










Servicios Personalizados
Revista

Articulo
 Inglés (pdf)
Inglés (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkIntersecciones en antropología
versión On-line ISSN 1850-373X
Intersecciones antropol. vol.15 supl.1 Olavarría dic. 2014
ARTÍCULOS
Taphonomy in the kitchen: culinary practices and processing residues of native tuberous plants of the south-central Andes
María del Pilar Babot, Julia Lund and Adriana Valeria Olmos
María del Pilar Babot. Instituto de Arqueología y Museo, Facultad de Ciencias Naturales e Instituto Miguel Lillo, Universidad Nacional de Tucumán (IAM, FCN e IML, UNT). Instituto Superior de Estudios Sociales - Consejo Nacional de Investigaciones Científicas y Técnicas-UNT (ISES-CONICET-UNT). San Lorenzo 429 (4000), San Miguel de Tucumán, Tucumán, Argentina. E-mail: pilarbabot@yahoo.com
Julia Lund. Instituto de Arqueología y Museo, Facultad de Ciencias Naturales e Instituto Miguel Lillo, Universidad Nacional de Tucumán (IAM, FCN e IML, UNT). San Martín 1545 (4000), San Miguel de Tucumán, Tucumán, Argentina. E-mail: julialund13@yahoo.es
Adriana Valeria Olmos. Instituto de Arqueología y Museo, Facultad de Ciencias Naturales e Instituto Miguel Lillo, Universidad Nacional de Tucumán (IAM, FCN e IML, UNT). San Martín 1545 (4000), San Miguel de Tucumán, Tucumán, Argentina. E-mail: adriana.valeria.olmos@gmail.com
Received 20 August 2013.
Accepted 14 February 2014
ABSTRACT
We present comparative material for the identification of culinary residues of cooked tubers of Solanum sp., Oxalis tuberosa and Ullucus tuberosus. We use a broad concept of taphonomy that includes the study of plant modifications resulting from the preparation of food, in this case the boiling and cooking al rescoldo of fresh tubers. We undertake a number of controlled cooking experiments and compare the results with fresh samples. We discuss morphological and optical modifications of tissue fragments and intracellular particles resulting from our cooking experiments. Finally, we discuss the possibility of recognizing cooking techniques from microscopic analysis of tuber remains.
Keywords: Taphonomy; Culinary techniques; Plant processing; Starch; Tubers.
RESUMEN
Tafonomía en la cocina: prácticas culinarias y residuos del procesamiento de plantas tuberosas nativas de los Andes centro-sur. Presentamos material comparativo para la identificación de residuos culinarios de tubérculos cocidos de Solanum sp., Oxalis tuberosa y Ullucus tuberosus. Partimos de un concepto amplio de tafonomía que incluye el estudio de las modificaciones de las plantas resultantes de la preparación de alimentos; en este caso, aquellas que se deben al hervido y cocción al rescoldo de tubérculos frescos. Realizamos experimentos de cocción controlados y comparamos los resultados con muestras frescas. Describimos las modificaciones en los atributos morfológicos y ópticos de tejidos y partículas intracelulares resultantes de nuestros experimentos de cocción. Finalmente, discutimos la posibilidad de reconocer las técnicas de cocción a partir del análisis microscópico de vestigios de tubérculos.
Palabras clave: Tafonomía; Técnicas culinarias; Procesamiento de plantas; Almidón; Tubérculos.
A TAPHONOMIC APPROACH OF CULINARY TRANSFORMATIONS
This work is part of a series of studies initiated over a decade ago (Babot and Korstanje 2001; Babot 2003), aimed at generating comparative material for identifying plant processing techniques from microscopic analysis of preserved residues of plants native to the south-central Andes. It is worth noting that the transformations and agents involved in such preparations (e.g., cooking) go beyond the scope of food, as medicinal or ritual uses can also be important. Thus, although the perspective of this work is culinary, this is only circumstantial; the emphasis is on how techniques, as modes of transformation, affect matter (Pazzarelli 2012). This study deals with the cooking of tuberous plants of the genus Solanum L. (Solanaceae), "potatoes"; Oxalis tuberosa Mol. (Oxalidaceae), "oca" and Ullucus tuberosus Caldas (Basellaceae), "ulluco" or "papa lisa". We began with an ethnobotanical approach that considers the traditional uses of plants in the southcentral Andean region and the various means by which tubers become food. We developed experiments to obtain tubers cooked al rescoldo and boiled tubers, which may themselves constitute simple foods, based on a single ingredient, or be part of complex preparations with other plant, animal and mineral ingredients. We rely on a broad concept of "recipe" "[...] as a more or less flexible or open formula to achieve a preparation" (Babot et al. 2012: 242). The components of a recipe are its ingredients in relative amounts and combinations, the modes and techniques of preparation and service, the circumstances of its use, the implements required throughout the process, and the spatio-temporal context and the actors involved in the culinary performance (Babot et al. 2012).
Of all these components, this paper deals with tubers as ingredients in recipes, and with methods of tuber preparation. Preparation sequences are not rigid or linear because a food element can be associated with multiple recipe pathways of different length and complexity (Babot 2009). "So, when we refer to old recipes, we refer to approximations to the modes of preparing foods, rather than to strict and closed models of the ingredients, their preparation and presentation [...]" (Babot et al. 2012: 241).
The taphonomic study of food residues gives us a unique perspective on how plants, animals and minerals were transformed into food and their subsequent history to the point of archaeological recovery and analysis. According to the "unrestricted taphonomy" approach (Borrero 2011) we use a broad definition of the concept as the study of the decomposition "or modification" of organisms, their parts or products, and of the processes leading to their accumulation and differential preservation in archaeological contexts, so that they can be genuine sources of archaeological information (Lewarch and O'Brien 1981; Borrazzo 2006). We are particularly interested in assessing how such a perspective can inform our interpretation of past human practices. Thus, we consider the modifications of plants' useful parts associated with different methods of food preparation, which condition the appearance and integrity of such food remains as recovered from archaeological contexts. Such modifications can be explained in terms of different physico-chemical processes that affect the structure and the properties of starch and other vegetable intracellular particles, and histological elements (Babot 2003). Previous work has demonstrated the utility of taphonomic and experimental approaches to the macroscopic and microscopic analyses of the modes and procedures for preparing animal or vegetable foods (e.g., González and Frère 2004; Gong et al. 2011; López et al. 2011; Lovis et al. 2011; Raviele 2011; Babot et al. 2012 and references therein; Lantos et al. 2012). In particular, our approach focuses on the micro-morphological characterization of such modifications.
In this study, we characterized the anthropic/ culinary modifications of micro-particles and tissue fragments in different varieties of tubers of three genera, which allowed us to formulate expectations for archaeological cases. Thus, the various stages of investigation aimed to: 1) characterize fresh tissues of different varieties of potato, oca and ulluco, since these are ingredients in traditional culinary preparations and recipes; 2) observe how aspects of the fresh tissue of potato, oca and ulluco change or disappear as a consequence of two traditional food preparation methods (boiling and cooking al rescoldo until completely cooked), and identify which new elements appear in processed foods, explaining these on the basis of the physico-chemical processes involved; and 3) compare the two sets of observations to identify taphonomic signatures of these culinary practices in archaeological situations.
THE TAPHONOMY OF MICROFOSSILS
Current Perspectives
The taphonomic study of plant microfossils and their archaeological associations is a versatile approach. Some taphonomic researches focus on the ways various biological (e.g., soil fauna and roots) and environmental agents (e.g., pH, moisture and temperature conditions, salt precipitation and transport) alter the microfossil record and affect its survival and integrity (Therin 1994; Hart 2003; Humphreys et al. 2003; Haslam 2004; Barton and Matthews 2006; Osterrieth et al. 2013; Musaubach and Babot 2014, among others).
These studies also consider humans, through the manipulation of the natural environment, as an agent of transformation, since they can modify plants and their microfossil remains through residential use of certain places, agriculture, or burning (Therin 1994; Parr et al. 2008; Osterrieth et al. 2009, among others). These kind of taphonomic researches have focused on analysis of biosiliceous particles, calcium phytoliths and starch from the perspective of paleoecological, pedological and archaeological problems. Previous work has also included study of the effects of different laboratory manipulations on the integrity and composition of microfossil assemblages that are ultimately observed under the microscope. Many of these recent studies also involved revision of sample extraction and treatment protocols with particular attention to archaeological applications (Babot and Korstanje 2001; Coil et al. 2003; Korstanje 2003; Babot 2007; Korstanje and Babot 2007; Torrence and Barton 2007, etc.). Other taphonomic studies closely related to the analysis of use residues on archaeological artifacts assess the survival rates and prospects of starch -one of the main microfossils useful for these purposes- under different conditions of entrapment, depth, sediment compaction and exposure, and considering the passage of time according to the nature of the processed substance (Lu 2003; Babot and Bru de Labanda 2005; Barton and Matthews 2006; Langejans 2010, among others). The potential for post-depositional contamination has also been discussed (Barton et al. 1998). Related to taphonomic studies but following different approaches are archaeologically or bromatologically inclined histological studies designed to understand the feasibility and reliability of identifying plant tissues exhibiting different degrees of fragmentation and preservation (Pochettino and Scattolin 1991; Cortella and Pochettino 1994).
We are optimistic about the possibility of obtaining archaeological information from taphonomic and contextual approaches and we consider residues (meaning remains in general, rather than a priori use residues) as dynamic systems that can provide data about use, the contemporaneous context for such use, processes subsequent to deposition, and the manufacture of associated artifacts (the latter when microfossils are included in the artifact raw materials) (Babot and Haros 2008; Hart 2011; Babot et al. 2012). Thus, when considering plant residues located on an artifact or in an archaeological matrix, it would hypothetically be possible to identify interfaces. In the first case, such interfaces would be artifact/residue/ sediment matrix after deposition, where the microfossil "signal" attributable to the raw materials would decrease from the artifact mass to its surface; the "signal" related to use would decrease from the surface of the artifact to the sediment matrix, and the "signal" of the matrix would follow an opposite tendency to that of use. Hence, precautions taken during sample extraction and the criteria used to interpret manufacture, use and context are very important (Babot and Haros 2008; Zucol and Loponte 2008; Hart 2011). The exchanges, gains, losses and modifications of material that occur at the interfaces between object, use residue and matrix must be considered, including other processes that may occur between the time archaeological remains are extracted and analytical data are obtained. Sometimes, burial conditions can promote the preservation of residues, degradation byproducts and new products resulting from interactions with the sedimentary matrix (Jones 2009). This applies to different types of elements contained in the residues, whether chemical, histological or intracellular.
In this regard, we consider that interpreting the nature of a residue requires knowing the particular history of the object under analysis. In other words, a residue could indicate 1) the last recorded use of an artifact (Haslam 2006), or 2) successive use episodes recorded in the stratification of the remains (Musaubach and Beron 2012), or 3) random averaging (in a broad sense rather than referring to statistical significance) resulting from repeated use, partial cleaning between successive uses, differential decay of components and exchanges between the environment, the object's mass and the residue, plus laboratory treatments (Babot 2007; Babot et al. 2012).
A taphonomic approach to plant microfossils and anthropogenic manipulations
A small number of studies have discussed the transformation of tissues and intracellular particles resulting from anthropogenic manipulation of plants. Examples include the identification of striae on the surface of silica phytoliths obtained during threshing of silica-rich taxa (e.g., European grains); the transformation of calcium oxalate crystals into calcite pseudomorphs, which occurs during combustion (Juan-Tresserras 1992); the fracture of biosiliceous particles as a result of activities such as grinding (Checa et al. 1999); and the tearing and disarrangement of plant fibers during mastication (Musaubach and Babot 2014). The formation of new products during cooking (e.g., micro-charcoals and particle clumps) has also been documented (Babot 2003; Tassara and Osterrieth 2008, among others). Starch is one of the most studied intracellular particles, both in archaeology and other fields. Analyses of the physico-chemical processes that can alter starch have a long history, linked initially to biological and food industry interests (e.g., Radley 1943; Whistler et al. 1984). These studies have addressed aspects including starch grains' loss of structure and birefringence properties through contact with caustic chemicals, heat in the presence of water, the action of enzymes during seed germination, and the sprouting of underground organs. The first applications of this knowledge to archaeological situations were related to questions about food, particularly brewing, grinding and boiling of starchy plants (e.g., Juan- Tresserras 1992; Loy 1994; Checa et al. 1999). These studies, along with the work of Babot (2003), Beck and Torrence (2006), Fullagar (2006), Samuel (2006), Henry et al. (2009), Gong et al. (2011) and Crowther (2012), are directly related to our research, as they consider the effects past anthropogenic manipulation of plants had on their starches. Babot's (2003) research aimed to develop modern comparative standards for application to archaeological cases (Babot 2009; Babot et al. 2012, etc.), and has focused on the optical and morphological effects of various processing techniques on starch (e.g., aeration, sun drying, toasting, ashing, freezing, desaponification/washing -i.e., removal of saponins-, and grinding). Babot's research also includes recognition of various cooking practices from starchy residues. Beck and Torrence (2006) considered the paths of starch when used for different purposes in various cultural contexts. Fullagar (2006) evaluated the role of experiments in the functional assessment of archaeological artifacts, and Samuel (2006) discussed various ways in which modified starch has been preserved, including bread and yeast residues. Henry et al. (2009) documented the progression of starch damage due to the baking, boiling, drying, and fermentation of domesticated legumes and grass seeds, and found boiling to be the most harmful cooking technique for starch. Gong et al. (2011) studied food remains from exceptionally well preserved mortuary offerings, and developed experiments to aid identification of culinary preparations. Crowther (2012) discussed the influence of moisture on starch during cooking.
These studies of cultural manipulations have shown that, although they are modified, starch and other intracellular elements can survive multiple food preparation processes. Drying, heating, the breaking down of tissues by trampling and friction, and the formation of ice crystals, all generate physico-chemical changes in those particles, modify their completeness and degree of crystallinity, and produce optical and morphological changes in some grains (Babot 2003). While the damage caused by different food processing techniques may look similar, and although sometimes the same process can alter the starch from different biological sources in different ways (Babot 2003; Henry et al. 2009), it has been shown that, in general, distinct damage patterns seem to result from different processes (Babot 2003).
Also, it has been documented that the more intense the process, the greater the severity of the damage. In addition, it is sometimes possible to infer the consecutive stages of processing, which are present as damage patterns superimposed on the same individual starch granule or on different granules within a sample. Damaged starch grains are more susceptible to hydrolytic agents and fungal and bacterial activity than well-preserved starch. This is attributed to infiltration of the grains' interior via cracks associated with the damage (Radley 1943; Cortella and Pochettino 1994) and to structural weaknesses resulting from physicochemical changes to the granules. However, research on ancient starch has shown that it is possible to recover damaged grains from both modern and archaeological contexts, as this carbohydrate's pseudocrystalline structure survives the passage of time even when damaged. Ancient starch analyses with a taphonomic perspective have increasingly focused on damaged starch grains (alongside those recovered intact), as well as the natural and cultural causes of the observed changes. In addition, appropriate and specific terminologies for the description of alteration processes and damage have been adopted (Babot 2003; The International Code for Starch Nomenclature [ICSN 2011]), referring to the so-called modified starch (sensu Samuel 2006; ICSN 2011). This category and one referred to as resistant starch1 have been distinguished from native starch, which refers to unaltered grains. Such studies have recognized certain traits that have a taphonomic rather than taxonomic origin, and have proposed that damaged starch is a source of information regarding cultural practices and postdepositional processes. Damage patterns have also been used to evaluate contamination by assessing the coherence between the types of damage and the archaeological context from which the starch was recovered (Babot 2003). Previous publications have proposed descriptors for modified starches, but when studying human practices we suggest it is possible to rank those descriptors by grouping them following hierarchical categories: a) techniques: modes of transformation implemented by humans; b) agents other than humans -the latter included in a- ; c) physico-chemical processes triggered by human activities and their results or the final state of the mass of material processed, and, finally, d) morphological and optical damage and modifications or changes that are verified in starch grains within the mass (Table 1). Following Lyman's (1994) terminology, applied techniques and resulting physico-chemical processes constitute taphonomic processes, where the taphonomic agents are the physical, chemical and biological factors involved in the transformation of grains. In this case humans are the default participants in technical manipulation. Finally, the taphonomic effects are the results of these processes and the specific patterns of damage and changes that may be identified histologically and described in morphological and optical terms (Table 1). Thus, for example, what we observe as a paste or a mixture of cooked food that includes resistant and modified starch "damaged in various ways" is a modification due to the retrogradation that occurs when boiled dough is left to rest, or a modification due to the melting by heating in a medium with low water content (Radley 1943; Johnson et al. 1990).
Table 1. Techniques, agents, physico-chemical processes and results, and morphological and optical modifications relevant to the study of human manipulation of starchy substances

The two main physico-chemical processes we address in this paper, gelatinization and melting, are described in previous works. The effects of heating in a wet medium have been studied extensively, especially for the generation of baseline information on the properties of starch for industrial purposes. It leads to a non-reversible process called gelatinization (Radley 1943). For dry heating or for heating with low moisture content, we consider the "dry-cooking series", toasting/ cooking al rescoldo/cooking on embers/roasting/ charring2, wherein the relative intensity of the heat and, therefore, the thermal alteration of the granules, increase from toasting to charring (Radley 1943; Babot 2003). Direct contact with the heat source differentiates roasting, charring and cooking al rescoldo from toasting and cooking on embers, with heat contact being indirect in the latter.
PROCEDURES AND MATERIALS
Tuber samples were obtained from traditional vending stalls in Jujuy Province (Argentina) in 2010 and 2012 and Villazón (Bolivia) in 2012. We obtained 19 ethnovarieties of potato (Solanum L.), 4 of oca (Oxalis tuberosa Mol.) and 3 of ulluco (Ullucus tuberosus Caldas), all from rural areas. Here we selected 10 varieties of potato, 2 of oca and 1 of ulluco, which were sampled and processed for culinary purposes. The purpose of using different ethno-varieties was to assess whether there were variations if different kinds of potatoes were subjected to different processing techniques, particularly since they could have been used in different traditional culinary practices (Castro 2008).
In all ethno-varieties, cortex and periderm were sampled separately from the vascular parenchyma and medullary tissue. Cortex and periderm were separated under the assumption that they were part of the "shell" that was eventually removed or peeled prior to consumption, while the parenchyma and the medullary tissue constitute the soft part intended for consumption. Three types of samplings were used for the fresh tubers to obtain reference material: 1) soft scrapings and histological cuts done freehand with a scalpel to obtain thin sections (Babot 2007), 2) diaphanization (a technique in which the tissue is treated with an oxidizing agent to make it transparent whilst retaining fabric), and 3) dry ashing (Piperno 1988). Some washing of the ash material was incorporated as well to assess tissue loss by this procedure. The first two sampling techniques provided references regarding tissue appearance and various elements of tissues, such as ergastic substances (starch, cellulose and crystalline calcium salts), of interest to archeology because they survive as microfossils. These procedures allowed us to observe intact tissue and fragments in different stages of disintegration, as well as free cells and intracellular elements. Controlled diaphanization at temperatures below 10°C thinned the material for better observation. Dry ashing allowed for recovery of the silicified tuber tissues. Due to the low proportion of silica produced by these plants, after the first trials, we avoided to wash ash samples because doing so results in a significant loss of material. Cooking experiments included: a) boiling until cooking was complete, transforming the starch-rich parenchyma and medullary tissue by heating it to boiling in a humid medium with sufficient water; b) cooking al rescoldo (in direct contact with embers and ashes) until cooking was complete, transforming tissues by heating them at moderate temperature in a dry medium or in the presence of tuber residual moisture. Cooking enables consumption, increases energetic value of tubers and eliminates the causes of indigestion (Wandsnider 1997).
Boiling was performed using a stove; tubers were placed in a metal container filled with potable water to avoid contamination. Unpeeled tubers were boiled separately, reaching an average of 99ºC during the time needed to soften the parenchyma and medullary tissue (between 15-20 minutes). Tubers started to loss their peel and stain water before completely cooked. Pink oca stained the water and started to loss their peel after 15 minutes in the boiling water. Nine minutes later it was completely cooked. White oca lost their peel at 17 minutes and started to smell like boiled sweet potato. Eight minutes later it was completely cooked. Ulluco took 19 minutes to begin staining the water, losing a minimal amount of peel. Two minutes later was completely cooked. Potatoes took 24 minutes to cook completely.
Cooking al rescoldo was done in a backyard fire. The fire was started with quebracho blanco (Aspidosperma quebracho-blanco Schltr.) wood. Red hot embers were produced and gradually covered with ash, at which point the charcoal was not as hot as in its red-hot state. A layer of embers and ashes was dispersed on the firebrick floor. Whole, unpeeled tubers were set over the embers and ashes and exposed to the heat source until the parenchyma and medullary tissue were softened, simulating the effect of placing tubers on the periphery of an active fire. This is a dry-cooking technique different from others such as toasting, cooking on the flame or on embers with a grate, or in ovens. Cooking on the flame involves direct contact of food with the heat source; placing a grate between the food and flame or red-hot embers separates food from the heat source. Toasting requires an intermediate object (e.g., a stone or vessel) to avoid direct contact with fire and buffer heat (Pazzarelli 2012). Cooking al rescoldo is a moderatetemperature procedure traditional in northwestern Argentina, where food is put into embers and ashes, coming into direct contact with the heat source. In our experiment, cooking al rescoldo took between 10- 15 minutes depending on the type and size/form of the tuber and degree of exposure to the heat source. Ocas cooked most quickly, followed by ullucos and longer/thinner potatoes and, finally, thicker potatoes. During this process the tubers were not covered, so the periderm was carbonized where directly in contact with embers. Differences in the humidity of the cooked mass were observed in areas near carbonized and noncarbonized periderm. Oxygen and temperature might have fluctuated between the periphery and the center of the ember and ash layer within the range of 100- 200ºC. Quebracho blanco provided a strong, slow, constant and non-sparking charcoal combustion, with low ash production.
Boiled tubers and those cooked al rescoldo were wrapped in aluminum foil and refrigerated until sampling was complete. Sampling followed the same procedures used for fresh specimens (see above). Fresh, dry-ashed and cooked specimens were mounted on slides for viewing and photographing using a polarizing microscope (200X to 630X). Assemblage analysis focused on the various histological elements and intracellular particles present. Characteristics recorded for fresh samples include: cell shape in two- and three-dimensions, arrangement of tissue cells (tiled, linear, concentric, etc.), color with and without polarizer, birefringence, cell size, presence of conduction elements and their characteristics, presence of cellulose in the cell walls; presence, kind, shape, color, presentation and location of calcium salt bodies within the tissues (isolated, grouped); and the occurrence of starch in the reserve parenchyma and the medullary tissue, and its disposition (isolated, in clusters, massively filling the tissue). The same characteristics were observed in food samples, with special attention to the features preserved, modified, and originated from culinary practices. In the case of native and modified starches, the specific variables summarized in the ICSN (2011) and previous studies were recorded (Korstanje and Babot 2007 and references therein; Henry et al. 2009). As mentioned, rather than a detailed description of individual starch grains, the presence of various attributes was confirmed (Table 1). Observations proceeded from questions such as: What is the general state of preservation of the intracellular particles and tissues in our samples? What is the degree of alteration? How does alteration vary with cooking techniques and their relative "aggressiveness"? Have the integrity, visibility, shape, size, color, spatial arrangement, optical properties or textures of tissues, cells and ergastic substances been altered? If so, how? Have any elements or distinctive features of fresh tissues disappeared? Have any new elements arisen as a result of processing?
Finally, we compared the two sets of observations, which allowed us to generate expectations for documenting foods made from potatoes, oca and ulluco based on archaeological residues (Tables 2 and 3).
Table 2. Characteristic features of the periderm and cortex of fresh and boiled tubers, and tubers cooked al rescoldo. Note: CE= Cellulose.

Table 3. Characteristic features of the parenchyma and the medullary tissue of fresh and boiled tubers, and tubers cooked al rescoldo. Note: CE = Cellulose; CR = Crystals; VR = Vessel elements; NP = New products.

RESULTS
Description of fresh tuber specimens Periderm and cortex tissues are turgid and present a tight or dense mosaic-like fabric of polyhedral cells (Figure 1; Table 2). The tissue is non-birefringent -with the exception of periderm cell walls that are cellulose enriched- and either colorless or colored by sections (Figure 1a-b). The presence of calcium salt crystals (primarily calciumoxalate) is variable (Figure 1). They occur as polyhedra in oca and ulluco (Figures1 i-n, p-r); tabular druses and crystal sand in oca, ulluco and Solanum (Figure 3o-p); and flat, rounded or globular particles in Solanum (Figure 1h).

Figure 1. Appearance of the fresh periderm and cortex of: a-h) Solanum, i-n) Oxalis tuberosa and o-r) Ullucus tuberosus. a-b) Pentaoca, c-d) Abajeña, e-f) Huanco Suyo, g-h) Solanum 1a, i-l) Pink oca, m-n) White oca, o-r) Ulluco. The contiguous twin micrographs correspond to views with parallel (left) and crossed (right) nicols of the same tissue

Figure 3. Appearance of the periderm and cortex of Oxalis tuberosa cooked al rescoldo. a-j) White Oca, k-s) Pink Oca. The contiguous twin micrographs correspond to views with parallel (left) and crossed (right) nicols of the same sample.
Parenchymal and fresh medullary tissue cells are polyhedral and globular, with thin and less celluloseenriched membranes than the periderm. They are closely packed and contain numerous starch grains of the type described previously ( e . g . , Korstanje and Babot 2007 and references therein), completely filling reserve tissue (Figure 5u, Table 3). Abundant calcium crystals are observed in ulluco and oca parenchyma (Figure 7i-n); in Solanum, crystal abundance depends on the variety (Figures 4c, e and 5ef). Dense clusters of birefringent conducting elements are observed, especially in the section corresponding to the vascular ring (Figures 5a-b and 7f-g). Description of tuber specimens cooked al rescoldo Carbonized or highly thermally altered sections of the periderm and cortex appear opaque to slightly translucent under the microscope. The internal structures and content of some tissues are difficult to observe, but in other sections, birefringent elements stand out well (Figures 2d-f, o-r and 3k-m; Table 2). Parts of the tissues that were not in direct contact with fire are less thermally altered (Figure 2a-c, h-I). Silicified parts stand out from the tissues around them due to a higher relief than the periphery. Occasionally, they appear as isolated polyhedral or globular siliceous particles (Figure 3k, g-h). Some deformation, fragmentation, crack and folds are observed, but this kind of damage is less pronounced among tubers cooked al rescoldo than among those that were boiled (Figures 2c-d, p). Tissues significantly affected by heat become brittle, dehydrated and shrunken. They are stained reddish-brown to black (in carbonized areas ) , which accentuates surface irregularities regardless of the original periderm color. Microcharcoals are abundant throughout the tissue; some of them are still recognizable in the inner layers (Figures 2a-c, h-i, k-n, g and 3d-f, j, q). Typically, it is possible to observe isolated clumps, or clumps attached to the tissue, within an unctuous, reddishbrown matrix, against which crystals and occluded micro - charcoals stand out. This pattern occurs in both extensive and restricted sections of tissue (Figures 2g-h, k-n and 3d-f, g-h, q). Cell wall cellulose birefringenceis retained or increased. In the cells of oca, birefringenceis wide spread , not restricted to cellulose accumulation (Figure 3n, r). Conducting tissues remain unaltered. Calcium crystals also remain largely unaltered, although their birefringence stands out (Figures 2r and 3n, p). In a few cases, partial melting of the druses' crystallites is observed (Figure 3i).

Figure 2. Appearance of the periderm and cortex cooked al rescoldo of: a-o) Solanum sp. and p-r) Ullucus tuberosus. a-c) Desireé, d-i) Abajeña, j-n) Malcacha, o) Huaico potato, p-r) Ulluco. The contiguous twin micrographs correspond to views with parallel (left) and crossed (right) nicols of the same sample.
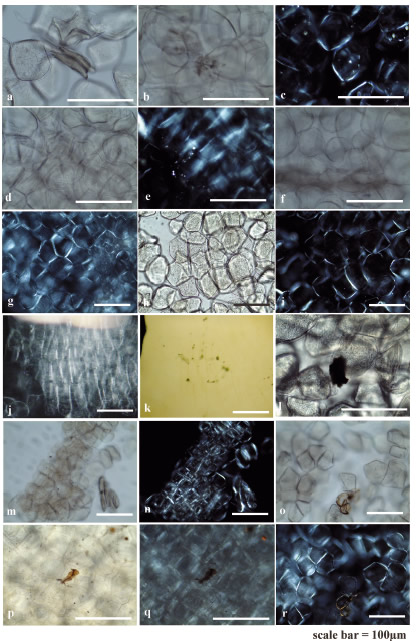
Figure 4. Appearance of parenchyma and medullary tissue of Solanum sp. cooked al rescoldo. a-c) Abajeña, d-g) Desireé, h-i) Malcacha, j-k) Pentaoca, l-r) Huaico potato. The contiguous twin micrographs correspond to views with parallel (left) and crossed (right) nicols of the same sample.
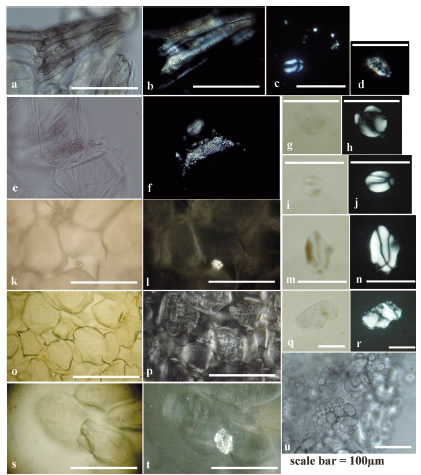
Figure 5. Starch grains, calcium crystals and conduction elements in parenchyma and medullary tissue cooked al rescoldo of: a-n) Solanum sp. and o-t) Oxalis tuberosa. Appearance of the fresh parenchyma and medullary tissue of Solanum sp. (Huaico potato) is in (u). a-c) Abajeña, d) Huaico potato, e-f) Desireé, g-n) Pentaoca, o-t) White oca. The contiguous twin micrographs correspond to views with parallel (left) and crossed (right) ni cols of the same sample.

Figure 7. Appearance of boiled parenchyma and medullary tissue of: a-h) Solanum sp. (Pentaoca) and i-n) Ullucus tuberosus. The contiguous twin micrographs correspond to views with parallel (left) and crossed (right) nicols of the same tissue.
Parenchyma and medullary tissues may retain their original structure. In other cases, cells remain close without completely attaching to each other, or are entirely detached. Their surfaces are dented, rough, contracted, cracked, and have lower relief than fresh internal tissues (Figures 4-5; Table 3). Despite being dehydrated, they retain a degree of turgor, though some are partially or fully collapsed (Figure 4a). Starches are in the process of melting/gelatinization (Figures 4e-g, j, o-r and 5o-p, s-t) or absent, giving the tissue the appearance of being almost totally empty (Figure 4c, i). Clumps (Figure 5s-t) and isolated grains of modified starch with damage from dry-cooking are occasionally observed (Table 3) (Figure 5g-j, m-n), and unaltered, resistant starch is scarce or virtually absent (Figure 5c, k-l). The birefringent cellulose of cell walls is retained. Although the crystals can maintain their location within the tissues (Figure 4c, e), clusters of multiple crystallites are often found in the intercellular spaces due to their expulsion from the interior of the cells (Figure 5e-f). Vessel elements are preserved unaltered , with high birefringence, appearing isolated or in clusters (Figure 5a-b).
Description of boiled tuber specimens
Periderm maintains its configuration and original morphology in some portions, although there is more fragmentation of the tissue than when cooking al rescoldo (Figure 6, Table 2). Highly fragmented, torn or folded sections are observed (Figures 6c, m-r). In general, boiling causes distension, loss of tissue turgor, an increase in size, and loss of relief. Tissues become transparent and noticeably thinner (Figure 6a-f). Colorless to gray silicified tissue and isolated polyhedral cells can be seen. If not detached, they stand out from surroundings tissues due to their higher relief (Figure 6e-f). A watery or diluted pigmentation remains unaltered or lost by sections in periderm cells (Figure 6e-f, I, m). Brown to reddish staining can appear by sections (Figure 6p). Birefringence of thickened cell wall cellulose is retained (Figure 6j, l, n, r). In pink oca, this phenomenon is widespread, as with cooking al rescoldo (Figure 6g). Calcium crystals may be unaltered and retain their position in the tissue (Figures 6m, q-r) or be released into the inter- or extracellular space (Figure 6k, q-r). Occasionally, crystals are found melted, formed into clumps in which the individuals are not discernable (Figure 6j-k). No micro-charcoals or carbonized tissues are observed in the boiling specimens.

Figure 6. Appearance of the boiled periderm and cortex of: a-d, g-l) Oxalis tuberosa, e-f) Solanum sp. and m-r) Ullucus tuberosus. a-d) White oca, e-f) Pentaoka, g-l) Pink oca, m-r) Ulluco. The contiguous twin micrographs correspond to views with parallel (left) and crossed (right) nicols of the same tissue.
Globular or polyhedral parenchymal and medullary tissue cells with rounded edges are present in clusters -although not attached to each other-, or disaggregated and dispersed, and there are empty net spaces between them (Figures 7-8, Table 3). In general, there is a loss of turgor, and cells appear more distended than when cooked al rescoldo or fresh. In sections more affected by boiling, cells appear collapsed due to the partial or total loss of starch (Figure 7a-e and 8jk). There are cracks and fractures in the cell walls; the surfaces are rough or dented, resembling those of tissues cooked al rescoldo (Figures 7a, e and 8n, r). Increased transparency and areas with watery or diluted reddish-brown staining are observed (Figure 7i and 8b-e). Some nodular particles in the same tones are attached to tissue (Figures 7i-j and 8a, c). Discontinuous masses of gelatinized, amorphous starch are observed in the interior or exterior of the tissues (Figure 8g-h, j-k). Resistant starch occurs as scarce, isolated individual grains (Figures 7c-e, h and 8g-i, l). Modified starch is in the process of gelatinization (Table 3), isolated (Figure 8no, r-s), forming clumps of individuals that have lost their original shape (Figures 7a-b and 8pq), and associated with starch exudates within the tissues or expelled from them (Figure 8j-k). The survival of resistant starch and of partially gelatinized individuals is greater in the close proximity of cortex and periderm than in the parenchymal and medullary tissue. Vessel elements within the vascular parenchyma preserve their birefringence and structure forming dense packages with a persistent linear orientation (Figures 7f-g and 8a, c-d, f). In some cases the crystals maintain their original location within the tissues (Figures 7i-j and 8f-h) or are expelled from them to the inter- or extracellular spaces (Figures 7k-n and 8m). Some are partially solubilized (Figure 7n).
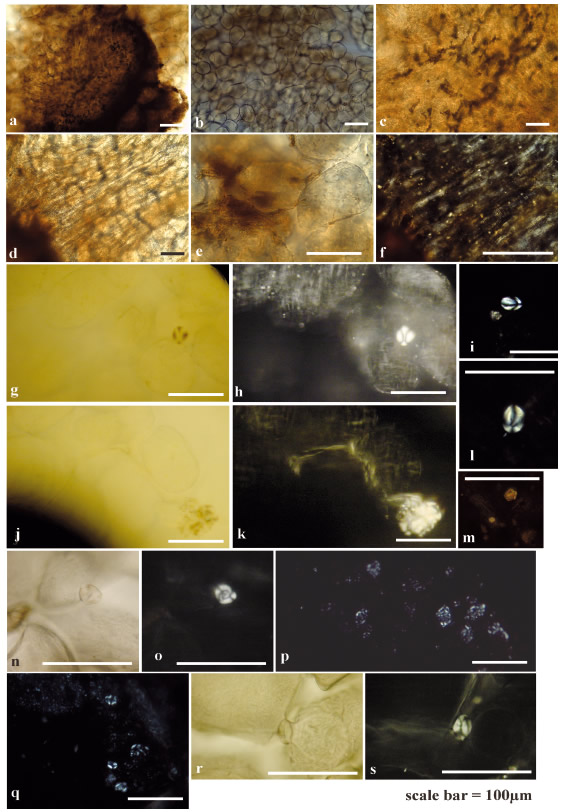
Figure 8. Appearance of boiled parenchyma and medullary tissue of Oxalis tuberosa. a-q) White Oca, r-s) Pink Oca. The contiguous twin micrographs correspond to views with parallel (left) and crossed (right) nicols of the same tissue.
DISCUSSION AND CONCLUSIONS
Experimental results indicate the potential for differentiating two cooking techniques: heating in humid and dry (or low moisture) environments. Based on previous studies, we expected these techniques to be similar in their effects on tubers since both involve the same agents of modification: heat and moisture. In the case of boiling, water is abundant in the cooking environment. By contrast, water is scarce but not totally absent in the "dry-cooking series" techniques. Among the latter, there is an increase in both the temperature gradient and exposure of the matter to temperature. While there are similarities in the effects of the cooking techniques reviewed, the abundance of water clearly leads to gelatinization, while its absence leads, in theory, to melting due to loss of starch humidity, although some degree of gelatinization is also possible. Generally speaking, certain reactions are common among tubers cooked al rescoldo and those heated in a moisturerich medium, the latter being a more aggressive transformation technique that compromises the integrity of plant tissues and particles to a corresponding degree. However, damage to the morphology and optical proper t ies of starch caused by cooking al rescoldo is greater than that caused by toasting, as reported elsewhere (Babot 2003). Such damage includes denaturation and a very low specimen count. In the case of cooking al rescoldo, this is influenced by the fact that plant matter is in direct contact with the heat source and subjected to higher temperatures than i n tasting . Although both groups of cooking techniques analyzed here cause similar modifications and damage, our results indicate that certain characteristics can be used to differentiate them and also, to distinguish cooked from fresh samples. For them to be applicable in archaeological contexts, we need to evaluate the survival over time of the characteristics reported here.
The periderm and cortex generally survive and retain their original fabric. They react differently to cooking al rescoldo (characterized by reddishbrown staining of tissues; unctuous clumps -isolated, or attached to tissue- with crystals and occluded micro-charcoals; contraction and tissue aging) versus boiling (characterized by diaphanous and thin material, partial preservation of the original watery color, tissue distension). However, the natural color of tuberous plants' shell could be confused with the effects of cooking on color and mistakenly attributed to anthropogenic modification. In the external tissues, massive carbonization occurs by sections, and the presence of micro-charcoals distributed throughout the tissue occurs only with cooking al rescoldo. Results obtained for the most external tissues are important for supplementing the data regarding the parenchyma and medullary tissue, where the starches are primarily located. While the latter are distinctively altered by particular processes and, therefore, play an important role in identifying anthropogenic processes, under certain circumstances they can be denatured or absent, making information about the periderm and cortex particularly relevant. Additionally, the periderm and cortex tissue can be preserved in archaeological contexts , both inside cooking containers where whole tubers were cooked and as discarded peelings (particularly those of bitter tuber varieties). The parenchyma and medullary tissue suffer greater damage than the periderm and cortex. Contraction occurs in cooking al rescoldo, whereas distension is observed with boiling. In the latter case, a diluted reddish-brown staining of the interior tissues may appear, as well as the occurrence of some nodular particles in the same tones. Observed starch alterations are consistent with those reported elsewhere, but a higher intensity of the modifications was verified when cooking al rescoldo, with respect to what was previously documented for toasting (Babot 2003). Despite the poor survival of starch as isolated, identifiable grains, both inside or outside the cooked mass, resistant starch specimens are more common in boiled tubers than in those cooked al rescoldo.
Both techniques result in modified starch grains. Calcium crystals, conducting tissues and cellulose survive both techniques. Melted or diluted crystals and crystals expelled from the tissues generally indicate manipulation in a medium with heat. Nonetheless these changes are more intense in the presence of water. Wet cooking is also more aggressive to cellulose than cooking al rescoldo. After cooking, cellulose allows excellent visualization of the original tissue fabric. Silica deposits, located in conduction and peridermal tissues and usually of little diagnostic value when taken alone, are not affected by the processes studied; on the contrary, they tend to stand out from the rest of the tissue, either in situ or detached from it. No significant differences in changes produced by the processing techniques considered here were observed between the three types of tubers and their varieties. Thus, the expectations generated should apply to a large number of tuberous plants and potentially other resources as well.
Finally, we must acknowledge that the daily character of kitchens and food preparation methods suggests intermediate instances that turn black and white results from controlled experiments into grays which cannot be ignored. For instance, tubers and other items might be cooked in their own liquid, without additional water, which, strictly speaking, is neither boiling nor toasting. Such exceptions and the integrity of the archaeological remains, determine the "grain" of the inferences that can made using experimental data. Despite these caveats, experimental work continues to contribute positively to our understanding of past practices.
Acknowledgments
To S. Hocsman, C. Codemo and A. Calisaya for providing the potatoes, ocas and ullucos analyzed in this study. To V. Bajales and J. Vildoza, who participated in sampling of the tubers. To anonymous reviewers for their useful comments on the content and language of this paper. To editors, K. Borrazzo and C. Weitzel for inviting us to publish in this volume and for their help with translation issues. This research was supported by projects CIUNT 26/G404 and PIPCONICET 464 directed by C. Aschero.
1 Resistant starch is defined as "[...] starch that is not digested in the small intestine of humans (Champ 2004; Biliaderis 2009). Useful term for explaining why some starches survive and others do not" (INSC 2011). In this paper, we used the term generically to refer to any starch grain preserved virtually unchanged.
2 We consider charring as a special cooking technique used in ritual performances (e.g., in feeding Pachamama) (Pazzarelli 2012).
REFERENCES
1. Babot, M. P. 2003 Starch grain damage as an indicator of food processing. In Phytolith and starch research in the Australian-Pacific-Asian regions: the state of the art, edited by D. M. Hart and L. A. Wallis, pp. 69-81. Terra Australis 19, The Australian National University, Canberra. [ Links ]
2. Babot, M. P. 2007 Granos de almidón en contextos arqueológicos: posibilidades y perspectivas a partir de casos del Noroeste argentino. In Paleoetnobotánica del Cono Sur: estudios de casos y propuestas metodológicas, compiled by M. B. Marconetto, M. P. Babot and N. Oliszewski, pp. 95-125. Museo de Antropología, Universidad Nacional de Córdoba, Córdoba. [ Links ]
3. Babot, M. P. 2009 Procesamiento de tubérculos y raíces por grupos agropastoriles del Noroeste argentino prehispánico: análisis de indicadores en residuos de molienda. In La alimentación en la América precolombina y colonial: una aproximación interdisciplinaria, compiled by A. Capparelli, A. Chevalier and R. Piqué, pp. 67-81. Treballs d'Etnoarqueologia 7, Instituto Milà y Fontanals, Consejo Superior de Investigaciones Científicas (CSIC), Madrid. [ Links ]
4. Babot, M. P. and E. Bru de Labanda 2005 Analysis of three factors that have an influence on the preservation of microfossils in archaeological artifacts. The Phytolitharien 17 (2): 4-5. [ Links ]
5. Babot, M. P. and M. C. Haros 2008 Interpreting content, context and manufacture from use-residues in ceramic vessels from Southern Argentinean Puna. In Abstracts of the 7th international Meeting on Phytolith Research, edited by M. Osterrieth, M. Fernández Honaine y N. Borrelli, pp. 44-45. Universidad Nacional de Mar del Plata, Mar del Plata. [ Links ]
6. Babot, M. P., S. Hocsman, R. E. Piccón Figueroa and M. C. Haros 2012 Recetarios prehispánicos y tradiciones culinarias. Casos de la Puna argentina. In Las manos en la masa. Arqueologías, antropologías e historias de la alimentación en Suramérica, edited by M. P. Babot, M. Marschoff and F. Pazzarelli, pp. 235-269. Instituto de Arqueología de Córdoba and Instituto Superior de Estudios Sociales, CONICET, Córdoba. [ Links ]
7. Babot, M. P. and M. A. Korstanje 2001 On starch taphonomy: some issues on physical, chemical and possible laboratory damage. In Program of the Conference: The state of the art in phytolith and starch research in the Australian-Pacific-Asian regions, pp. 7-9. Centre for Archaeological Research, Australian National University, Canberra. [ Links ]
8. Barton, H. and P. J. Matthews 2006 Taphonomy. In Ancient starch research, edited by R. Torrence and H. Barton, pp. 75-94. Left Coast Press, Walnut Creek. [ Links ]
9. Barton, H., R. Torrence and R. Fullagar 1998 Clues to stone tool function re-examined: comparing starch grain frequencies on used and unused obsidian artifacts. Journal of Archaeological Science 25: 1231-1238. [ Links ]
10. Beck, W. and R. Torrence 2006 Starch Pathways. In Ancient starch research, edited by R. Torrence and H. Barton, pp. 53-74. Left Coast Press, Walnut Creek.
11. Biliaderis, C. G. 2009 Structural transitions and related physical properties of starch. In Starch: Chemistry and Technology, 3rd ed., edited by J. R. BeMiller and R. L. Whistler, pp. 293-373, Academic Press, Amsterdam. [ Links ]
12. Borrazzo, K. 2006 Tafonomía lítica en dunas: una propuesta para el análisis de los artefactos líticos. Intersecciones en Antropología 7: 247-261. [ Links ]
13. Borrero, L. A. 2011 La función transdisciplinaria de la arqueozoología en el siglo XXI: restos animales y más allá. Antípoda 13: 267-274. [ Links ]
14. Castro, M. V. 2008. La papa (Solanum sp.): contexo social e ideológico en sus zonas de desarrollo originarias. Revista Chagual 6: 33-43. [ Links ]
15. Checa, A., A. Jimeno, J. Juan-Tresserras, J. P. Benito and A. Sanz 1999 Molienda y economía doméstica en Numancia. In Actas del IV Simposio sobre Celtíberos. Economía, pp. 63- 68. Institución "Fernando el Católico", CSIC, Zaragoza. [ Links ]
16. Coil, J., M. A. Korstanje, S. Archer and C. A. Hastorf 2003 Laboratory goals and considerations for multiple microfossil extraction in archaeology. Journal of Archaeological Science 30: 991-1008. [ Links ]
17. Cortella, A. R. and M. L. Pochettino 1994 Starch grain analysis as a microscopic diagnostic feature in the identification of plant material. Economic Botany 48 (2): 171-81. [ Links ]
18. Crowther, A. 2012 The differential survival of native starch during cooking and implications for archaeological analyses: a review. Archaeological and Anthropological Sciences 4 (3): 221-235. [ Links ]
19. French, D. 1984 Organization of starch granules. In Starch: Chemistry and Technology, edited by R. Whistler, J. BeMiller and E. Paschall, pp. 183-247. Academic Press, Florida. [ Links ]
20. Fullagar, R. 2006 Starch on artifacts. In Ancient starch research, edited by R. Torrence and H. Barton, pp. 177-204. Left Coast Press, Walnut Creek. [ Links ]
21. Gong, Y., Y. Yang, D. K. Ferguson, D. Tao, W. Li, C. Wang, E. Lüand and H. Jiang 2011 Investigation of ancient noodles, cakes and millet at the Subeixi Site, Xinjiang, China. Journal of Archaeological Science 38: 470-479. [ Links ]
22. González, M. I. and M. M. Frère 2004 Analysis of potsherd residues and vessel use in hunter-gatherer-fisher groups (Pampean region, Argentina). In Proceedings of the XIV UISPP Congress, pp. 27-36. BAR International Series 1270, Archaeopress, Oxford. [ Links ]
23. Hart, D. M. 2003 The influence of soil fauna on phytolith distribution in an Australian soil. In Phytolith and starch research in the Australian-Pacific-Asian regions: the state of the art, edited by D. M. Hart and L. A. Wallis, pp. 83-91. Terra Australis 19, The Australian National University, Canberra. [ Links ]
24. Hart, T.C. 2011 Evaluating the usefulness of phytoliths and starch grains found on survey artifacts. Journal of Archaeological Science 38: 3244-3253. [ Links ]
25. Haslam, M. 2004 The decomposition of starch grains in soils: implications for archaeological residue analyses. Journal of Archaeological Science 31: 1715-1734. [ Links ]
26. Haslam, M. 2006 An archaeology of the instant? Action and narrative in archaeological residue analyses. Journal of Social Archaeology 6 (3): 402-424. [ Links ]
27. Henry, A. G., H. F. Hudson and D. R. Piperno 2009 Changes in starch grain morphologies from cooking. Journal of Archaeological Science 36: 915-922. [ Links ]
28. Humphreys, G. S., D. M. Hart, N. Simons and R. J. Field 2003 Phytoliths as indicators of process in soils. In Phytolith and starch research in the Australian-Pacific- Asian regions: the state of the art, edited by D. M. Hart and L. A. Wallis, pp. 93-104. Terra Australis 19, The Australian National University, Canberra. [ Links ]
29. ICSN 2011 The International Code for Starch Nomenclature, http://www.fossilfarm.org/ICSN/Code.html (accessed 19/10/2011). [ Links ]
30. Jacobson, M. R., M. Obanni and J. N. BeMiller 1997 Retrogradation of starches from different botanical sources. Cereal Chemistry 74: 511-518. [ Links ]
31. Jones, P. J. 2009 A microstratigraphic into the longevity of archaeological residues, Sterkfontein, South Africa. In Archaeological science under a microscope. Studies in residue and ancient DNA analysis in honour of T.H. Loy, edited by M. Haslam, G. Robertson, A. Crowther, S. Nugent and L. Kirkwood, pp. 29-46. Terra Australis 30, Australian National University E-Press, Canberra. [ Links ]
32. Juan-Tresserras, J. 1992 Procesado y preparación de alimentos vegetales para consumo humano. Aportaciones del estudio de fitolitos, almidones y lípidos en yacimientos arqueológicos prehistóricos y protohistóricos del cuadrante NE de la Península Ibérica. PhD Dissertation, Universitat de Barcelona. [ Links ]
33. Korstanje, M. A. 2003 Taphonomy in the laboratory: starch damage and multiple microfossil recovery from sediments. In Phytolith and starch research in the Australian-Pacific- Asian regions: the state of the art, edited by D. M. Hart and L. A. Wallis, pp. 105-118. Terra Australis 19, The Australian National University, Canberra. [ Links ]
34. Korstanje, M. A. and M. P. Babot 2007 Microfossils characterization from south Andean economic plants. In Plants, people and places: recent studies in phytolith analysis, edited by M. Madella and D. Zurro, pp. 41-72. Oxbow Books, Cambridge. [ Links ]
35. Langejans, G. H. J. 2010 Remains of the day-preservation of organic microresidues on stone tools. Journal of Archaeological Science 37: 971-985. [ Links ]
35. Lantos, I., M. Maier and N. Ratto 2012 Recreando recetas: primeros resultados de una experimentación con variedades nativas de maíz del Noroeste argentino. In Las manos en la masa. Arqueologías, antropologías e historias de la alimentación en Suramérica, edited by M. P. Babot, M. Marschoff and F. Pazzarelli, pp. 527-552. Instituto de Arqueología de Córdoba and Instituto Superior de Estudios Sociales, CONICET, Córdoba. [ Links ]
36. Lewarch, D. and M. O´Brien 1981 The Expanding Role of Surface Assemblages in Archaeological Research. Advances in Archaeological Method and Theory 4: 297-342. [ Links ]
37. López, M. L., A. Capparelli and A. Nielsen 2011 Traditional post-harvest processing to make quinoa grains (Chenopodium quinoa var. quinoa) apt for consumption in Northern Lipez (Potosí, Bolivia): Ethnoarchaeological and archaeobotanical analyses. Journal of Anthropological and Archaeological Science 3 (1): 49-70. [ Links ]
38. Lovis, W. A., G. R. Urquhart, M. E. Raviele and P. Hart 2011. Hardwood ash nixtamalization may lead to false negatives for the presence of maize by depleting bulk ä13C in carbonized residues. Journal of Archaeological Science 38: 2726-2730. [ Links ]
39. Loy, T. 1994 Methods in the analysis of starch residues on prehistoric stone tools. In Tropical archaeobotany: Applications and new developments, edited by J. Hather, pp. 86-114. Routledge, New York. [ Links ]
40. Lu, T. 2003 The survival of starch residue in a subtropical environment. In Phytolith and starch research in the Australian-Pacific-Asian regions: the state of the art, edited by D. M. Hart and L. A. Wallis, pp. 119-126. Terra Australis 19, The Australian National University, Canberra. [ Links ]
41. Lyman, R. L. 1994 Vertebrate taphonomy. Cambridge Manuals in Archaeology, Cambridge University Press, Cambridge. [ Links ]
42. Messner, T. and B. Schindler 2010 Plant processing strategies and their affect upon starch grain survival when rendering Peltandra virginica (L.) Kunth, Araceae edible. Journal of Archaeological Science 37: 328-336. [ Links ]
43. Musaubach, M. G. and M. P. Babot 2014 Uso de las plantas entre los cazadores-recolectores pampeanos: estudio de microfósiles recuperados de tártaro dental humano, sitio Chenque 1. In El sitio Chenque. Un cementerio prehispánico en la Pampa Occidental. Estilo de vida e interacciones culturales de cazadores-recolectores del Cono Sur Americano, edited by M. Berón. Sociedad Argentina de Antropología, Buenos Aires, in press. [ Links ]
44. Musaubach, M. G. and M. A. Berón 2012 Cocinando en ollas en la Pampa Occidental. Datos desde la etnohistoria, el registro arqueológico y la arqueobotánica. In Las manos en la masa. Arqueologías, antropologías e historias de la alimentación en Suramérica, edited by M. P. Babot, M. Marschoff and F. Pazzarelli, pp. 599-620. Instituto de Arqueología de Córdoba and Instituto Superior de Estudios Sociales, CONICET, Córdoba. [ Links ]
45. Osterrieth, M., M. Madella, D. Zurro and M. F. Álvarez 2009 Taphonomical aspects of silica phytoliths in the loess sediments of the Argentinean Pampas. Quaternary International 193 (1-2): 70-79. [ Links ]
46. Osterrieth, M., M. Fernández Honaine, F. Álvarez, N. Borrelli and L. Benvenuto 2013 Procesos tafonómicos y silicofitolitos en secuencias pedoestratigráficas y pedoarqueológicas de distintos ambientes en Argentina. In Anais do Museu Nacional- UFRJ, Serie Livros 49: 218. Resúmenes del 5º Encontro Latinoamericano de Fitólitos. Rio de Janeiro. [ Links ]
47. Pardo, O. and J. L. Pizarro 2008 Alimentos: conservación y almacenamiento en el Chile Precolombino. Parina, Arica. [ Links ]
48. Parr, J. F., G. Kerr, J. Arthur and K. H. Taffs 2008 Impact of fire on peatlands in northeastern NSW Australia: implications for the interpretation of microfossil assemblages and fire histories. In Matices interdisciplinarios en estudios fitolíticos y de otros microfósiles, edited by M. A. Korstanje and M. P. Babot, pp. 47-53. BAR International Series 1870, Hedges, Oxford. [ Links ]
49. Pazzarelli, F. 2012 Arqueología de la comida. Cultura material y prácticas de alimentación en Ambato. PhD Dissertation, Universidad Nacional de Córdoba, Córdoba. [ Links ]
50. Piperno, D. M. 1988 Phytolith analysis: an archaeological and geological perspective. Academic Press, San Diego. [ Links ]
51. Pochettino, M. L. and M. C. Scattolin 1991 Identificación y significado de frutos y semillas carbonizados de sitios arqueológicos de la ladera occidental del Aconquija, Prov. de Catamarca, Rca. Argentina. Revista del Museo de La Plata (nueva serie) Antropología 9 (71): 169-181. [ Links ]
52. Radley, J. A. 1943 Starch and its derivatives. 2nd. edition, Chapman and Hall, London. [ Links ]
53. Raviele, M. 2011 Experimental assessment of maize phytolith and starch taphonomy in carbonized cooking residues. Journal of Archaeological Science 38 (10): 2708-2713. [ Links ]
54. Reichert, C. T. 1913 The differentiation and specificity of starches in relation to genera, species, etc. Publication 173, Carnegie Institution of Washington D.C., Washington D.C. [ Links ]
55. Samuel, D. 2006 Modified starch. In Ancient starch research, edited by R. Torrence and H. Barton, pp. 205-216. Left Coast Press, Walnut Creek. [ Links ]
56. Tassara, G. and M. Osterrieth 2008 Silicofitolitos en artefactos de molienda de sitios arqueológicos del Área Interserrana, Buenos Aires. Un estudio preliminar. In Matices interdisciplinarios en estudios fitolíticos y de otros microfósiles, edited by M. A. Korstanje and M. P. Babot, pp. 163-171. BAR International Series 1870, Hedges, Oxford. [ Links ]
57. Therin, M. 1994 Subsistence through starch: the examination of subsistence changes on Garua Island, West New Britain, Papua New Guinea, through the extraction and identification of starch from sediments. BA (Honours) Thesis, Sydney University, Sidney. [ Links ]
58. Torrence, R. and H. Barton (editors) 2007 Ancient starch research. Left Coast Press, Walnut Creek. [ Links ]
59. Wandsnider, L. A. 1997 The roasted and the boiled: food composition and heat treatment with special emphasis on pit-hearth cooking. Journal of Anthropological Archaeology 16: 1-48. [ Links ]
60. Whistler, R. L., J. BeMiller and E. Paschall (editors) 1984 Starch: Chemistry and Technology. Academic Press, Florida. [ Links ]
61. Williams, M. R. and P. Bowler 1982 Starch gelatinization: A morphological study of Triticeae and other starches. Starch/Stärke 34: 221-223. [ Links ] [ Links ]

