










Services on Demand
Journal

Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO
Related links
-
 Similars in
SciELO
Similars in
SciELO
Share

 Permalink
PermalinkIntersecciones en antropología
On-line version ISSN 1850-373X
Intersecciones antropol. vol.16 no.2 Olavarría Sept. 2015
ARTÍCULOS
Procesamiento y consumo de pinnípedos: el caso de las ocupaciones canoeras tempranas del sitio Imiwaia I (Tierra del Fuego, República Argentina)
María Paz Martinoli
María Paz Martinoli. Centro Austral de Investigaciones Científicas (CADIC). Consejo Nacional de Investigaciones Científicas y Técnicas (CONICET). Bernardo Houssay 200 (Ushuaia, Tierra del Fuego). E-mail: mpmartinoli@yahoo.com.ar
Recibido 27 de noviembre 2013.
Aceptado 7 de junio 2014
RESUMEN
El objetivo general de este artículo es analizar los patrones de procesamiento y consumo de pinnípedos en las ocupaciones canoeras tempranas de la región del canal Beagle. A tal efecto, se evalúan las modalidades explotación de dichos mamíferos marinos a través de los conjuntos óseos recuperados en depósitos arqueológicos del sitio Imiwaia I (Tierra del Fuego, Argentina) con cronologías cercanas a 6000 AP. Se presentan los perfiles de edad y sexo de los individuos explotados, así como el estudio detallado de las modificaciones óseas. Los resultados muestran: a) un predominio de especímenes juveniles/subadultos de sexo indeterminado y machos adultos de Arctocephalus australis; b) la mayoría de las unidades anatómicas de tales individuos están representadas; y c) las marcas de corte se encuentran concentradas sobre las regiones proximales y distales de las diáfisis de huesos largos y sobre sectores proximales y mediales de las costillas. Estos datos permitirían sostener que la captura de la generalidad de las presas identificadas posiblemente no se produjo en loberías de reproducción. Los individuos probablemente ingresaron enteros al sitio, donde sufrieron una desarticulación primaria y luego la reducción en porciones más pequeñas.
Palabras clave: Canal Beagle; Ocupaciones tempranas; Pinnípedos; Explotación.
ABSTRACT
Pinniped processing and consumption: early littoral occupations at Imiwaia I site, Tierra del fuego, República Argentina.
The main objective of this article is to analyze the processing and consumption patterns of pinnipeds in the early human occupations of the Beagle Channel region. In order to achieve this aim, the modes of exploitation of these marine mammals is evaluated on the basis of the bone assemblages recovered from archaeological deposits at Imiwaia I site (Tierra del Fuego, Argentina), dated to around 6000 years BP. The age and sex profiles of exploited animals is presented, as well as a detailed study of bone modifications. The results show: a) a clear predominance of juvenile/subadult indeterminate sex and male adult specimens of Arctocephalus australis; b) all anatomical units of these individuals are present; and c) cut marks are mostly represented on the proximal and distal regions of long bone diaphyses and on the proximal and medial areas of ribs. These data support the view that it is unlikely that most of the identified prey were captured in rookeries. The animals arrived whole at the site, where they underwent primary disarticulation and then were butchered into smaller portions.
Keywords: Beagle Channel; Early occupations; Pinnipeds; Exploitation.
INTRODUCCIÓN
Los trabajos arqueológicos que se han desarrollado en la región del canal Beagle permitieron generar un sólido modelo de "adaptación a la vida litoral" (Schiavini 1990, 1993; Orquera y Piana 1999: 96, 2005, 2009). Este hace énfasis principalmente sobre dos aspectos. Primero, que los pinnípedos fueron las presas que aportaron el mayor ingreso energético a la dieta de los grupos cazadores-recolectores que habitaron allí durante el Holoceno medio y tardío (Schiavini 1990, 1993; Orquera y Piana 1999, 2005; Zangrando 2003, 2009a y b; Tivoli 2010; Tivoli y Zangrando 2011). Segundo, que tales sociedades emplearon tecnología especializada que permitió aumentar la eficacia y previsibilidad de captura de estos mamíferos marinos (Schiavini 1990, 1993; Orquera y Piana 1999). Aunque estas pautas son conocidas desde momentos iniciales de la secuencia arqueológica, aún no se ha profundizado sobre algunos aspectos fundamentales relacionados con los patrones de procesamiento y consumo de estos recursos. El objetivo general de este trabajo es evaluar las pautas de captura, procesamiento y consumo de pinnípedos a través de los conjuntos óseos recuperados en las capas basales del sitio Imiwaia I (Tierra del Fuego, Argentina) con fechados cercanos a 6000 años AP. En términos generales y de manera simplificada, la explotación de una presa implica como mínimo cuatro etapas: 1) la captura; 2) el transporte; 3) el procesamiento; y 4) el consumo (Lyman 1992). Cada una de ellas puede ser entendida considerando determinada cantidad de variables, pero evidentemente se encuentran interrelacionadas y se influyen recíprocamente. En lo que respecta a la captura de pinnípedos, se han considerado como especialmente importantes dos factores para analizar las estrategias de aprovisionamiento en el archipiélago fueguino: el comportamiento y la distribución de los pinnípedos en el ambiente (Vidal y Winograd 1986; Lanata y Winograd 1988; Lyman 1989, 1995, 2003; Schiavini 1990, 1993; Lanata y Borrero 1994; Muñoz 1996, 2002, 2004, 2005; Orquera y Piana 1999) y la tecnología empleada para su explotación (Lanata y Winograd 1988; Lyman 1989, 1995; Schiavini 1990, 1993; Hildebrandt y Jones 1992; Orquera y Piana 1999). Los pinnípedos que están actualmente disponibles en la región del canal Beagle pertenecen a dos especies, ambas de la familia Otariidae: los lobos marinos de dos pelos o lobos finos sudamericanos (Arctocephalus australis) y los lobos marinos de un pelo o leones marinos (Otaria flavescens). Ambas especies son poligínicas y su ciclo anual se divide en una etapa reproductiva corta y un período de visitas regulares a áreas costeras (Crespo et al. 2008a y b). Para entender las estrategias de aprovisionamiento de pinnípedos practicadas por los cazadores-recolectores de la región del canal Beagle, es necesario especificar el comportamiento reproductivo y la distribución socioespacial. La temporada de parición de los lobos marinos de dos pelos se da entre los meses de noviembre y diciembre, mientras que la de los lobos marinos de un pelo se produce en los meses de enero y febrero, acorde con los datos publicados por Sielfeld (1983, 1999).
No obstante, según Schiavini (1990) las pariciones de A. australis se dan principalmente en diciembre, y las de O. flavescens, en enero. Durante estos períodos reproductivos se ha observado el establecimiento de dos tipos distintos de apostaderos: a) de juveniles (sin actividad reproductiva); y b) de cría (predominio de cachorros, hembras y machos adultos territoriales) (Crespo et al. 2008a y b). Fuera del período reproductivo y como parte del ciclo (Campagna 1985), dichos otáridos forman apostaderos de descanso, que pueden coincidir o no con las áreas de colonias de reproducción, cuya estructura social presenta cierta variabilidad (edades y sexos variables) (Crespo et al. 2008a y b). Sin embargo, en términos generales, los machos adultos no territoriales (periféricos) y los individuos inmaduros son segregados agrupándose en asentamientos específicos, pero con una tendencia a permanecer más tiempo en el agua que en tierra (efectúan largos desplazamientos en busca de alimentos), mientras que las hembras regresan continuamente a las zonas de crías para amamantar a los cachorros, siendo sus movimientos más restringidos en tiempo y espacio (Vaz Ferreira y Vallejo 1981; Crespo et al. 2008a y b; Bowen et al. 2009). Los lobos marinos de dos pelos se ubican preferentemente sobre costas rocosas y con cierta exposición al oleaje (Sielfeld 1983); en el caso de Tierra del Fuego, la mayoría de las colonias reproductivas se encuentran en sectores de islas y costas exteriores del archipiélago (Schiavini 1990; Crespo et al. 2008a: 2), tal es el caso de las múltiples áreas de parición y cría en Isla de los Estados e Isla Observatorio (Schiavini y Raya Rey 2001). Sin embargo, se han relevado apostaderos de descanso en sectores interiores del canal Beagle, como por ejemplo en el islote Les Eclaireurs Oeste (Crespo et al. 2008a: 3), que en un censo realizado en 1992 mostraba un total de 895 individuos y una clara tendencia hacia el crecimiento poblacional (Schiavini y Raya Rey 2001). Por el contrario, los lobos marinos de un pelo eligen costas con declives más suaves y más reparadas para establecer sus colonias reproductivas y no reproductivas, por lo cual pueden ser halladas tanto en islas e islotes dentro del canal Beagle (e.g., Islas Becasses), como en sectores exteriores (e.g., colonia Las Chapas, Península Mitre) (Sielfeld 1983, Schiavini 1990; Schiavini y Raya Rey 2001; Crespo et al. 2008b: 9).
Extensos trabajos previos llevados a cabo en la región indagaron directamente sobre las estrategias de subsistencia de los grupos que habitaron el área en momentos tempranos de la secuencia de ocupación (6000-4500 años AP). En el Segundo Componente de Túnel I (capas D y E), más del 65% (en términos de NISP) de los restos de fauna corresponden a pinnípedos. En dicho conjunto, se registra un MNI de Arctocephalus australis de 344 individuos y de 9 para Otaria flavescens. De dichos totales, el 80% corresponde a machos, de los cuales el 69% son ejemplares subadultos (Schiavini 1990; Orquera y Piana 1999; Orquera 2011). Dicho perfil etario y de sexo es, para los autores, compatible con la captura en el agua de individuos aislados -dada la tecnología de arpones de punta desmontable y canoas-, ya que predominan los machos adultos y subadultos de A. australis -los cuales, como se explicó previamente, se encuentran ligados por períodos más cortos a las loberías-, y sus apostaderos reproductivos se encuentran en sectores exteriores del archipiélago fueguino. Pero principalmente, el análisis de los cortes delgados realizados en dientes indicó que el 90% fue consumido en los seis meses comprendidos entre marzo y septiembre, y a partir de esta determinación, Schiavini (1990, 1993) propuso que los grupos cazadores-recolectores importaban energía hacia el sistema costero durante el invierno a través del consumo de presas pelágicas que ingresaban de manera recurrente al canal Beagle. En lo que refiere a las pautas de procesamiento y consumo, pocos trabajos arqueológicos de Tierra del Fuego presentan información al respecto, aunque sí es de destacar el desarrollo de este tipo de análisis en el sector norte de Península Mitre, en el área de la costa atlántica (Muñoz 1996, 2002, 2004, 2005) y en la costa norte del Estrecho de Magallanes y mares interiores (San Román 2010). Sin embargo, a pesar de estas excepciones, en términos generales, muchos autores sólo se manejan en el nivel de NISP y en pocas oportunidades presentan información correspondiente a MNI, MNE o MAU% (Muñoz 2011). En el caso específico de la región del canal Beagle, en dos sitios se avanzó sobre la descripción de marcas de procesamiento en restos de pinnípedos, ambos conjuntos pertenecientes a la Fase Reciente de ocupación regional (sensu Orquera y Piana 1999): Ajej I, con una cronología de 1400 ± 90 años AP (Piana et al. 2006, Piana et al. 2008) y Túnel VII del S. XIX (Estévez y Martínez 1995).
Las presas, una vez capturadas, son eventualmente transportadas (enteras o en porciones) a las áreas de procesamiento y consumo. Pues bien, en las decisiones generales relacionadas con el transporte intervienen sin lugar a dudas un cúmulo de consideraciones a tener en cuenta (Lupo 2006), entre las que es posible enumerar: el tamaño de la presa (Binford 1978), la distancia entre el lugar de matanza y el campamento residencial (Binford 1978), los costos asociados a la adquisición y procesamiento del recurso (Hawkes y O´Connell 1985) y la tecnología disponible para llevar a cabo dicho transporte (Lupo 2006), entre otros. Asimismo, en el caso específico de Tierra del Fuego, la estrategia de procesamiento y consumo de pinnípedos se puede relacionar tanto con el tipo de aprovisionamiento practicado (explotación de apostaderos vs. explotación de individuos marginales; captura en tierra vs. captura en el agua); y a su vez, con las decisiones sobre el transporte (Borrero y Lanata 1988; Lanata y Winograd 1988; Lanata y Borrero 1994; Orquera y Piana 1999), así como con cuestiones propias de la anatomía de la presa (Lyman 1992; Muñoz 1996, 2005) y nuevamente con la tecnología disponible (Lanata y Winograd 1988; Orquera y Piana 1999), puntos que sin lugar a dudas son interdependientes. Como se señaló previamente, en el ámbito del canal abundan dos especies de otáridos (A. australis y O. flavescens), ambos tienen comportamientos reproductivos similares, pero sus tamaños corporales, abundancias y distribuciones en el ambiente fueguino son diferentes. El lobo fino sudamericano (Arctocephalus australis) alcanza las siguientes dimensiones, según Vaz Ferreira (1979): crías entre 3,5 y 5,5 kg; machos adultos: 1,9 m y 159 kg; hembras adultas: 1,4 m y 48,5 kg; estos datos pertenecen a pinnípedos de las Islas Malvinas. Los cálculos presentados por Schiavini (1990) sobre esta misma especie postulan dimensiones promedio menores: entre 80,9 y 68,2 kg en el caso de los machos adultos y 40,6 kg respecto de las hembras adultas. Los tamaños del lobo común o lobo marino de un pelo (Otaria flavescens) son los siguientes: crías entre 10,2 y 14,8 kg.; machos adultos: 2,56 m y 300- 340 kg.; hembras adultas: 2 m y 144 kg, datos correspondientes a pinnípedos de las costas uruguayas y de las Islas Malvinas (Vaz Ferreira 1979; Schiavini 1990). Respecto de lo antes dicho, cabe señalar que los machos adultos de Otaria flavescens (300 kg) por lo menos duplicarían en tamaño a los de A. australis (160 kg). Este no es un aspecto menor en cuanto a los potenciales aportes individuales de ambas especies a la dieta de los grupos cazadores-recolectores de la región, pero también tiene interesantes implicaciones en relación con la forma en que cada una de ellas pudo ser transportada y procesada. Por lo tanto, sería esperable un procesamiento diferencial entre las especies atendiendo a sus tamaños para su transporte (Binford 1981; Gifford-González 1989; Lyman 1989, 1992, 1994, 2003; Gifford-González y Sunseri 2009), y a su posible abundancia relativa diferencial en las costas interiores del canal Beagle (Sielfeld 1983; Lanata y Winograd 1988; Schiavini 1990; Schiavini y Raya Rey 2001; Crespo et al. 2008a y b).
Proponemos que en el caso de los pinnípedos podrían existir distintas estrategias de transporte de presas considerando la variabilidad de tamaños entre especies, sexos y grupos etarios, donde sería factible esperar lo siguiente: las hembras adultas de O. flavescens, los machos y hembras adultas de A. australis, los juveniles/subadultos de ambos sexos y especies, y los cachorros de ambos sexos y especies podrían ser trasportados enteros desde los sitios de matanza a los espacios residenciales y todas las actividades de desposte se reflejarían en el mismo, mientras que machos adultos de O. flavescens sufrirían un trozamiento inicial en el área de aprovisionamiento, para luego realizar un transporte selectivo de porciones (Binford 1984; Lyman 1992). Para cumplimentar con el objetivo general del trabajo y considerando lo antes dicho: 1) examiné la composición taxonómica y anatómica de los restos de pinnípedos en las capas M, L y K del sitio Imiwaia I (6000-5700 años AP); 2) relevé la influencia de los procesos naturales en la formación del conjunto óseo considerando la relación entre la densidad ósea y las representaciones anatómicas, y cuantifiqué las modificaciones óseas no antrópicas; 3) establecí los perfiles de edad y sexo de los individuos explotados; 4) a través de la descripción de los perfiles anatómicos de los conjuntos (MNE y MAU%) y su covariación con la importancia económica de las distintas partes esqueletarias y con el relevamiento cuantitativo y cualitativo de las modificaciones óseas antrópicas presentes, analicé las posibles pautas de procesamiento y consumo de pinnípedos.
IMIWAIA I: CARACTERÍSTICAS GENERALES
Imiwaia I es un sitio multicomponente en la bahía Cambaceres Interior, Tierra del Fuego, Argentina (54º52´26´´ latitud sur; 67º17´59´´ longitud oeste) (Figura 1). El objetivo de su estudio fue el de cumplimentar una de las etapas planteadas en el marco del Proyecto Arqueológico Canal Beagle, bajo la dirección de L. Orquera y E. Piana, la cual consistía en la comparación entre sitios con cronologías comparables (Segundo Componente de Túnel I) pero ubicados en microambientes con una oferta distinta entre sí de recursos (Orquera y Piana 1999, 2000). La bahía Cambaceres Interior ofrece costas de poco declive, baja presencia de guijarros, con un estrato submareal barroso y relativamente protegido contra los fuertes vientos de la región. Estas características particulares permitieron a los investigadores del proyecto generar un conjunto de expectativas sobre los recursos disponibles en el área y su posible uso (Orquera y Piana 2000: 442).

Figura 1. Mapa con indicación del sitio Imiwaia I (54º 52´26´´ latitud sur; 67º 17´59´´ longitud oeste, en la bahía Cambaceres Interior).
En la Tabla 1 se describe la información cronológica y estratigráfica completa del sitio, que surgió a partir de tres campañas de excavación realizadas en 1998, 1999 y 2002. El total de la superficie excavada en el conchal fue de 50 m² (Orquera y Piana 2000; Zangrando 2009a). En los años 2009 y 2011 continuaron las excavaciones en el sitio aunque por fuera del área de conchales (capa S) (completando unos 74 m²) (Piana et al. 2010; Piana et al. 2012). El sitio Imiwaia I se encuentra conformado por tres estratos de conchales. En este trabajo se presentará el análisis de los conjuntos zooarqueológicos de pinnípedos de los concheros inferiores (capas M, L y K) con una cronología de 6000-5700 años AP. Optamos por presentar la información zooarqueológica de las tres capas en conjunto por dos razones: a) los fechados calibrados a años calendario (Stuiver y Reimer 1993) se superponen parcialmente (Figura 2); y b) se realizaron remontajes entre especímenes provenientes de las distintas capas que conforman los concheros inferiores.
Tabla 1. Información cronológica y estratigráfica del sitio Imiwaia I (los fechados se realizaron sobre carbón) (Orquera y Piana 1999; Zangrando 2009a; Tivoli 2010; Piana et al. 2010).
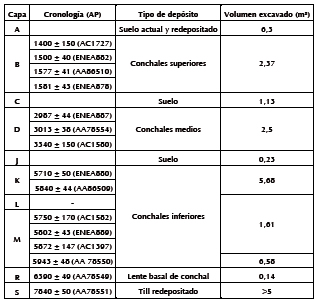

Figura 2. Fechados radiocarbónicos de las capas K y M calibrados AP, CALIB RADIOCARBON CALIBRATION PROGRAM, curva SHcal04, 2σ (Stuiver y Reimer 1993).
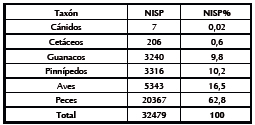
El total de los restos óseos identificados a nivel taxonómico recuperados de las capas M, L y K es de 32479 especímenes (NISP). Las categorías taxonómicas que se han utilizado corresponden en términos generales a cánidos, cetáceos, guanacos, pinnípedos, aves y peces (Tabla 2).
Tabla 2. Composición taxonómica de los concheros inferiores de Imiwaia (Martinoli et al. 2011).
MATERIALES Y MÉTODOS
La identificación taxonómica de los individuos de A. australis y de O. flavescens se realizó utilizando las colecciones de referencia disponibles en el Centro Austral de Investigaciones Científicas, Tierra del Fuego, Argentina (CADIC), y en el Museo Acatushún de Aves y Mamíferos Australes, y empleando criterios morfológicos (King 1954; Sielfeld 1983; Legoupil 1989; Pérez García 2003). Se emplearon los siguientes indicadores morfológicos correspondientes al cráneo (King 1954; Legoupil 1989; Sielfeld 1983; Schiavini 1990): a) la forma del hueso palatino (Legoupil 1989: 103:Figura 3); b) la forma de la arcada dentaria superior (Sielfeld 1983) (Legoupil 1989: 105: Figura 4); c) la forma de la fosa masetera y del proceso coronoide (Legoupil 1989: 105: Figura 4); y d) la forma del canino (Legoupil 1989). Para la identificación taxonómica de huesos del poscráneo se utilizaron los lineamientos metodológicos propuestos por Pérez García (2003), quien describe un conjunto de caracteres discriminantes entre las dos especies de otáridos a través de ejemplares colectados en Uruguay. Tal definición es posible a partir de los huesos de las cinturas de los miembros y de atlas, axis y sacro, específicamente en sectores de articulaciones e inserciones musculares (e.g., Pérez García 2003: 8: Figura 1). Para determinar el sexo de los especímenes, por un lado, se observó la forma de los coxales en su extremo posterior (conformado por el isquion y el pubis), ya sea que estén fusionados o no, pues dicho elemento anatómico permite una determinación inequívoca (King 1983). Y por otro lado; en el caso de A. australis, se midió el ancho de la corona de los caninos; trabajos previos muestran que valores menores/iguales a 8 mm corresponden a hembras, mientras que medidas mayores a 8mm conciernen a machos (Schiavini 1990: 280: Figura 28). De forma subsidiaria, y a modo de complemento, dado el dimorfismo sexual que presentan estas especies, es posible utilizar también la combinación entre estados de fusión (Orquera 2000; Pérez García 2008; Borella et al. 2013) y la longitud de los huesos largos (húmeros, cúbitos, radios, ulnas, fémures y tibias) para determinar el sexo (Orquera 2000).
Las tres categorías de edad utilizadas en este trabajo, definidas a partir del tamaño y del nivel de epifización de los elementos identificados (Pérez García 2008; Borella et al. 2013) son: neonatos (por tamaño y fusión), juveniles/subadultos y adultos (Lyman 1988, Crespo et al. 2008; Borella et al. 2013). Dicha clasificación es la que actualmente, a nuestro entender, describe mejor la relación entre el desarrollo osteológico de los individuos y los cambios en el comportamiento acorde, por un lado, con la finalización del período de amamantamiento, y por otro lado, con el desarrollo de la capacidad reproductiva. A los fines de medir la abundancia relativa de los diferentes taxones identificados, así como de establecer la diversidad de las partes esqueletarias presentes, se han utilizado los indicadores usualmente empleados en los análisis zooarqueológicos: MNI, MNE, MAU y MAU% (Binford 1981; Grayson 1984; Mengoni Goñalons 1988, 1999; Lyman 1994). La frecuencia de partes esqueletarias (expresada como MAU%) obtenidas para los restos de pinnípedos fue correlacionada a través del coeficiente de Spearman rho, tanto con la densitometría ósea de cada unidad anatómica (se promediaron todas las porciones del hueso para obtener un único valor de densidad ósea por elemento) (Borella et al. 2007), como con un índice de utilidad económica (%MUI) calculado por San Román (2009) a partir de un ejemplar de Otaria flavescens. Para analizar la fragmentación de los conjuntos se utilizó la relación NISP/MNE (Lyman 1994; Mengoni Goñalons 1999) y un índice de media ponderada denominado WMI (weighted mean index) (Zohar et al. 2001). Este último se calcula sobre la base de la porción del hueso representada.
Las modificaciones óseas antrópicas relevadas fueron las marcas de corte, machacado o raspado, así como su ubicación sobre los huesos y las termoalteraciones sobre la base de la coloración adquirida por el hueso (Brain 1981; Lyman 1994; Mengoni Goñalons 1999). Con respecto a las modificaciones óseas de origen natural se documentó el grado de meteorización de los especímenes considerando los lineamientos propuestos por Behrensmeyer (1978) sobre la base de distintas etapas o estadios por los que pasa la desintegración de un hueso (cero a cinco). También se relevaron posibles marcas de carnívoros, descriptas en la bibliografía como surcos (scoring), hoyuelos (pitting) y pozos (punctures) (Binford 1981). Para finalizar, se registraron las improntas dejadas por raíces, ya que podrían ser importantes agentes naturales modificadores del registro óseo (Lyman 1994; Marean 1995; Mengoni Goñalons 1999).
RESULTADOS
Abundancia taxonómica y perfil de edad y sexo
En lo que respecta a la composición taxonómica de los concheros inferiores de Imiwaia I (Tabla 3), es notorio el predominio de los restos de Arctocephalus australis (MNI = 32) por sobre los de Otaria flavescens (MNI = 4), en todas las clases de edad y sexo. Con respecto al perfil de mortalidad de A. australis, cabe destacar que es similar la cantidad representada de adultos (MNI = 17) y de juveniles/ subadultos (MNI = 14), determinados a partir del estado de fusión de los huesos largos. Asimismo, se ha podido identificar un sólo individuo correspondiente a la categoría neonato (menos de un año de edad). En lo que refiere a O. flavescens, de los cuatro individuos definidos, tres corresponden a adultos y uno es un subadulto. Las representaciones de las categorías de sexo en el caso de individuos adultos que se han podido describir a través de la forma y fusión de los coxales, y del ancho de la corona de los caninos, en complemento con la medición de los huesos largos (en conjunto con su estado de fusión), muestran que en el caso de A. australis, preponderan claramente los machos (MNI = 14) sobre las hembras (MNI = 3). Para O. flavescens adultos, si bien son muy pocos los individuos identificados, se determinó la presencia de un macho y dos hembras. En la categoría juveniles/subadultos de A. australis se identificaron, por la forma de los coxales, tres machos y una hembra, sin embargo, a diferencia de los adultos, en la mayoría de los individuos (MNI = 10), no se ha podido definir el sexo por tres motivos: a) muchos de los coxales sin fusionar se encuentran fragmentados en la parte diagnóstica; b) no es posible utilizar como guía exclusiva los criterios morfométricos para identificar huesos largos dado el solapamiento de tamaños entre especies y categorías de edad y sexo (Legoupil 1989; Orquera 2000); y c) los caninos que integran la muestra son pocos (MNE = 30), y algunos de ellos se encuentran fragmentados y no es posible medir el ancho de la corona. Para el taxón O. flavescens, el individuo subadulto identificado a partir de un radio, un fragmento de escápula y un fragmento de coxal corresponde a un macho (Orquera 2000).
Tabla 3. Perfil de edad y sexo discriminado por especie (A. australis y O. flavescens) en conjuntos de pinnípedos de los concheros inferiores de Imiwaia I.

Preservación ósea y procesos posdepositacionales
La relación NISP/MNE promediada entre todas las unidades anatómicas de Arctocephalus australis adultos de ambos sexos y de juveniles/subadultos de sexo indeterminado representadas es de 1,2 (Tabla 4), mientras que el WMI es de 77% (Tabla 4). Es decir, ambos índices muestran un alto porcentaje de representación (Zohar et al. 2001); incluso si consideramos por separado los juveniles/subadultos, tal relación es similar (Martinoli 2013). En lo que concierne a los restos identificados como O. flavescens, la relación NISP/MNE es de 1,2, mientras que el WMI es del 88,6%, por lo que también se observa un estado de fragmentación bajo. En lo que refiere a los niveles de meteorización (sensu Behrensmeyer 1978) en las tres capas que conforman los concheros inferiores de Imiwaia I, la mayoría de los restos óseos se encuentran en los estadios 0 y 1 (NISP% = 91,9), mientras que los huesos restantes fueron clasificados dentro del estadio 2 (NISP% = 8,1). Con respecto a las improntas dejadas por la acción de las raíces, menos del 20% de los huesos están afectados. No se registró la acción de carnívoros.
Tabla 4. Identificación de elementos óseos y relación NISP/MNE en A. australis adultos y juveniles/sudadultos.

Representación de partes esqueletarias
La mayoría de las unidades anatómicas que conforman el esqueleto de los Arctocephalus australis machos y hembras adultos y juveniles/subadultos de ambos sexos están representadas en los concheros inferiores de Imiwaia I, sin embargo, no todas en proporciones similares (Tabla 4). En términos generales, la porción que muestra mayores porcentajes de representación es la del esqueleto axial, especialmente las vértebras lumbares (92,4%), las escápulas (81,8%), las esternebras (74,9%) y los coxales (74,5%). Asimismo, los otros huesos que conforman tal porción presentan un MAU% mayor al 50%. En cuanto a los huesos de las extremidades, también es posible observar una alta representación. En el caso de los miembros anteriores, los húmeros, las ulnas y los radios se presentan en 74,5%, 92,7% y 100% respectivamente, mientras que en miembros posteriores el porcentaje de fémures es de 87,3%, el de tibias de 50,9%, y el de peronés, de 70,9%. Las unidades con porcentajes menores de representación son: articulares (carpianos/tarsianos), metacarpianos y metatarsianos. En lo que respecta a Otaria flavescens, se han identificado pocas unidades anatómicas (MNE = 14). Las dos hembras adultas se encuentran representadas a partir de: dos fragmentos de maxilares inferiores, dos fragmentos de escápulas, un húmero, un radio, dos cúbitos y un fémur. El macho adulto y el macho juvenil/ subadulto fueron determinados por la presencia de un coxal y un fragmento de maxilar inferior, en el primer caso, y de un radio, de un fragmento de escápula y de un fragmento de coxal sin fusionar, en el segundo. Pues bien, en el caso de A. australis adultos de ambos sexos y juveniles/subadultos de sexo indeterminado, aunque están representadas la mayoría de las unidades anatómicas, es posible notar diferencias moderadas en sus frecuencias. Si comenzamos evaluando los valores de fragmentación, aunque algunos elementos se ubican en el rango comprendido entre 1-2 (maxilares inferiores, costillas, coxales, húmeros, ulnas, radios, metacarpianos indeterminados, fémures, rótulas, tibias, falanges indeterminadas y vértebras), en tanto específicamente las escápulas presentan un nivel bastante más alto de fragmentación (3,5), la mayoría de las unidades muestran una relación de 1 (Tabla 4), es decir que no están fragmentados. Por lo tanto, a pesar de las diferencias que acabamos de marcar, la fragmentación parece no estar afectando la abundancia en términos de MAU%, ya que los radios con un coeficiente de fragmentación 1,8 son los elementos más representados (MAU% = 100), seguidos por las ulnas con 92,7% (y sólo 1,5 de índice de fragmentación) y las escápulas con 81,2% y 3,5 de fragmentación. Ahora bien, si analizamos la preservación diferencial de partes esqueletarias considerando a la densidad ósea como uno de los posibles factores estructurantes de los conjuntos analizados, debe notarse que la relación testeada no arroja resultados estadísticamente significativos (rs = 0,30 p>0,05). La Figura 3 muestra que la correlación entre las variables se expresa a partir de una dispersión relativamente difusa a lo largo de los dos vectores. La única unidad anatómica que está representada en un bajo porcentaje (MAU% = 21,1) y que posee una densidad ósea baja (0,6) son las vértebras dorsales. Tampoco en el caso de los especímenes que se han podido identificar efectivamente como juveniles/subadultos existe una preservación diferencial considerando la densidad ósea como factor de preservación, ya que el coeficiente de correlación es rs = 0,03 p>0,05. Asimismo, la diferencia en las representaciones esqueletarias en A. australis adultos de ambos sexos y de juveniles/subadultos de sexo indeterminado tampoco puede ser relacionada con un transporte diferencial de porciones anatómicas si se considera su valor económico, ya que el resultado de la correlación arroja un valor estadísticamente no significativo (rs = -0,43 p>0,5). En el caso de todos los restos de Otaria flavescens identificados, los aportes cárnicos de la mayoría de las unidades anatómicas identificadas también tienen valores económicos de medios a bajos (fémur = 6,2, húmero = 20,2%, pelvis = 19,2%, cabeza = 26,7%, radio/ ulna = 15,5%); la excepción son las escápulas, que tienen un valor mayor al 40%.
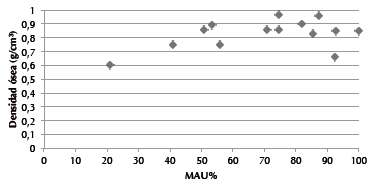
Figura 3. Gráfico de dispersión entre la densidad ósea (g/cm3) y el MAU%.
Procesamiento
Las modificaciones óseas de origen antrópico que se han documentado son las marcas de corte; no se han observado marcas de raspado, machacado o fracturas. Del total de los huesos analizados (3316), el 11,9% de los especímenes se encuentran afectados. Las unidades anatómicas con mayor incidencia de marcas son metacarpianos (12,6%), falanges anteriores/ posteriores (10,9%) y costillas (10,4%) (Tabla 5); y particularmente en los miembros anteriores: húmeros 6,3% (Figura 4) y radios 5,6% (Tabla 5). En el caso específico de los restos de Otaria flavescens (MNE = 14), no se observaron marcas antrópicas de ningún tipo. Con respecto a la ubicación de las marcas de corte en los huesos, el conjunto muestra el mayor porcentaje de trazas en zonas proximales y distales sobre las diáfisis (Figura 5), lo que podría ser compatible con actividades de desarticulación (Binford 1981; Lyman 1992); sin embargo, las epífisis prácticamente no muestran trazas. Las escápulas muestran una alta incidencia de marcas de corte en la porción proximal de la hoja (60%), los húmeros sobre la diáfisis distal (32,5%), y los radios y ulnas sobre las diáfisis proximales (57,1% y 44,4% respectivamente). En los especímenes correspondientes a los miembros posteriores: a) los fémures se ven más afectados sobre las regiones distales de las diáfisis (41,7%); b) las tibias, en una proporción similar sobre todas las porciones de las diáfisis (28,6%); y c) los peronés, sobre la diáfisis en su sector medial (50%). Las falanges anteriores y posteriores muestran concentración de trazas sobre las porciones medias de la diáfisis (38,1%), en tanto metacarpianos y metatarsianos se ven más afectados sobre la región proximal (50,9% y 50% respectivamente). En las vértebras, la mayoría de las trazas de corte se aglutinan sobre las apófisis (más del 50% en todos los casos), mientras que, en las costillas, sobre la porción medial del cuerpo (40,5%). Por último, es necesario destacar el bajo porcentaje de especímenes en los cuales se observa alteración térmica. De los 3316 especímenes, sólo 41 (1,2%) muestran rastros de termoalteración (4 fragmentos carbonizados y 37 fragmentos quemados sensu Lyman 1994).
Tabla 5. Cuantificación de marcas de corte por unidad anatómica en A. australis adultos y juveniles/subadultos.

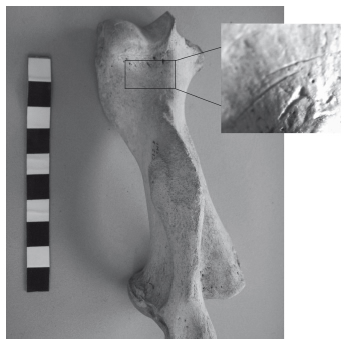
Figura 4. Húmero de hembra adulta de A. australis con detalle de marcas de corte (lupa binocular x 0.63) sobre porción distal de la diáfisis.
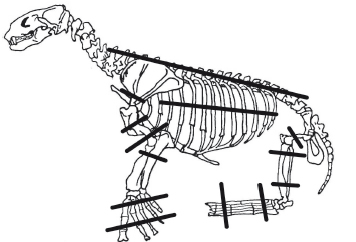
Figura 5. Esqueleto de pinnípedo con indicación de porciones con mayor porcentaje de marcas de corte (Arctocephalus australis adultos y juveniles/ subadultos de sexo indeterminado).
DISCUSIÓN
En cuanto a la preservación de los restos óseos, es posible sostener que Imiwaia I presenta niveles generales buenos. Por un lado, la fragmentación expresada a través de la relación NISP/MNE es cercana a 1 en la mayoría de los casos, mientras que el WMI es mayor al 70%. Por otro lado, los conjuntos muestran poca perturbación por acción de agentes como raíces y no se observaron marcas de carnívoros. Sin embargo, si evaluamos la preservación a través de los niveles de meteorización (sensu Behrensmeyer 1978), es necesario remarcar que el 8% del NISP total de la muestra se encuentra en el estadio 2 de meteorización, lo que señala un grado significativo de exposición aérea de los conjuntos óseos; sus implicaciones serán analizadas con mayor detalle en el futuro a partir de estudios actualísticos que se encuentran en curso para la región. Ahora bien, con respecto a la preservación diferencial de partes esqueletarias, si consideramos a la densidad ósea como un factor mediador en la preservación de los conjuntos analizados, debe notarse que la relación testeada no arrojó resultados estadísticamente significativos. Un punto de suma importancia para comenzar a describir las posibles modalidades de explotación de pinnípedos fue la determinación de los perfiles etarios y de sexo. En el conjunto se observa un predominio de adultos machos de Arctocephalus australis, una baja proporción de hembras y la presencia de un único individuo neonato (menor a un año). Esta composición no es compatible con la explotación en loberías de reproducción. Como se dijo previamente, a partir del análisis realizado en el Segundo Componente de Túnel I (Schiavini 1990, 1993; Orquera y Piana 1999) se sostiene que el predominio de machos adultos y subadultos puede responder a la captura en el agua de individuos aislados. En el caso de los concheros inferiores de Imiwaia I, no se ha podido definir el sexo en muchos de los individuos juveniles/subadultos, lo cual dificulta la interpretación. En este sentido, puede considerarse que las presas podrían haber sido obtenidas también en apostadores de descanso y luego ser transportadas a las áreas residenciales (Lanata y Winograd 1988; Moreno 2008).
En el caso de Otaria flavescens -si bien son muy pocos los restos que se han podido asignar a esta especie- es al menos interesante remarcar la presencia de dos hembras adultas, considerando que esta especie, en la actualidad, sí utiliza islas o islotes ubicados en el canal Beagle para el establecimiento de sus apostaderos reproductivos (Schiavini 1993). Sin lugar a dudas, en este punto sería necesario plantear nuevas hipótesis e incorporar más datos para ajustar las interpretaciones. El segundo grupo de implicancias relacionadas directamente con la estrategia de captura es el que hace a las representaciones esqueletarias. En lo que respecta únicamente a los restos de Arctocephalus australis adultos de ambos sexos y de juveniles/subadultos de sexo indeterminado, en el conjunto analizado es posible observar que la mayoría de las unidades anatómicas están representadas aunque no en la misma proporción. Cabe aclarar que en el caso particular de carpianos y tarsianos, la baja representación podría responder a una preservación o pérdida diferencial, considerando que son huesos pequeños, y particularmente que son cartilaginosos en juveniles y subadultos (Legoupil 1989), los cuales representan una gran parte del conjunto estudiado. En cuanto a los restos de O. flavescens, son muy pocos los elementos identificados. Ahora bien, las diferencias en las representaciones esqueletarias en ninguna de las dos especies pueden ser relacionadas con un transporte diferencial de porciones anatómicas considerando como variable principal su valor económico, ya que no se hallaron relaciones estadísticamente significativas con ese factor. En resumen, los perfiles anatómicos registrados para A. australis adultos de ambos sexos y juveniles/subadultos de sexo indeterminado -al estar representadas todas las unidades esqueletarias- no permiten defender una estrategia que involucre el procesamiento inicial de las carcasas fuera del sitio; por el contrario, parece que estas habrían ingresado enteras (o, al menos, que en diferentes episodios se introducían indistintamente porciones anatómicas diversas de los individuos sucesivamente capturados). Esta interpretación se complementa bien con argumentos previos (Schiavini 1990; Orquera y Piana 1999) que señalan que la captura de esta especie habría ocurrido mayormente sobre individuos aislados en el agua, empleando arpones de punta desmontable, y posteriormente el transporte utilizando medios de navegación. No obstante, no debe dejar de evaluarse la posibilidad de la captura en tierra de individuos marginales (desagregados), o la explotación de grupos pequeños apostados de modo semipermanente u ocasional en sectores de islas e islotes cercanos a los sitios residenciales costeros como Imiwaia I, considerando que el costo de transporte sería relativamente bajo por la disponibilidad de tecnología de navegación (Binford 1978). Los individuos de O. flavescens están representados por muy pocas unidades anatómicas, por lo que resulta más difícil hacer una evaluación sobre su transporte al sitio.
En lo que concierne específicamente al procesamiento de pinnípedos, los restos óseos de A. australis hembras/machos adultos y de juveniles/subadultos de sexo indeterminado que conforman los concheros inferiores de Imiwaia I muestran incidencia de marcas de cortes proporcionalmente semejantes entre sí, mientras que en los restos de O. flavescens no se relevaron marcas de origen antrópico. La distribución de las trazas de corte sobre los huesos que corresponden a los primeros es más o menos homogénea entre todas las porciones. Sin embargo, las regiones que muestran proporciones más altas de esos rastros son relacionables con actividades de desarticulación. En los restos que corresponden a los miembros anteriores es posible observar marcas que indicarían la separación en tres partes: en primer lugar, la porción que corresponde a la escápula (marcas sobre la cara proximal); en segundo lugar, la porción superior del miembro anterior (marcas sobre la zona distal del húmero y sobre las regiones proximales y mediales de los radios y las ulnas, respectivamente); y por último, el seccionamiento de la aleta (marcas sobre la porción proximal de metacarpianos). Esto último posiblemente indique la conservación/consumo del grueso panículo adiposo ubicado en ese sector -más del 50% del tejido que conforma dicha porción es grasa (Martinoli et al. 2014). No obstante, también podrían ser marcas compatibles con actividades de cuereo, según información de actividades de loberos comerciales del siglo XIX (Cartajena y Labarca 2007). En cambio, los miembros posteriores presentan marcas compatibles con el fraccionamiento en dos secciones; los fémures concentran una mayor cantidad de marcas sobre las porciones distales, mientras que las tibias y peronés sobre las regiones proximales de la diáfisis; también los huesos de las aletas, particularmente los metatarsianos, se ven más afectados sobre sectores proximales.
En el caso del esqueleto axial, la columna vertebral muestra una gran concentración de marcas de corte en las apófisis, lo cual podría indicar actividades de desarticulación. En cuanto a las costillas, se observa la mayor incidencia de trazas sobre sectores mediales. Estas huellas podrían estar relacionadas con la extracción de carne, también considerando como referencia la actividad lobera del siglo XIX (Cartajena y Labarca 2007). Por lo tanto, probablemente la actividad principal de trozamiento fue la desarticulación: en primer término, la separación de miembros anteriores y posteriores del esqueleto axial y a su turno la reducción de los primeros a porciones más pequeñas (marcas sobre zonas articulares de húmeros, radio, ulnas, fémures, tibias y peronés). A su vez, el procesamiento de aletas para el consumo del panículo adiposo podría haber sido una práctica recurrente, ya que metacarpianos y metatarsianos muestran trazas sobre zonas donde la tarea extractiva de la grasa y carne adherida es más dificultosa (Martinoli et al. 2014). Por último, como se dijo previamente, es muy bajo el porcentaje de restos óseos con evidencias de termoalteración, lo cual debería ser evaluado en trabajos futuros que indaguen directamente sobre las posibles actividades de cocción. Las evidencias enumeradas con respecto al perfil anatómico y las marcas de corte serían entonces posibles indicadores de que todas las actividades de procesamiento y consumo se llevaron a cabo en el área residencial (Binford 1980). Si introducimos una breve comparación con sitios pertenecientes a la fase tardía de ocupación de la región del canal Beagle pareciera no haber cambios en el procesamiento de pinnípedos. En el sitio Ajej I, los autores observan posibles pautas de desmembramiento de los miembros anteriores y posteriores en porciones más pequeñas y la conservación para el consumo de los panículos adiposos que se ubican en las aletas (Piana et al. 2006; Piana et al. 2008). Asimismo, la columna vertebral muestra marcas de corte en las regiones ventrales y sólo algunas esternebras están termoalteradas (Piana et al. 2006; Piana et al. 2008). En el caso de Túnel VII, es posible observar la representación de la mayoría de las unidades anatómicas, por lo que los autores suponen el ingreso de las presas enteras al sitio (Estévez y Martínez 1995).
Las marcas de corte documentadas (estrías longitudinales) son compatibles con prácticas de desmembramiento. Para este sitio los autores proponen un proceso continuo de trabajo desde el descuartizamiento, pasando por la desarticulación hasta llegar al descarnado (Estévez y Martínez 1995). En los últimos años se ha desarrollado un modelo (Zangrando 2009a y b; Tivoli 2010; Tivoli y Zangrando 2011) que da cuenta de los cambios en las representaciones faunísticas a lo largo de toda la secuencia de ocupación de la región del canal Beagle, las cuales muestran una tendencia general hacia la disminución en la proporción de consumo de pinnípedos y un aumento relativo en las representaciones de peces y aves -especialmente en aquellas especies de peces de aguas profundas- Sin embargo, más allá de las posibles diferencias en el consumo de pinnípedos entre conjuntos antiguos (concheros inferiores de Imiwaia I) y recientes (Ajej I y Túnel VII) no parece haber cambios en las pautas de procesamiento. Debemos resaltar, no obstante, que lo antes dicho es una evaluación preliminar que debería ser contrastada sumando evidencia correspondiente a más sitios.
CONCLUSIONES
En este trabajo se evaluaron las pautas de captura, procesamiento y consumo de pinnípedos en los conjuntos canoeros tempranos del sitio Imiwaia I, considerando el modo en que las decisiones sobre la explotación de tales presas pueden ser relacionadas con aspectos tales como su biología, su comportamiento y su disponibilidad en el ambiente (Lanata y Winograd 1988; Lyman 1989; Hildebrant y Jones 1992; Jones y Hildebrant 1995; Colten y Arnold 1998; Orquera y Piana 1999). Concretamente, si evaluamos los perfiles de edad y sexo relevados es posible observar que en el caso de A. australis efectivamente hubo poco consumo de hembras adultas y neonatos, en tanto predominan notoriamente los restos de machos adultos, lo cual no sería compatible con la explotación de loberías reproductivas. No obstante, no fue posible determinar el sexo en la mayoría de los individuos juveniles/ subadultos. En lo que respecta a O. flavescens, la identificación de sólo 14 elementos no permite realizar a una interpretación adecuada de los datos.
El otro factor que se ha considerado fundamental para comprender las pautas de procesamiento y consumo ha sido el del transporte de porciones esqueletarias. En el caso de los restos de A. australis adultos de ambos sexos y de juveniles/subadultos de sexo indeterminado, hemos observado la representación de la mayoría de las porciones anatómicas; mientras que en el caso de los huesos de O. flavescens hay una presencia parcial del esqueleto. Posiblemente hayan existido variaciones en el procesamiento de las carcasas de tales presas, considerando el ingreso diferencial de partes anatómicas al sitio. Incluso, en trabajos futuros debería ser evaluada la posibilidad de un aprovechamiento oportunístico de los machos adultos de O. flavescens. No obstante, al ser tan pocos los huesos pertenecientes a tal categoría las conclusiones son sumamente preliminares, y podrían responder a un problema exclusivamente estadístico derivado del tamaño de la muestra de huesos de O. flavescens en contraposición a los restos óseos de A. australis. Por último, las modificaciones óseas de origen antrópico relevadas en los restos óseos de A. australis de ambos sexos y de juveniles/subadultos de sexo indeterminado se corresponden con actividades de desarticulación, y, en menor medida, de cuereo y extracción de carne, todas ellas probablemente realizadas en el sitio, lo cual es compatible con la captura de las presas en el agua o de presas disponibles en áreas de costa relativamente cercanas al sitio y el posterior transporte de las carcasas enteras a las áreas residenciales.
A través del estudio de los conjuntos óseos recuperados en las capas M, L y K (6000-5700 años AP) hemos descripto la forma en que los pinnípedos pueden haber sido capturados, procesados y consumidos en los momentos tempranos de las ocupaciones canoeras. En trabajos futuros sería necesario considerar dos aspectos importantes: a) el estudio de las pautas de procesamiento y consumo de pinnípedos en otros conjuntos canoeros tempranos de la región para generar datos que correspondan a diversos contextos (e.g., sitios en otros microambientes); y b) conocer las pautas de procesamiento y consumo de pinnípedos en términos diacrónicos, teniendo en cuenta los cambios regionales propuestos en trabajos antecedentes; y c) integrar estos estudios sobre pinnípedos con otros análisis que evalúen los distintos componentes de la alimentación humana canoera.
Agradecimientos
Este trabajo surge de la Tesis de Licenciatura "Modalidades de explotación de pinnípedos en las ocupaciones canoeras tempranas de la región del canal Beagle: el sitio Imiwaia I (Tierra del Fuego, Argentina)", Facultad de Filosofía y Letras, Universidad de Buenos Aires (marzo de 2013), en el marco de los proyectos PICT 2071/2006 (MINCYT-ANPCYTN) y PIP 0395/10 (CONICET). Quiero agradecer a Francisco Zangrando y a Luis Orquera por los aportes realizados sobre las distintas versiones de este trabajo. A Angélica Tívoli y Vanesa Parmigiani por la colaboración en el montado de las figuras. A Lucas Bustos por facilitarme material bibliográfico. A Natalie Goodall por permitirme acceder al material osteológico de referencia del Museo Acatushún de Aves y Mamíferos Marinos Australes. A los evaluadores por sus enriquecedoras sugerencias.
REFERENCIAS CITADAS
1. Behrensmeyer, A. K. 1978 Taphonomic and Ecologic Information from Bone Weathering. Paleobiology 5: 150-162. [ Links ]
2. Binford, L. 1978 Nunamiut ethnoarchaeology. Academic Press, Nueva York. [ Links ]
3. Binford, L. 1981 Bones: Ancient Men and Modern Myths. Academic Press, Nueva York. [ Links ]
4. Binford, L. 1984 Faunal Remains from Klasies River Mouth. Academic Press, Nueva York. [ Links ]
5. Borella, F., M. Gutiérrez, H. Fodere y J. Merlo 2007 Estudio de densidad mineral ósea para dos especies de otáridos frecuentes en el registro arqueofaunístico patagónico (Otaria flavescens y Arctocephalus australis). En Arqueología de Fuego- Patagonia. Levantando piedras, Desenterrando huesos... y Develando Arcanos, editado por F. Morello, A. Prieto, M. Martinic y G. Bahamonde, pp. 421-426. Centro de Estudios del Cuaternario de Fuego Patagonia y Antártica (CEQUA), Punta Arenas. [ Links ]
6. Borella, F., F. Grandi, D. Vales, N. Goodall y E. Crespo 2013 Esquema preliminar de fusión epifisaria en huesos de lobos marinos (Arctocephalus australis y Otaria flavescens), su contribución en los análisis zooarqueológicos. En Tendencias teórico-metodológicas y casos de estudio en la arqueología de la Patagonia, editado por A. Zangrando, R. Barberena, A. Gil, G. Neme, M. Giardina, L. Luna, C. Otaola, S. Paulides, L. Salgán y A. Tivoli, pp. 39-51. Sociedad Argentina de Antropología (SAA) e Instituto Nacional de Antropología y Pensamiento Latinoamericano (INAPL), Buenos Aires. [ Links ]
7. Borrero, L. y J. Lanata 1988 Estrategias adaptativas representadas en los sitios de Estancia María Luisa y Cabo San Pablo. Precirculados IX Congreso Nacional de Arqueología: 166-174. ICA. [ Links ]
8. Bowen, W., C. Beck y D. Austin 2009 Pinniped Ecology. En Encyclopedia of marine mammals, 2da. ed., editado por W. Perrin, B. Würsig y J. Thewissen, pp. 852-861. Academic Press, Nueva York. [ Links ]
9. Brain, C. K. 1981 The hunters or the hunted? An introduction to African cave taphonomy. Chicago, Chicago University Press. [ Links ]
10. Campagna, C. 1985 The breeding cycle of the southern sea lion, Otaria byronia. Marine Mammal Science 1 (3): 210-218. [ Links ]
11. Cartajena, I. F. y R. Labarca 2007 Patrones históricos de cuereo en lobos marinos: La Evidencia de los Abrigos 1 y 2 de Playa Yámana (Isla Livingston, Antártica Chilena). Magallania 35 (1): 95-105. [ Links ]
12. Colten, R. H. y J. E. Arnold 1998 Prehistoric Marine Mammal Hunting on California´s Northern Channel Island. American Antiquity 63: 679-701. [ Links ]
13. Crespo E., N. García, S. Dans y S. Pedraza 2008a Arctocephalus australis. Atlas de Sensibilidad Ambiental de la Costa y el Mar Argentino. Mamíferos Marinos. http://atlas.ambiente.gov.ar (marzo 2013). [ Links ]
14. Crespo E., N. García, S. Dans, S. Pedraza, M. Lewis y C. Campagna 2008b Otaria flavescens. Atlas de Sensibilidad Ambiental de la Costa y el Mar Argentino. Mamíferos Marinos. http://atlas.ambiente.gov.ar (marzo 2013). [ Links ]
15. Estévez, J. y J. Martínez 1995 Reflexiones metodológicas a partir del análisis de los restos de pinnípedos de Túnel VII. En Encuentros en los conchales fueguinos, vol. 1, editado por J. Estévez Escalera y A. Vila Mitjà, pp. 183-193. Universidad Autónoma de Barcelona, Barcelona. [ Links ]
16. Gifford-González, D. 1989 Ethnographic analogues for interpreting modified bones: some cases from East Africa. En Bone Modification, editado por R. Bonnichsen y M. H. Sorg, pp. 179-246. Centre for the Study of the First Americans, Maine. [ Links ]
17. Gifford-Gonzalez, D. y C. K. Sunseri 2009 An Earlier Extirpation of Fur Seals in the Monterey Bay Region: Recent Findings and Social Implications. Proceedings of the Society for California Archaeology 21: 89-102. [ Links ]
18. Grayson, D. 1984 Quantitative zooarchaeology. Academic Press, Nueva York. [ Links ]
19. Hawkes, K. y O'Connell, J. 1985 Optimal foraging models and the case of the !Kung. American Anthropologist 87: 401-404. [ Links ]
20. Hildebrandt, W. R. y T. L. Jones 1992 Evolution of Marine Mammal Hunting: A view from the California and Oregon Coast. Journal Anthropological Archaeology 11 (4): 360-401. [ Links ]
21. Jones, T. L y W. R. Hildebrandt 1995 Reasserting a Prehistoric Tragedy of the Commons: Reply to Lyman. Journal of Anthropological Archaeology 14 (1): 78-98. [ Links ]
22. King, J. 1954 The Otariid seals of the Pacific Coast of America. Bulletin of the British Museum (Natural History) 2 (10): 311-337. [ Links ]
23. King, J. 1983 Seals of the world. Oxford University Press, Londres. [ Links ]
24. Lanata, J. y L. Borrero 1994 Riesgo y Arqueología. Arqueología de Cazadores- Recolectores, límites, casos y apertura, Arqueología Contemporánea 5: 129-143. [ Links ]
25. Lanata, J. y A. Winograd 1988 Gritos y Susurros, Aborígenes y Lobos Marinos en el litoral de la Tierra del Fuego. En Arqueología de las Américas. 45 Congreso Internacional de Americanistas: 227-246. Fondo Promoción de la Cultura, Bogotá [ Links ].
26. Legoupil, D. 1989 La identificación de los mamíferos marinos en los sitios canoeros de Patagonia: problema y constataciones. Anales del Instituto de la Patagonia 19: 101-113. [ Links ]
27. Lupo, K. 2006 What Explains the Carcass Field Processing and Transport Decisions of Contemporary Hunter-Gatherers? Measures of Economic Anatomy and Zooarchaeological Skeletal Part Representation. Journal of Archaeological Method and Theory 13 (1): 19-66. [ Links ]
28. Lyman, R. L. 1988 Zoogeography of Oregon coast marine mammals: the last 3000 years. Marine Mammals Science 4 (3): 247-264. [ Links ]
29. Lyman, R. L. 1989 Seal and Sea-lion Hunting: A Zooarchaeological Study from the Southern Northwest Coast of North America. Journal of Anthropological Archaeology 8: 68-99. [ Links ]
30. Lyman, R. L. 1992 Prehistoric Seal and Sea-lion Butchering on the Southern Northwest Coast. American Antiquity 57 (2): 246-261. [ Links ]
31. Lyman, R. L. 1994 Vertebrate Taphonomy. Cambridge University Press, Cambridge. [ Links ]
32. Lyman, R. L. 1995 On the Evolution of Marine Mammal Hunting on the West Coast of North America. Journal of Anthropological Archaeology 14: 45-77. [ Links ]
33. Lyman, R. L. 2003 Pinniped Behavior, Foraging Theory, and the Depression of Metapopulation and Nondepression of a Local Population on the Southern Northwest Coast of North America. Journal of Anthropological Archaeology 22 (4): 376-388. [ Links ]
34. Marean, C. 1995 Of taphonomy and zooarchaeology. Evolutionary Anthropology 4: 64-72. [ Links ]
35. Martinoli, M. P. 2013 Modalidades de explotación de pinnípedos en las ocupaciones canoeras tempranas del canal Beagle: el sitio Imiwaia I (Tierra del Fuego, Argentina). Tesis de Licenciatura inédita. Facultad de Filosofía y Letras, Universidad de Buenos Aires, Buenos Aires. [ Links ]
36. Martinoli, M. P., A. Tívoli, D. Alunni y A. F. Zangrando 2014 Anatomía económica de A. australis. En preparación. [ Links ]
37. Martinoli, M. P., A. Tivoli, E. Piana y L. Orquera 2011 Conjuntos zooarqueológicos del sitio Imiwaia I (Tierra del Fuego): una visión global sobre las representaciones taxonómicas. Libro de resúmenes de II Congreso Nacional de Zooarqueología Argentina: 57. Olavarría. [ Links ]
38. Mengoni Goñalons, G. 1988 Análisis de materiales faunísticos de sitios arqueológicos. Xama 1: 71-120. [ Links ]
39. Mengoni Goñalons, G. 1999. Cazadores de guanacos de la estepa patagónica. SAA, Buenos Aires. [ Links ]
40. Moreno, J. E. 2008 Arqueología y etnohistoria de la Costa Patagónica Central en el Holoceno Tardío. Rawson, Fondo Editorial Provincial, Secretaría de Cultura del Chubut. [ Links ]
41. Muñoz, S. 1996 Explotación de pinnípedos en la costa Atlántica de Tierra del Fuego. Arqueología 6: 199-222. [ Links ]
42. Muñoz, S. 2002 La explotación de mamíferos por Cazadores- Recolectores Terrestres de Tierra del Fuego. Tesis Doctoral inédita. Facultad de Filosofía y Letras, Universidad de Buenos Aires, Buenos Aires. [ Links ]
43. Muñoz, S. 2004 La explotación de pinnípedos en el sitio arqueológico Punta María 2, Isla Grande de Tierra del Fuego. En Contra viento y marea. Arqueología de la Patagonia, editado por T. Civalero, P. Fernández y G. Guráieb, pp. 489-496. SAA, Buenos Aires. [ Links ]
44. Muñoz, S. 2005 Zooarqueología del Sector Atlántico de la Isla Grande de Tierra del Fuego. La utilización de camélidos y pinnípedos por los cazadores-recolectores fueguinos. Relaciones de la Sociedad Argentina de Antropología XXX: 59-77. [ Links ]
45. Muñoz, S. 2011 Pinniped Zooarchaeological Studies in Southern Patagonia: Current Issues and Future Research Agenda. En Trekking the Shore. Changing Coastlines and the Antiquity of Coastal Settlement, editado por N. Bicho, J. Haws y L. Davis, pp. 305-331. Springer, Nueva York. [ Links ]
46. Orquera, L. A. 2000 Instrucciones para el procesamiento de restos óseos de pinnípedos. MS. [ Links ]
47. Orquera, L. A. 2011 Comunicación en el Primer Taller Internacional "Pinnípedos del Neotrópico y Antártida: zooarqueología, historia y biología". Puerto Santa Cruz. MS. [ Links ]
48. Orquera L. A. y E. L. Piana 1999 Arqueología de la región del canal Beagle (Tierra del Fuego, Argentina). SAA, Buenos Aires. [ Links ]
49. Orquera L. A. y E. L. Piana 2000 Imiwaia I: un sitio de canoeros del sexto milenio AP en la costa norte del canal Beagle. En Desde el País de los Gigantes. Perspectivas arqueológicas en Patagonia, t. II, editado por J. Belardi, F. Carballo y S. Espinosa, pp. 441-453. Universidad Nacional de la Patagonia Austral, Río Gallegos. [ Links ]
50. Orquera L. A. y E. L. Piana 2005 Mid-Holocene littoral adaptation at the southern end of South America. Quaternary International 132: 107-115. [ Links ]
51. Orquera L. A. y E. L. Piana 2009 Sea Nomads of the Beagle Channel in Southernmost South America: Over Six Thousand Years of Coastal Adaptation and Stability. Journal of Island & Coastal Archaeology 4: 61-81. [ Links ]
52. Pérez García, M. I. 2003 Osteología comparada del esqueleto postcraneano de dos géneros de otariidae del Uruguay. Boletín de la Sociedad Zoológica del Uruguay 14: 1-16. [ Links ]
53. Pérez García, M. I. 2008 Ontogenia del postcráneo de Arctocephalus australis (mammalia, carnivore, otariidae). Boletín de la Sociedad Zoológica del Uruguay 17: 1-19. [ Links ]
54. Piana, E. L., M. Vázquez y M. Álvarez 2008 Nuevos resultados del estudio del sitio Ajej I: un aporte a la variabilidad de estrategias de los canoeros fueguinos. Runa 29: 101-121. [ Links ]
55. Piana, E. L., M. Vázquez, M. Álvarez y N. Rua 2006 El sitio Ajej I: excavación de rescate en la costa del canal Beagle. En Arqueología Argentina en los inicios de un nuevo siglo, t. I, editado por F. Oliva, N. de Grandis y J. Rodríguez, pp. 345-356. Universidad Nacional de Rosario, Rosario. [ Links ]
56. Piana, E., F. Zangrando y L. Orquera 2010 La evidencia arqueológica de la capa S de Imiwaia I (Canal Beagle, Tierra del Fuego): una ocupación del holoceno temprano en el extremo austral del continente Americano. V Simposio Internacional del Hombre Temprano en América. La Plata. [ Links ]
57. Piana, E., F. Zangrando y L. Orquera 2012 Early occupations in Tierra del Fuego and the evidences from S layer in Imiwaia I site (Beagle Channel, Argentina). Current Research in the Pleistocene, Center for the Study of First Americans, en prensa. [ Links ]
58. San Román, M. 2009 Anatomía económica de Otaria flavescens. En Zooarqueología en el confín del mundo, editado por P. López, I. Cartajena, C. García y F. Mena, pp. 169-180. Facultad de Estudios del Patrimonio Cultural de la Universidad Internacional SEK-Chile, Área de Arqueología, Santiago de Chile. [ Links ]
59. San Román, M. 2010 La explotación de recursos faunísticos en el sitio Punta Santa Ana 1: estrategias de subsistencia de grupos cazadores marinos tempranos de Patagonia meridional. Magallania 38 (1): 183-198. [ Links ]
60. Schiavini, A. 1990 Estudio de la relación entre el hombre y los pinnípedos en el proceso adaptativo humano del canal Beagle (Tierra del Fuego, Argentina). Tesis Doctoral inédita. Facultad de Ciencias Exactas y Naturales, Universidad de Buenos Aires, Buenos Aires. [ Links ]
61. Schiavini, A. 1993 Los lobos marinos como recursos para cazadoresrecolectores marinos: el caso de Tierra del Fuego. Latin American Antiquity 4: 346-366. [ Links ]
62. Schiavini, A. y A. Raya Rey 2001 Aves y Mamíferos Marinos en Tierra del Fuego. Estado de situación, interacción con actividades humanas y recomendaciones para su manejo. Informe preparado bajo contrato con el proyecto "Consolidación e Implementación del Plan de Manejo de la Zona costera Patagónica", proyecto ARG/97/G31 GEF/PNUD/MRECIC CADIC-CONICET. [ Links ] Sielfeld, W.
63. Sielfeld, W. 1983 Mamíferos Marinos de Chile. Ediciones de la Universidad de Chile, Santiago. [ Links ]
64. Sielfeld, W. 1999 Estado del conocimiento sobre conservación y preservación de Otaria flavescens (Shaw, 1800) y Arctocephalus australis (Zimmermann, 1783) en las costas de Chile. Estudios Oceanológicos 18: 81-96.
65. Stuiver, M. y P. Reimer 1993 Calib Radiocarbon Calibration Program. Radiocarbon 35: 215-230. [ Links ]
66. Tivoli, A. 2010 Las aves en la organización socioeconómica de cazadores-recolectores-pescadores del extremo sur sudamericano. Tesis Doctoral inédita. Facultad de Filosofía y Letras, Universidad de Buenos Aires, Buenos Aires. [ Links ]
67. Tivoli, A. y F. Zangrando 2011 Subsistence variations and landscape use among maritime hunter-gatherers. A zooarchaeological analysis from the Beagle Channel (Tierra del Fuego, Argentina). Journal of Archaeological Science 38 (5): 1148-1156. [ Links ]
68. Vaz Ferreira, R. 1979 South American sea lion. FAO Fisheries Series 5 (2): 9-12. [ Links ]
69. Vaz Ferreira, R. y S. Vallejo 1981 Algunos aspectos del comportamiento de Arctocephalus australis (Zimmerman) lobo de dos pelos de Sudamérica (pinnipedia-otariidae) en el Uruguay. En VI Jornadas Argentinas de Zoología, Simposios: 223- 236. La Plata. [ Links ]
70. Vidal H. J. y A. Winograd 1986 El factor humano. Modificaciones antropogénicas en la paleoecología de otáridos en Tierra del Fuego. En Anais da 2a Reuniâo de trabalho de especialistas em mamíferos aquáticos da América do Sul. Río de Janeiro. [ Links ]
71. Zangrando, A. F. 2003 Ictioarqueología del canal Beagle. Explotación de peces y su implicación en la subsistencia humana. SAA, Buenos Aires. [ Links ]
72. Zangrando, A. F. 2009a Historia evolutiva y subsistencia de cazadoresrecolectores marítimos de Tierra del Fuego. SAA, Buenos Aires. [ Links ]
73. Zangrando, A. F. 2009b Is fishing intensification a direct route to huntergatherer complexity? A case study from the Beagle Channel region (Tierra del Fuego, southern South America). World Archaeology 41 (4): 589-608. [ Links ]
74. Zohar, I., T. Dayan, E. Galili, y E. Spanier 2001 Fish processing during the early Holocene: A taphonomic study. Journal of Archaeological Science 28: 1041-1053. [ Links ]
