas desde la matanza hasta el descarte final, pudiendo involucrar situaciones de consumo diferido vía almacenamiento, reparto entre unidades domésticas o limpieza de desechos de los espacios intramuros, entre otras. Esta diversidad de situaciones plantea como expectativa una mayor selección en los perfiles anatómicos para los contextos domésticos, la cual será evaluada en este trabajo a través de diferentes marcos de referencia.
Es importante mencionar sin embargo que una comprensión detallada de la variabilidad zooarqueológica intrasitio, y en particular en la configuración material de distintos tipos de contextos, precisa la incorporación de un mayor número de muestras que permitan aislar los factores responsables de tales contrastes. En este trabajo presentamos un primer análisis de esta variabilidad, en la medida en que es un paso necesario para la definición las unidades de análisis válidas para el estudio comparativo del uso y circulación de recursos faunísticos en sitios tardíos. Entendemos que un análisis de estas características, basado en unidades espaciotemporales de resolución fina, puede contribuir a generar una imagen más completa sobre el papel que desempeñaron los distintos productos animales entre las sociedades prehispánicas de valles y quebradas.
Para llevar a cabo esta propuesta, nos centraremos en el estudio de la selección y descarte de camélido y de sus unidades anatómicas en los distintos conjuntos estudiados de Bilcapara. Previamente, presentamos una caracterización del área de estudio, del sitio en cuestión y de los contextos de procedencia de las muestras.
BILCAPARA Y SU MARCO GEOGRÁFICO
La CMRGSJ (Figura 1) se ubica en la vertiente oriental de los Andes, entre el norte de la actual provincia argentina de Jujuy y el extremo sudoeste del departamento boliviano de Potosí. Este espacio comprende el valle del río Grande de San Juan y una serie de quebradas tributarias altas, con cotas que oscilan entre los 3000 a 3600 msnm. Estas quebradas y el propio valle principal se encuentran interpuestas por el extremo norte de la Puna jujeña y los diversos cordones montañosos que componen la Cordillera de Chichas. Las investigaciones realizadas a la fecha permiten postular, para momentos prehispánicos tardíos, la presencia en la región de poblaciones sedentarias de magnitud, cuya alfarería predominante corresponde a la denominada "Cultura Yavi" (Krapovickas y Aleksandrowicz 1986) o "estilo alfarero yavi" (Ávila 2009), contrastante a la vez con aquélla recuperada en ámbitos próximos como el altiplano de Lípez, la cuenca del río Miraflores o la Quebrada de Humahuaca. La intensidad de ocupación durante momentos tardíos se manifiesta en la presencia de numerosos sitios residenciales a la vera de las quebradas más importantes (Krapovickas y Cigliano 1964; Nielsen et al. 2014, 2010b). Se trata principalmente de asentamientos conglomerados, emplazados en terrazas fluviales relativamente elevadas. Asimismo, en torno a estas localidades se han registrado extensos sectores agrícolas que indican un uso variable e intensivo de los espacios cultivables que pueden ser cronológicamente atribuidos a momentos prehispánicos tardíos.

Figura 1. CMRGSJ y ubicación de sitios mencionados en el texto.
Bilcapara, con aproximadamente 1,2 ha de extensión y emplazado a ca. 3450 msnm, es uno de los asentamientos más importantes documentados hasta el momento en el sector superior del río Grande de San Juan. Los trabajos realizados en el mismo y en zonas aledañas revelaron la presencia de áreas domésticas y de uso público, así como agrupamientos de bloques grabados, construcciones destinadas a la funebria/almacenamiento y estructuras agrícolas en las inmediaciones (Krapovickas y Cigliano 1964; Nielsen et al. 2014). Por otra parte, las investigaciones recientes confirmaron la alta frecuencia de cerámica yavi en el área, lo cual emparenta a Bilcapara, al menos desde su cultura material más conspicua, con los sitios ubicados en otros sectores de la CMRGSJ como el bolsón de Yavi o las quebradas de Talina y San Lorenzo (Nielsen et al. 2014). Dos fechas radiocarbónicas sitúan cronológicamente las ocupaciones del sitio alrededor del siglo XIV DC (Nielsen et al. 2014), por lo que se corresponde con la fase tardía (sensu Nielsen 2007) del PDR. Asimismo, la ausencia de artefactos diagnósticos de momentos previos (como alfarería Calahoyo) o de filiación Inka, así como de remodelaciones en la arquitectura del sitio, indican una ocupación relativamente acotada al lapso señalado (Nielsen et al. 2014). Esta situación es concordante con el panorama que ofrecen distintas regiones de la vertiente oriental surandina, consistente en un proceso de concentración poblacional en torno a los corredores principales, en asentamientos elevados y de difícil acceso, durante los siglos XIII y XIV.
Los conjuntos faunísticos analizados fueron obtenidos de excavaciones llevadas a cabo en dos sectores diferenciados del sitio. En los dos casos se trata de sondeos estratigráficos de 2 m2 de superficie realizados en densos depósitos de basura, los cuales fueron excavados hasta alcanzar el substrato estéril. Todos los sedimentos fueron tamizados utilizando mallas de 3,2 mm. Asimismo, ambos depósitos revelaron la presencia de un único componente arqueológico producto principalmente del descarte de materiales cerámicos y óseos. De ellos proceden las fechas radiocarbónicas mencionadas, que permitieron no sólo precisar la cronología del sitio sino también postular la contemporaneidad de los conjuntos recuperados.
El primero de ellos (Bil-R1), fue efectuado en la esquina de un recinto de dimensiones relativamente grandes (6 x 8 m), perteneciente a un complejo doméstico. Su tamaño, junto a la ausencia de restos de techumbre y sus muros bajos, sin evidencias de derrumbe, permitió caracterizar al recinto como un patio descubierto (Nielsen et al. 2014). En torno a este patio se agrupan recintos menores de tamaños variables, conformando un patrón recurrente tanto en Bilcapara como en otros sitios residenciales de la región (Nielsen et al. 2010b). El segundo conjunto (Bil-S2), por el contrario, se recuperó en una vía de circulación adyacente a un recinto de gran tamaño, identificado como una plaza (Nielsen et al. 2014). La edificación del mismo tiene la particularidad de contar con numerosos bloques de cuarzo blanco, rasgo que contrasta con la arquitectura general del sitio, donde predominan esquistos de color oscuro. Esta misma situación también se presenta en el principal espacio público de Chipiwaykho, uno de los asentamientos más importantes de la región situado a una distancia lineal de 50 km (Nielsen et al. 2010b).
Los materiales descartados en este contexto, entonces, pueden ser asociados a las actividades llevadas a cabo en el principal espacio público de Bilcapara.
RESULTADOS
Se analizaron un total de 1904 restos óseos, los cuales conforman la totalidad de los materiales recuperados en ambos depósitos (número total de restos, Tabla 1). El número de especímenes identificados (NISP), observándose el predominio de camélidos en los dos conjuntos (Tabla 1). En Bil-S2 se registró un mayor número de categorías taxonómicas con respecto a Bil-R1, aunque no puede descartarse que esto sea resultado del desigual tamaño de cada una de las muestras. Asimismo, estas diferencias no opacan el predominio de los artiodácti los, incluyendo camélidos, siendo que ambas categorías sumadas superan el 85% de los especímenes identificados en los dos conjuntos.
Tabla 1. Composición taxonómica de Bilcapara.

Las diferencias en la representación de camélidos y artiodáctilos indeterminados entre los conjuntos puede explicarse como resultado del variable grado de fragmentación en cada uno de ellos y de sus consecuentes diferencias en la identificabilidad de los restos. Mientras que en Bil-R1 un 63% de la muestra fue identificada al menos a nivel de orden, en Bil-S2 esta proporción alcanzó apenas el 45%. Es sugerente entonces que el conjunto que presenta un nivel más alto de fragmentación posee, a la vez, mayor abundancia de artiodáctilos indeterminados. En este sentido, es probable que aquellos especímenes asignados a artiodáctilos indeterminados sean camélidos con un grado de identificabilidad bajo. De esto se desprende que, al igual que en la amplia mayoría de los casos analizados para el Sur Andino correspondientes a este segmento temporal, los camélidos fueron el principal recurso faunístico consumido. Al margen de la similitud existente en este nivel, discutiremos a continuación algunos indicadores tafonómicos empelados para camélidos y artiodáctilos indeterminados. Estos estuvieron dirigidos a evaluar comparativamente la ocurrencia de procesos de destrucción posdepositacional entre ambos conjuntos. En la Tabla 2 se observa que tanto en Bil-R1 como en Bil-S2 la mayoría de los especímenes presentan estadios de meteorización entre 0 y 1 (siguiendo a Behrensmeyer 1978). Debemos mencionar además que en ninguno de los dos conjuntos se registraron especímenes con estadios de meteorización superiores a 2. Por otra parte, la incidencia de roedores y carnívoros se presentó baja en ambos conjuntos, aunque levemente superior en Bil-S2. Otro indicador de utilidad para medir la destrucción diferencial dentro de este grupo fue el grado de completitud de huesos carpianos y tarsianos. Al ser poco proclives a seleccionarse para el consumo, su fragmentación puede expresar procesos de destrucción posdepositacional (Marean 1991). En Bil-R1 se hallaron seis especímenes completos sobre un total de siete, mientras que en Bil-S2 esta relación fue de seis sobre ocho, siendo en ambos minoritaria la presencia de huesos fragmentados.
Tabla 2. Indicadores tafonómicos en artiodáctilos (camélidos e indeterminados).
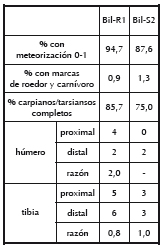
Por último, comparamos la relación entre porciones epifisiarias proximales y distales tanto en húmeros como en tibias. Al presentar las primeras valores de densidad mineral ósea claramente inferiores a las segundas (Elkin 1995; Stahl 1999), y por lo tanto menores chances de preservación, pueden constituir indicadores útiles de destrucción posdepositacional (Binford 1981). De acuerdo con esto, en Bil-R1 no se observó una clara sobrerrepresentación de aquellas porciones con mayores chances de supervivencia. Lo mismo ocurrió en Bil-S2 con respecto a la tibia. Sin embargo, en este conjunto se destaca la ausencia de porciones proximales de húmero, habiéndose registrado la presencia de dos porciones distales. Esto último sugiere niveles más altos de destrucción in situ para los materiales de Bil-S2, lo que es consistente con: (1) la menor identificabilidad general del conjunto; (2) la presencia de más especímenes en el estadio más alto de meteorización; (3) la levemente mayor fragmentación registrada en carpianos y tarsianos; y (4) la también levemente mayor proporción de especímenes con marcas de carnívoros y roedores.
La concurrencia de estos indicadores apoyaría en principio la existencia de peores condiciones de preservación en Bil-S2. Debe señalarse sin embargo que las diferencias entre ambos conjuntos no fueron sustantivas en todos los indicadores utilizados y que, en líneas generales, estos sugieren bajos niveles de destrucción posdepositacional. Si bien esto habilita el análisis comparativo aquí propuesto, advierte sobre condiciones distintas en la formación de ambos conjuntos que retomaremos más adelante.
Diferenciación específica de camélidos
Para discriminar a nivel de especie los restos de camélido se emplearon dos técnicas osteométricas, comparando especímenes arqueológicos con valores actuales de referencia siguiendo el criterio del gradiente de tamaño registrado en camélidos actuales (Mengoni Goñalons y Yacobaccio 2006). En este caso, dicho criterio fue central para tratar la variación taxonómica interconjunto dada la ausencia de otras líneas de evidencia con probado potencial en ese sentido, tales como la caracterización de fibras (Yacobaccio et al. 1997-1998; Reigadas 2008) o la morfología de incisivos (Izeta 2004, entre otros). Los protocolos para la selección, medición y registro de especímenes fueron tomados de Elkin (1996). La primera técnica se basó en la comparación, en una escala logarítmica (Meadow 1999), de una medida para cada hueso arqueológico con respecto al mismo valor de un ejemplar de guanaco actual procedente de las Cumbres Calchaquíes (Salta) medido por Mengoni Goñalons y Elkin (G. Mengoni Goñalons, comunicación personal 2011). Los valores que oscilaron entre ±0,02, denominados intermedios, fueron asignados a la categoría Lama sp. ya que aunque pertenecen al "grupo grande" pueden corresponder tanto a guanacos como a llamas pequeñas. Se midieron las siguientes dimensiones: ancho del extremo proximal de falange proximal y medial, ancho y espesor de metapodio distal, ancho de extremo proximal de radioulna y espesor de tibia proximal.
La segunda técnica empleada consistió en la comparación, a través de gráficos de dispersión, de dos medidas por cada hueso arqueológico con las mismas medidas en camélidos actuales. Los valores de referencia utilizados fueron, además del ejemplar de guanaco ya mencionado, un guanaco procedente de la Falda Occidental del Aconquija, dos vicuñas (procedentes de Jujuy, Argentina, y Oruro, Bolivia) y dos llamas procedentes de las provincias de Jujuy y Catamarca (Yacobaccio et al. 1997-1998; Izeta 2004; Izeta et al. 2009a; G. Mengoni Goñalons, comunicación personal 2011). En este caso se consideraron los siguientes pares de medidas: ancho y espesor del extremo proximal de falanges primeras y segundas, ancho y espesor de los cóndilos distales de metapodios y ancho máximo y espesor de metapodios distales, este último calculado como promedio del espesor de ambos cóndilos.
En la Figura 2 se presenta la dispersión de valores arqueológicos (N = 19) con respecto al estándar de guanaco andino, detallada para cada hueso. En principio, hay que mencionar que ambos conjuntos difieren en la proporción de elementos que pudieron ser analizados por osteometría. En Bil R-1 estos alcanzan el 6,6% del NISP de camélido (N = 10), mientras que en Bil-S2 llegan al 3% (N = 9). Esto puede ser explicado por los mayores niveles de fragmentación y las peores condiciones de preservación detectadas en este último conjunto e impone ciertos recaudos a la hora de comparar la distribución de valores entre ambas muestras. Más allá de esto, debe destacarse que en Bilcapara están presentes los distintos grupos de tamaño, incluyendo una cantidad de valores "intermedios", los cuales alcanzan casi la mitad de la muestra.

Figura 2. Diferencias logarítmicas entre valores arqueológicos y estándar de referencia.
En las Figuras 3 y 4 se muestran las dispersiones de los pares de valores arqueológicos y actuales. Aunque sólo una parte del conjunto medible (N = 14) pudo ser analizado mediante esta técnica, la misma permitió una mayor resolución en las asignaciones taxonómicas de algunos huesos pertenecientes al grupo grande. El análisis bivariado confirmó la tendencia previamente observada.
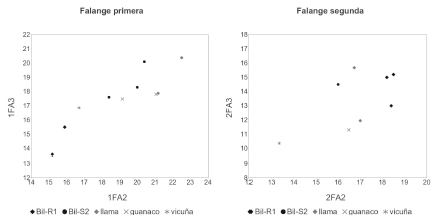
Figura 3. Dispersión de medidas de falanges de camélido (expresadas en mm).

Figura 4. Dispersión de medidas de metapodios de camélido (expresadas en mm).
Resulta importante señalar que ambos conjuntos difieren en cuanto a los grupos de tamaño presentes en cada uno. De acuerdo con el máximo nivel de resolución alcanzado a partir ambas técnicas el análisis osteométrico arrojó la presencia de camélidos pequeños y grandes en proporciones idénticas en Bil-R1 (N = 10). Asimismo, de los cinco especímenes asignados a este último grupo al menos cuatro superan los tamaños conocidos de guanaco norandino, por lo que muy probablemente correspondan a llamas de porte grande. Distinta es la situación en Bil-S2 (N = 9), el basural adyacente al espacio público de Bilcapara, ya que allí no se han registrado especímenes pequeños, asignables a vicuña. En este conjunto, cinco especímenes pudieron ser asignados a llama en función de su tamaño, mientras que los cuatro restantes se encuentran dentro del rango actual de guanaco/llama pequeña, por lo que su adscripción taxonómica a nivel específico no es clara. El panorama para Bilcapara indica entonces la presencia de todos los grupos de tamaño, aunque desde una perspectiva centrada en las variaciones contextuales se observa una distribución restringida de ejemplares de vicuña, acotados exclusivamente a Bil-R1 (patio doméstico), pero en proporciones altas.
El tamaño relativamente pequeño de las muestras y las diferencias en las proporciones de huesos medibles en cada una de ellas obliga a ser cautelosos en la interpretación cuantitativa de estos resultados. En términos cualitativos, sin embargo, podemos afirmar que ambos conjuntos evidencian un contraste claro en la distribución de vicuñas así como la ubicua presencia de llamas, aunque no es posible descartar la presencia de camélidos silvestres en Bil-S2 debido a la existencia de valores "intermedios".
Composición anatómica
El análisis de los perfiles anatómicos involucró exclusivamente a los restos identificados como camélidos (ver Tabla 1). En primera instancia, se cuantificó cada conjunto a partir de unidades anatómicas inclusivas, las cuales agrupan distintos elementos según las unidades resultantes del trozamiento primario de camélidos observadas en pastores andinos actuales (Miller 1979, y casos presentados en Yacobaccio et al. 1998). Estas unidades son luego segmentadas progresivamente hasta su descarte final, siguiendo criterios variables que dependen básicamente de las modalidades de consumo implicadas en cada caso. Por este motivo preferimos emplear con fines exploratorios aquellas categorías anatómicas que nos sitúan desde la perspectiva de las primeras instancias de desorganización anatómica. Este tipo de agrupamiento ha sido empleado en análisis semejantes en nuestro país (Yacobaccio et al. 1997-1998, De Nigris 2004). En esta oportunidad hemos definido seis unidades anatómicas para analizar las diferencias entre conjuntos, cuantificadas a partir del número mínimo de elementos (MNE, calculados según zonas diagnósticas, sensu Mengoni Goñalons 1999), que fue de 88 en Bil-R1 y de 147 en Bil-R2.
La Figura 5 muestra la abundancia relativa de cada unidad anatómica expresada como la razón entre el MNE observados y aquellos presentes en un esqueleto completo, estandarizada en una escala del 0 al 100. En ella se aprecia que en ambos casos todas las unidades anatómicas están presentes pero registran marcadas diferencias en su abundancia relativa. Si bien todas las unidades fueron descartadas, los huesos de las patas y de la cabeza predominan en los dos depósitos en detrimento de las costillas y los pies, siendo más variable la representación de huesos de la columna. Las semejanzas que se perciben entre los dos conjuntos y la presencia ubicua, aunque variable, de todas las unidades anatómicas permite asumir que en ambos casos estamos probablemente ante materiales descartados luego del consumo final y no frente a contextos que reflejan distintas instancias en la secuencia de procesamiento de los animales, aun cuando restan evaluar otros indicadores al respecto (e.g., fracturas intencionales o huellas de corte). Por otra parte, al comparar ambos conjuntos entre sí se observa que los principales contrastes se relacionan con las unidades correspondientes al esqueleto axial. Esto no parece responder a problemas de identificabilidad resultantes de la fragmentación diferencial en cada depósito. En concordancia con lo observado para la categoría Camelidae, los huesos axiales en Artiodáctilos Indeterminados alcanzan un 10,5% en Bil-S2 (N = 30) y apenas un 1,5% en Bil-R1 (N = 1).

Figura 5. Frecuencia relativa de unidades anatómicas de camélido en Bilcapara.
Considerando esta situación, y las eventuales divergencias en la trayectoria de los elementos anatómicos luego de su trozamiento primario, se analizaron las frecuencias de los elementos particulares, tratando de forma independiente la región axial y apendicular por un lado y, por otro lado, considerando el esqueleto completo (Tabla 3). Para esto se emplearon como marco de referencia diversos modelos que jerarquizan cada elemento según distintos criterios de utilidad. Ellos son los índices de carne (IC, Mengoni Goñalons 1991) y de médula (IM, Mengoni Goñalons y De Nigris 1999), que ordenan las partes anatómicas en función de los recursos que ofrecen desde la perspectiva del consumo inmediato. Por otra parte se empleó el índice de secado (ISC), que jerarquiza cada parte según su aptitud para el secado, basándose en las proporciones entre carne, hueso y médula (De Nigris y Mengoni Goñalons 2005). Este índice permite evaluar la presencia de situaciones de consumo diferido. Es importante notar que el potencial para el secado así definido implica la existencia de técnicas de conservación de carne que involucran la presencia de hueso en el producto final, aun cuando se ha registrado entre pastores andinos una amplia variabilidad en este aspecto (ver por ejemplo Miller 1979; Yacobaccio et al. 1998; Stahl 1999). Nuestras observaciones indican que actualmente en la región la técnica de conservación utilizada, denominada localmente charqui, consiste en el secado a la intemperie de carne adherida al hueso y previamente salada. Todos los elementos son charqueados para su almacenamiento a excepción de la cabeza, que incluye cráneo y mandíbula, la cual es consumida de forma inmediata. Antes de evaluar los resultados de estos marcos de referencia, se controló la ocurrencia de patrones de representación que puedan ser dependientes de la densidad mineral ósea (DO), empleando los valores de Elkin (1995) para un total de 22 pares de variables. Todos estos índices se correlacionaron con las frecuencias anatómicas expresadas en MAU (Lyman 2008). En casi todos los casos las correlaciones entre los valores de DO y la representación de partes son positivas, aunque bajas o moderadas y no significativas, siendo la única excepción la situación de Bil-R1 cuando se considera la totalidad del esqueleto (Tabla 3). Aunque se trata de una correlación débil, esto podría ser indicativo de la actuación más acusada de procesos de selección no antrópicos en la formación de este conjunto, en contraste con lo observado para Bil-S2. Sin embargo, esta situación es contradictoria con los resultados obtenidos a partir de otros indicadores tafonómicos, que sugerían un menor nivel de destrucción posdepositacional para Bil-R1 (ver arriba). Si bien en este conjunto la subrepresentación de algunos elementos podría estar condicionada por sus chances de preservación, no parece ser este un factor central en el pautamiento observado. En apoyo a esta idea debemos mencionar que, como se ha expuesto previamente, no se han detectado signos de una intensa destrucción in situ a partir de otros parámetros. Por otra parte, la correlación de Bil-R1 con las frecuencias esperadas según el modelo de destrucción mediada por la DO es estadísticamente significativo sólo en una escala de análisis (el esqueleto completo) y la misma presenta un valor moderado (Tabla 3). Esta serie de factores, puestos además en la perspectiva comparativa entre conjuntos, permiten defender que las diferencias en el pautamiento de ambas muestras son potencialmente informativas de la actividad humana.
Tabla 3. Correlaciones entre frecuencias anatómicas de camélido (MAU) e índices de utilidad y densidad mineral ósea.
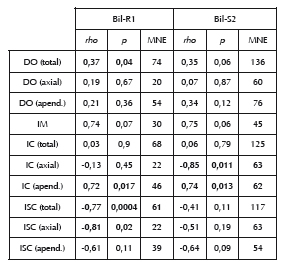
Nota: DO = densidad ósea; IM = Índice de Médula; IC = Índice de Carne; ISC = Índice de Secado. Las correlaciones estadísticamente significativas (p < .05) se destacan en negrita.
Las frecuencias de huesos largos en ambas muestras presentan una correlación positiva y alta, aunque no significativa, con el IM. Por otro lado, la representación del esqueleto apendicular en los dos conjuntos arrojó correlaciones positivas, altas y significativas con el IC. Finalmente, para esta misma región las correlaciones con el ISC son inversas y no significativas. Esto permite sostener que la frecuencia de partes de la región apendicular, semejante en ambos depósitos como fue observado previamente, puede explicarse por el consumo inmediato de carne y probablemente médula. Distinta es la situación del esqueleto axial, para el cual se observan marcadas diferencias interconjunto. En Bil-R1 la utilidad de carne no parece ser un factor determinante en la conformación de las muestras óseas, mientras que con el ISC se presentan correlaciones inversas y significativas. Esto se vincula con la alta frecuencia de partes poco propensas al secado, como el cráneo y la mandíbula, en detrimento de las vértebras y las costillas. Por su parte, en Bil-S2 no se observa para la región axial la incidencia de la selección en función del potencial de secado, aunque sí en función de las partes menos ricas en carne, siendo la correlación con el IC alta, inversa y significativa (Tabla 3).
Si se considera el esqueleto completo como unidad de análisis no se observan patrones de selección en ninguno de los dos conjuntos, al menos desde la perspectiva de los modelos empleados. La única excepción es la alta frecuencia de partes poco aptas para el secado en Bil-R1. En líneas generales, sin embargo, puede argumentarse que ambas regiones esqueletarias siguieron trayectorias diferentes antes de su descarte final. El esqueleto apendicular se comporta de la misma manera en los dos conjuntos, siendo descartados preferentemente los huesos más ricos en carne y eventualmente médula. En cambio, las variaciones contextuales son ostensibles cuando se analiza el tratamiento de las partes axiales. Los contextos y modos de consumo de ciertas unidades anatómicas pueden explicar el descarte diferencial de elementos en uno y otro depósito. A continuación presentamos una discusión integrada de los resultados taxonómicos y anatómicos recién descriptos.
DISCUSIÓN Y CONCLUSIONES
Desde una perspectiva comparativa el registro zooarqueológico de Bilcapara muestra algunas semejanzas pero también diferencias importantes. En términos taxonómicos, y más allá de la variable riqueza presente en los conjuntos -que puede atribuirse provisoriamente al desigual tamaño de muestras- los camélidos predominan en ambos depósitos. A este fenómeno, observado recurrentemente en muestras tardías del Sur Andino, se le suma la ubicuidad de especies identificadas como llamas en ambos conjuntos. De esta manera, el consumo de recursos pastoriles es visible y recurrente en los distintos espacios hasta ahora muestreados de Bilcapara.
Más allá de esta observación general sobre el componente pastoril, es destacable el contraste observado en la distribución de especímenes asignados a vicuña. Estos están restringidos exclusivamente al contexto doméstico (Bil-R1). Si bien la identificación de camélidos silvestres mediante osteometría tiene como límite la resolución de los especímenes pequeños del grupo grande, la circulación de ciertos recursos de caza, en este caso vicuñas, parece no ser homogénea. La alta frecuencia de camélidos silvestres en Bil-R1 se asemeja a lo observado en otros contextos domésticos de sociedades agropastoriles de valles y quebradas (Izeta y Scattolin 2006; Izeta et al. 2009b). Esto es consistente con una estrategia dirigida a minimizar el sacrificio del propio rebaño, observada en familias de pastores puneños actuales (Göbel 1994; Yacobaccio et al. 1998). Aunque actualmente esta estrategia se lleva a cabo a través de la diversificación del rebaño y de la alternancia en las especies a sacrificar, se ha planteado que durante momentos prehispánicos la minimización en las tasas de matanza pudo haber sido resuelta a través de la caza de camélidos silvestres (Yacobaccio et al. 1998). La ausencia de vicuñas en Bil-S2 no va en detrimento de la presencia de otra fauna silvestre, aunque el consumo parece haberse centrado en camélidos domésticos. Con respecto a este punto, el consumo mayoritario de animales domésticos en ámbitos ceremoniales puede ser abordado en el futuro considerando su valor como riqueza social, perspectiva que se ha mostrado promisoria en otros contextos culturales (Russell 1998). De todos modos, la evaluación de cualquier hipótesis al respecto precisa no sólo de muestras más amplias procedentes de los espacios públicos, sino también una mayor comprensión de estos contextos.
Los patrones de descarte de unidades anatómicas en cada depósito también ofrecen información valiosa al respecto. El conjunto de Bil-R1, procedente de un patio doméstico, puede ser explicado como resultado de dos tendencias. La primera consiste en el aprovechamiento inmediato de las partes más ricas en carne y con presencia de médula del esqueleto apendicular y de aquellas unidades con menor potencial de secado del esqueleto axial (cráneo, mandíbula y algunas vértebras). Por otro lado se observa una tendencia a la escasez general de partes aptas para el secado, las cuales se corresponden principalmente con la región axial (Tabla 3). Esto sugiere que estas unidades anatómicas pueden haber sido preparadas para su consumo diferido, lo que habría resultado en una incongruencia espacial en su descarte final. Esto tiene sentido si nos situamos desde la perspectiva del uso de recursos alimenticios a nivel doméstico, el cual involucra posiblemente escalas de consumo acotadas, sobre todo en contextos donde la producción agrícola es relevante. El consumo inmediato de ciertas unidades y la conservación y consumo diferido de otras, cuya trayectoria posmatanza sería más prolongada y su descarte final espacialmente variable, es concordante con los patrones de representación de partes anatómicas observados en este caso. Por otra parte, los huesos axiales del basurero Bil-S2, contiguo y asociado al principal espacio público del sitio, no fueron descartados en relación a su potencial de secado. En este caso, y esto puede generalizarse a toda la muestra, el descarte diferencial en función del consumo diferido no parece haber jugado un papel relevante en la conformación del conjunto. El argumento a favor del consumo inmediato, como lo sugieren los modelos empleados, es concordante con las particularidades funcionales del contexto en el que se recuperó la muestra. Dado el carácter público de dicho contexto, es esperable que la acumulación del conjunto óseo de Bil-S2 resulte de eventos de consumo que involucraron mayor cantidad de personas, en contraste con la situación observada para el patio doméstico de Bil-R1.
Comparando ambos conjuntos, la relación variable entre llamas y vicuñas y sus distintos modos de utilización puede concurrir en la explicación de la composición anatómica de las muestras. A diferencia de lo que ocurre con los camélidos domésticos, el aprovechamiento de vicuñas implica una incongruencia entre los lugares de matanza y consumo final. El transporte y tratamiento diferencial preconsumo de los elementos axiales y apendiculares de estos animales hacia los sitios residenciales puede entonces dar cuenta de los contrastes observados en las representaciones anatómicas.
De cualquier manera, los resultados aquí presentados sugieren que solo algunos indicadores zooarqueológicos varían en función del contexto de procedencia. Los conjuntos óseos estudiados en Bilcapara poseen como propiedades comunes el marcado predomino de camélidos por sobre otros taxones y, dentro de estos, la ubicua presencia de llamas. A esto se le suma la alta frecuencia de individuos sacrificados entre los 12 y los 36 meses, observada previamente a partir del estado de fusión de huesos apendiculares (Maryañski 2010: 102). Dichos elementos, que no parecen ser dependientes del contexto intrasitio, permiten sostener la importancia del pastoreo de camélidos para estas sociedades, al menos durante la fase tardía del PDR.
Por otro lado, la presencia de vicuñas y la frecuencia relativa de partes anatómicas de camélidos en general parecen ser elementos susceptibles de variar en función de los contextos locales. Estos contextos, asociados a espacios públicos y domésticos, difieren en cuanto al carácter y a la escala de las actividades implicadas. El registro analizado en Bilcapara sugiere que dichos aspectos incidieron en las modalidades de uso de los recursos animales, como resultado tanto de la selección variable de especies particulares como del tratamiento diferencial de las carcasas. A lo largo de este trabajo se planteó la importancia de evaluar la variabilidad local en el registro zooarqueológico, bajo la hipótesis de que esta es significativa e informa, potencialmente, sobre las estrategias de uso de la fauna entre las sociedades prehispánicas. Esta variabilidad es particularmente relevante en los grandes sitios residenciales del PDR, ya que se trata de localidades que poseen diferencias funcionales internas marcadas, al menos con respecto a otro tipo de sitios contemporáneos o a asentamientos residenciales más tempranos. Al mismo tiempo, estas diferencias se asocian normalmente a acumulaciones óseas que conforman una oportunidad privilegiada para estudiar las formas de uso de la fauna a escala local, como complemento a las perspectivas de resolución más gruesa.
El lugar ocupado por los camélidos silvestres y domésticos en la economía de las sociedades tardías de valles y quebradas ha sido una temática ampliamente abordada en la literatura zooarqueológica surandina. Sostenemos que más allá de la evaluación de las representaciones taxonómicas en términos cuantitativos, sea útil considerar su ocurrencia en contextos de consumo específicos como vía complementaria para caracterizar los usos de estos animales. Asimismo, los datos obtenidos en Bilcapara sugieren que el contraste entre espacios públicos y domésticos es una distinción válida para construir unidades de observación del registro arqueofaunístico y explicar su variabilidad, no solo desde el punto de vista del consumo sino también desde la perspectiva de la producción y/o extracción de recursos animales. Esta situación advierte sobre los problemas de extrapolar la información procedente de un depósito a la construcción de un panorama regional amplio, sobre todo cuando tratamos con localidades que, como fue mencionado, poseen diferencias internas ostensibles. Finalmente, consideramos que la ampliación de los estudios a múltiples escalas en la CMRSJ permitirá distinguir las propiedades del registro zooarqueológico localmente variables de aquellas de alcance más amplio, con la finalidad de generar hipótesis regionales sobre las estrategias económicas de momentos prehispánicos.
Agradecimientos
Agradezco a Axel Nielsen, Mariana De Nigris y a tres evaluadores anónimos por los comentarios y sugerencias realizados sobre este trabajo. Lo aquí expuesto, sin embargo, es de mi exclusiva responsabilidad.
REFERENCIAS CITADAS
1. Ávila, F. 2009 Interactuando desde el estilo. Variaciones en la circulación espacial y temporal del estilo alfarero Yavi. Estudios Atacameños 37: 29-50. [ Links ]
2. Behrensmeyer, A. K. 1978 Taphonomic and Ecologic Information From Bone Weathering. Paleobiology 4 (2): 150-162. [ Links ]
3. Binford, L. 1981 Bones. Ancient Men and Modern Myths. Academic Press, Nueva York. [ Links ]
4. Dantas, M. 2010 Arqueología de los animales y procesos de diferenciación social en el Valle de Ambato, Catamarca, Argentina. Tesis Doctoral inédita. Facultad de Filosofía y Humanidades, Universidad Nacional de Córdoba, Córdoba. [ Links ]
5. DeFrance, S. 2009 Zooarchaeology in Complex Societies: Political Economy, Status, and Ideology. Journal of Archaeological Research 17 (2): 105-168. [ Links ]
6. De Nigris, M. 2004 El consumo en grupos cazadores recolectores. Un ejemplo zooarqueológico de Patagonia Meridional. Sociedad Argentina de Antropología, Buenos Aires. [ Links ]
7. De Nigris, M. y G. L. Mengoni Goñalons 2005 The Guanaco as a Source of Meat and Fat in the Southern Andes. En The Zooarchaeology of Milk and Fats. Proceedings of the 9th Conference of the ICAZ, editado por J. Mulville y A. Outram, pp. 160-166. Oxbow, Durham. [ Links ]
8. Dietler, M. y B. Hayden 2001 Feasts: Archaeological and Ethnographic Perspectives on Food, Politics, and Power. Smithsonian, Washington. [ Links ]
9. Elkin, D. 1995 Volume density of south american camelid skeletal parts. International Journal of Osteoarchaeology 5: 29-37. [ Links ]
10. Elkin, D. 1996 Arqueozoología de Quebrada Seca 3: Indicadores de subsistencia temprana en la Puna Meridional argentina. Tesis Doctoral inédita. Facultad de Filosofía y Letras, Universidad de Buenos Aires, Buenos Aires. [ Links ]
11. Göbel, B. 1994 El manejo del riesgo en la economía pastoril de Susques. En Zooarqueología de camélidos, vol. 1, editado por D. Elkin, C. Madero, G. L. Mengoni Goñalons, D. Olivera, M. Reigadas y H. Yacobaccio, pp. 43-56. Del Tridente, Buenos Aires. [ Links ]
12. Izeta, A. 2004 Zooarqueología del sur de los Valles Calchaquíes. Estudio de conjuntos faunísticos del período Formativo. Tesis Doctoral inédita. Facultad de Ciencias Naturales y Museo, Universidad Nacional de La Plata, La Plata. [ Links ]
13. Izeta, A 2008 Late Holocene Camelid Use Tendencies in Two Different Ecological Zones of Northwestern Argentina. Quaternary International 180: 135-144. [ Links ]
14. Izeta, A. y M. C. Scattolin 2006 Zooarqueología de una unidad doméstica formativa. El caso de Loma Alta (Catamarca, Argentina). Intersecciones en Antropología 7: 193-205. [ Links ]
15. Izeta, A., C. Otaola y A. Gasco 2009a Estándares métricos y variabilidad en falanges proximales de camélidos sudamericanos. Su importancia como conjunto comparativo para interpretaciones en arqueología. Revista del Museo de Antropología 2 (1): 169-180. [ Links ]
16. Izeta, A., S. Urquiza y L. Baldini 2009b La arqueofauna del período Tardío en el NOA. Una aproximación desde los conjuntos de Molinos I (Provincia de Salta, R. Argentina). Arqueología 15: 63-84. [ Links ]
17. Jackson, H. y S. Scott 2003 Patterns of Elite Faunal Utilization at Moundville, Alabama. American Antiquity 68 (3): 552-572. [ Links ]
18. Krapovickas, P. y S. Aleksandrowicz 1986 Breve visión de la cultura de Yavi. Anales de Arqueología y Etnología 41-42: 83-127. [ Links ]
19. Krapovickas, P. y E. Cigliano 1964 Investigaciones arqueológicas en el valle del río Grande de San Juan (Puna Argentina). Anales de Arqueología y Etnología 17-18: 71-118. [ Links ]
20. Lema, V., M. Giovannetti, C. Deschamps, A. Capparelli y R. Raffino 2011 Análisis de restos faunísticos en el sitio inkaiko El Shincal (Catamarca, Argentina). Comparación con información arqueobotánica y análisis cerámico. Treballs D'Etnoarqueologia 7: 97-112. [ Links ]
21. Lyman, R. L. 2008 Quantitative Paleozoology. Cambridge Manuals in Archeology, Nueva York. [ Links ]
22. Madero, C. 1992 Análisis faunístico de Huachichocana III (Jujuy): Identificación arqueológica de la caza y el pastoreo de camélidos. Palimpsesto 2: 107-122. [ Links ]
23. Madero, C. 1993 Explotación faunística, tafonomía y economía en Humahuaca antes y después de los Yupanqui. En Inka: Arqueología, historia y urbanismo del Altiplano Andino, editado por R. Raffino, pp. 145-168. Corregidor, Buenos Aires. [ Links ]
24. Marean, C. W. 1991 Measuring the Post-Depositional Destruction of Bone in Archaeological Assemblages. Journal of Archaeological Science 18: 677-694. [ Links ]
25. Maryañski, J. 2010. Una contribución a la economía de la caza y el pastoreo en los Valles Chichas (Depto. de Potosí, Bolivia) a lo largo del período de Desarrollos Regionales (ca. 900-1450 d.C.). Tesis de Licenciatura inédita. Departamento de Ciencias Antropológicas, Universidad de Buenos Aires. [ Links ]
26. Meadow, R. 1999 The Use of Size Index Scaling Techniques for Research on Archaeozoological Collections from the Middle East. En Historia Animalium ex Ossibus, Festschrift für Angela von den Driesch. Beiträge Zur Paläoanatomie, Archäologie, Ägyptologie, Ethnologie und Geschichte der Tiermedizin, editado por C. Becker, H. Manhart, J. Peters y J. Schibler, pp. 285-300. [ Links ]
27. Mengoni Goñalons, G. L. 1991 La llama y sus productos primarios. Arqueología 1: 179-196. [ Links ]
28. Mengoni Goñalons, G. L. 1999 Cazadores de guanacos de la estepa patagónica. Sociedad Argentina de Antropología, Buenos Aires. [ Links ]
29. Mengoni Goñalons, G. L. 2010 El aprovechamiento de la fauna en sociedades complejas: aspectos metodológicos y su aplicación en diferentes contextos arqueológicos del NOA. En Al borde del Imperio. Paisajes sociales en áreas periféricas del Qollasuyu, editado por V. Williams y B. Cremonte. En prensa. [ Links ]
30. Mengoni Goñalons, G. L. y M. De Nigris 1999 Procesamiento de huesos largos de guanaco en Cerro de los Indios 1 (Santa Cruz). Soplando en el viento: Actas de las Terceras Jornadas de Arqueología de la Patagonia, editado por Comité editorial de las Terceras Jornadas de Arqueología de la Patagonia, pp. 461-475. Universidad del Comahue-Instituto Nacional de Antropología y Pensamiento Latinoamericano (INAPL), Neuquén y Buenos Aires. [ Links ]
31. Mengoni Goñalons, G. L. y H. Yacobaccio 2006 The Domestication of South American Camelids. A View from the South-Central Andes. En Documenting Domestication. New Genetic and Archaeological Paradigms, editado por M. Zeder, D. Bradley, E. Emshwiller y B. Smith, pp. 228-244. University of California Press, Berkeley. [ Links ]
32. Mercolli, P. 2011 El truco está en la diversidad: Estrategias de manejo pastoril en la Quebrada de Humahuaca, Provincia de Jujuy, Argentina, Entre el 900 y el 1200 AD. Arqueología 17: 81-99. [ Links ]
33. Mercolli, P. y V. Seldes 2007 Las sociedades del Tardío en la Quebrada de Humahuaca. Perspectivas desde los registros bioarqueológico y zooarqueológico. En Producción y circulación prehispánicas de bienes en el Sur Andino, editado por A. Nielsen, M. C. Rivolta, V. Seldes, M. M. Vázquez y P. Mercolli, pp. 259-276. Brujas, Córdoba. [ Links ]
34. Miller, G. 1979 An Introduction to the Ethnoarchaeology of the Andean Camelids. Tesis Doctoral inédita. Graduate Division, University of California, Berkeley. [ Links ]
35. Miller, G. y R. Burger 1995 Our Father the Cayman, Our Dinner the Llama: Animal Utilization at Chavín de Huantar, Perú. American Antiquity 60 (3): 421-458. [ Links ]
36. Miracle, P. 2002 Mesolithic Meals From Mesolithic Middens. En Consuming Passions and Patterns of Consumption, editado por P. Miracle y N. Millner, pp. 65-88. Mc Donald Institute for Archaeological Research, Cambridge. [ Links ]
37. Nielsen, A. E. 2006 Plazas para los antepasados: Descentralización y poder corporativo en las formaciones políticas pre-incaicas de los Andes Circumpuneños. Estudios Atacameños 31: 63-89. [ Links ]
38. Nielsen, A. E. 2007 El período de Desarrollos Regionales en la Quebrada de Humahuaca: aspectos cronológicos. En Sociedades precolombinas surandinas: Temporalidad, interacción y dinámica cultural del NOA en el ámbito de los Andes Centro Sur, editado por V. Williams, B. Ventura, A. Callegari y H. Yacobaccio, pp. 235-251. S/d, Buenos Aires. [ Links ]
39. Nielsen, A. E., P. Mercolli y N. Nasif 2010a Ocupaciones temporarias y explotación faunística en la región lacustre altoandina. Actas del XVII Congreso Nacional de Arqueología Chilena 2: 1365- 1378. Valdivia. [ Links ]
40. Nielsen, A. E., F. Ávila y M. Vázquez 2010b Notas Sobre la arqueología de la Cuenca Media del río Grande de San Juan (1000-1450 d.C.). Trabajo presentado en el XVII Congreso Nacional de Arqueología Argentina, Mendoza. [ Links ]
41. Nielsen, A. E., C. Angiorama, J. Maryañski, F. Ávila y M. L. López 2014 Paisajes prehispánicos tardíos en San Juan Mayo (frontera Argentina-Bolivia). Arqueología, en prensa. [ Links ]
42. Olivera, D. y J. Grant 2009 Puestos de Altura de la Puna Argentina: Zooarqueología de Real Grande 1 y 6 y Alero Tomayoc. Revista del Museo de Antropología 2: 151-168. [ Links ]
43. Raffino, R. 2007 Poblaciones indígenas en Argentina. Urbanismo y proceso social precolombino, 3a. ed. Emecé, Buenos Aires. [ Links ]
44. Reigadas, M. C. 2008 Explotación de recursos animales y producción textil durante el Holoceno en Antofagasta de la Sierra. Estudios Atacameños 35: 35-48. [ Links ]
45. Rodríguez Loredo, C. 1997-1998 Estudio arqueozoológico del sitio Inca Potrero-Chaquiago, Barrios La Solana y Retambay, Andalgalá, Pcia. de Catamarca (Argentina). Relaciones de la Sociedad Argentina de Antropología XXII-XXIII: 203-245. [ Links ]
46. Russell, N. 1998 Cattle as Wealth in Neolithic Europe: Where's the Beef? En The Archaeology of Value: Essays on Prestige and the Process of Valuation, editado por D. Bailey, pp. 42-54. BAR International Series 730, Archaeopress, Oxford. [ Links ]
47. Stahl, P. 1999 Structural Density of Domesticated South American Camelid Skeletal Elements and the Archaeological Investigation of Andean Ch'arki. Journal of Archaeological Science 26: 1347-1368. [ Links ]
48. Yacobaccio, H., C. Madero y M. Malmierca 1998 Etnoarqueología de pastores surandinos. Grupo de Zooarqueología de Camélidos, Buenos Aires. [ Links ]
49. Yacobaccio, H., C. Madero, M. Malmierca y M. C. Reigadas 1997-1998 Caza, domesticación y pastoreo de camélidos en la Puna Argentina. Relaciones de la Sociedad Argentina de Antropología XXII-XXIII: 389-418. [ Links ]

