










Services on Demand
Journal

Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO
Related links
-
 Similars in
SciELO
Similars in
SciELO
Share

 Permalink
PermalinkIntersecciones en antropología
On-line version ISSN 1850-373X
Intersecciones antropol. vol.16 no.1 Olavarría June 2015
ARTÍCULOS
Explotación de fauna dulceacuícola en el valle inferior del río Chubut (Patagonia central) durante el Holoceno tardío
Ariadna Svoboda y Julieta Gómez Otero
Ariadna Svoboda. Centro Nacional Patagónico (CENPAT) y Consejo Nacional de Investigaciones Científicas y Técnicas (CONICET). Boulevard Brown 2915, Puerto Madryn (9120), Chubut, Argentina. E-mail: svoboda@cenpat-conicet.gob.ar
Julieta Gómez Otero. CENPAT-CONICET. Universidad Nacional de la Patagonia San Juan Bosco (UNPSJB). Boulevard Brown 2915, Puerto Madryn (9120), Chubut, Argentina. E-mail: julieta@cenpat-conicet.gob.ar
Recibido 3 de abril 2013.
Aceptado 18 de septiembre 2013
RESUMEN
En este trabajo se presentan y discuten los resultados de la ampliación del estudio arqueofaunístico de dos sitios arqueológicos del valle inferior del río Chubut cuya cronología se extiende entre 1500 y 1200 años AP. Estos contextos se destacan por el aprovechamiento de fauna dulceacuícola (peces, aves y coipos) por sobre el guanaco, el recurso principal para los cazadores-recolectores de Patagonia. Se discute su relación con procesos de intensificación económica y factores situacionales, como inundaciones repentinas y catastróficas.
Palabras clave: Explotación de fauna dulceacuícola; Cazadores-recolectores; Valle inferior del río Chubut; Holoceno tardío.
ABSTRACT
Evidence of freshwater faunal exploitation in the lower valley of the Chubut River, central Patagonia, during the late holocene.
This article presents and discusses the results of a new archaeofaunal study of two archaeological sites from the lower valley of the Chubut River, dated to 1500 -1200 BP. These assemblages stand out because of the exploitation of freshwater fauna (fish, birds and coipo) over the guanaco, the main resource for hunters-gatherers from Patagonia. The relationship between the faunal remains, intensification processes, and situational factors, such as sudden and catastrophic river floods, is discussed.
Keywords: Freshwater faunal exploitation; Hunter-gatherers; Lower Valley of the Chubut River; Late Holocene.
INTRODUCCIÓN
El estudio arqueológico de la fauna menor aporta valiosa información para discutir aspectos paleoambientales y también conductuales tales como la amplitud del espectro alimenticio, la estacionalidad de las ocupaciones y los métodos de captura (Wheeler y Jones 1989; Stahl 1996; síntesis en Cruz et al. 2007). Para Patagonia septentrional y central se cuenta con trabajos que consignan la existencia de restos de fauna menor dulceacuícola (moluscos, peces, aves y coipos) en sitios del Holoceno tardío localizados en ambientes fluviales y lacustres. En algunos trabajos sólo se mencionó su presencia (e.g., Arrigoni 1991; Menegaz 1996; Silveira 1996); en otros se infirió aprovechamiento humano. La explotación de recursos dulceacuícolas fue determinada para sitios del curso inferior del río Colorado (Martínez et al. 2005; Martínez 2008- 2009; Stoessel 2012); el valle medio del río Negro (Prates 2008; Prates y Acosta Hospitaleche 2010); el bosque y ecotono bosque-estepa de la provincia de Río Negro (Hajduk y Albornoz 1999; Hajduk et al. 2004) y la cuenca del Lago Musters (Moreno y Pérez Ruiz 2010). No obstante, la presencia de restos de peces también podría deberse a transporte y acumulación por carnívoros, tal como lo planteó Fernández (2008) para el área de Piedra Parada. En los sitios donde se ha comprobado la intervención humana, la abundancia relativa de la fauna dulceacuícola es baja y prevalecen los recursos terrestres. La excepción la configuran tres sitios del Holoceno tardío. Uno es El Tigre, ubicado en el curso inferior del río Colorado (Martínez et al. 2005); los otros -Loma Grande 1 (Gómez Otero 1994) y Cinco Esquinas 1 (Gómez Otero et al. 2010)- están emplazados en el valle inferior del río Chubut. Las explicaciones para estos contextos difieren: el primer caso fue vinculado con un proceso de diversificación dietaria (incorporación de recursos menores) y de intensificación en la explotación de peces, guanaco y vegetales (Martínez 2008-2009; Stoessel 2012).
En cambio, para los dos sitios del valle inferior del río Chubut se hipotetizó una explotación oportunista de los recursos fluviales -percas, aves acuáticas y coipos- como una estrategia de adecuación a grandes crecidas del río que impedían el acceso a las presas terrestres, especialmente guanacos (Gómez Otero 1994; Gómez Otero et al. 2010). En función de lo anterior, los objetivos principales de este trabajo son: 1) presentar los resultados de un nuevo y más amplio estudio arqueofaunístico de los sitios Loma Grande 1 y Cinco Esquinas 1 del valle inferior del río Chubut; 2) identificar los procesos de formación y los agentes posdepositacionales; y 3) discutir las implicaciones de la evidencia en relación con procesos de intensificación económica en el Holoceno tardío y estrategias oportunísticas como respuesta a inundaciones catastróficas y repentinas.
EL ÁREA DE ESTUDIO Y MARCO ECOLÓGICO
El área de estudio corresponde al valle inferior del río Chubut (VIRCH), que se inicia en la cola del embalse Florentino Ameghino y desemboca en Bahía Engaño (Figura 1). El clima es árido y ventoso, con precipitaciones medias anuales de 200 mm (Beeskow et al. 1987). Fitogeográficamente corresponde a la subunidad del Monte Austral de la Provincia del Monte (León et al. 1998), dominada por jarilla (Larrea sp.), quilimbay (Chuquiraga sp.) y varias especies de algarrobo, principalmente Prosopis alpataco (Beeskow et al. 1987). La fauna pertenece al dominio Patagónico (Cabrera y Yepes 1960). Entre las aves terrestres sobresale el choique (Pterocnemia pennata) y entre los mamíferos, el guanaco (Lama guanicoe). También hay mamíferos terrestres de menor porte, como el gato montés (Felis geoffroyi), los zorros gris y colorado (Pseudalopex sp.), el peludo (Chaetophractus villosus), el piche (Zaedyus pichiy), la mara (Dolichotis patagonum) y microroedores (Ctenomys sp., Microcavia australis, Galea leucoblephara, entre otros). En lo que respecta a los recursos dulceacuícolas, la fauna íctica nativa presenta un ensamble de baja diversidad específica compuesto principalmente por perca (Percichthys trucha) (sensu Ruzzante et al. 2011), pejerrey patagónico (Odontesthes hatcheri) y bagres (Diplomystes sp. y Hatcheria macrei) (Gosztonyi 1988). Las aves acuáticas incluyen patos (Anas spp.), cisnes (Cygnus sp.), cauquén (Chloephaga sp.), biguá (Phalacrocorax olivacius), gallareta (Fulica sp.), garza blanca (Egretta alba) y flamenco austral (Phoenicopterus chilensis). Entre los mamíferos se destaca el coipo (Myocastor coypus).
DESCRIPCIÓN DE LOS SITIOS
Los sitios Loma Grande 1 y Cinco Esquinas 1 se ubican en el segmento oriental del VIRCH (Figura 1). En este sector, la geomorfología se caracteriza por una llanura aluvial donde predominan áreas topográficas bajas e inundables, limitadas lateralmente por terrazas que conforman escalones de escasa altura relativa. Antes de la construcción del dique Florentino Ameghino, en 1960, las inundaciones en el río eran frecuentes. Según los colonos galeses, entre fines del siglo XIX y principios del XX tuvieron lugar cuatro crecidas repentinas y catastróficas, que anegaron todo el valle arrasando con las viviendas y los sembrados (Matthews 1992; Hughes 1993). Las inundaciones ocurrían entre junio y julio, y el valle permanecía inundado hasta septiembre o noviembre, pero podían quedar lagunas ribereñas hasta cuatro años después (Hughes 1993: 176). Los colonos se refugiaban en las lomas de la margen norte hasta que el nivel del río descendía. Por su parte, el ganado vacuno y caballar era arrastrado por la corriente o huía fuera del valle (Matthews 1992: 57). Estudios hidrogeológicos para los últimos 100 años determinaron una periodicidad de 4,7 a 8 año para las crecidas altas (desborde de los sectores ribereños), de 8 a 10,4 años para las extraordinarias (inundación completa del valle) y una altura máxima que no superó los 5,50 m (Pronsato 1950).
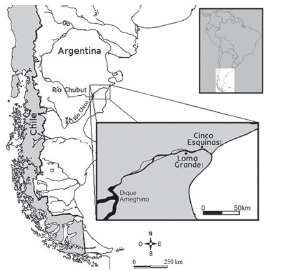
Figura 1. Área de estudio y localización de los sitios arqueológicos.
Loma Grande 1 (LG1)
Se trata de una localidad a cielo abierto (300 x 100 m) ubicada sobre un albardón a 10 msnm, a unos 500 m del cauce actual del río y a 30 km de la costa marina. Se identificaron distintos rasgos de ocupación: entierros humanos múltiples, materiales arqueofaunísticos y líticos en capa y en superficie, y alta densidad y concentración de huellas de fogón (Gómez Otero 1994). Sobre un sector con huellas de fogones se planteó un sondeo de 1 m x 1,50 m de lado y se determinaron dos niveles estratigráficos:
Nivel 1 (0 - 6 cm): sedimento muy fino limoarcilloso, concrecionado, con granos de cuarzo. Presentaba espículas de carbón que le conferían un color castaño-grisáceo oscuro. Se observaron manchones de tierra quemada y dos fogones de límites definidos: el Fogón 1 (30 x 19 cm) y el Fogón 2 (31 x 39 cm). En su interior se registraron restos arqueofaunísticos, microdesechos de talla (algunos con evidencias de estrés térmico), piedras termóforas y dos pequeños molinos planos en la base del Fogón 1.
Nivel 2 (6 - 13,5 cm): sedimento fino más suelto, color castaño, con un manchón color negro compactado en su parte inferior, que sería una huella de fogón.
Se obtuvieron tres dataciones radiocarbónicas: 1390 ± 60 AP (LP-844; hueso; ä13C= -20) y 1400 ± 50 AP (LP-1028; hueso; ä13C= -20) para dos individuos humanos, y 1210 ± 60 AP (LP-1641; carbón; ä13C= -25) para el Nivel 1 del Fogón 1.
Cinco Esquinas 1 (CE1)
Cinco Esquinas es una localidad arqueológica a cielo abierto que se emplaza en la Chacra 84 sobre un relieve de albardones a 10 msnm, a 800 m del cauce actual del río y a unos 18 km de la costa marina. Pronsato (1950) destaca que esta chacra y la 85 permanecieron como áreas secas durante los grandes eventos de inundación de 1932 y 1945. El lugar muestra importante impacto antrópico (Gómez Otero et al. 2010) a causa de su uso como cantera de áridos y fábrica de ladrillos. Al igual que en LG1, se reconocieron entierros humanos múltiples, numerosas huellas de fogón y materiales arqueofaunísticos, líticos y cerámicos. Todos estos contextos estaban parcialmente expuestos en superficie por erosión eólica e hídrica (pluvial), o por acción antrópica, en el caso de los restos humanos. En dos campañas (1996 y 2009) se efectuaron muestreos y excavaciones en diferentes sectores (Gómez Otero et al. 2010) -ver Materiales y métodos-. En 1996 se excavó una cuadrícula de un metro de lado sobre una huella de fogón del Sector 1 (F1S1), y se reconoció un único nivel cultural en una profundidad máxima de 13,5 cm. El sedimento asociado era arcilloso, estaba muy compactado y mezclado con pequeños carbones. Se rescataron materiales faunísticos, carbones y escasos microdesechos de talla. En 2009 se trabajaron los sectores 2 y 3. En el sector 2 se realizó un muestreo sistemático de superficie de cuatro metros de lado (M1S2), que aportó restos de fauna y artefactos líticos. En el sector 3 se excavaron el Fogón 2 (F2S3) y la Concentración Cerámica 1 (CC1). Para la excavación del Fogón 2 (de 30 x 70 cm de superficie) se planteó una cuadrícula de un metro de lado y en una potencia máxima de 10 cm se identificó una única capa estratigráfica del mismo tipo que la de F1S1. En ese fogón se registraron restos faunísticos y artefactos líticos. La concentración cerámica fue relevada mediante dos cuadrículas consecutivas (A y B) de un metro de lado. Los materiales faunísticos estaban concentrados en la cuadrícula A, asociados con pequeños carbones y escasos tiestos.
Se obtuvieron dos dataciones: 1560 ± 90 (LP-2282; carbón; ä13C= -24) para la concentración cerámica y 1260 ± 90 (LP-2294; hueso; ä13C= -20) para un individuo humano hallado en el Sector 2 (Gómez Otero et al. 2010). Se interpretó que el lugar fue usado como campamento residencial (Gómez Otero et al. 2010).
MATERIALES Y MÉTODOS
Los materiales arqueofaunísticos aquí analizados provienen de los siguientes contextos: los fogones 1 y 2 de LG1, y todos los muestreos (F1S1, F2S3, CC1A y M1S2) de CE1. Como se aclaró antes, este trabajo es una ampliación de estudios preliminares llevados a cabo por Gómez Otero y colaboradores (Gómez Otero 1994; Gómez Otero et al. 2010). En la excavación se tomaron tridimensionales y se usaron zarandas de malla fina: 2 mm en LG1 y de 1,3 mm en CE1. Dada la frecuencia de concreciones sedimentarias por el alto contenido de limo y arcilla, en laboratorio se aplicó el método de levigación en agua, que permite la flotación de los materiales y/o su separación del sedimento que los contiene. El análisis faunístico consistió en la identificación taxonómica y anatómica, para la que se utilizó la colección de referencia del Laboratorio de Arqueología del Centro Nacional Patagónico (Puerto Madryn). En los casos en que sólo se pudo determinar la Clase, los especímenes fueron asignados a categorías discriminadas de acuerdo con el peso. Sobre la base de la propuesta de Mengoni Goñalons (1999: 42), los mamíferos se clasificaron en: pequeño (< 3 kg), mediano- pequeño (entre 3 y 15 kg; aquí se incluyó al coipo y la mara); mediano-grande (entre 15 y 50 kg) y grande (> 50 kg). Para las aves se empleó la clasificación de Giardina (2012): pequeña (Columbidae y Passeriformes), mediana (la mayoría de las aves acuáticas, terrestres y marinas) y grande (choique). Se estimaron el NISP (Número de especímenes identificado por taxón), el NUSP (Número de especímenes no identificados) y el NSP (NISP + NUSP) (Lyman 2008). Para la determinación del MNI de mamíferos y aves se consideraron la lateralidad y/o el estado de fusión (Mengoni Goñalons 1999). Para la abundancia anatómica absoluta y relativa se emplearon las unidades derivadas MNE y MAU%, y se contemplaron la lateralidad y las zonas diagnósticas (Binford 1984; Mengoni Goñalons 1999). Las mismas unidades de cuantificación se aplicaron para los peces según la metodología de Zangrando (2009: 134-140). La primera clasificación de restos de peces de LG1 fue realizada en 1993 por el Dr. Atila Gosztonyi, ictiólogo del CENPAT, quien determinó un MNI de 19 percas, a partir de la primera vértebra (Gómez Otero 1994: 67); lamentablemente esos restos se extraviaron.
Se evaluaron también los procesos de formación del conjunto y los posdepositacionales a través del análisis de distintas variables tafonómicas a toda la muestra (NSP) (Behrensmeyer 1978; Lyman 1994, entre otros). Mediante observaciones macroscópicas y microscópicas de hasta 50x con una lupa trinocular Karl Zeiss, se exploraron trazas sobre la superficie ósea para determinar la acción de distintos agentes naturales y antrópicos (Lyman 1994). Respecto del estado de meteorización ósea se contemplaron los atributos definidos por Behrensmeyer (1978) para mamíferos mayores a 5 kilos; por Muñoz y Savanti (1998) para aves, y por Svoboda y Moreno (2014) para perca. En los especímenes menores a tres centímetros y/o termoalterados no se midió esta variable. La determinación del grado de alteración térmica tuvo en cuenta la apariencia macroscópica a partir del color (Shipman 1984, en Mengoni Goñalons 1999). La extensión de la fragmentación se evaluó a partir de la proporción de especímenes anatómicamente completos (Lyman 1994: 333, 2008). Para la identificación de huellas y fracturas de origen antrópico se consideraron el tipo y su ubicación en el hueso (Mengoni Goñalons 1999). Finalmente, la estación de captura fue estimada sobre la base de la propuesta de Kaufmann (2009) para guanaco y la lectura de otolitos de perca (Svoboda 2013a).
RESULTADOS
Loma Grande 1
Preservación del conjunto
Se destaca el alto porcentaje de fragmentación de la muestra total (NSP = 2918), que alcanza tanto a los especímenes identificados taxonómica y anatómicamente (NISP = 1035; 46%) como a los no determinados (NUSP = 1883; 54%) (Tabla 1). El 80% del NUSP corresponde a especímenes menores a un centímetro, y el resto, a fragmentos de entre uno y tres centímetros. Entre los especímenes identificados la extensión de la fragmentación es diferencial según los taxa: la proporción de elementos completos en los peces es de 55%, y en las aves y los mamíferos mediano-pequeños apenas llega a 11%. Independientemente del taxón, el grado de meteorización es bajo a nulo (estadios 0 a 1) (Tabla 2): los restos de peces no muestran fisuras ni exfoliaciones significativas; los de aves y mamíferos presentan el tejido intacto y sin agrietamientos. Si bien no se evaluó la meteorización en los especímenes de menos de tres centímetros, su estado de preservación es similar al de los identificados taxonómicamente. Las improntas de raíces se registraron sólo en 39 especímenes. No se observaron marcas de masticado de carnívoros (e.g., pozos, hoyuelos, etc.) (Binford 1981), acción de roedores (surcos cortos y profundos), corrosión gástrica (Andrews 1990) o abrasión geológica (Gutiérrez 2001).
Tabla 1. Abundancia taxonómica de los conjuntos faunísticos de Loma Grande 1 y Cinco Esquinas 1.
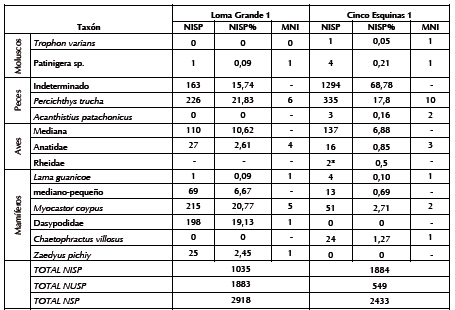
Nota: *= corresponden a fragmentos de cáscaras de huevo.
Diversidad y abundancia taxonómica
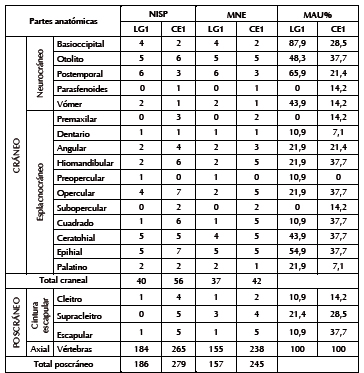
Entre los 2918 restos (NSP) se identificó una valva completa del gasterópodo marino (Patinigera sp.), dos fragmentos de cáscaras de huevo de origen indeterminado (que por densidad, grosor y tamaño no corresponderían a choique, sino a un ave mediana), y 2915 especímenes óseos, de los cuales 1035 fueron determinados taxonómicamente (Tabla 1). En la muestra predomina P. trucha (NISP = 226), cuyo MNI es de seis (calculado a partir de la primera vértebra); pero ese total se elevaría a 25 si se suman los 19 individuos calculados anteriormente por el Dr. Gosztonyi a partir del mismo elemento. Las tallas estimadas van de 22,7 a 35,3 cm de largo total (Svoboda 2013b). Esta especie está representada por todos los elementos del esqueleto craneal y poscraneal, de los cuales el segmento axial es del de mayor abundancia relativa (Tabla 3). Además se identificaron las espinas de la aleta dorsal y anal. Los restos de peces indeterminados (NISP = 163) corresponden a elementos con bajo grado de identificabilidad taxonómica: costillas, pterigióforos y radios de las aletas. Dado que P. trucha es la única especie íctica determinada, estos especímenes podrían atribuirse a ella.
Tabla 2. Principales variables tafonómicas.
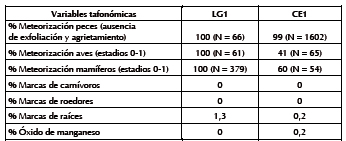
Tabla 3. Medidas de abundancia anatómica de P. trucha.
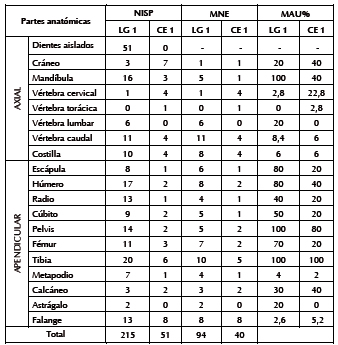
Se estableció la presencia de Anatidae (NISP = 27) con un MNI de cuatro (estimado a partir del húmero proximal izquierdo) y de aves medianas indeterminadas (NISP = 110) con un MNI de dos (contabilizado a partir del coracoides y la escápula). Entre los anátidos los elementos más frecuentes corresponden al esqueleto apendicular (especialmente húmero y coracoides); mientras que el cráneo y el resto del esqueleto axial están ausentes (Tabla 4). Entre las aves medianas predomina el esqueleto apendicular (sólo tres vértebras caudales) y la proporción de cilindros es alta (n = 79). Entre los mamíferos, M. coypus es el más abundante (NISP = 215). Se estimaron cinco individuos sobre la base de la mandíbula. Está representado todo el esqueleto, con predominio de la mandíbula y las extremidades (excepto el autopodio) (Tabla 5). Algunas carillas de vértebras (n = 12), astillas (n = 25) y otros fragmentos indeterminados (n = 32) -adjudicados a mamífero mediano-pequeño (NISP = 69)- corresponderían a esta especie. Por último, se asignaron a Z. pichiy placas de bandas móviles y una hemimandíbula (NISP = 25); a Dasypodidae varios escudos (NISP = 198) y a L. guanicoe un tarsiano.
Tabla 4. Medidas de abundancia anatómica de Anatidae.
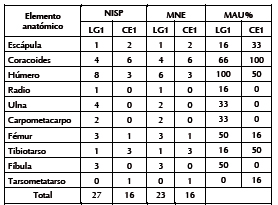
Tabla 5. Medidas de abundancia anatómica de M. coypus.
Evidencias de procesamiento y termoalteración
Del NSP sólo se identificaron huellas en 14 especímenes de Anatidae y M. coypus (Tabla 6). Entre los anátidos se reconocieron evidencias de machacado en una diáfisis distal de radio; también, huellas de corte transversales en una diáfisis distal de fémur y un coracoides, y otras oblicuas y agrupadas en una diáfisis proximal de húmero (Figura 2a y b). Asimismo, se detectaron fracturas transversales en nueve cilindros (radio, ulna y tibiotarso) y marcado perimetral en dos cilindros de radio. Con relación a M. coypus, se hallaron huellas de corte en la mandíbula, fémur, escápula, tibia y astillas (Tabla 6 y Figura 3). A su vez, se observaron fracturas transversales en dos radios, cinco cúbitos, tres fémures, cuatro tibias y dos metapodios, fracturas helicoidales en una tibia y un radio, y marcado perimetral en un metapodio.
Tabla 6. Alteraciones antrópicas en la superficie ósea de distintos taxa. Número de Especímenes Identificados por Taxón con: huellas de corte (NISPc) y con evidencias de alteración térmica (NISPt).
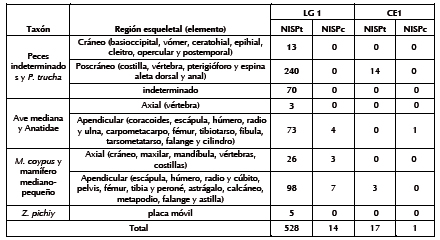
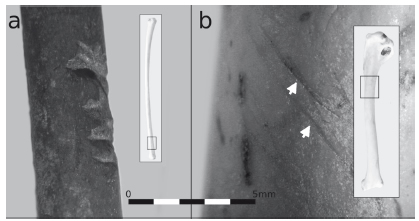
Figura 2. Huellas de procesamiento en Anatidae: a) machacado en radio distal; b) corte en diáfisis proximal de húmero.
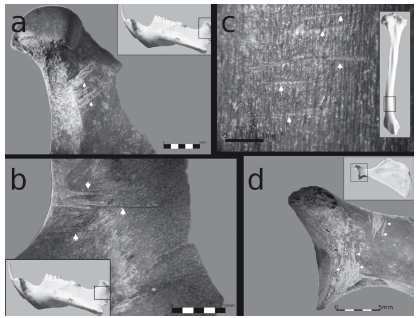
Figura 3. Huellas de procesamiento en M. coypus: a y b) en mandíbula; c) en diáfisis distal de tibia; d) en escápula.
Del total de especímenes, 1959 (67%) presentan evidencias de combustión: 528 (51%) de los 1035 identificados y 1431 (76%) de los 1883 no identificados. La acción del fuego afectó a todos los taxa y a todas las partes del esqueleto (Tabla 6). En P. trucha y peces indeterminados la termoalteración alcanzó a 323 especímenes (83%), de los cuales un 75% está quemado (color rojizo); 17%, carbonizado (color negro); y sólo 7%, calcinado (grisazulado). En los anátidos y aves medianas se observaron evidencias de combustión en 76 especímenes (59%); entre ellos, el 67% está quemado; el 18%, carbonizado; y el 14%, calcinado. La termoalteración es pareja y completa en todos los huesos. La excepción está dada por cinco fragmentos de Anatidae -tres húmeros, dos ulnas y un fémur- que están afectados sólo en los extremos de la diáfisis. Con relación a M. coypus, la acción del fuego impactó en 124 especímenes (44%): 33%, quemado; 37%, carbonizado; y 29%, calcinado. Al igual que en las aves, la combustión cubrió la totalidad de cada espécimen, salvo una costilla, un radio y un cúbito. Por último, sólo se registraron cinco placas quemadas de Z. pichiy, mientras que el tarsiano de guanaco no presenta huellas de procesamiento ni de combustión.
Estacionalidad de ocupación
Las bandas de crecimiento anual de dos otolitos de P. trucha presentan el anillo del borde opaco, lo que señala captura en primavera- verano (Svoboda 2013a). La presencia de cáscaras de huevo, probablemente Anatidae, refuerza la determinación de estacionalidad.
Cinco Esquinas 1
Preservación del conjunto
A diferencia de LG1, los especímenes de todos los muestreos (NSP = 2433) presentan baja fragmentación, lo que permitió identificar el 77% (NISP = 1884) contra el 23% (NUSP = 549) de indeterminados (Tabla 1). Al igual que en el sitio anterior, la magnitud de la fragmentación varió según los taxa, aunque es menor en las aves y los mamíferos: la proporción de elementos completos es de 45% en los peces, 22% en los mamíferos mediano-pequeños y 18% en las aves. En cuanto a la meteorización (Tabla 2), en los peces no se vio agrietamiento ni exfoliación, y la mitad de los especímenes de aves y mamíferos presenta buena preservación (estadios 0 y 1). El resto de aves y mamíferos, concentrados en M1S2, presenta blanqueamiento por acción solar, y líneas y grietas de desecación y exfoliación de la superficie cortical (estadios 2 y 3). Son muy pocas las evidencias de actividad de raíces (n = 7) y dendritas de manganeso (n = 7); tampoco se registraron corrosión gástrica, abrasión geológica ni marcas de carnívoros y roedores (Tabla 2).
Diversidad y abundancia taxonómica
Entre los 2433 del NSP se contabilizaron cinco fragmentos de valvas marinas (géneros Patinigera y Trophon), dos cáscaras de huevo de rheido y 2426 especímenes óseos, de los cuales, 1884 fueron identificados taxonómicamente y corresponden principalmente a restos ictioarqueológicos (Tabla 1). La distribución de restos por muestreo es numéricamente diferente: F2S3 (n = 2005), F1S1 (n = 221), M1S2 (n = 197) y CC1A (n = 10). Los restos de peces se concentran en F2S3 (n = 1580), los de aves en F1S1 (n = 83) y M1S2 (n = 72), los de coipo y mamífero medianopequeño en F1S1 (n = 40), M1S2 (n = 24) y F2S3 (n = 1), los armadillos sólo en F1S2 (n = 24), y los de guanaco en M1S1 (n = 4).
Entre los peces se destaca P. trucha (NISP = 335) con un MNI de diez (estimado a partir de la primera vértebra), y tallas estimadas de entre 24 y 45 cm de largo total (Svoboda 2013b). Si bien se encuentra la totalidad de su esqueleto, hay dominio de la región axial (Tabla 3), que se presenta frecuentemente articulada. También se determinaron dos individuos de la especie marina Acanthistius patachonicus (mero) a partir del otolito izquierdo. La mayoría de los especímenes de peces (NISP = 1294) no pudo ser determinada porque c orresponde a elementos poco diagnósticos: fragmentos de costillas, pterigióforos y radios de aletas. En relación con las aves, se identificaron escasos especímenes de Anátidos (NISP = 16) pero con un MNI proporcionalmente elevado de cuatro ejemplares, calculado a partir del coracoides. En términos anatómicos, en los Anátidos se destacan los elementos del esqueleto apendicular como el coracoides, húmero y tibiotarso (Tabla 4). Los demás especímenes corresponden a ave mediana indeterminada (NISP = 137), representada por elementos diagnósticos como esternón, coracoides, costillas, carpometacarpo, tarsometatarso y falanges; cilindros (n = 61) y fragmentos indeterminados (n = 50). Es muy probable que los restos asignados a ave mediana correspondan a Anatidae.
En cuanto a los mamíferos, predomina M. coypus (NISP = 51), del que se estimó un mínimo de dos individuos sobre la base de la hemipelvis. Se hallan presentes todas las partes anatómicas, con una supremacía del cráneo y el esqueleto apendicular (Tabla 5). Vértebras y falanges asignadas a mamífero medianopequeño (NISP = 13) podrían pertenecer a esta especie. Le siguen en abundancia los restos de C. villosus (NISP = 24), ejemplificado por un húmero y placas móviles, y de L. guanicoe (NISP = 4), representados por una pelvis no fusionada, una costilla fragmentada y una astilla que, a juzgar por su tamaño y densidad ósea, pertenecerían a este taxón (Gómez Otero et al. 2010).
Evidencias de procesamiento y termoalteración
Sólo se identificaron cortes en una astilla de diáfisis de mamífero mediano-pequeño, probablemente coipo. Es muy baja la proporción de modificaciones producidas por combustión: sólo 25 (1,02%) del NSP. Los especímenes afectados provienen en su mayoría del Fogón F2S3, y en los casos en que pudieron ser identificados (n = 15), corresponden a elementos carbonizados y calcinados del poscráneo de peces (costilla y vértebra) y a una pelvis de coipo parcialmente carbonizada (Tabla 6). En el M1S2 se detectó carbonización en una epífisis distal de fémur y una falange de coipo. También se halló una placa de peludo calcinada en la concentración cerámica (CC1). No se identificaron termoalteraciones en el resto de los taxa. En cuanto al patrón de fractura, 13 especímenes de ave mediana y cinco de mamífero mediano-pequeño presentan fracturas astilladas. En las aves, sólo una fractura es del tipo helicoidal; no se determinaron atributos relacionados con percusión. Por último, entre los no identificados (astillas de menos de tres centímetros) no se registraron huellas de corte ni de impacto. Estacionalidad de ocupación La lectura de los anillos de crecimiento anual de los otolitos de P. trucha del F2S3 señala muerte en otoño-invierno (anillo del borde hialino), mientras que la pelvis no fusionada de guanaco del M1S2 indica captura entre septiembre y abril (Kaufmann 2009). Por último, las cáscaras de huevo remiten a primavera.
DISCUSIÓN Y CONCLUSIONES
En la muestra de Loma Grande 1 se observa predominio absoluto de fauna dulceacuícola -percas, coipo y aves- y casi ausencia de fauna terrestre, representada por placas dérmicas de armadillo y un hueso de guanaco. La presencia de cinco valvas marinas indica interacción con la costa. Todo el conjunto muestra ausencia de meteorización ósea, lo que indicaría rápido enterramiento de los huesos. Otro aspecto destacable es el alto grado de fragmentación tanto entre los especímenes determinados como entre los no identificados, lo que, dadas la ausencia de meteorización y de marcas de carnívoros y la baja incidencia de raíces, no se debería a factores naturales. La menor fragmentación se observó en la ictiofauna, mientras que es muy alta entre las aves y el coipo. Esta diferencia podría relacionarse con distintas modalidades de procesamiento de los recursos en función de sus características. En este sentido, los peces chicos -como los de esta muestra- no necesitan ser fracturados para su consumo; según Zangrando (2009), la fractura ósea en peces raramente es ocasionada por acción antrópica. Tampoco se observaron huellas de procesamiento, lo que sería esperable en perciformes de menos de 80 cm de largo (Stewart 1991). Los ejemplares podrían haberse asado sobre el cuero y luego manipulado con las manos durante el consumo. No obstante, estudios experimentales con perciformes demostraron que el trozamiento puede producir huellas de corte en vértebras, espinas, cleitro, opercular y preopercular (Willis et al. 2008).
Entre las aves y los coipos hay muchos restos fragmentados pero pocos evidencian procesamiento antrópico: las huellas detectadas en anátidos indican desarticulación y descarne de la región pectoral (Laroulandie 2005); las observadas en coipo, cuereado, desarticulación y extracción de carne (Escosteguy y Vigna 2010). Por su parte, las fracturas transversales y helicoidales con negativos de impacto en los especímenes de aves y coipo sugieren el aprovechamiento de la médula ósea. Si se tiene en cuenta que la tecnología cerámica ya habría estado en uso en el área después de 1500 AP (Gómez Otero et al. 2010), estos fragmentos pudieron hervirse en recipientes cerámicos para así maximizar su rendimiento en grasa ósea y médula. En cambio, la fractura transversal de los radios de ave parece más orientada a la elaboración de cuentas óseas, como las halladas en el canal Beagle (Orquera y Piana 1999). Dado el altísimo porcentaje de huesos quemados, un factor que podría explicar la alta fragmentación es el contacto directo con el fuego. Experimentaciones con huesos sometidos a combustión en fogón mostraron abundante producción de astillas menores a tres centímetros (Lanza 2010: 505) y la disipación de los rasgos identificables de cada elemento luego de la depositación (Stiner et al. 1995: 235). En el caso de animales pequeños, cuya determinación taxonómica es de por sí difícil (Stahl 1996), los efectos de la combustión dificultarían aún más su identificación. Entre los especímenes determinados de LG1 se observó que la acción del fuego afectó por igual a todos los taxa y a todas las partes de su esqueleto. Trabajos experimentales de cocción por combustión de vertebrados pequeños mostraron que esta afecta porciones restringidas de algunos elementos (Hockett y Ferreira Bicho 2000; Laroulandie 2005; Moreno et al. 2007; Medina et al. 2012). Esto no se condice con lo observado en este sitio, sino con los patrones esperados para incendios naturales o actividades de descarte posconsumo (Hockett y Ferreira Bicho 2000). Dado que la arqueofauna termoalterada está directamente asociada con una estructura de combustión de límites acotados, se infiere que estos restos se descartaron en el fogón luego del consumo y que esto incrementó su fragmentación.
Por su parte, la representación integral del esqueleto de los peces y el coipo permite inferir que las presas ingresaron completas al sitio. El predominio del esqueleto axial en los peces se relacionaría con factores culturales, ya que ofrece mayor masa muscular que el cráneo. La presencia de todas las unidades anatómicas de coipo sugiere que no se seleccionaron los elementos con mayor rendimiento económico, como el costillar, la pelvis y el sacro (León y Bonomo 2011). Entre los anátidos y aves indeterminadas, la ausencia del cráneo y el esqueleto axial podría vincularse con causas intrínsecas a su anatomía, como la baja densidad mineral ósea, tal como consignó Elkin (1990, en Savanti 1994) para Chloephaga melanoptera. Sin embargo, tampoco se encuentra el esternón, cuya densidad mineral ósea es baja (Elkin 1990, en Savanti 1994) y su rendimiento cárnico y calórico es alto en Chloephaga picta o cauquén común (Tívoli y Pérez 2009). Quizás el esternón fue descartado fuera del fogón o sufrió tan alta fragmentación que la identificación no pudo ser efectuada de manera segura. En síntesis, diversos rasgos permiten sostener el origen antrópico primario en la depositación de estos restos: 1) su ubicación en un fogón de límites definidos; 2) el alto porcentaje de huesos afectados por combustión; 3) la presencia de huellas que indican distintas etapas del procesamiento; 4) la ausencia de marcas naturales (carnívoro, roedores, raíces); y 5) la asociación contextual con materiales líticos también termoalterados. De manera similar a LG1, el sitio Cinco Esquinas 1 presenta marcada superioridad en la proporción de fauna dulceacuícola y casi nula representación del guanaco y otros recursos terrestres. También aquí se hallaron restos de valvas marinas que, según Gómez Otero y coautores (2010), habrían sido usadas como materia prima. La meteorización diferencial entre los muestreos se relacionaría con distintas historias posdepositacionales. En F2S3 y F1S1 es nula, lo que sugiere que los restos fueron enterrados rápidamente; en cambio, el avanzado grado de deterioro y el blanqueamiento entre las aves y coipos en M1S1 señala mayor tiempo de exposición subaérea. A su vez, las marcas de raíces y depósitos de manganeso en especímenes de todos los muestreos indican un microambiente depositacional de humedad elevada (Gutiérrez 2001). La representación integral del esqueleto de los peces y del coipo permite inferir que las presas entraron completas al sitio. El predominio del espinazo en los peces y la presencia de vértebras articuladas en el fogón F2S3 sugieren extracción de este segmento y posterior descarte en el fuego. En el caso del coipo, al igual que en LG1, no se determinó la selección de partes con mayor rendimiento económico. Entre las aves, la ausencia del esqueleto axial podría explicarse por la baja densidad mineral ósea de este segmento.
En la generalidad de todos los muestreos se destaca la muy baja frecuencia de huellas de acción antrópica (0,04%), lo que puede generar incertidumbre sobre su origen cultural. Los dos contextos más dudosos son CC1A y M1S2: el primero aportó escasos restos faunísticos; el segundo corresponde a un contexto de superficie con unos pocos especímenes quemados. Una explicación alternativa sería que la acumulación de estos restos se debió a la acción hídrica derivada de los grandes episodios de inundación del río anteriores a 1960. En estas crecientes, el río podría haber depositado carcasas de distintos recursos dulceacuícolas, que luego se sedimentarían por diversos procesos diagenéticos e ingresarían al registro arqueológico, mezclándose con materiales culturales. No obstante, otros rasgos permiten defender el origen primario antrópico de los conjuntos zooarqueológicos: a) en CC1 estaban asociados con carbones y tiestos cerámicos; b) en F1S1 y F2S3 se asocian con fogones de límites definidos; c) en F2S3 hay varios huesos termoalterados y la densidad de restos ícticos (1580 NISP/m²) es notablemente más alta que la esperada para depósitos naturales (Zohar et al. 2001). Más allá de esto, el argumento más contundente se apoya sobre la localización del sitio en una cota de 10 msnm, que, según estudios de Pronsato (1950), no habría sido alcanzado por el río en las grandes crecidas. Justamente, la presencia de entierros humanos múltiples sugiere que no eran lugares inundables. Esto estaría reforzado por la ausencia de alteraciones por acción hídrica (e.g., abrasión geológica) en los restos faunísticos.
En síntesis, para ambos sitios se comprobó la explotación casi exclusiva de recursos dulceacuícolas como percas, aves y coipos. Dada su cercanía al río, estas presas se habrían obtenido en el área inmediata o camp surrounding (Binford 1982) y en distintas estaciones del año. La representación integral del esqueleto en percas y coipo; las evidencias de fractura (especialmente en LG1) y corte en aves y coipo y la presencia de especímenes óseos termoalterados sobre todo en huellas de fogón, indican que las presas ingresaron enteras a los sitios, donde fueron procesadas, consumidas, y descartados sus huesos en el fuego. Las modalidades de cocción pudieron haber sido dos: al rescoldo y/o en recipientes cerámicos. Este último método habría permitido optimizar el aprovechamiento de la grasa ósea y la médula, lo que representaría un suplemento nutricional. El contundente predominio de fauna menor dulceacuícola en estos sitios podría vincularse con procesos de intensificación económica (Binford 2001), que buscan aumentar la tasa neta de rendimiento económico por unidad de área ante situaciones de declinación en la disponibilidad de las presas principales. Una de las estrategias es la diversificación en la dieta mediante la incorporación de recursos de baja tasa de retorno energético, como plantas y pequeños animales (Smith 1983). Otra es la especialización en un recurso. La aplicación de ambas estrategias implica el aumento en los costos de obtención, de procesamiento y hasta de almacenamiento, lo que promueve el desarrollo de tecnologías orientadas a tales propósitos, tales como la fabricación de artefactos especializados, recipientes cerámicos, utensilios de molienda, construcción de escondrijos, entre otros.
Para otras cuencas fluviales del norte de Patagonia también se han discutido registros posteriores a 1000 AP de similares características a los tratados en este trabajo. En el caso del curso inferior del río Colorado Martínez (2008-2009) y Stoessel (2012) propusieron la hipótesis de intensificación económica para explicar cambios en la explotación de plantas y recursos faunísticos. Los autores plantean una diversificación alimenticia con la incorporación de fauna pequeña de ambientes terrestres, marinos y fluviales, que complementaron una dieta que hasta entonces había estado basada en guanacos y complementada con venados, choiques y plantas. También establecen un aprovechamiento más intensivo de vegetales, peces marinos y fluviales y guanaco (Stoessel 2012). Por su parte, para la cuenca media del río Negro, Prates (2008) determinó una dieta de amplio espectro para todo el Holoceno tardío final, caracterizada por presas de gran porte -guanaco, venado y ñandú- y recursos complementarios como vegetales, moluscos, reptiles, peces, aves dulceacuícolas, cuises y armadillos. No obstante, Prates y Acosta Hospitaleche (2010: 16) consideran que no se puede aplicar a este caso la hipótesis de diversificación. Una de las razones es la carencia de registro arqueológico para tiempos anteriores al Holoceno tardío; otra, la gran biodiversidad disponible en las cuencas fluviales de los ambientes áridos como los de Patagonia, que desde siempre habría propiciado la aplicación de estrategias oportunistas. Para el VIRCH y la costa norte de Chubut, Gómez Otero (2006) también planteó intensificación económica a partir de 1000 AP, especialmente orientada a maximizar la explotación de plantas. Para ello se apoya sobre distintas líneas de evidencia: a) estudios de isótopos estables de carbono 13 de adherencias orgánicas sobre piezas cerámicas que indicaron presencia de vegetales (Gómez Otero 2007); b) el hallazgo frecuente de artefactos de cerámica y molienda en los sitios de esa antigüedad (Gómez Otero 2006); y c) análisis de salud bucal en restos humanos que muestran un leve aumento de caries y disminución significativa del desgaste dentario y de abscesos en comparación con los individuos del período anterior (Gómez Otero y Novellino 2011).
En cuanto a la aplicación de la hipótesis de diversificación dietaria a los sitios aquí tratados, en principio no sería viable porque no se dispone de datos arqueofaunísticos para tiempos previos a 1500 AP en el VIRCH. No obstante, dado que los análisis isotópicos en restos humanos han mostrado la interacción costa-interior a partir de 6000 AP (Gómez Otero 2006, 2007), una opción válida es discutir información relativa a la desembocadura del río y costa adyacente. Los conjuntos arqueofaunísticos procedentes de estos sectores señalan la explotación preponderante de guanacos y moluscos (en muy pocos casos predominan los pinnípedos) desde el Holoceno medio hasta tiempos inmediatamente anteriores al contacto con los europeos. Los recursos menores como peces, aves y pequeños mamíferos, también están presentes pero no se observan tendencias claras en cuanto a un incremento en la intensidad de su utilización a través del tiempo, en especial a partir del Holoceno tardío final (Gómez Otero 2006; Gómez Otero et al. 2009, 2013). Esto se observó en dos localidades arqueológicas de la desembocadura: Los Cangrejales Sur y Barranca Norte. En la primera se hallaron restos de peces, aves indeterminadas y armadillos en varias capas culturales datadas entre 2290 AP y 840 AP (Gómez Otero et al. 2009); en Barranca Norte, un nivel (B2N2) de ca. 3000 AP señaló la mayor abundancia de peces respecto de otros niveles más tardíos (Gómez Otero 2006).
Por lo expuesto, es factible seguir sosteniendo la hipótesis planteada inicialmente por Gómez Otero y coautores (2010: 19-21), que propone explotación oportunista de recursos dulceacuícolas durante las relativamente periódicas grandes crecientes del río. Estas inundaciones generarían un ambiente ribereño de aguas poco profundas muy favorable para la fauna fluvial pero inapropiado para los animales terrestres, entre ellos el guanaco y el choique. Esto habría disminuido la disponibilidad de estas grandes presas en el valle y propiciado la aplicación de estrategias para minimizar el riesgo mediante la incorporación de recursos de menor jerarquía a la dieta. Más allá de esto, como agenda para el futuro, sería importante explorar si la perduración de estos escenarios ribereños durante meses o años pudo haber propiciado estrategias de subsistencia más sistemáticas, como la obtención de recursos de agua dulce en el valle y de guanacos y otras presas terrestres en las zonas secas y altas de las mesetas circundantes.
Agradecimientos
A Isabel Cruz y Paula Escosteguy por el aporte bibliográfico, a Ulyses Pardiñas por el préstamo de material comparativo, a Atilio Francisco Zangrando, quien asesoró en la identificación taxonómica de los peces en Cinco Esquinas 1, y al Ing. Gabriel Kaless por la bibliografía édita e inédita y el asesoramiento. También queremos agradecer a los evaluadores anónimos, cuyos comentarios y sugerencias ayudaron a mejorar sustancialmente la presentación. Este trabajo contó con fondos de subsidios de PIP-CONICET 6470 y 11401000100210 otorgados a Julieta Gómez Otero, y de una beca doctoral Tipo I del CONICET adjudicada a Ariadna Svoboda.
REFERENCIAS CITADAS
1. Andrews, P. 1990 Owls, caves and fossils. Natural History Museum Publications, Londres. [ Links ]
2. Arrigoni, G. 1991 Interpretación de la ocupación del Alero del Chamán. Shincal 3: 216-220. [ Links ]
3. Beeskow, A. M., H. F. Del Valle y C. M. Rostagno 1987 Los sistemas fisiográficos de la región árida y semiárida de la provincia del Chubut. Delegación Regional Patagonia, Secretaría de Ciencia y Tecnología (SECyT), Puerto Madryn. [ Links ]
4. Behrensmeyer, A. K. 1978 Taphonomic and Ecological Information from Bone Weathering. Paleobiology 4 (2): 150-162. [ Links ]
5. Binford, L. R. 1981 Bones: Ancient Men and Modern Myths. Academic Press, Nueva York. [ Links ]
6. Binford, L. R. 1982 The Archaeology of Place. Journal of Anthropological Archaeology 1: 5-31. [ Links ]
7. Binford, L. R. 1984 Faunal Remains from Klasies River Mouth. Academic Press, Orlando. [ Links ]
8. Binford, L. R. 2001 Constructing Frames of Reference: An analytical Method for Archaeological Theory Building using Hunter-gatherer and Environmental Data Sets. University of California Press, Berkeley. [ Links ]
9. Cabrera, A. L. y J. Yepes 1960 Mamíferos sudamericanos. Ediar, Buenos Aires. [ Links ]
10. Cruz, I., S. Muñoz y F. Zangrando 2007 La interpretación de los restos de animales pequeños en la arqueología patagónica: estado de la cuestión y perspectivas. En Arqueología de Fuego- Patagonia. Levantando piedras, desenterrando huesos. y develando arcanos, editado por F. Morello, M. Martinic, A. Prieto y G. Bahamonde, pp. 15-22. Centro de Estudios del Cuaternario, Fuego, Patagonia y Antártica (CEQUA), Chile. [ Links ]
11. Escosteguy, P. D. y M. Vigna 2010 Experimentación en el procesamiento de Myocastor coypus. En Mamül Mapu: pasado y presente desde la arqueología pampeana, editado por M. Berón, L. Luna, M. Bonomo, C. Montalvo, C. Aranda y M. Carrera Aizpitarte, pp. 293-307. Del Espinillo, Ayacucho. [ Links ]
12. Fernández, P. M. 2008 Taphonomy and zooarchaeology in the Neotropics: A view from northwestern Patagonian forest and steppe. Quaternary International 180: 63-74. [ Links ]
13. Giardina, M. A. 2012 Intensificación en el sur de Mendoza: un enfoque avifaunístico. Archaeofauna 21: 219-234. [ Links ]
14. Gómez Otero, J. 1994 Parada 4. Sitio Loma Grande. En Guía de Campo de la VII Reunión de Campo del CADINQUA, pp. 66-67. Centro Nacional Patagónico (CENPAT), Puerto Madryn. [ Links ]
15. Gómez Otero, J. 2006 Dieta, uso del espacio y evolución en poblaciones cazadoras-recolectoras de la costa centro-septentrional de Patagonia durante el Holoceno medio y tardío. Tesis Doctoral inédita. Facultad de Filosofía y Letras, Universidad de Buenos Aires, Buenos Aires. [ Links ]
16. Gómez Otero, J. 2007 Isótopos estables, dieta y uso del espacio en la Costa Atlántica Centro-septentrional y el Valle inferior del río Chubut (Patagonia Argentina). En Arqueología de Fuego-Patagonia. Levantando piedras, desenterrando huesos. y develando arcanos, editado por F. Morello, M. Martinic, A. Prieto y G. Bahamonde, pp. 151-162. CEQUA, Chile. [ Links ]
17. Gómez Otero, J. y P. Novellino 2011 Diet, Nutritional Status and Oral Health in Hunter- Gatherers from the Central-Northern Coast of Patagonia and the Chubut River Valley, Argentina. Journal of Osteoarchaeology 21: 643-659. [ Links ]
18. Gómez Otero, J., N. Weiler y J. E. Moreno 2009 Localidad Arqueológica Los Cangrejales: evidencias de ocupaciones humanas y de variaciones de la línea de costa en el Holoceno tardío. En Arqueología de Patagonia: una mirada desde el último confín, t. II, editado por M. Salemme, F. Santiago, M. Álvarez, E. Piana, M. Vázquez y E. Mansur, pp. 1023-1036. Utopías, Ushuaia. [ Links ]
19. Gómez Otero, J., E. Moreno y V. Schuster 2010 Ocupaciones tardías en el valle inferior del río Chubut: primeros resultados del sitio Cinco Esquinas 1. Actas del XVII Congreso Nacional de Arqueología Argentina, t. V: 1917-1922. Mendoza. [ Links ]
20. Gómez Otero, J., N. Weiler, A. Banegas y J. E. Moreno 2013 Ocupaciones del Holoceno medio en Bahía Cracker, costa atlántica de Patagonia Central. En Tendencias teórico-metodológicas y casos de estudio en la arqueología de la Patagonia, editado por A. F. Zangrando, R. Barberena, A. Gil, G. Neme, M. Giardina, L. Luna, C. Otaola, S. Paulides, L. Salgán y A. Tívoli, pp. 177-186. Museo de Historia Natural de San Rafael, San Rafael. [ Links ]
21. Gosztonyi, A. E. 1988 Peces del río Chubut inferior, Argentina. Physis. Secc. B 46 (110): 41-50. [ Links ]
22. Gutiérrez, M. A. 2001 Bone Diagenesis and Taphonomic History of the Paso Otero 1 Bone Bed, Pampas of Argentina. Journal of Archaeological Science 28: 1277-1290. [ Links ]
23. Hajduk, A. y A. Albornoz 1999 El sitio Valle Encantado 1. Su vinculación con otros sitios: un esbozo de la problemática local del Nahuel Huapi. En Soplando en el Viento. Actas de las III Jornadas de Arqueología de la Patagonia, pp. 371- 391. Instituto Nacional de Antropología y Pensamiento Latinoamericano (INAPL)-Universidad Nacional del Comahue, Neuquén-Buenos Aires. [ Links ]
24. Hajduk, A., A. Albornoz y M. Lezcano 2004 El "Mylodon" en el patio de atrás. Informe preliminar sobre los trabajos en el sitio El Trébol, ejido urbano de San Carlos de Bariloche, Provincia de Río Negro. En Contra viento y marea. Arqueología de Patagonia, editado por M. T. Civalero, P. M. Fernández y A. G. Guráieb, pp. 715-731. INAPL y Sociedad Argentina de Antropología (SAA), Buenos Aires. [ Links ]
25. Hockett, B. y N. Ferreira Bicho 2000 The rabbits of Picareiro Cave: small mammal hunting during the late upper paleolithic in the Portuguese Estremadura. Journal of Archaeological Science 27: 715-723. [ Links ]
26. Hughes, W. M. 1993 A orillas del río Chubut en la Patagonia. El Regional, Rawson. [ Links ]
27. Kaufmann, C. 2009 Estructura de edad y sexo en guanacos: estudios actualísticos y arqueológicos en Pampa y Patagonia. SAA, Buenos Aires. [ Links ]
28. Lanza, M. 2010 Arqueología experimental y conjuntos arqueofaunísticos en sitios históricos. En Zooarqueología a principios del siglo XXI: aportes teóricos, metodológicos y casos de estudio, editado por M. De Nigris, P. M. Fernández, M. Giardina, A. F. Gil, M. A. Gutiérrez, A. Izeta, G. Neme y H. D. Yacobaccio, pp. 499-509. Del Espinillo, Buenos Aires. [ Links ]
29. Laroulandie, V. 2005 Bird exploitation pattern: the case of Ptarmigan Lagopus sp. in the Upper Magdalenian site of La Vache (Ariège, France). En Feathers grit and symbolism, Birds and humans in the ancient Old and New World. Proceedings of the 5th meeting of the ICAZ Bird Working Group, editado por G. Grupe, y J. Peters, pp. 165-178. Verlag Marie Leidorf, Rahden. [ Links ]
30. León, C. y M. Bonomo 2011 Índices de rendimiento económico de coipo (Myocastor coypo). Segundo Congreso Nacional de Zooarqueología Argentina. UNICEN, Olavarría. [ Links ]
31. León, R., D. Bran, M. Collantes, J. M Paruelo y A. Soriano 1998 Grandes unidades de vegetación de la Patagonia extra andina. Ecología Austral 8 (2): 75-308. [ Links ]
32. Lyman, L. R. 1994 Vertebrate Taphonomy. Cambridge University Press, Nueva York. [ Links ]
33. Lyman, L. R. 2008 Quantitative Paleozoology. Cambridge University Press, Nueva York. [ Links ]
34. Martínez, G. A. 2008-2009 Arqueología del curso inferior del río Colorado: estado actual del conocimiento e implicaciones para la dinámica poblacional de cazadores-recolectores pampeano-patagónicos. Cazadores-recolectores del Cono Sur 3: 71-92. [ Links ]
35. Martínez, G., A. F. Zangrando y L. Stoessel 2005 Sitio El Tigre (Pdo. de Patagones, Pcia. de Buenos Aires, Argentina): evidencias sobre la explotación de peces en el curso inferior del Río Colorado e implicaciones para los sistemas de subsistencia. Magallania 33: 127-142. [ Links ]
36. Matthews, A. 1992. Crónica de la colonia galesa de la Patagonia. El Regional, Rawson. [ Links ]
37. Medina, M. E., P. Teta y D. Rivero 2012 Burning damage and small-mammal human consumption in Quebrada del Real 1 (Córdoba, Argentina): an experimental approach. Journal of Archaeological Science 39: 737-743. [ Links ]
38. Menegaz, A. 1996 Análisis del material faunístico procedente del sitio Piedra del Águila 11 (Neuquén, Argentina). Praehistoria 2: 147-172. [ Links ]
39. Mengoni Goñalons, G. 1999 Cazadores de guanacos en la estepa patagónica. SAA, Buenos Aires. [ Links ]
40. Moreno, J. E., B. Videla, H. Pérez Ruiz, L. Asencio y V. Leonforti 2007 Búsqueda de indicadores de diversificación económica prehistórica en la Cuenca del Lago Musters (Chubut, Argentina), primeros resultados. En Arqueología de Fuego-Patagonia. Levantando piedras, desenterrando huesos. y develando arcanos, editado por F. Morello, M. Martinic, A. Prieto y G. Bahamonde, pp. 23-32. CEQUA, Chile. [ Links ]
41. Moreno, J. E. y H. Pérez Ruiz 2010 Evidencias de utilización prehispánicas de recursos fluviales en la cuenca del lago Musters (Chubut, Argentina). Actas del XVII Congreso Nacional de Arqueología Argentina, t. I: 345-350. Mendoza. [ Links ]
42. Muñoz, A. y F. Savanti 1998 Observaciones tafonómicas sobre restos avifaunísticos de la costa noreste de Tierra del Fuego. Revista del Museo de Historia Natural de San Rafael 20 (1-2): 107-121. [ Links ]
43. Orquera, L. A. y E. L. Piana 1999 Arqueología de la región del canal Beagle (Tierra del Fuego, República Argentina). SAA, Buenos Aires. [ Links ]
44. Prates, L. 2008 Los indígenas del río Negro, un enfoque arqueológico. SAA, Buenos Aires. [ Links ]
45. Prates, L. y C. Acosta Hospitaleche 2010 Las aves de sitios arqueológicos del Holoceno tardío de Norpatagonia, Argentina. Los sitios Negro Muerto y Angostura 1 (Río Negro). Archaeofauna 19: 7-18. [ Links ]
46. Pronsato, A. D. 1950 Estudio geohidrológico del Río Chubut. Parte II. Dirección General de Agua y Energía Eléctrica. Revista Agua y Energía 29 y 30. [ Links ]
47. Ruzzante, D., S. Walde, P. Macchi, M. Alonso y J. Barriga 2011 Phylogeography and phenotypic diversification in the Patagonian fish Percichthys trucha: the roles of Quaternary glacial cycles and natural selection. Biological Journal of the Linnean Society 103: 514-529. [ Links ]
48. Savanti, F. 1994 Las aves en la dieta de los cazadores-recolectores terrestres de la costa fueguina. Temas de Arqueología. Programa de Estudios Prehistóricos, Buenos Aires. [ Links ]
49. Silveira, J. M. 1996 Alero Los Cipreses (Provincia del Neuquén, Republica Argentina). En Arqueología Solo Patagonia, editado por J. Gómez Otero, pp. 107-118. CENPAT, Puerto Madryn. [ Links ]
50. Smith, E. A. 1983 Anthropological Applications of Optimal Foraging Theory. Current Anthropology 24: 625-651. [ Links ]
51. Stahl, P. W. 1996 The Recovery and Interpretation of Microvertebrate Bone Assemblages from a Archaeological Contexts. Journal of Archaeological Methods and Theory 3 (1): 31-75. [ Links ]
52. Stewart, K. M. 1991 Modern Fishbone Assemblages at Lake Turkana, Kenya: A Methodology to Aid in Recognition of Hominid Fish Utilization. Journal of Archaeological Science 18: 579-603. [ Links ]
53. Stiner, M. C., S. L. Khun, S. Weiner y O. Bar-Yosef 1995 Differential Burning, Recrystallization, and Fragmentation of Archaeological Bone. Journal of Archaeological Science 22: 223-237. [ Links ]
54. Stoessel, L. 2012 Consumo de peces en el área ecotonal áridasemiárida del curso inferior del río Colorado (provincia de Buenos Aires) durante el Holoceno tardío. Relaciones de la Sociedad Argentina de Antropología XXXVII (1): 159-182. [ Links ]
55. Svoboda, A. 2013a Método para determinar la estacionalidad de ocupación de sitios arqueológicos de ambientes fluviolacustres de Patagonia a partir de la observación de otolitos de percas. La Zaranda de Ideas, Revista de Jóvenes Investigadores en Arqueología 9 (2). En prensa. [ Links ]
56. Svoboda, A. 2013b Disponibilidad cárnica, rendimiento energético y estimación de la talla de Percichthys trucha (perca criolla) a partir de la morfometría de huesos diagnósticos y su aplicación a los conjuntos ictioarqueológicos de Patagonia Central. Cuadernos de Antropología 9: 251-266. [ Links ]
57. Svoboda, A. y J. E. Moreno 2014 Experimentación sobre los efectos de la meteorización en la supervivencia de elementos óseos de Percichthys trucha: implicaciones ictioarqueológicas para el sitio DV1, Lago Musters (Prov. de Chubut, Argentina). Revista Chilena de Antropología 29: 60-67. [ Links ]
58. Tívoli, A. M. y A. F. Pérez 2009 Rendimiento económico del cauquén común (Chloephaga picta, Familia: Anatidae). En Arqueología de Patagonia: una mirada desde el último confín, t. II, editado por M. Salemme, F. Santiago, M. Álvarez, E. Piana, M. Vázquez y E. Mansur, pp. 853-864. Utopías, Ushuaia. [ Links ]
59. Wheeler, A. y A. K. Jones 1989 Fishes. Cambridge University Press, Cambridge. [ Links ]
60. Willis, L. M., M. I. Eren y T. C. Rick 2008 Does butchering fish leave cut marks? Journal of Archaeological Science 35: 1438-1444. [ Links ]
61. Zangrando, A. F. 2009 Historia Evolutiva, tiempos y subsistencia humana en la región del Canal Beagle. Una aproximación zooarqueológica. SAA, Buenos Aires. [ Links ]
62. Zohar,I., T. Dayan, E. Galili y E. Spanier 2001 Fish Processing During the Early Holocene: a Taphonomic Case Study from Coastal Israel. Journal of Archaeological Science 28: 1041-1053 [ Links ]
