










Servicios Personalizados
Revista

Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkIntersecciones en antropología
versión On-line ISSN 1850-373X
Intersecciones antropol. vol.16 no.1 Olavarría jun. 2015
ARTÍCULOS
Camélidos domésticos en el Valle de Mauro (Norte Semiárido, Chile): múltiples análisis para un mismo problema
Patricio López M., Isabel Cartajena F., Boris Santander, Daniel Pavlovic B. y Daniel Pascual
Patricio López M. Universidad de Chile, Facultad de Ciencias Sociales, Departamento de Antropología, Calle Ignacio Carrera Pinto N° 1045, Ñuñoa, Santiago, Chile. E-mail: patriciolopezmend@gmail.com
Isabel Cartajena F. Universidad de Chile, Facultad de Ciencias Sociales, Departamento de Antropología, Calle Ignacio Carrera Pinto N° 1045, Ñuñoa, Santiago, Chile. E-mail: isabel.cartajena@gmail.com
Boris Santander. Departamento de Antropología. Universidad Alberto Hurtado. Almirante Barroso 10, Santiago, Chile; Departamento de Historia, Universitat Rovira i Virgili e IPHES,España. E-mail: bdsantander@uahurtado.cl
Daniel Pavlovic B. Departamento de Antropología, Facultad de Ciencias Sociales, Universidad de Chile e Instituto de Estudios de Montaña de Aconcagua. Eduardo Castillo Velasco 2788, Ñuñoa, Santiago, Chile. E-mail: daniel.pavlovic@gmail.com
Daniel Pascual. Arzobispo Fuenzalida 2918, Ñuñoa, Santiago, Chile. E-mail: danipascual79@gmail.com
Recibido 2 de junio 2013.
Aceptado 18 de octubre 2013
RESUMEN
Valle de Mauro (31° S) tiene una larga historia de ocupaciones humanas desde el Arcaico medio (~7.500 años AP) hasta la actualidad. Para momentos prehispánicos, este valle presenta una extensa secuencia ocupacional que refleja gran parte de las múltiples manifestaciones culturales identificadas para el interior de la zona meridional del Norte Semiárido de Chile. A lo largo de toda la secuencia temporal estudiada, los camélidos son el recurso cárnico principal. Al respecto, fueron identificadas evidencias de cambios en su explotación desde el Arcaico tardío hasta el periodo Tardío (influencia del Tawantinsuyu). Uno de estos principales cambios es la aparición de camélidos domésticos en el registro, por lo que se discute si estos habrían sido introducidos o domesticados localmente. Por ende, una pregunta que surge es cuándo comienzan estos cambios y qué factores fueron los desencadenantes. A partir del cruce de los resultados de los análisis contextuales, zooarqueológicos, osteométricos, de isótopos y paleoambientales, se observa una correlación directa entre los cambios de tamaño y dieta desde el Arcaico tardío hasta el periodo Tardío en el valle, y refuerza la idea de una introducción de camélidos domésticos y la crianza local a partir del periodo Intermedio tardío que se afianza durante el Tardío.
Palabras clave: Norte semiárido; Chile; Camélidos; Osteometría; Isótopos Estables.
ABSTRACT
Domestic camelids in Mauro valley (Semiarid north, Chile): multiple analyses of the same problem
The Valley of Mauro (31° S) has a long occupational history from the Middle Archaic (c. 7500 years BP) to the present. Most of the pre-Hispanic cultural manifestations of the inland southern zone of the Semiarid North of Chile were identified in this valley. Camelids were exploited during the entire occupational sequence, principally as the main meat resource. However, changes in their exploitation can be observed from the Late Archaic through the Late Period, especially under the influence of Tawantinsuyu. One of the main transformations is the appearance of domestic camelids in the record. The paper discusses whether they were introduced or locally domesticated as well as which factors could have triggered their incorporation. Results of the zooarchaeological and osteometric analyses were compared with stable isotopes results and paleoenvironmental data. A direct correlation between changes in camelid size and diet was observed from the Late Archaic to the Late Period in the valley, reinforcing the idea of the introduction of domestic camelids and local breeding beginning in the Intermediate Late Period and consolidating during the Late Period.
Keywords: Semiarid North; Chile; Camelids; Osteometry; Stable Isotopes.
INTRODUCCIÓN
La prehistoria del Norte Semiárido de Chile se ha focalizado principalmente en tradiciones culturales asociadas a bloques temporales fragmentarios. En la medida que se han ido identificando nuevos yacimientos y reevaluando antiguos contextos, se observan grandes vacíos de información que den cuenta de las transformaciones sufridas por estos grupos humanos, particularmente, datos referentes a la presencia de grupos sustentados en una economía agroganadera, a partir de la incorporación de camélidos domésticos. Si bien hallazgos como los entierros de Plaza Coquimbo sugirieron la importancia de los camélidos domésticos para las comunidades del Complejo Cultural Ánimas (Castillo 1984), nuevos trabajos señalarían que la incorporación de la llama (Lama glama) es relativamente tardía y que iría de la mano con la influencia del Tawantinsuyu (Becker 2004; Troncoso et al. 2009). Gran parte de las dificultades para abordar este problema se deben a la escasa evidencia disponible y a la falta de trabajos referidos al análisis de restos arqueofaunísticos en esta región. A lo anterior se une lo complejo de trabajar problemas como la crianza y domesticación de animales, ya que su estudio involucra aspectos relacionados con cambios morfológicos y etológicos, además de nuevas respuestas culturales (como corrales) o espaciales (delimitar áreas de pastoreo), entre otras. Estos estudios han sido profusamente abordados para el caso de los camélidos sudamericanos. En ellos, a pesar de las dificultades planteadas para la diferenciación de las cuatro especies tanto domésticas (llama/alpaca) como silvestres (guanaco/vicuña) a partir de restos zooarqueológicos (Hesse 1982; Kent 1982; Wheeler 1982; Elkin 1996; Mengoni Goñalons 2008), la utilización de métodos cuantitativos basados en la osteometría -especialmente la aplicación de diversos análisis estadísticos- ha permitido delimitar diversos grupos de tamaño atribuibles a diferencias taxonómicas (Cartajena 2009, 2013; Izeta et al. 2009a; Izeta 2009, 2010; Grant 2010; L`Heureux 2010; Yacobaccio 2010; Dantas 2012, entre otros). No obstante, tal como lo señala Izeta (2009), la mayor dificultad radica en la interpretación de los datos, ya que en general se cuenta con muy pocos ejemplares comparativos.
En el presente trabajo se aborda la presencia de camélidos domésticos en el Valle de Mauro (Norte Semiárido, Chile), a través de un ejercicio que tuvo por fin cruzar distintos tipos de fuentes de información, considerando el problema de la domesticación o introducción de animales domésticos a partir de una larga secuencia temporal que va desde el Arcaico tardío hasta las influencias Tawantinsuyu durante el periodo Tardío. A lo largo de su historia cultural, este valle ha sido escenario de las principales manifestaciones arqueológicas del Norte Semiárido de Chile, y por ende, refleja en parte los procesos culturales y ecológicos acaecidos en esta franja intermedia entre el Norte Árido y Chile Central. Dentro de los recursos explotados, los camélidos fueron una de las fuentes económicas más relevantes en la región surandina, con un valor agregado como animal de carga -en el caso de la llama- y como agente articulador de distintas manifestaciones sociales. Para abordar este problemática, se llevó a cabo un ejercicio que incorporó cuatro variables. En primer lugar, se analizaron las muestras zooarqueológicas de todos los sitios asignados al Arcaico tardío (AT), periodo Alfarero temprano (PAT), periodo Intermedio tardío (PIT) y periodo Tardío (PT) en función de su frecuencia, diversidad y perfiles etarios, con el fin de identificar diferencias entre los contextos más relevantes. Un segundo análisis realizado fue el osteométrico, con el objetivo de identificar las diferencias de tamaño desde el Arcaico tardío hasta el periodo Tardío, seguido por el estudio de isótopos estables (carbono y nitrógeno) para dar cuenta de los cambios en la dieta a lo largo de la secuencia. Por último, estos datos fueron relacionados con los datos ambientales y sus posibles implicancias en los cambios de tamaño y dieta de los camélidos estudiados.
EL VALLE DE MAURO, ANTECEDENTES Y PROBLEMAS DE INVESTIGACIÓN
El Norte Semiárido (NSA en adelante) es un área intermedia entre el gran Norte Árido y la Zona Central de Chile. A lo largo de su extensión (26°-33° S), refleja tanto en su clima como en su vegetación rasgos transicionales entre el desierto absoluto del norte y de ambientes mediterráneos que se manifiestan más hacia el sur (Veit 1993). Las ocupaciones AT del NSA se hacen extensivas en parte en el Valle de Mauro a partir del registro arqueológico de dos sitios, a saber, MAU085 y MAU033. El primero corresponde a una extensa área de actividades de procesamiento de camélidos, retoque de artefactos líticos, recambios de puntas de proyectil fracturadas, trabajo en fibras, entre otras (Santander y López 2012). Por su parte, MAU033 es un pequeño montículo con múltiples contextos funerarios que van desde el Arcaico medio hasta el PAT. El AT ha sido estudiado con mayor detalle en la zona costera, mientras que para el interior la información es escasa (Cornejo y Jackson 2004), por lo que los sitios identificados en Mauro son relevantes para la comprensión de las poblaciones arcaicas y su transición a sociedades alfareras. En la costa, las ocupaciones AT se caracterizan por una movilidad residencial en función de los recursos, con una economía eminentemente cazadora y recolectora (Méndez y Jackson 2004).
Para el PAT se han reconocido ocupaciones tanto en la costa como en el interior (Pavlovic 2004). Al interior del valle del Choapa, los campamentos base se emplazan en quebradas tributarias y en las zonas altas de los valles, mientras que los sitios de tareas efímeras se ubican en terrazas bajas y quebradas secundarias (Pavlovic 2004: 40). Por su parte, los estudios de Méndez y Jackson (2008) en Combarbalá, a partir de sitios PAT, identificaron patrones de alta movilidad con estrategias tecnológicas conservadoras, especialmente las líticas, muy similares a las del AT, y una dependencia en la caza y recolección. Evidencias cerámicas Agrelo-Calingasta sugieren además una movilidad entre ambas vertientes andinas, mientras que las evidencias cerámicas locales indican que las vasijas estarían relacionadas con el traslado a baja escala de recursos vegetales (Méndez et al. 2009). Para el Valle de Mauro, el registro arqueológico sugiere durante el PAT estrategias expeditivas y una alta movilidad, aunque las evidencias se encuentran en proceso de estudio. Por su parte, durante el PIT se destaca el desarrollo de la denominada Cultura Diaguita (~1000-1400 años DC), caracterizada por grupos con una alta movilidad orientada a la obtención de diversos recursos, con un mayor desarrollo agrícola y un patrón de asentamiento disperso centrado en las terrazas fluviales (Troncoso 2001). Este patrón de asentamiento se complementa además con un consumo de quinoa, maíz y la caza básicamente de guanaco (Lama guanicoe), junto con el aprovechamiento de recursos malacológicos en las tierras interiores (Troncoso 2001). La posible presencia de llama, además del aumento del consumo de peces y una explotación mayor y diversa de recursos costeros, forman parte de las transformaciones económicas de esta cultura a lo largo de su desarrollo (Becker 2004). Estas transformaciones se acentúan con la llegada del Inca durante el PT, y es clara para este periodo la aparición de la llama como recurso económico y estratégico para unir sectores distantes. Además, estos cambios trasuntan en el paso de pequeños núcleos campesinos a una organización ligada a la lógica incaica, sobre todo en la producción de excedentes y tributos e integrando a la sociedades PIT en una compleja red de intercambio y circulación de bienes (Troncoso 2004).
El Valle de Mauro se emplaza en la zona meridional del NSA (31º 57`S-71º 01`O, ~900 msnm) y corresponde a la cuenca preandina formativa del estero Pupío, que desemboca en el océano Pacífico, a unos 40 km al poniente, y que se inserta dentro de un área dominada por un clima de estepa templado (Figura 1). Este valle fue estudiado profusamente, y se registraron en él un total de 107 sitios arqueológicos cuyas excavaciones, en un número no menor, fueron intensivas. A partir de estos trabajos de terreno y los posteriores análisis en laboratorio, fue posible identificar ocupaciones desde inicios del Arcaico medio (7490 ± 30 años AP sin calibrar) hasta tiempos históricos, aunque las evidencias arqueofaunísticas se registran a partir del AT (3040 ± 25 a 2670 ± 25 años AP sin calibrar) en sitios como MAU085 y MAU033 (Santander y López 2012). Una de las primeras preguntas que surgieron en el estudio de este valle fue sobre el modo de explotación de la fauna propia del sector, y particularmente sobre la visibilidad de las evidencias arqueológicas referentes a la introducción, crianza y/o domesticación de camélidos. Si se consideran todas las evidencias disponibles para el NSA, el problema de la domesticación es complejo, y hasta ahora, la hipótesis más aceptada indica que la llama fue introducida en momentos tardíos, durante la influencia del Tawantinsuyu (Troncoso et al. 2009). Sin embargo, cabe preguntarse si para el PIT e incluso para el PAT existen indicios de que la presencia de animales domésticos pudiera haber sucedido más tempranamente y cómo es posible identificar tal fenómeno (Meadow 1989; Miller y Burger 1995).

Figura 1. A. Ubicación del Valle de Mauro en la IV Región de Coquimbo con detalles de ambientes aledaños. B. Ubicación de los sitios del valle mencionados en el texto.
Al respecto, la osteometría es un método frecuentemente aplicado para este tipo de problemas de investigación, aunque presenta una serie de dificultades para su correcta interpretación. La primera y la más ampliamente discutida es la dificultad para la separación entre las especies domésticas de camélidos sudamericanos (llama y alpaca) y silvestres (guanaco y vicuña) puesto que si bien es posible separar entre el conjunto de camélidos grandes (guanaco/llama) y pequeños (vicuña/ alpaca), al interior de cada grupo existen amplias áreas de traslape (Cartajena 2002; Cartajena et al. 2007). Una segunda dificultad se relaciona con las diferencias de tamaño entre ambientes como la costa y el interior, y por diferencias latitudinales; mientras que finalmente, los estándares utilizados como referentes para separar entre animales silvestres versus domesticados (Cartajena et al. 2007, 2012). Por esto, y para disminuir el rango de error, es necesario contar con una secuencia temporal lo más amplia posible y que refleje los morfotipos más tempranos, con el fin de evaluar la historia de cambios morfométricos a lo largo del tiempo a partir de un conjunto local. Por lo anterior, si bien los cambios observados pueden haber sido inducidos por control antrópico, es necesario diferenciarlos del potencial impacto de cambios en el ambiente (Cartajena et al. 2007; Cartajena 2013). Es necesario recalcar que el estudio de estos cambios requiere de un cruce necesario con otro tipo de información, como la paleoambiental y contextual, a las cuales, en los últimos años, se ha agregado la dieta de los camélidos como un proxy importante, ya que refleja en parte el manejo de rebaños y áreas de captura de estos animales (Finucane et al. 2006; Mengoni Goñalons 2007; Izeta et al. 2009b; Yacobaccio et al. 2009; Dantas 2012, entre otros). Una de las hipótesis iniciales, que presentamos en este trabajo, señala que los cambios morfométricos en los restos de camélidos desde el AT hasta el PT observados en trabajos previos (Cartajena et al. 2012) tienen correlato directo con posibles cambios en la dieta, explicables principalmente por la introducción y la mantención de animales domésticos. Hasta la fecha, los análisis isotópicos realizados en el presente trabajo son los primeros referentes para todo el NSA.
SITIOS ESTUDIADOS Y METODOLOGÍA DE ANÁLISIS
Los sitios estudiados y sus asignaciones cronológicas y culturales se detallan en las Tablas 1 y 2. En total, el registro analizado corresponde a 286.473 especímenes óseos (NISP). Este conjunto se distribuye de modo heterogéneo para cada contexto, por cuanto sitios como MAU085, MAU033, MAU062, MAU083, MAU091, MAU092, MAU094 y MAU067 acumulan la mayor parte de especímenes y muestras usadas para el análisis osteométrico e isotópico; mientras que los restantes sitios representan un porcentaje muy menor de restos óseos. De acuerdo con los objetivos explicitados anteriormente, el análisis zooarqueológico consideró como unidad de medición el %MAU y variables como el Índice de Utilidad (Borrero 1990 y modificado por Lyman 1994), además de Índices de Cavidad Medular y Secado (Mengoni Goñalons 1996; De Nigris y Mengoni Goñalons 2004), con el fin de evaluar tendencias en las muestras de camélidos de los sitios más relevantes para cada periodo analizado. Los perfiles de edad se realizaron mediante los rangos propuestos por Kauffman (2004). Con la comparación de todos estos periodos se busca identificar algunas tendencias entre el patrón de asentamiento y las estrategias de procesamiento y consumo, sobre todo para sitios PIT-PT debido a la influencia del Tawantinsuyu y sus repercusiones en la estratificación social.
Tabla 1. Taxones identificados en los sitios MAU085 y MAU033 asignados al Arcaico tardío (AT) y periodo Alfarero temprano (PAT) del Valle de Mauro (sitios MAU076, MAU087, MAU077, MAU066 y MAU062).

Referencia: *Con un componente datado para el PIT-PT.
Tabla 2. Taxones identificados en los sitios MAU065, MAU083, MAU092 y MAU091 asignados al periodo Intermedio tardío (PIT) y con componentes del Periodo Tardío (PT) del Valle de Mauro (MAU064, MAU068, MAU094 y MAU067).

En cuanto al análisis isotópico, este va de la mano con el estudio osteométrico ya que las muestras seleccionadas -en su mayoría- corresponden a huesos medidos. Dentro de estas muestras se incluyen principalmente primeras y segundas falanges, además de astrágalos, y en el caso de no contar con estos especímenes, huesos cortos y otros que no fueron medidos debido a lo escaso de la muestra disponible. La totalidad de los análisis isotópicos fueron realizados sobre colágeno de hueso en el Center for Applied Isotope Studies (CAIS) de la University of Georgia. Para mantener un control sobre los valores isotópicos se midió la concentración de C (C%) y N (N%) y se calculó la razón C/N para corroborar la correcta preservación del colágeno en los huesos. Los valores de ä13Ccol (13C/12C) se expresan en partes por mil () siguiendo el estándar internacional PeeDee Belemnite (PDB) con un error menor al 0.1, y las del ä15Ncol (15N/14N) en partes por mil () según el estándar internacional para el Nitrógeno Atmosférico (Air) con un error menor de 0.2. Para estos análisis se utilizó un Espectrómetro de Masas modelo Finnigan MAT252, y previamente se realizó un tratamiento de las muestras para eliminar contaminantes que se hayan incorporado al tejido óseo. A las muestras de faunas actuales, que fueron utilizadas como referentes, se les aplicó un factor de corrección de + 1.5 debido al denominado "efecto industrial" (Marino y McElroy 1991). Para el análisis osteométrico se utilizó un total de 279 especímenes óseos, principalmente primeras y segundas falanges tanto anteriores como posteriores, además de astrágalos completos, o bien secciones que permitiesen la obtención de más de un par de medidas. Todas estas medidas fueron obtenidas a partir de muestras de animales adultos y utilizando los estándares de Von den Driesch (1999); se privilegiaron muestras datadas directamente mediante 14C y analizadas isotópicamente. Para visualizar distribuciones de pares de medidas se utilizaron gráficos de dispersión a partir del par de variables que discriminó de mejor manera los grupos. Las medidas del material zooarqueológico fueron comparadas con esqueletos actuales de Lama guanicoe (guanaco macho adulto utilizado de estándar) de Chile central y de Lama glama (llama hembra adulta) del norte del país, ambos depositados en el Departamento de Antropología de la Universidad de Chile. Asimismo, se utilizó el Índice de Tamaño Logarítmico (LSI), el cual posibilitó maximizar el número de medidas, ya que se consideraron en conjunto variadas secciones del esqueleto (Meadow 1999). Este conjunto de herramientas tuvo por finalidad comparar las tendencias en los cambios de tamaño a lo largo de toda la secuencia.
Perfiles etarios y explotación de camélidos
Los cambios en la explotación de camélidos son un referente para el estudio de procesos como la domesticación, puesto que las sociedades inmersas en este sistema socioeconómico aplican una serie de mecanismos para el control de hatos de animales, como el manejo de espacios de pasturas, control de natalidad, consumo, entre otros. En MAU085 (AT) el registro de camélidos (guanacos) está dominado por animales adultos y neonatos, y con un menor porcentaje de juveniles (Figura 2A). Si consideramos las evidencias contextuales, artefactuales y ecofactuales, MAU085 está relacionado con actividades de caza intensiva en la que la selección del sitio fue preponderante dada su ubicación estratégica en un pequeño cerro isla que domina el valle y la cuenca del estero aledaño. Ambos sitios arcaicos (MAU085 y MAU033) se emplazan en zonas levemente elevadas, con evidencias que indican un énfasis en la caza y recolección. Para el PAT, la baja diversidad de unidades anatómicas en camélidos no permite discutir las tendencias en la explotación de camélidos. Lo escaso de este registro deriva en una pregunta evidente, la cual se relaciona con las causas de la baja frecuencia de huesos para este periodo. Un problema que debe abordarse a futuro es el tipo de procesamiento utilizado en el PAT, con un énfasis hipotético en el aprovechamiento casi completo de una carcasa con fracturas intensivas en huesos largos y, probablemente, con asentamientos localizados estratégicamente para la caza y procesamiento primario, tal como acontece en el AT. Para el PIT, la frecuencia de restos óseos no es homogénea. El sitio con mayor número de evidencias corresponde a MAU092, en el cual dominan restos de animales adultos por sobre los juveniles. En la representación anatómica abundan restos de cráneos y huesos cortos, los cuales ingresaron al sitio adosados a huesos largos. En todos los sitios estudiados se observa una baja diversidad de restos óseos (ver ejemplo del sitio MAU092 en Figura 2B) y dominan restos de animales adultos, los cuales -debido a lo bajo de su representación- no aportan antecedentes para la discusión. Sin embargo, si se considera la hipótesis de crianza para el PIT, es plausible un control de hatos mediante el sacrificio de machos fértiles, con el fin de evitar luchas o competencias por las hembras en la época de celo (Vásquez et al. 2012). Además, en este periodo la caza de guanacos sigue siendo un sustento económico importante, por lo que el sacrificio de animales en proceso de crianza puede explicarse en el control de hatos más que para el consumo.

Figura 2. Representación anatómica expresada en %MAU en camélidos juveniles y adultos, estructura etaria expresada en MNI y resultados estadísticos entre el %MAU y Densidad Ósea (DO), Índice de Utilidad (IM), Cavidad Medular (CM) de huesos largos (húmeros, radioulnas, fémures, tibias, metapodios y primeras falanges) e Índice de Secado (IS). A. MAU085, B. MAU092 y C. MAU094. En negro, correlaciones significativas al nivel 0,05.
Para el PIT-PT, las evidencias son complejas, puesto que los sitios marcan diferencias notables en cuanto a la frecuencia y diversidad taxonómica. Considerando los restos de camélidos para el sitio MAU094 (contexto que posee un número elevado de restos faunísticos) se observa un alto porcentaje de animales adultos (59%). En este sitio se evidencia un faenamiento intensivo, como lo denota la gran cantidad de restos, a pesar del valor inverso del Índice de Cavidad Medular (Figura 2C), por lo que la fragmentación no se relacionaría con la obtención de médula. El número de animales identificados es considerable (MNI = 17) con una mayor representación de animales adultos, lo que podría asociarse a la caza de grupos familiares en época estival en el caso de los guanacos o bien al sacrificio deliberado de adultos, en el caso de los animales domésticos.
Análisis osteométrico
Mediante el análisis osteométrico, durante el AT se observa que el 85% de los especímenes medidos superan en talla tanto a los guanacos actuales de Chile Central, así como a las llamas del Norte Árido (Tabla 3 A y B, Figura 3). Sin embargo, considerando la adscripción cronológica del periodo y los datos zooarqueológicos existentes para este lapso de tiempo, los restos tanto de MAU085 como MAU033 fueron clasificados como camélidos silvestres con un morfotipo de gran tamaño. Es difícil establecer la correspondencia con la subespecie de guanaco descrita para la región, la que correspondería a Lama guanicoe huanacus, que se distribuye entre 22° y 38° S. Sin embargo, se ha discutido la validez de las subespecies y sus límites geográficos (González et al. 2006; Marín et al. 2008) como también las relaciones filogenéticas y las rutas de radiación y dispersión a lo largo de los Andes sureños (Palma et al. 2001). Cabría esperar un aumento en relación con el tamaño corporal de los guanacos del extremo norte del tamaño corporal en un gradiente norte-sur de acuerdo con la Ley de Bergmann. Esta ley señala que dentro de una especie o un grupo cercano de especies, el tamaño corporal aumenta con el decrecimiento del promedio de la temperatura anual expresada en el incremento de la latitud. No obstante, este principio ha sido cuestionado por numerosos investigadores (ver Geist 1986), y además es necesario considerar que las variaciones de tamaño también se encuentran influenciadas por la disponibilidad y calidad del forraje, la competencia intraespecífica, la relación con los predadores, entre otros (Davis 1981; Ducos y Horwitz 1997). Se sugiere que, si bien supera el tamaño del estándar utilizado, se trataría de guanacos, considerando especialmente la evidencia contextual, lo que indica grupos eminentemente cazadores y recolectores (Santander y López 2012). Para el PAT no se obtuvieron muestras medibles, debido a lo fragmentario de los huesos de este periodo. Por su parte, las medidas obtenidas para el PIT indican que los especimenes mantienen el rango observado en el AT, aunque hay muestras de menor tamaño, lo que denota un mayor rango de variabilidad. Si se consideran huesos asignables al PIT no datados directamente, se observa un conjunto en la parte inferior del rango, lo que también acontece para algunos especímenes asignados al PIT/PT (Cartajena et al. 2012). Para este periodo, destaca una primera falange posterior que presenta un tamaño muy pequeño (Figura 3A), aunque es mayor en comparación a las alpacas modernas utilizadas como referencia y otros conjuntos contemporáneos con restos de camélidos pequeños (Cartajena 2002, 2009), lo que podría indicar la presencia de un tercer taxón (vicuña/alpaca). Por otra parte aparecen por primera vez especímenes que superan en tamaño a los camélidos del AT, y se observa un rango de variabilidad mucho mayor al observado en los periodos anteriores.
Tabla 3. Resumen de mediadas obtenidas para las unidades anatómicas de cada periodo consideradas para el LSI.

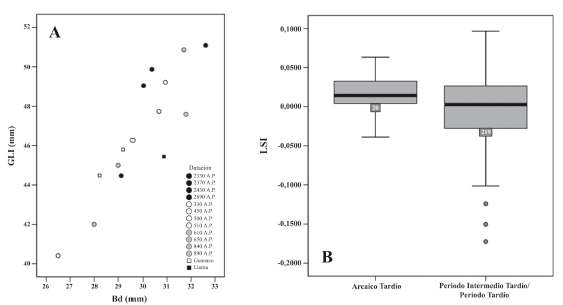
Figura 3. A. Representación de las medidas largo lateral (GLl sensu von den Driesch 1999) y ancho distal (Bd sensu von den Driesch 1999) para el astrágalo. Las medidas representadas corresponden a fechas taxón obtenidas para los astrágalos de los periodos AT, PIT y PT. Se incluyen medidas de referencia de guanaco (Gua) y llamas (Lla) actuales. B. Gráfico de cajas del Índice de Tamaño Logarítmico (LSI) que representa medidas obtenidas de las primeras, segundas falanges y astrágalos de los periodos Arcaico tardío e Intermedio tardío/Tardío.
En el caso del Índice de Tamaño Logarítmico, se consideraron las medidas de las primeras (N = 75), segundas falanges (N = 117), y astrágalos (N = 47) en conjunto, los cuales en su mayoría provienen del PIT y PT (Tabla 3). Para el caso del AT, la mediana es mayor que el esqueleto de guanaco usado como estándar; no obstante, si se considera un mayor número de medidas la dispersión es menor indicando una homogeneidad de los restos medidos. En cuanto al PIT-PT, la mediana decrece, y la varianza aumenta, producto de un conjunto heterogéneo, sobre todo en la parte inferior y superior del rango en menor medida (Figura 3B).
Isótopos estables
A partir del catastro de plantas realizado en Valle de Mauro, se identificaron especies de las familias Berberidaceae, Baraginaceae, Brassicaceae, Dryopteridaceae, Woodsiaceae, Ephedraceae, Apiaceae, Asclepiadaceae, Asteraceae, Cactaceae, Cakycereaceae, Camoanulaceae, Caryophyllaceae, Chenopodiaceae, Convolvulaceae, Euphorbiaceae, Fabaceae, Gentianaceae, Hydrophyllaceae, Onagraceae, Oxalidaceae, Phytolacaceae, Plantaginaceae, Portulacaceae, Ranunculaceae, Rosaceae, Rubiaceae, Santalaceae, Scrophulariaceae, Solanaceae, Valerianaceae, Verbenaceae, Violaceae, Viviniaceae, Alliaceae, Amaryllidaceae, Cyparaceae, Iriadaceae, Juncaceae, Orchidaceae y Poaceae (Gramineae) (Belmar y Quiroz 2012). En su gran mayoría, estas plantas poseen un ciclo fotosintético de tipo C3, con una escasa inclusión de plantas del tipo C4, como algunas especies de Portulacaceae, además de plantas del tipo CAM. La totalidad de las muestras (N = 34) presentan una razón C/N con rangos aceptables (2.8 y 3.6), lo que se aprecia también en las concentraciones de C y N, por lo que los resultados isotópicos son confiables por una buena preservación del colágeno (Ambrose 1990; Pate 1994). En la Figuras 4A y 4B se observa la distribución de los resultados de ä13Ccol y ä15Ncol tanto por sitio como por periodo según asociaciones culturales y distribuciones estratigráficas, considerando el AT, PAT y PIT-PT por separado. La media de ä13Ccol para los camélidos es de -19.99 ± 1.69, mientras que para el ä15Ncol la media es de 5.79 ± 1.09, con una correlación negativa (r = -0.36, p = 0.19) entre la totalidad de los valores de ä13Ccol y ä15Ncol, sin embargo, no es estadísticamente significativa. Los valores del AT y PAT mantienen intervalos similares, y la dieta de los animales de ambos periodos se concentra en plantas C3, no obstante, en el caso del PAT se observan algunos outliers. Para el PIT/ PT, los valores tanto de carbono como de nitrógeno presentan un mayor intervalo con un valor de ä13Ccol máximo de -16.6 y un valor mínimo de -24.9, mientras que en el caso de ä15Ncol el valor máximo es 8.7, con un mínimo de 3.7. Si se consideran únicamente los huesos fechados directamente por 14C los valores isotópicos presentan prácticamente una misma distribución en comparación con la separación realizada por periodo, desde el AT hasta las últimas ocupaciones incaicas en el valle (Figuras 5A y 5B).

Figura 4. A. Gráfico de dispersión con los valores de ä13Ccol y ä15Ncol (camélidos) por sitio. B. Gráfico de cajas con los valores de ä13Ccol y ä15Ncol (camélidos) por periodo.

Figura 5. A. Relación entre datos cronológicos e isótopos de carbono en muestras de camélidos datados directamente por 14C. B. Relación entre datos cronológicos e isótopos de nitrógeno en m uestras de camélidos datados directamente por 14C.
En cambio, la varianza en los isótopos de ä15Ncol hacia el PT presenta dos patrones diferentes muy claros a partir del 500 DC. Lo mismo sucede con los isótopos de ä13Ccol, donde se observa la misma tendencia pero un poco más temprana alrededor del 1000 DC. Llama la atención que estos cambios en la alimentación coinciden con los observados en la composición taxonómica de los camélidos. En relación con los datos osteométricos, como se mencionó anteriormente para el AT, los camélidos corresponden a un morfotipo silvestre de gran tamaño, el cual se mantiene hacia el PIT-PT, aunque con la inclusión para este periodo de animales de mayor y menor talla, posiblemente llamas de gran tamaño (cargueras) y animales domésticos de morfotipo pequeño, incluso algunos similares en talla a la alpaca. Los cambios isotópicos se vincularían más bien con posibles diferencias en la alimentación debido a la utilización de áreas de pastura y, en el caso de aquellas muestras con leve inclusión de C4, podrían reflejar una procedencia distinta, que se reflejaría en una alimentación diferente.
Paleoambiente
Los antecedentes paleoambientales para el NSA señalan que hacia los ~4200 años cal. AP se produce una expansión de la vegetación de humedales, y en los ~3200 años cal. AP la contracción del bosque, lo que coincide con las ocupaciones AT; entre los ~1800 a 1300 años cal. AP se produce la expansión de la vegetación arbustiva, evento que fue el marco para las ocupaciones PAT, mientras que para el PAT/PIT hacia los ~1300 años cal. AP se estabilizan las condiciones actuales (Maldonado y Villagrán 2002, 2006). Estos antecedentes indican que los cambios climáticos no fueron determinantes en las modificaciones en la composición de la dieta de los camélidos del valle, por cuanto reflejan transformaciones relacionadas con la expansión y contracción de la vegetación arbustiva, y no con cambios radicales en la cobertura. El cambio más drástico se vincula a las ocupaciones AT, debido a la contracción del bosque, lo que constituye un aspecto a investigar a futuro y que puede relacionarse con la gran talla alcanzada por los guanacos para este periodo.
CONCLUSIONES
Dentro de las variables utilizadas en el presente análisis, tanto la osteometría como los análisis isotópicos son los que aportaron un mejor cuerpo de datos para la discusión acerca de la presencia de camélidos domésticos en el valle. Al respecto, la economía durante el PIT está fuertemente vinculada a la actividad agrícola, con asentamientos dispersos y una explotación a baja escala, en los que la caza de camélidos cumplió un rol relevante (Becker 2004). No obstante, la aparición de camélidos más pequeños a los observados durante el AT sugiere que se trataría de la incorporación de taxones domésticos a baja escala, acorde con asentamientos dispersos con sustento agrícola. Pensamos que el modelo podría corresponder al comienzo de una economía mixta (Olivera 2001; Izeta 2007; Figueroa 2013). Nos parece importante recoger la crítica en cuanto en general existe siempre la supremacía de una actividad sobre la otra (Figueroa 2013). Entendemos que en el caso de las poblaciones del PIT, la agricultura habría jugado un rol central y que la ganadería habría sido un complemento de ella, donde los animales pastan a campo abierto en áreas marginales de la agricultura junto con la caza. La incorporación de animales domésticos podría relacionarse con el aumento de las redes de interacción, incluso con tradiciones trasandinas con una economía de base agrícola complementada con la recolección, la caza y con el pastoreo de camélidos (Bárcena 2001). No se observan los cambios aparejados a un proceso local de domesticación (Meadow 1989); además, la fecha es muy tardía en relación con lo observado en otros centros de domesticación. La caza del guanaco seguiría siendo una actividad relevante, mientras que la crianza de animales domésticos se enfocaría a la mantención de pequeños conjuntos de animales para el consumo y obtención de fibras para trabajo textil, como lo denotan los artefactos utilizados para este tipo de actividad en sitios PIT del NSA (López et al. 2012). Los antecedentes isotópicos indican variaciones que pueden relacionarse con camélidos que habitaban regiones fitográficas de altitudes distintas y/o con variaciones intertaxonómicas en la dieta de estos animales (Fernández y Panarello 1999-2001; Gil et al. 2010), sobre todo para el PIT y PT.
La escasa variación morfométrica e isotópica para el AT y el PAT indica más bien un énfasis en la cacería de guanacos de gran tamaño y sin rastros de evidencias de crianza. Para el PT, con la llegada del Inca comienza una mayor intensificación en la explotación de recursos asociados a un incremento en la población y diversificación de roles sociales. Para este periodo, la presencia de llama especializada como animal de carga es clara y esperable para momentos asociados a la presencia del Inca (Mengoni Goñalons 2007). No obstante, lo interesante es la mayor diversificación en los tamaños y dieta de los camélidos, que refleja una mayor complejidad en el manejo de rebaños, complementados mediante la caza de guanacos que habitaron en el valle. Sobre lo mismo, debemos considerar múltiples escenarios, tales como los manejos incaicos de sus propios rebaños, la influencia de estas técnicas en las poblaciones locales y la mantención de sistemas de crianza por las comunidades diaguitas, y que no sufrieron cambios durante el PT. Para el primer escenario, es esperable para las llamas cargueras una crianza en áreas disímiles y, por tanto, una dieta diversificada, mientras que otro grupo puede presentar una dieta más controlada, menos diversa y con animales asociados espacialmente a sitios habitados por elites. Para el segundo escenario, es decir, poblaciones diaguitas vinculadas al Tawantinsuyu, son esperables rebaños de animales mantenidos localmente en asentamientos periféricos y encargados de la producción de bienes. Bajo este contexto, es esperable para los rebaños una movilidad menos extensa, restringida dentro del mismo valle y sectores aledaños (costa- interior, por ejemplo). El último escenario ha sido discutido anteriormente, y sus principales implicancias arqueológicas radican en morfotipos de menor tamaño criados en las cercanías de los asentamientos para la producción a baja escala de textiles y como recursos alimenticios, además de la conexión con la costa para el traslado de recursos. Esto podría reflejar un cambio agrícola complementado con la ganadería, que da lugar al desarrollo de un nuevo sistema productivo, de acuerdo con el cual el forrajeo de camélidos es restringido artificialmente por los humanos (Figueroa 2013). Estos escenarios requieren de un análisis contextual más detallado al realizado en el presente trabajo, en el que se diferencie funcionalidad de sitios, asociaciones con otras evidencias y un mayor correlato con rastros como coprolitos, micro y macrorrestos vegetales, entre otros. El ejercicio realizado en primera instancia por medio de los datos referentes a la explotación y perfiles etarios de camélidos de los sitios más relevantes por periodo aporta algunas pistas para esta discusión, pero dichos datos no son resolutivos. Por lo mismo, este es un problema de investigación que a futuro debe abordarse con más detalle. Las evidencias de llamas cargueras son claras para el PT, e implican que en el valle se mantuvieron rebaños de estos animales y por ende, a futuro, se deben discutir con mayor detalle evidencias contextuales relacionadas con esta mantención de camélidos para la carga. Como se mencionó anteriormente, el sacrificio de camélidos adultos (tanto llamas como guanacos) dentro de la lógica Inca no es explicable únicamente por el consumo nutricional, sino que también estuvo inmersa dentro de lógicas simbólicasrituales (Brotherson 1989). En el caso de las muestras analizadas, este último punto es complejo de abordar arqueológicamente en el valle, mientras que para el consumo es evidente para algunos sitios la producción de ch'arki y el uso de huesos para la elaboración de artefactos para la producción textil al igual que en el PIT (López et al. 2012).
Cabe destacar que el análisis secuencial realizado en el presente trabajo no se enmarca bajo una lógica de desarrollo unilineal, sino que más bien aporta a la discusión y ratifica la complejidad de trabajar con muestras de camélidos a distintas escalas. Un buen referente de lo anterior es el análisis osteométrico, ya que las evidencias de un morfotipo de guanaco de gran tamaño, sobre todo para el AT y PAT, reflejan la dificultad de trabajar con estándares y patrones que no representan la totalidad de los escenarios de cambios en las poblaciones animales. Por esto, los resultados del presente trabajo no son extrapolables a todo el NSA, donde cada cuenca y valle requieren de un análisis propio con el fin de identificar tendencias regionales como particularidades para cada área de estudio. Las evidencias aportadas, a su vez, motivan a futuro nuevas líneas de trabajo, como el uso diferenciado del espacio así como la construcción de paisajes en función de una economía agrícola y criancera, lo que diferiría de los modelos agroganaderos anteriormente propuestos.
Agradecimientos
El presente estudio fue financiado por el Proyecto Arqueológico Mauro. Nuestros agradecimientos a Daniela Villalón, Douglas Jackson, Bárbara Rivera, Camila Opazo, María Paz Casorzo y Javiera Bastías por su asistencia durante el proceso de análisis del material de los sitios abordados en el presente trabajo. Por último, nuestros agradecimientos a los tres evaluadores anónimos, que permitieron mejorar el manuscrito original, y al equipo editorial de Revista Intersecciones en Antropología.
REFERENCIAS CITADAS
1. Ambrose, S. H. 1990 Preparation and characterization of bone and tooth collagen for stable carbon and nitrogen isotope analysis. Journal of Archaeological Science 17: 430-451. [ Links ]
2. Bárcena, R. 2001 Prehistoria del Centro-Oeste Argentino. En Historia Argentina Prehispánica, editado por E. E. Berberián y A. E. Nielsen, pp. 561-634. Brujas, Córdoba. [ Links ]
3. Becker, C. 2004 Animales que cuentas historias. Chungara 36, Volumen Especial (1): 359-364. [ Links ]
4. Belmar, C. y L. Quiroz 2012 Informe de los restos arqueobotánicos del Valle de Mauro. Proyecto Arqueológico Mauro, Santiago. MS. [ Links ]
5. Borrero, L. A. 1990 Fuego-Patagonian Bone Assemblages and the Problem of Communal Guanaco Hunting. En Hunters of the Past, editado por L. B. Davis y B. O. K. Reeves, pp. 373-399. Unwin Hyman, Londres. [ Links ]
6. Brotherson, G. 1989 Andean pastoralism and Inka ideology. En The walking leader. Patterns of domestication, pastoralism and predation, editado por J. Clutton-Brock, pp. 240- 255. One World Archaeology, Unwin Hyman, Londres. [ Links ]
7. Cartajena, I. 2002 Los conjuntos arqueofaunísticos del Arcaico Temprano en la Puna de Atacama, Norte de Chile. Tesis Doctoral inédita. Freie Universität Berlin. Impresa en microfilm, ABESY Vertriebs GmbH. [ Links ]
8. Cartajena, I 2009 Explorando la variabilidad morfométrica del conjunto de camélidos pequeños durante el Arcaico Tardío y el Formativo Temprano en Quebrada Tulan, norte de Chile. Revista del Museo de Antropología 2: 199-212. [ Links ]
9. Cartajena, I 2013 Faunal assemblages from the Middle Holocene: environmental and cultural variability in the western slope of the Puna de Atacama. Quaternary International 307: 31-37. [ Links ]
10. Cartajena, I., L. Nuñez y M. Grosjean 2007 Camelid domestication in the western slope of the Puna de Atacama, Northern Chile. Anthropozoologica 42 (2): 155-173. [ Links ]
11. Cartajena, I., P. López, B. Rivera y B. Santander 2012 Introducción de taxones domésticos y control de camélidos en el Norte Semiárido: variabilidad osteométrica en el Valle de Mauro, IV Región, Chile. Revista Chilena de Antropología 30: 98-103. [ Links ]
12. Castillo, G. 1984 Un cementerio del Complejo Las Ánimas en Coquimbo: Ejemplo de relaciones con San Pedro de Atacama. Estudios Atacameños 7: 199-209. [ Links ]
13. Cornejo, L. y D. Jackson 2004 Un Panorama del Patrón de Asentamientos en los Esteros Conchalí-Pupío (Los Vilos): Entre el Interior y la Costa. Revista Werken 5: 83-87. [ Links ]
14. Dantas, M. 2012 Identificación interspecífica de camélidos en el valle de Ambato (Catamarca, Argentina): una aproximación a la problemática de distintas líneas de análisis. Revista del Museo de Antropología 5: 259-268. [ Links ]
15. Davis, S. J. 1981 The effects of temperature change and domestication on the body size of late Pleistocene to Holocene mammals of Israel. Paleobiology 7: 101-114. [ Links ]
16. De Nigris, M. y G. L. Mengoni Goñalons 2004 El guanaco como fuente de carne y grasas en Patagonia. En Contra viento y marea. Arqueología de Patagonia, editado por M. T. Civalero, P. M. Fernández y A. G. Guráieb, pp. 469-476. Sociedad Argentina de Antropología (SAA) e Instituto Nacional de Antropología y Pensamiento Latinoamericano (INAPL), Buenos Aires. [ Links ]
17. Driesch, A. von den 1999 A guide to measurement of animal bones from archaeological sites. Peabody Museum Bulletins 1. Harvard University, Cambridge. [ Links ]
18. Ducos, P. y L. K. Horwitz 1997 The Influence of climate on artiodactyl size during the Late Pleistocene-Early Holocene of the Southern Levant. Paléorient 23 (2): 229-247. [ Links ]
19. Elkin, D. C. 1996 Arqueolozoología de Quebrada Seca 3. Indicadores de subsistencia humana temprana en la Puna Meridional Argentina. Tesis Doctoral inédita. Facultad de Filosofía y Letras, Universidad de Buenos Aires, Buenos Aires. [ Links ]
20. Fernández, J. y H. Panarello 1999-2001 Isótopos del carbono en la dieta de herbívoros y carnívoros de los Andes jujeños. Xama 12- 14: 71-85. [ Links ]
21. Figueroa, G. 2013 Estrategias productivas en Aguada de Ambato (Catamarca, Argentina). Relaciones de la Sociedad Argentina de Antropología XXXVIII (1): 111-135. [ Links ]
22. Finucane, B., P. Maita Agurto y W. Isbell 2006 Human and animal diet at Conchopata, Peru: stable isotope evidence for maize agriculture and animal management practices during the Middle Horizon. Journal of Archaeological Science 33: 1766-1776. [ Links ]
23. Geist, V. 1986 Bergmann´s rule is invalid. Canadian Journal of Zoology 65: 1035-1038. [ Links ]
24. Gil, A., G. Neme y R. Tykot 2010 Isótopos estables y consumo de maíz en el centro occidente argentino: tendencias temporales y espaciales. Chungara 42 (2): 497-513. [ Links ]
25. González, B., E. Palma, B. Zapata y J. C. Marín 2006 Taxonomic and biogeographical status of guanaco Lama guanicoe (Artiodactyla, Camelidae). Mammal Review 36 (2): 157-178 [ Links ]
26. Grant, J. 2010 Aporte de distintas técnicas osteométricas para la diferenciación interespecífica de camélidos sudamericanos. En Zooarqueología a principios del siglo XXI. Aportes teóricos, metodológicos y casos de estudio, editado por M. Gutiérrez, M. De Nigris, P. Fernández, M. Giardina, A. Gil, A. Izeta, G. Neme y H. Yacobaccio, pp. 17-28. Libros del Espinillo, Buenos Aires. [ Links ]
27. Hesse, B. 1982 Archaeological evidence of camelid explotation in the Chilean Andes. Säugetierkundliche Mitteilungen 30 (2): 201-211. [ Links ]
28. Izeta, A. D. 2007 Zooarqueología del sur de los valles Calchaquíes (Provincia de Catamarca y Tucumán, República Argentina). Análisis de conjuntos faunísticos del primer milenio A.D. BAR International Series 1612. Archaeopress, Oxford. [ Links ]
29. Izeta, A. D. 2009 Introducción al Dossier: Osteometría de camélidos. Aportes metodológicos desde la arqueología sudamericana. Revista de Antropología 2: 125-127 [ Links ]
30. Izeta, A. D. 2010 Variabilidad osteométrica en sitios arqueológicos del NOA. En Zooarqueología a principios del siglo XXI. Aportes teóricos, metodológicos y casos de estudio, editado por M. Gutiérrez, M. De Nigris, P. Fernández, M. Giardina, A. Gil, A. Izeta, G. Neme y H. Yacobaccio, pp. 29-38. Libros del Espinillo, Buenos Aires. [ Links ]
31. Izeta, A. D., C. Otaola y A. Gasco 2009a Estándares métricos y variabilidad en falanges proximales de camélidos sudamericanos. Su importancia como conjunto comparativo para interpretaciones en arqueología. Revista del Museo de Antropología 2: 169-180. [ Links ]
32. Izeta, A., A. Laguens, B. Marconetto y M. Scattolin 2009b Camelid handling in the meridional Andes during the first millennium AD: A preliminary approach using stable isotopes. International Journal of Osteoarchaeology 19: 204-214. [ Links ]
33. Kaufmann, C. 2004 La fusión ósea como indicador de edad y estacionalidad en guanaco (Lama guanicoe). En Contra viento y marea: Actas de las V Jornadas de Arqueología de la Patagonia, editado por T. Civalero, G. Guraieb y P. Fernández, pp. 477-487. SAA e INAPL, Buenos Aires. [ Links ]
34. Kent, J. D. 1982 The Domestication and exploitation of the South American camelids: methods of análisis and their application to circum-lacustrine archaeological sites in Bolviia and Peru. Tesis Doctoral inédita. Washington University, St. Louis. [ Links ]
35. L`Heureux, G. L. 2010 Morfometría de camélidos sudamericanos modernos. La variabilidad morfológica y diversidad taxonómica. En Zooarqueología a principios del siglo XXI. Aportes teóricos, metodológicos y casos de estudio, editado por M. Gutiérrez, M. De Nigris, P. Fernández, M. Giardina, A. Gil, A. Izeta, G. Neme y H. Yacobaccio, pp. 39-50. Libros del Espinillo, Buenos Aires. [ Links ]
36. López, P., I. Cartajena, B. Santander, B. Rivera y C. Opazo 2012 Explotación de camélidos de un sitio Intermedio Tardío (1.000-1.400 d.C.) y Tardío (1.400-1.536 d.C.) del Valle de Mauro (IV Región, Chile). Boletín de la Sociedad Chilena de Arqueología 41-42: 91-108. [ Links ]
37. Lyman, R. L. 1994 Vertebrate Taphonomy. Cambridge University Press, Cambridge. [ Links ]
38. Maldonado, A. y C. Villagrán 2002 Paleoenvironmental changes in the semiarid coast of Chile (~32ºS) during the last 6200 cal years inferred from a swampforest pollen record. Quaternary Research 58: 130-138. [ Links ]
39. Maldonado, A. y C. Villagrán 2006 Variability of the Northern limit of the Southern Westerlies over the last 9900 cal yr BP from a swamp forest pollen record along the semiarid coast of Chile (32º05'S). Quaternary Research 66: 246-258. [ Links ]
40. Marín J. C., A. Spotorno, B. A. González, C. Bonacic, J. C. Wheeler, C. S. Casey, M. W. Bruford, E. Palma y E. Poulin 2008 Mitochondrial DNA variation and systematics of the Guanaco (Lama guanicoe, Artiodactyla: Camelidae). Journal of Mammalogy 89 (2): 269-281. [ Links ]
41. Marino, B. y M. McElroy 1991 Isotopic composition of atmospheric CO2 inferred from carbon in C4 plant cellulose. Nature 349: 127-131. [ Links ]
42. Meadow, R. H. 1989 Osteological evidence for the process of animal domestication. En The Walking Larder: Patterns of Domestication, Pastoralism, and Predation, editado por J. Clutton-Brock, pp. 980-990. Unwin Hyman, Londres. [ Links ]
43. Meadow, R. H. 1999 The use of size index scaling techniques for research on archaeozoological collections from the Middle East. En Historia Animalium ex Ossibus. Beiträge zur Paläoanatomie, Archäologie, ägyptologie, Ethnologie und Geschichte der Tiermedizin, editado por C. Becker, H. Manhart, C. Peters y J. Schibler, pp. 285-300. Festschrift für Angela von den Driesch zum 65. Geburtstag. Internationale Archäologie, Studia Honoraria 8. Rahden/Westfalen. [ Links ]
44. Méndez, C. y D. Jackson 2004 Ocupaciones humanas del Holoceno Tardío en Los Vilos (IV Región, Chile): Origen y características conductuales de la población local de cazadores recolectores del litoral. Chungara 36 (2): 279-293. [ Links ]
45. Méndez, C. y D. Jackson 2008 La ocupación prehispánica de Combarbalá (Norte Semiárido, Chile): una propuesta sintética. Chungara 40 (2): 5-17. [ Links ]
46. Méndez, C., A. Troncoso, D. Jackson y D. Pavlovic 2009 Movilidad y uso del espacio entre cazadoresrecolectores tardíos en espacios cordilleranos del Norte Semiárido de Chile. Intersecciones en Antropología 10 (2): 311-326. [ Links ]
47. Mengoni Goñalons, G. L. 1996 La domesticación de los camélidos sudamericanos y su anatomía económica. En Zooarqueología de Camélidos 2, editado por D. Elkin, C. Madero, G. Mengoni-Goñalons, D. Olivera, M. Reigadas y H. Yacobaccio, pp. 3-45. Grupo de Zooarqueología de Camélidos, Buenos Aires. [ Links ]
48. Mengoni Goñalons, G. L. 2007 Camelid management during Inca times in N.W. Argentina: models and archaeozoological indicators. Antropozoologica 42 (2): 129-141. [ Links ]
49. Mengoni Goñalons, G. L. 2008 Camelids in ancient Andean societies: A review of the zooarchaeological evidence. Quaternary International 185: 59-68. [ Links ]
50. Miller G. y R. Burger 1995 Our father the cayman, our dinner the llama: animal utilization at Chavín de Huantar, Peru. American Antiquity 60 (3): 421-458. [ Links ]
51. Olivera, D. E. 2001 Sociedades agropastoriles tempranas: El Formativo Inferior del Noroeste Argentino. En Historia argentina prehispánica, editado por E. E. Berberián y A. E. Nielsen, pp. 83-125. Brujas, Córdoba. [ Links ]
52. Palma, R. E., J. C. Marín, A. E. Spotorno, y J. L. Galaz 2001 Phylogenetic relationship among South American subspecies of camelids based on sequences of the cytochrome b mitochondrial genes. En Progress in South American camelids research. Proceedings of the 3rd European Symposium on South American Camelids and SUPREME European Seminar (27-29 Mai 1999, Göttingen), editado por M. Gerken y C. Reinieri, pp. 44-52. EAAP Publication 105, Göttingen. [ Links ]
53. Pate, F. D. 1994 Bone chemistry and paleodiet. Journal of Archaeological Method and Theory 1: 161-209. [ Links ]
54. Pavlovic, D. 2004 Dejando atrás la tierra de nadie: Asentamientos, contextos y movilidad de las comunidades Alfareras Tempranas del Choapa. Werken 5: 39-46. [ Links ]
55. Santander, B. y P. López 2012 Análisis de Microhuellas de Uso Mediante Microscopio Electrónico de Barrido (MEB) de Artefactos Óseos de un Sitio Arcaico Tardío del Valle de Mauro (Región de Coquimbo, Chile): Aportes para una Reconstrucción Contextual. Revista Chilena de Antropología 26: 129-150. [ Links ]
56. Troncoso, A. 2001 La cultura Diaguita en el 2001, problemas y perspectivas desde el Choapa. Actas del IV Congreso Chileno de Antropología, t. II: 1351-1356. Colegio de Antropólogos de Chile, Santiago. [ Links ]
57. Troncoso, A. 2004 Relaciones socio-culturales de producción, formas de pensamiento y ser en el mundo: un acercamiento a los Periodos Intermedio Tardío y Tardío en la cuenca del Río Choapa. Werken 5: 61-67. [ Links ]
58. Troncoso, A., C. Becker, D. Pavlovic, P. González, J. Rodríguez y C. Solervicens 2009 El sitio LV099-B Fundo Agua Amarilla y la ocupación del periodo incaico en la costa de la provincia del Choapa. Chungara 41 (2): 241-259. [ Links ]
59. Vásquez, V., T. Rosales, I Rey y G. Dorado 2012 Pruebas zooarqueológicas e isotópicas de crianza local de camélidos en la Zona Urbana Moche de los complejos arquitectónicos Huacas del Sol y de la Luna en Trujillo (Perú). Archaeobios 6 (1): 19-32. [ Links ]
60. Veit, H. 1993 Upper Quaternary landscape and climate evolution in the Norte Chico (Northern Chile): An overview. Mountain Research Development 13: 139-144. [ Links ]
61. Wheeler, J. C. 1982 Aging llamas and alpacas by their teeth. Llama World I: 12-17. [ Links ]
62. Yacobaccio, H. D. 2010 Osteometría de llamas (Lama glama) y sus consecuencias arqueológicas. En Zooarqueología a principios del siglo XXI. Aportes teóricos, metodológicos y casos de estudio, editado por M. Gutiérrez, M. De Nigris, P. Fernández, M. Giardina, A. Gil, A. Izeta, G. Neme y H. Yacobaccio, pp. 65-76. Del Espinillo, Buenos Aires. [ Links ] [ Links ]
