










Serviços Personalizados
Journal

Artigo
 Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares em
SciELO
Similares em
SciELO
Compartilhar

 Permalink
PermalinkIntersecciones en antropología
versão On-line ISSN 1850-373X
Intersecciones antropol. vol.17 no.3 Olavarría dez. 2016
ARTÍCULOS
Isótopos estables en camélidos y vegetales modernos de Antofagasta de la Sierra: hacia una ecología isotópica de la Puna Meridional argentina
Jennifer Grant
Consejo Nacional de Investigaciones Científicas y Técnicas (CONICET), Instituto Nacional de Antropología y Pensamiento Latinoamericano (INAPL). 3 de Febrero 1378 (C1426BJN), Ciudad Autónoma de Buenos Aires, Argentina. E-mail: jennygrantlett@gmail.com
Recibido 2 de octubre 2015.
Aceptado 28 de diciembre 2015
RESUMEN
En este trabajo se dan a conocer los primeros resultados obtenidos en el marco de un proyecto de investigación orientado a construir una ecología isotópica de la Puna Meridional argentina, susceptible de ser empleada como marco de referencia para la interpretación del registro arqueológico. Se presentan 34 pares de valores de ä13C y ä15N medidos sobre colágeno óseo de camélidos modernos (Vicugna vicugna y Lama glama) y 28 pares de valores de ä13C y ä15N medidos sobre las pasturas consumidas por dichos animales en diferentes unidades vegetales de Antofagasta de la Sierra (Catamarca, Argentina). Los resultados obtenidos en especies vegetales demuestran la existencia de ambientes con diferentes marcas isotópicas, que dependen principalmente de las variaciones altitudinales presentes en el área de estudio. Estos contrastes entre vegetación ubicada en diferentes cotas altitudinales se ven reflejados asimismo en diferencias en los valores de ä13C y ä15N en camélidos, principalmente silvestres. En el caso de los animales domésticos, la práctica moderna de complementar la dieta de los rebaños con alfalfa (Medicago sativa), una planta C3, resultó en valores más negativos que los esperados.
Palabras clave: Ecología isotópica; Isótopos estables de carbono y nitrógeno; Camélidos; Plantas; Puna Meridional Argentina.
ABSTRACT
Stable isotopes in camelids and modern plants from Antofagasta de la Sierra: towards an isotopic ecology of the southern Argentine Puna
This article presents the first results obtained within the framework of a research project focused on the development of an isotopic ecology of the Southern Argentine Puna. In turn, this isotopic ecology study will provide a valuable reference tool for the interpretation of archaeological data. Here we present 34 pairs of ä13C and ä15N values taken from modern camelid bone collagen (Vicugna vicugna and Lama glama) and 28 pairs ä13C and ä15N values taken from pastures consumed by these animals across the different plant units of Antofagasta de la Sierra (Catamarca, Argentina). The results thus obtained show the existence of habitats with different isotope footprints, based primarily on the different altitudinal floors existing in the study area. The contrasts observed in the plants across the different altitudinal floors are reflected in the difference in ä13C and ä15N values of camelids, especially among the wild ones. In the case of domesticated animals, the modern practice of complementing the diet of the herd with alfalfa (Medicago sativa), a C3 plant, resulted in more negative than expected values.
Keywords: Isotopic ecology; Carbon and nitrogen stable isotopes; Camelids; Plants; Southern Argentine Puna.
INTRODUCCIÓN
Los isótopos estables constituyen una línea de investigación que ha contribuido de manera fundamental al estudio y reconstrucción de los estilos de vida del pasado. En la Argentina, la información obtenida por esta vía de análisis ha sido incorporada en las interpretaciones de distintos estudios arqueológicos desde comienzos de la década de 1990. A partir de entonces, se ha producido un importante incremento en cuanto a variedad de temas, cantidad de valores disponibles, regiones geográficas abarcadas y líneas de evidencia puestas en comparación con este registro (Barberena et al. 2009; Tessone 2010). En el área Andina en general, y en la Puna argentina en particular, la aplicación de esta técnica estuvo orientada, inicialmente, a la obtención de valores en restos humanos con el fin de determinar dietas pasadas (Burger y van der Merwe 1990; Yacobaccio et al. 1997; Olivera y Yacobaccio 1999; Finucane 2007; Killian Galván y Olivera 2008; Pérez y Killian Galván 2011). Posteriormente, numerosos estudios comenzaron a analizar la dieta de poblaciones animales en el pasado, ya sean domésticas o silvestres, a partir de la comparación entre valores de isótopos estables actuales y aquellos obtenidos en arqueofaunas (Fernández y Panarello 1999-2001; Finucane et al. 2006; Mengoni Goñalons 2007; Izeta et al. 2009; Mondini et al. 2010; Yacobaccio et al. 2010; Thorton et al. 2011; Samec 2012, 2014; Samec et al. 2013; Dantas et al. 2014; Szpak et al. 2014; Panarello y Mondini 2015). Dichos estudios han abierto la posibilidad de enfocar distintas problemáticas desde una nueva perspectiva, tales como estrategias de movilidad pastoril, estructura de rebaños domésticos, zonas de forrajeo, áreas de captación de recursos silvestres, entre otras, lo que ha permitido contrastar hipótesis surgidas de otro tipo de registros, como el zooarqueológico o el etnohistórico (Mengoni Goñalons 2008; Yacobaccio et al. 2009, 2010; Samec 2012).
Asimismo, la medición de valores isotópicos en especies vegetales y tejidos correspondientes a especies animales, tanto actuales como arqueológicas, es considerada como un requisito previo para una mejor interpretación de los valores obtenidos en restos humanos recuperados en contextos arqueológicos (Barberena 2007; Tessone 2010; Szpack et al. 2013). Esto es así puesto que los valores isotópicos no tienen un significado intrínseco, sino que deben ser interpretados contextualmente (Barberena 2007). En este sentido, la ecología isotópica de una región provee dicho contexto (Barberena 2007; Tessone 2010). Esta es la subdisciplina que estudia la distribución natural de valores isotópicos de distintas especies animales y vegetales y los factores de fraccionamiento correspondientes, y que permite establecer las relaciones de naturaleza trófica existentes en un ecosistema (Fry 2006). Por lo tanto, la medición de valores isotópicos en especies animales actuales no sólo permite abordar problemáticas ligadas, por ejemplo, a la movilidad de las poblaciones animales, sino que opera como un marco de referencia (sensu Binford 2001) necesario para la interpretación de los datos arqueológicos, ya sean estos obtenidos a partir del análisis de arqueofaunas o de restos humanos (Yacobaccio et al. 2009; Samec 2012).
Ante la ausencia de información isotópica actual para la microrregión de Antofagasta de la Sierra (Catamarca, Argentina) que permita proveer de un contexto para la interpretación de diversos contextos arqueológicos (aunque ver infra, Killian Galván y Salminci 2014; Killian Galván et al. 2014; Panarello y Mondini 2015), el objetivo de este trabajo es comenzar con un estudio sistemático de la ecología isotópica de la Puna Meridional argentina. Para cumplir con el objetivo planteado, se midieron las composiciones isotópicas de carbono y de nitrógeno de plantas, y de colágeno óseo en camélidos silvestres y domésticos actuales recuperados en diversas localidades de la microrregión de Antofagasta de la Sierra. A partir de los resultados obtenidos se discuten los factores que inciden en la variabilidad isotópica de la dieta de camélidos domésticos y silvestres modernos y sus áreas de alimentación, como paso necesario para la interpretación de los conjuntos arqueológicos. Estos resultados pueden plantearse como un marco de referencia para la discusión de las estrategias de pastoreo y de caza empleadas por las poblaciones humanas que ocuparon esta misma área en el pasado.
ÁREA DE ESTUDIO
Antofagasta de la Sierra, ubicada en el ángulo noroeste de la provincia de Catamarca, entre los 25º 50' y 26º 10'S y entre los 67º 30' y 67º 10'O, se incluye dentro de la Puna Salada o Meridional (Figura 1). Para este sector de la Puna se advierte, en términos generales, una extrema aridez (clima árido andino puneño), con precipitaciones de régimen estival inferiores a 100-150 mm anuales, aunque ausentes en algunas temporadas. Por su parte, la temperatura media anual es de 9,5º C, con gran amplitud térmica diurna/nocturna y estacional, y baja presión atmosférica. La red hidrográfica es endorreica, alimentada por las escasas precipitaciones, el régimen de deshielo (de noviembre a marzo) y las aguas subterráneas. Como en el resto de la Puna, se observa un desarrollo de distintas comunidades vegetales determinadas por la variable altitudinal. En este sentido, se destaca que a menos de 3800 msnm aparece el Tolar, caracterizado por comunidades de arbustos compuestas por la familia de las Asteraceae (i.e., Fabiana spp., Bacharis boliviensis, Adesmia spp.). Entre los 3900 a 5000 msnm se extiende el Pajonal (estepa de altura), compuesto por la familia de las Poaceae, principalmente Festuca spp. y Stipa spp. Entre el Pajonal y el piso de Tolar –3800/3900 msnm– hay una zona de ecotono con una comunidad mixta de arbustos y estepa. Estos pisos constituyen el 99% de la matriz del desierto; el restante 1% está ocupado por los humedales conocidos como vegas (Tchilinguirian y Olivera 2009). Estas últimas pueden ser encontradas a diversas altitudes, y se dividen, en el área de estudio, en dos subunidades: vega prepuneña (3400- 3800 msnm), compuesta principalmente por Juncus sp., Juncus balticus, Tanuculus cymbajaria, Nitrophila australis y Distichlis humilis; y la vega puneña (3800 a 4500 msnm), donde el taxón más abundante es Juncus sp., aunque también abundan Ranunculus cynbaria, Ranunculus sp., Festusca desertícola var. Paupera y Hypsella ologophila (Haber 1991).
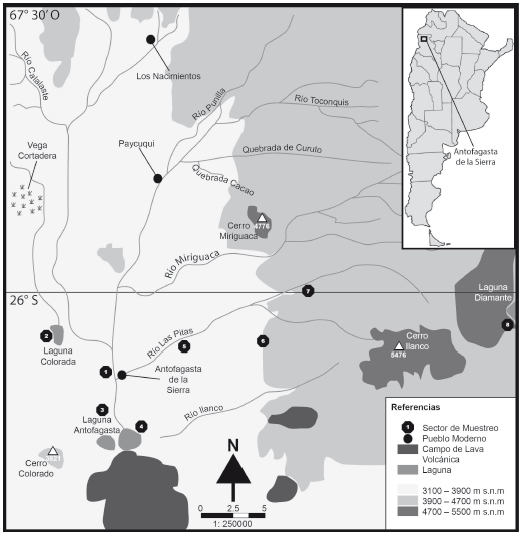
Figura 1. Área de investigación (modificado de Aschero et al. 2012), donde se indica la procedencia de las muestras: 1. río Punilla, 2. laguna/Salar Colorada, 3. cerros del Coypar, 4. laguna Antofagasta este, 5. río Las Pitas, 6. camino Laguna Diamante, 7. Real Grande, y 8. laguna Diamante.
En general, las especies vegetales presentes corresponden a las tres vías fotosintéticas existentes: C3, C4 y CAM. Estas últimas se encuentran representadas principalmente por cactáceas, las que, si bien forman parte de la dieta de los camélidos, lo hacen en una muy baja proporción. Las plantas C3 se hallan representadas por la totalidad de las especies arbustivas, numerosas hierbas y gramíneas, y los vegetales que siguen la vía C4 se encuentran integrados mayormente por gramíneas (Haber 1991; Fernández y Panarello 1999- 2001; Samec et al. 2015). La presencia de una serie de microambientes con recursos vegetales, animales y minerales bien definidos en relación con las diferencias altitudinales ha permitido distinguir entre tres sectores principales dentro de la microrregión (Olivera 2006): fondo de cuenca (3400-3550 msnm), sectores intermedios (3550-3900 msnm) y quebradas de altura (3900-4600 msnm).
MATERIALES Y SU PROCEDENCIA
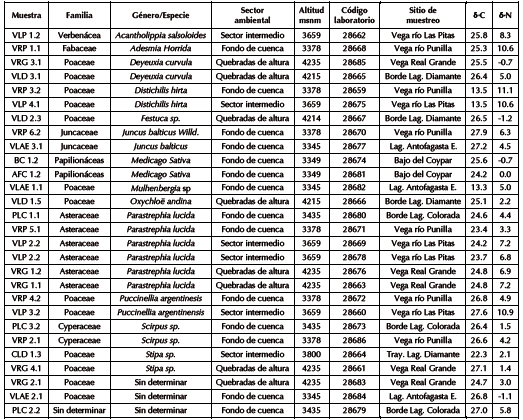
En este trabajo se presentan los valores ä13C y ä15N correspondientes a 28 plantas herbáceas, arbustivas, gramíneas y juncáceas recolectadas a partir de la consideración de los tres sectores microambientales ubicados a distintas cotas altitudinales de Antofagasta de la Sierra. Las especies seleccionadas para el análisis corresponden en su mayoría a aquellas preferidas por los camélidos, aunque el muestreo también fue diseñado para recolectar las plantas más representativas de cada sector de muestreo. De esta manera, se incorporaron al estudio taxones como Parastrephia sp., que, si bien no suelen ser los preferidos de los camélidos, se encuentran muy representados en las distintas comunidades muestreadas, por lo que pueden ser ingeridos en bajas proporciones (ver también Samec et al. 2015). En este mismo sentido, se recolectó al menos un espécimen de cada especie reconocida en el campo, y más de uno en caso de estar altamente representada en el sector de muestreo. Los especímenes provenientes del fondo de cuenca (n = 14) fueron recolectados a partir de transectas llevadas a cabo en la vega del río Punilla (VRP), en el borde de la laguna/ Salar Colorada (PLC), al pie de los cerros del Coypar (BC) y en la vega laguna Antofagasta este (VLAE). Para los sectores intermedios (n = 6), las especies vegetales fueron recolectadas en transectas realizadas en la vega del río Las Pitas (VLP) y en el ecotono tolar/ pajonal camino a laguna Diamante (CLD). Por último, los especímenes representantes de las quebradas de altura (n = 8) fueron obtenidos a partir de transectas que cubrieron los sectores de vega y pajonal en Real Grande (VRG) y en la vega de laguna Diamante (VLD). La recolección de plantas en estas distintas localidades permitió abarcar un gradiente altitudinal entre los 3345 y 4235 msnm (Figura 1). De todas las muestras se seleccionaron hojas, las cuales se guardaron en una prensa entre papel tissue hasta ser enviadas para su identificación a la Cátedra de Botánica Agrícola, Facultad de Agronomía, Universidad de Buenos Aires. Sin embargo, cabe mencionar que no todas las muestras analizadas en este estudio pudieron ser identificadas a nivel especie o género.
Asimismo, se presentan aquí 32 pares de valores de ä13C y ä15N medidos sobre colágeno óseo de llamas y vicuñas actuales de Antofagasta de la Sierra. Cada par de valor analizado corresponde a un individuo, que es la principal unidad de observación en el análisis isotópico. En el caso de los animales domésticos (n = 15), el material óseo fue recolectado dentro o en las cercanías de asentamientos pastoriles en actividad de los que se conoce el área de captación de recursos. La conformación de una muestra de llamas actuales, con el objetivo de elaborar un marco de referencia para la interpretación de contextos pastoriles arqueológicos, se ha enfrentado con dos limitaciones principales. Estas últimas se relacionan con la naturaleza de las actividades pastoriles practicadas en la actualidad en Antofagasta de la Sierra. En primer lugar, es preciso destacar que la práctica tradicional andina de pastorear llamas en distintos pisos altitudinales –tal como nos fuera relatado por distintos pastores del área de estudio y como se menciona en la bibliografía de prácticas pastoriles andinas (Gundermann 1984; Göbel 1994, 2002; Yacobaccio et al. 1998, entre otros)– ha ido mermando en el área de estudio en las últimas décadas. Esto repercutió en la cantidad de muestras recuperadas en cada sector, obteniéndose una menor cantidad de especímenes óseos de camélidos domésticos en puestos de quebradas de altura (n = 4), en relación con aquellos obtenidos en los sectores de fondo de cuenca (n = 10). Asimismo, de aquellos individuos obtenidos en sectores de altura, únicamente dos fueron pastoreados siguiendo la práctica de alternancia entre distintos sectores microambientales. En cuanto a los otros dos ejemplares de Lama glama de quebradas de altura, uno fue pastoreado exclusivamente en la comunidad de pajonal, mientras que otro individuo fue alimentado en la comunidad de tolar de fondo de cuenca y sólo llevado al puesto de altura para su sacrificio y faenamiento. En segundo lugar, y en relación directa con los cambios en las prácticas de manejo pastoril, en la actualidad es común complementar la alimentación de las llamas con forraje artificial, en particular con alfalfa (Medicago sativa), lo que limita la posibilidad de conseguir carcasas de llamas alimentadas únicamente con base en pasturas naturales.
Para la obtención de la porción de la muestra que representa a los animales silvestres se consideraron los hábitos territoriales de las poblaciones al realizar el muestreo. De este modo, las muestras se encuentran integradas por carcasas recolectadas en el campo en función de criterios tales como la disponibilidad y la conservación, aspectos que condicionaron la cantidad de especímenes recuperados en cada sector, con un sesgo a favor de los elementos recuperados en las quebradas de altura. La muestra correspondiente a vicuñas quedó conformada entonces por un total de 18 individuos: cinco recuperados en fondo de cuenca, dos en sectores intermedios y doce en quebradas de altura.
TÉCNICAS DE LABORATORIO
Los trabajos de laboratorio se llevaron a cabo en el Instituto de Geocronología y Geología Isotópica (INGEIS) bajo la supervisión del Dr. Augusto Tessone. El procesamiento de las plantas actuales implicó una limpieza a partir de baños de ultrasonido con agua bidestilada. Posteriormente, se realizó un secado de las plantas en un horno por 24 horas a < 60° C. Por último, se procedió a la molienda de las hojas para su medición en un espectrómetro de masas. En el caso de los huesos, se seleccionaros aquellos con tejidos densos, preferentemente diáfisis. La preparación de las muestras para realizar las mediciones isotópicas en colágeno óseo siguió los procedimientos descriptos por Tykot (2004), con algunas modificaciones. Inicialmente, se realizó una limpieza física de la superficie del hueso con un torno y se lo sometió a lavados ultrasónicos con agua bidestilada a fin de remover la contaminación superficial y las zonas con potencial alteración diagenética. En forma posterior, se seleccionó aproximadamente un gramo de cada elemento óseo (uno por individuo) y se eliminaron los ácidos húmicos con hidróxido de sodio al 0,1 M durante 24 horas. Para la obtención del colágeno, el hueso fue desmineralizado en ácido clorhídrico al 2% durante 72 horas, el cual era renovado cada 24 horas. Finalmente, cada muestra fue inmersa en NaOH (0.1 M) por otras 24 horas. El material resultante fue lavado con agua deionizada y secado en horno a < 60º C durante 24 horas (Tykot 2004; Yacobaccio et al. 2010).
Tanto los vegetales como los colágenos óseos se pesaron en balanza analítica y se colocaron dentro de una cápsula de plata. Estas fueron introducidas, mediante un muestreador automático, en un analizador elemental CARLO ERBA EA1108, acoplado a un espectrómetro de masas para relaciones isotópicas (IRMS) Thermo Delta V Advantage mediante una interfaz CONFLO IV™. Los resultados se expresan como desviaciones isotópicas ä13C y ä15N respecto de las referencias internacionales VPDB y AIR, respectivamente. La incertidumbre (1ä) es ± 0,2‰ en ambos isótopos. En el caso del colágeno, la calidad de la señal isotópica de la muestra fue evaluada utilizando la relación C:N (De Niro 1985).
RESULTADOS
Vegetación
El análisis de las composiciones isotópicas de los 28 especímenes de plantas muestreadas arrojó como resultado que los valores ä13C oscilan entre -24,3‰ y -13,3‰, con un valor medio de -24‰ y los ä15N entre -1,2‰ y 11,1‰, con un valor medio de 4,6‰ (Tabla 1 y Figura 2). Del relevamiento realizado en este trabajo, la mayor parte de las especies corresponde a plantas con patrón fotosintético C3 (n = 25), las cuales fueron recuperadas en todos los pisos altitudinales. Los valores ä13C para las plantas C3 oscilan entre -27,9‰ y -22,3‰, con un valor medio de -25,6‰; mientras que los de ä15N oscilan entre -1,2‰ y 10,9‰, con un valor medio de 4,1‰. Por su parte, las plantas C4 fueron registradas en menor proporción (n = 3), únicamente por debajo de los 3900 msnm y fueron recolectadas en vegas prepuneñas. Estas se corresponden con los denominados pasto salado (Distichlis hirta phil.) y pasto vicuña (Mulhenbergia sp.), y los valores ä13C oscilaron entre -13,5‰ y 13,8 y los de ä15N entre 11,1‰ y 5‰. En la Figura 2 se observa la separación de los valores ä13C entre plantas C3 y C4. Dicha separación no ha sido observada en los valores de ä15N, si bien es preciso considerar las diferencias en el tamaño de muestra entre ambos tipos de plantas.
Con la finalidad de simplificar el análisis, se ha decidido separar las plantas por pisos altitudinales, utilizado como límite los 3900 msnm. Dicho límite marca la división entre los dos rangos altitudinales principales con diferentes comunidades vegetales del área de estudio: tolar y pajonal. El valor promedio para el ä13C de las especies vegetales de fondo de cuenca y sectores intermedios (<3900 msnm) es de -23,8‰, con un rango de distribución entre -27,9‰ y -13,3‰. Por su parte, las plantas de quebradas de altura (>3900 msnm) presentan un valor medio de -25,6‰, con valores mínimos y máximos de -27,1‰ a -24,7‰, lo cual implica un empobrecimiento de 1,7‰ y un menor rango de distribución de valores respecto del anterior conjunto (Figura 2). No obstante, si se toman como referencia las medianas, se observa un enriquecimiento de 0,3‰ de las plantas de altura (mediana de -25,3) respecto de las de menor altura (mediana de -25,6‰). Esto permite corroborar que las principales diferencias en los valores ä13C se deben a la presencia/ausencia de plantas C4 en los distintos sectores altitudinales (Figura 2). En lo que se refiere a la vegetación C3, no existe una diferencia significativa entre las dos principales unidades muestreadas (One Way ANOVA F=1.12 p=0,39 >0.05).

Figura 2. Valores δ13C y δ15N correspondientes a los 28 vegetales muestreados en los sectores de fondo de cuenca, sectores intermedios y quebradas de altura.
En el caso del ä15N, las plantas obtenidas por debajo de los 3900 msnm presentan un valor medio de 5,3‰, con un rango de distribución entre -1,1‰ y 11,1‰; mientras que para las plantas obtenidas por encima de los 3900 msnm, el valor medio es de 2,9‰, con valores mínimos y máximos de -1,2‰ y 7,2‰ respectivamente (Tabla 1). De esta forma, se registra un empobrecimiento del 2,4‰ del ä15N con la altitud, y se observa que, si bien los valores mínimos se superponen, los máximos se muestran enriquecidos en las plantas obtenidas por debajo de los 3900 msnm (Figura 2). Dicho patrón de empobrecimiento con la altitud se verifica al tomar en cuenta las medianas, de 5‰ para las plantas por debajo de los 3900 y de 2,6‰ por encima de dicha cota altitudinal. Por lo tanto, los resultados alcanzados muestran una correlación negativa y estadísticamente significativa (One Way ANOVA F=4.32 p=0,048 >0.05) entre la altitud y los valores ä15N, independientemente de la vía fotosintética de las especies vegetales consideradas, lo cual podría ser relacionado con la mayor disponibilidad de agua en las áreas más altas (Ambrose y De Niro 1986; Amundson et al. 2003; Samec et al. 2013, 2015).
Tabla 1. Valores de ä13C y ä13N de plantas actuales de Antofagasta de la Sierra.
Colágeno óseo
Los resultados obtenidos en colágeno óseo de camélidos actuales (N = 33) han arrojado valores de ä13C que oscilan entre -21,9 ‰ y -14,0 ‰, con una media de -19,8 ‰; y valores de ä15N que varían entre 1,5 ‰ y 8,4 ‰, con una media de 4,9 ‰ (Tabla 2 y Figura 3). Respecto de la integridad de los especímenes analizados, la razón C/N es coherente con el buen estado de conservación de las muestras, promediando 3,3 con un valor mínimo de 3,0 y uno máximo de 3,5 (Tabla 2). En general, se puede observar en la Figura 3 que las vicuñas presentan mayor variabilidad en su dieta en relación con las llamas, y que las vicuñas recuperadas por debajo de los 3900 msnm son las que presentan los valores ä13C y ä15N más altos (ver infra). A continuación, evaluamos los resultados obtenidos en colágeno óseo de vicuñas y llamas por separado, puesto que la información isotópica brindada para cada una de las especies responde a diferentes variables. En este sentido, es preciso considerar que la alimentación y el comportamiento territorial de las llamas están sujetos a las prácticas de manejo empleadas por los pastores actuales, mientras que tales factores se encuentran ausentes en el caso de las vicuñas (Samec 2012).
Tabla 2 Valores de ä13C y ä13N de llamas y vicuñas actuales de Antofagasta de la Sierra.

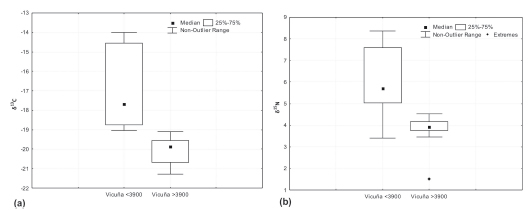
Figura 3. Valores ä13C y ä15N correspondientes a llamas y vicuñas actuales obtenidas en sectores de muestreo por encima y por debajo de los 3900 msnm.
Vicugna vicugna
El análisis de las composiciones isotópicas del colágeno óseo de vicuñas permitió registrar que estas ostentan una dieta variable, comprendida entre los valores de ä13C de -21,3 y -14,0‰, y valores de ä15N de 1,5‰ y 8,4‰ (Tabla 2 y Figura 3). Para analizar los factores que determinan dicha variabilidad, dividimos la muestra en dos subgrupos, empleando como límite los 3900 msnm (Figura 4), siguiendo los criterios utilizados en el análisis de especies vegetales. Los valores ä13C para vicuñas obtenidas por debajo de los 3900 msnm oscilan entre 19,1‰ a -14,0‰, con un valor medio de 17,1‰. Por su parte, las vicuñas recuperadas por encima de los 3900 msnm presentan un valor medio de -20,1‰, empobrecidas en un 3‰ respecto de los valores anteriores, y muestran un rango de distribución más acotado, con valores que oscilan entre -21,3‰ a -19,1‰ (Figura 4a). A fin de evaluar la significancia estadística de estos datos se ha realizado un test ANOVA de una vía (One Way ANOVA) resultando en un valor de F = 21,02 con un valor de p <0,01, lo que permite establecer la existencia de una variación estadísticamente significativa en la dieta de esta especie en función del rango altitudinal considerado, con un empobrecimiento a medida que aumenta la altitud.

Figura 4. Box-plots de vicuñas actuales separadas por sector de muestreo por encima y por debajo de los 3900 msnm: a) valores ä13C y b) valores ä15N.
Por su parte, los valores ä15N para vicuñas obtenidas por debajo de los 3900 msnm muestran un valor medio de 6,1‰, con un rango de distribución entre 3,4‰ y 8,4‰; mientras que, para los individuos obtenidos por encima de los 3900 msnm, este valor es de ä15N 3,9‰, con valores mínimos y máximos de 1,5‰ y 4,5‰ respectivamente. De esta forma, se registra un empobrecimiento del 2,3‰ en el valor medio de ä15N con la altitud. En la Figura 4.b se observa que el valor mínimo de ä15N de 1,5‰ para las vicuñas de sectores de altura corresponde a un outlier. Si se descarta este último valor, se observa una superposición entre los valores mínimos obtenidos para ambos subgrupos. Sin embargo, y en concordancia con lo registrado en las plantas actuales del área de estudio, los valores máximos se muestran enriquecidos en las vicuñas obtenidas por debajo de los 3900 msnm; por lo tanto, se registra una mayor variabilidad en su dieta en comparación con los individuos alimentados por encima de dicha cota altitudinal.
Lama glama
Los resultados obtenidos en llamas actuales arrojaron valores de ä13C de entre -21,9 y -19,1‰, con un valor medio de -20,9‰ y valores de ä15N de 2,9 a 6,9‰; la media presenta un valor de 5,3‰ (Tabla 2 y Figura 3). Tomados en conjunto, los valores de carbono indican que las llamas del área de estudio ostentan una dieta poco variable. Sin embargo, es preciso considerar que los valores obtenidos –en general empobrecidos en ä13C y señalando una dieta compuesta principalmente por vegetales C3– indicarían la influencia de las estrategias pastoriles actuales en la alimentación de las llamas. Esto se debe a la inclusión de la alfalfa como forraje suplementario en su dieta, que en el área de estudio presenta una media de ä13C de -24,9‰ (Tabla 1). En el caso del ä15N, la variación observada se corresponde con aquella registrada en las plantas recolectadas por debajo de los 3900 msnm, las cuales muestran un amplio rango de distribución. Por otra parte, el análisis exploratorio separando los individuos que se alimentan por debajo de los 3900 msnm de los pocos que lo hacen por encima de esta cota altitudinal y discriminando entre aquellos con o sin forraje complementario, permitió detectar ciertos patrones interesantes. Si se toman en cuenta las muestras de llamas cuya alimentación se basó íntegramente en pasturas naturales, se observa en la Figura 5.a que el individuo alimentado por debajo de los 3900 msnm (muestra AA 19) registra un valor ä13C de -19,1‰, mientas que aquel cuya dieta responde exclusivamente a la comunidad de pajonal por encima de los 3900 msnm (muestra AA 34) indica un valor ä13C de -21,9‰, empobrecido por lo tanto en un 2,8‰. Asimismo, los individuos cuya dieta se basó en una alternancia entre distintas áreas de pasturas (muestras AA 37 y 38) muestran un valor medio de -19,6‰, más enriquecidos respecto del valor obtenido para pajonal y acercándose a aquel registrado para la llama alimentada en la comunidad de tolar.

Figura 5. Box-plots de llamas actuales separadas por sector de muestreo por encima y por debajo de los 3900 msnm: a) valores ä13C y b) valores ä15N.
El mismo patrón es observado al considerar los valores ä15N, puesto que para el individuo alimentado por debajo de los 3900 msnm se obtuvo un valor de 6,4‰; mientras que para aquel obtenido por encima de los 3900 msnm es de ä15N 3,8‰ (Tabla 2 y Figura 5b). De esta forma, se registra un empobrecimiento del 2,6‰ de los valores ä15N con la altitud. En el caso de las llamas alimentadas en ambas cotas altitudinales (n = 2) se registra una mediana de 5,5‰. Por lo tanto, si bien basados por el momento en muy pocos valores, la tendencia detectada no contradice aquella esperada, con un empobrecimiento en los valores de ambos isótopos estables con la altitud. En el caso de las muestras de llamas obtenidas por debajo de los 3900 msnm y cuya dieta fue complementada con alfalfa (n = 11), se registra un valor medio de ä13C de -21,1‰, empobrecido por lo tanto en un 2‰ en relación con el individuo de fondo de cuenca alimentado únicamente con base en pasturas naturales. En cuanto al nitrógeno, el valor medio registrado es de 5,3‰, más bajo que el valor obtenido para llama de fondo de cuenca, aunque acercándose al valor medio obtenido en llamas alimentadas en distintas cotas altitudinales.
DISCUSIÓN
El análisis de las composiciones isotópicas de ä13C y ä15N en especies vegetales de la microrregión de Antofagasta de la Sierra permitió registrar la existencia de comunidades vegetales con señales isotópicas diferentes, estructuradas fundamentalmente por la variable altitudinal. Por un lado, los valores de ä13C permiten corroborar, tal como ha sido registrado en otras investigaciones realizadas en la Puna Argentina (Fernández y Panarello 1999-2001; Yacobaccio et al. 2009, 2010; Samec et al. 2013; Samec 2014; Panarello y Mondini 2015), la presencia de plantas C4 por debajo de los 3900 msnm, y la ausencia de plantas con este patrón fotosintético por encima de dicha cota altitudinal. La diferencia en las proporciones o en la presencia/ausencia de plantas C3 y C4 según el sector de muestreo resulta, por lo tanto, en valores medios ä13C más altos en las pasturas naturales ubicadas en cotas altitudinales más bajas y secas (ver también Szpack et al. 2013; Samec et al. 2015). Sin embargo, no se ha observado una correlación significativa entre la altitud y los valores ä13C en plantas C3, resultado que se acerca al obtenido en la Puna Septentrional (Samec et al. 2015).
Por otro lado, los valores de ä15N variaron en función de la altitud, dado que se registró un mayor rango de distribución y la presencia de valores más elevados en especies recogidas en los sectores más secos que se encuentran por debajo de los 3900 msnm, en fondo de cuenca y sectores intermedios; mientras que los más bajos se encuentran por encima de dicha cota, en la comunidad de pajonal de las quebradas de altura, donde se registra una mayor humedad relativa. Estos contrastes entre vegetación ubicada en diferentes cotas altitudinales se ven reflejados en los valores isotópicos obtenidos en vicuñas. De esta manera, se ha registrado la existencia de una correlación negativa entre los valores ä13C y ä15N en vicuñas y la altitud, puesto que a medida que esta última se incrementa, ambos marcadores isotópicos muestran valores más bajos y un menor rango de distribución. Por lo tanto, se observa cierta preferencia por parte de las vicuñas por una alimentación con mayor proporción de especies C4 cuando estas se encuentran disponibles; es decir, en aquellos ambientes situados por debajo de los 3900 msnm, aspecto que concuerda con la información etológica disponible para dicha especie (Borgnia et al. 2010). Asimismo, las diferencias detectadas en la dieta de ambos subgrupos –es decir, por encima y por debajo de los 3900 msnm– son las esperadas en función de los hábitos extremadamente territoriales de esta especie, en la que los patrones migratorios entre pisos altitudinales están prácticamente ausentes (Arzamendia y Vilá 2006; Samec 2012).
En el caso de las llamas, se ha mencionado previamente que la recolección de muestras correspondientes a ellas se ha visto limitada en su objetivo de analizar los patrones de movilidad entre distintos pisos altitudinales, puesto que en la actualidad la mayoría de los rebaños son alimentados con alfalfa cultivada en el área. Esto último se vio reflejado en valores ä13 en general empobrecidos, y marcando el predominio de especies vegetales C3. No obstante, los resultados del análisis exploratorio efectuado mediante la discriminación de las muestras de Lama glama según el tipo de dieta (con o sin forraje artificial suplementario) y sector altitudinal, si bien basado en muy pocos valores, permitieron confirmar la tendencia registrada previamente hacia un empobrecimiento y un menor rango de distribución en los valores de ambos isótopos estables con la altitud. Por otra parte, es interesante remarcar que la media registrada para los valores de ä13C en las llamas del área de estudio se encuentra empobrecida en un 1,7‰ con respecto a aquella obtenida del análisis de una muestra de 46 llamas procedentes de diferentes sectores altitudinales de la Puna Septentrional (PS), que presentaron un valor medio de 19,21‰ (Samec 2012). Es más, si la comparación se efectúa entre los valores medios de llamas de ambas punas cuyos territorios de alimentación se encuentran ubicados por debajo de los 3900 msnm, se registra un empobrecimiento del 3,4‰ en los valores ä13C en las llamas de la Puna Meridional (PM) (valor medio de 21,1‰) en relación con los de la Puna Septentrional (valor medio de 17,74‰). Esta diferencia obedecería principalmente al mayor aporte de plantas C3 en nuestra área de estudio a partir de la complementación de la dieta con alfalfa.
En este mismo sentido, al comparar únicamente los valores obtenidos para llamas que basaron su dieta en pasturas naturales (n = 4), la diferencia entre las medias de ambos conjuntos se reduce a un 0,7 ‰ y de las medianas a 0,3‰. Asimismo, cuando la comparación se realiza a partir de los valores medios de ä13C obtenidos para vicuñas de ambas punas (PS: 18,4‰; PM; 18,9‰), se observa que las del área de estudio se encuentran empobrecidas en tan sólo un 0,5‰ respecto de las de la Puna Septentrional. Por lo tanto, los datos isotópicos medidos sobre especímenes de Lama glama demuestran ser un ejemplo que ilustra la utilidad de estas técnicas en la evaluación de la intervención humana en la dieta de las poblaciones de camélidos domésticos, ya que parecerían reflejar diferencias respecto de las expectativas que se tenían sobre la dieta de esta especie en relación con los valores obtenidos en plantas actuales.
CONCLUSIONES
Los resultados presentados en este trabajo sobre el análisis de las relaciones isotópicas del carbono y nitrógeno de las plantas de la microrregión de Antofagasta de la Sierra permiten comenzar a establecer una línea de base de las cadenas tróficas terrestres del área de estudio y proveen de un contexto a partir del cual poder contrastar los datos generados del análisis isotópico del colágeno óseo de camélidos. A su vez, estos últimos llevan a confirmar, en consonancia con los datos publicados para otros sectores de la Puna Argentina (Fernández y Panarello 1999-2001; Yacobaccio et al. 2009, 2010; Samec et al. 2013; Samec 2014), que la dieta de vicuñas y, en menor medida, de las llamas del área de estudio, se correlaciona con la variación altitudinal, de manera que a medida que esta variable aumenta, los valores de ä13C y ä15N se vuelven más negativos. Dicha variación en las firmas isotópicas a través de las diferentes zonas ecológicas proveen un punto de partida a partir del cual reconstruir la dieta antigua de los camélidos de la microrregión de Antofagasta de la Sierra, tanto silvestres como domesticados, permitiéndonos de esta manera inferir las estrategias de pastoreo y zonas de predación utilizadas en el pasado (Yacobaccio et al. 2010). De este modo, se espera que las variables altitud e intervención humana en la dieta –ya sea mediante estrategias de movilidad entre distintos sectores o por su complementación mediante forrajes artificiales (e.g., maíz)– determinen las características de la alimentación de los camélidos domésticos en el pasado. Al mismo tiempo, se espera que la diferencia entre la alimentación de las especies domésticas y silvestres –en función de la utilización complementaria de ambientes por parte de los pastores y la territorialidad exhibida por las poblaciones de camélidos silvestres– se evidencie en el registro isotópico de los sitios pastoriles de Antofagasta de la Sierra.
En síntesis, el presente trabajo constituye un aporte a la configuración de un modelo ecológico actual de poblaciones animales y especies vegetales de la Puna Meridional Argentina. Puesto que representa uno de los primeros pasos en pos de obtener tal modelo, se vio sujeto a ciertas limitaciones, en especial en lo referido al tamaño de muestra empleada. No obstante, los resultados isotópicos actuales recabados hasta el momento permitieron observar tendencias interesantes y elaborar expectativas respecto de los factores determinantes en la dieta de las poblaciones de camélidos susceptibles de haber operado en contextos arqueológicos. Asimismo, cabe destacar que, a los estudios isotópicos aquí realizados, se le agregan aquellos que están siendo llevados a cabo por distintos investigadores del área de estudio (Killian Galván y Samec 2012; Killian Galván y Salminci 2014; Killian Galván et al. 2014; Panarello y Mondini 2015), que permitirán en un futuro próximo aumentar los valores disponibles y contribuir, junto con las investigaciones desarrolladas en la Puna Septentrional, a la configuración de un marco de referencia robusto para el trabajo en ecología isotópica en los Andes en general.
REFERENCIAS CITADAS
1. Ambrose, S. H. y M. J. De Niro 1986 The isotopic ecology of East African mammals. Oecologia 69: 395-406. [ Links ]
2. Amundson, R., A. T. Austin, E. A. G. Schuur, K. yoo, V. Matzek, C. Kendall, A. Uebersax, D. Brenner y W. T. Baisden 2003 Global patterns of the isotopic composition of soil and plant nitrogen. Global Biogeochemical Cycles 17: 10-31. [ Links ]
3. Arzamendia, Y. y B. L. Vilá 2006 Estudios etoecológicos de vicuñas en el marco de un plan de manejo sustentable: Cieneguillas, Jujuy. En Investigación, conservación y manejo de vicuñas. Proyecto MACS, editado por B. Vilá, pp. 69-84. INCO, Buenos Aires. [ Links ]
4. Aschero, C., A. Izeta y S. Hocsman 2012 New Data on South American Camelid Bone Size Changes during Middle-Late Holocene Transition: Osteometry at Peñas Chicas 1.5 (Antofagasta de la Sierra, Argentinian Puna). International Journal of Osteoarchaeology 9: 204-214. [ Links ]
5. Barberena, R. 2007 Arqueología y Biogeografía Humana en Patagonia Meridional. Tesis Doctoral inédita. Facultad de Filosofía y Letras, Universidad de Buenos Aires, Buenos Aires. [ Links ]
6. Barberena, R., A. Zangrando, A. Gil, G. Martínez, G. Politis, L. Borrero y G. Neme 2009 Guanaco (Lama guanicoe) isotopic ecology in southern South America: spatial and temporal tendencies, and archaeological implications. Journal of Archaeological Science 36: 2666-2675. [ Links ]
7. Binford, L. 2001 Constructing frames of reference. An analytical method for archaeological theory building using huntergatherer and environmental data sets. University of California Press, Berkeley. [ Links ]
8. Borgnia M., B. Vilá y M. Cassini 2010 Foraging ecology of Vicuña, Vicugna vicugna, in dry Puna of Argentina. Small Ruminant Research 88 (1): 44-53. [ Links ]
9. Burger, R. L. y N. van Der Merwe 1990 Maize and the origin of highland Chavin civilization: an isotopic perspective. American Anthropologist 92: 85-95. [ Links ]
10. Dantas, M., G. G. Figueroa y A. Laguens 2014 Llamas in the cornfield: prehispanic agro-pastoral system in the Southern Andes International Journal of Osteoarchaeology 24: 149-165. [ Links ]
11. De Niro, M. J. 1985 Postmortem preservation and alteration of in vivo bone collagen isotope ratios in relation to paleodietary reconstruction. Nature 317: 806-809. [ Links ]
12. Fernández, J. y H. O. Panarello 1999-2001 Isótopos del carbono en la dieta de herbívoros y carnívoros de los Andes Jujeños. Xama 12-14: 71-85. [ Links ]
13. Finucane, B. C. 2007 Mummies, maize, and manure: multi-tissue stable isotope analysis of late prehistoric human remains from the Ayacucho Valley, Peru. Journal of Archaeological Science 34: 2115-2124. [ Links ]
14. Finucane, B. C., P. Maita Agurto y W. H. Isbell 2006 Human and animal diet at Conchopata, Perú: stable isotope evidence for maize agriculture and animal management practices during the Middle Horizon. Journal of Archaeological Science 33: 1766-1776. [ Links ]
15. Fry, B. 2006 Stable isotope ecology. Nueva York, Springer. [ Links ]
16. Göbel, B. 1994 El manejo del riesgo en la economía pastoril de Susques. En Zooarqueología de camélidos 1, editado por D. C. Elkin, C. Madero, G. L. Mengoni Goñalons, D. E. Olivera, M. C. Reigadas y H. D. Yacobaccio, pp. 43-56. Grupo de Zooarqueología de Camélidos, Buenos Aires. [ Links ]
17. Göbel, B. 2002. La arquitectura del pastoreo: uso del espacio y sistema de asentamientos en la Puna de Atacama (Susques). Estudios Atacameños 23: 53-76. [ Links ]
18. Gundermann, H. 1984 Ganadería Aymara, Ecología y Forrajes: Evaluación Regional de una Actividad Productiva Andina. Chungara 12: 99-124. [ Links ]
19. Haber, A. 1991 La estructuración del recurso forrajero y el pastoreo de camélidos. Actas del XI Congreso Nacional de Arqueología Chilena: 139-150. Chile. [ Links ]
20. Izeta, A., A. Laguens, M. Marconetto y M. Scattolin 2009 Camelid Handling in the Meridional Andes during the First Millennium AD: A Preliminary Approach Using Stable Isotopes. International Journal of Osteoarchaeology 19: 204-214. [ Links ]
21. Killian Galván, V. A. y C. T. Samec 2012 A cada uno su verdad culinaria: patrones paleodietarios y variables ambientales en el NOA. En Entre Pasados y Presentes III. Estudios Contemporáneos en Ciencias Antropológicas, compilado por N. Kuperszmit, T. Lagos Mármol, L. Mucciolo, M. Sacchi, pp. 487-508. Colección Investigación y Tesis, Mnemosyne, Buenos Aires (publicado en CD-ROM). [ Links ]
22. Killian Galván, V. A. y P. Salminci 2014 Aportes a la ecología isotópica: información actual y sistemas de regadío arqueológicos en la microrregión de Antofagasta de la Sierra (Provincia de Catamarca, Argentina). Comechingonia. Revista de Arqueología 18: 51-72. [ Links ]
23. Killian Galván, V. A. y D. E. Olivera 2008 First ä13C values for human skeletal remains from South Western Puna (Jujuy, Argentina). Trabajo presentado en el VI South American Symposium on Isotope Geology, San Carlos de Bariloche, Argentina. [ Links ]
24. Killian Galván, V. A., N. Oliszewski, D. E. Olivera y H. O. Panarello 2014 Intraspecific variability in the ä13C and ä15N values of archaeological samples of Zea mays cobs (Northeastern Argentinean Puna). En Physical, Chemical and Biological Markers in Argentine Archaeology: Theory, Methods and Application, editado por D. M. Kligmann y M. R. Morales, pp. 39-51. BAR International Series 2678. Archaeopress, Oxford. [ Links ]
25. Mengoni Goñalons, G. L. 2007 Camelid management during Inca times in N. W. Argentina: models and archaeozoological indicators. Anthropozoologica 42 (2): 129-141. [ Links ]
26. Mengoni Goñalons, G. L. 2008 Camelids in ancient Andean societies: A review of the zooarchaeological evidence. Quaternary International 185: 59-68. [ Links ]
27. Mondini, M., H. Panarello Y A. Zangrando 2010 First isotopic evidence in southern Puna camelids throughout the Holocene. Póster presentado en el 11º ICAZ Conference (Session 2). París. [ Links ]
28. Olivera, D. 2006 Recursos bióticos y subsistencia en Sociedades Agropastoriles de la Puna Meridional Argentina. Comechingonia 9: 19-56. [ Links ]
29. Olivera, D. y H. Yacobaccio 1999 Estudios de paleodieta en poblaciones humanas de los Andes del Sur a través de isótopos estables. Trabajo presentado en el 5º Congreso Nacional de Paleopatología, Alcalá La Real, Jaén. MS. [ Links ]
30. Panarello, H. y M. Mondini 2015 Evidencia isotópica de camélidos y vegetales modernos en la puna sur. Relevancia para la arqueología de la región. Cuadernos del Instituto Nacional de Antropología y Pensamiento Latinoamericano 24 (2): 45-62. [ Links ]
31. Pérez, M. y V. A. Killian Galván 2011 Doncellas (Puna Septentrional, Jujuy, Argentina): Nuevos enfoques a partir del estudio cerámico y el análisis paleodietario. Estudios Atacameños 42: 79-100. [ Links ]
32. Samec, C. 2012 Variabilidad dietaria en camélidos de la Puna: un modelo actual a partir de la evidencia isotópica. En Entre pasados y presentes III. Estudios contemporáneos en Ciencias Antropológicas, compilado por N. Kuperszmit, T. Lagos Mármol, L. Mucciolo, M. Sacchi, pp. 666-683. Colección Investigación y Tesis. Mnemosyne (publicado en CD-ROM). [ Links ]
33. Samec, C. 2014 Ecología isotópica en la Puna Seca Argentina: un marco de referencia para el estudio de las estrategias de pastoreo en el pasado. Cuadernos del Instituto Nacional de Antropología y Pensamiento Latinoamericano - Series Especiales 2 (1): 61-85. [ Links ]
34. Samec, C., M. Morales y H. Yacobaccio 2013 Exploring Human Subsistence Strategies and Environmental Change through Stable Isotopes in the Dry Puna of Argentina. International Journal of Osteoarchaeology 24 (2): 134-148. [ Links ]
35. Samec, C. T., H. D. Yacobaccio y H. O. Panarello 2015 Carbon and nitrogen isotope composition of natural pastures in the dry Puna of Argentina: a baseline for the study of prehistoric herd management strategies. Archaeological and Anthropological Sciences 1-11. DOI:10.1007/s12520-015-0263-2 [ Links ]
36. Szpak, P., C. White, F. Longstaffe, J. Millaire y V. Vásquez Sánchez 2013 Carbon and nitrogen isotopic survey of northern Peruvian plants: baselines for paleodietary and paleoecological studies. PLoS One 8, e53763. [ Links ]
37. Szpak, P., J. F. Millaire, C. D. White y F. Longstaffe 2014 Small scale camelid husbandry on the north coast of Peru (Virú Valley): Insight from stable isotope analysis. Journal of Anthropological Archaeology 36: 110-129. [ Links ]
38. Tchilinguirian, P. y D. Olivera 2009 Paleoambientes y geoarqueología: sitios Corral Grande y Arroyo Las Pitas. Antofagasta de la Sierra (Prov. De Catamarca, Puna Meridional Argentina). Arqueometría Latinoamericana (2do.Congreso Nacional de Arqueometría y 1ro. Latinoamericano), 2: 544- 553. CONEA, Buenos Aires, Argentina. [ Links ]
39. Tessone, A. 2010 Arqueología y Ecología Isotópica. Estudio de isótopos estables de restos humanos del Holoceno tardío en Patagonia meridional. Tesis Doctoral inédita. Facultad de Filosofía y Letras, Universidad de Buenos Aires, Buenos Aires. [ Links ]
40. Thornton, E. K., S. D. Defrance, J. Krigbaum y P. R. Williams 2011 Isotopic Evidence for Middle Horizon to 16th Century Camelid Herding in the Osmore Valley, Peru. International Journal of Osteoarchaeology 21: 544-567. [ Links ]
41. Tykot, R. H. 2004 Stable Isotopes and Diet: You Are What You Eat. En Physics Methods in Archaeometry, Proceedings of the International School of Physics "Enrico Fermi" Course CLIV, editado por M. Martini, M. Milazzo y M. Piacentini, pp. 433-444. Società Italiana di Fisica, Bologna. [ Links ]
42. Yacobaccio, H. D., C. M. Madero, M. P. Malmierca y M. Del C. Reigadas 1997 Isótopos estables, dieta y estrategia de pastoreo. Arqueología 7: 105-109. [ Links ]
43. Yacobaccio, H. D., C. M. Madero y M. P. Malmierca 1998 Etnoarqueología de pastores surandinos. Grupo Zooarqueología de Camélidos, Buenos Aires. [ Links ]
44. Yacobaccio H. D., M. R. Morales y C. T. Samec 2009 Towards an isotopic ecology of herbivory in the Puna ecosystem: new results and patterns in Lama glama. International Journal of Osteoarchaeology 19 (2): 144-155. [ Links ] [ Links ]

