










Serviços Personalizados
Journal

Artigo
 Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares em
SciELO
Similares em
SciELO
Compartilhar

 Permalink
PermalinkIntersecciones en antropología
versão On-line ISSN 1850-373X
Intersecciones antropol. vol.17 no.3 Olavarría dez. 2016
ARTÍCULOS
Análisis tafonómico de una asociación faunística generada por pequeños carnívoros de la Región Pampeana
Cristian A. Kaufmann
Consejo Nacional de Investigaciones Científicas y Técnicas (CONICET)-Instituto de Investigaciones Arqueológicas y Paleontológicas del Cuaternario Pampeano (INCUAPA). Facultad de Ciencias Sociales, UNICEN. Av. Del Valle 5737 (B7400JWI). Olavarría, Buenos Aires, Argentina. E-mail: ckaufman@soc.unicen.edu.ar
Recibido 31 de agosto 2015.
Aceptado 28 de abril 2016
RESUMEN
El objetivo del presente trabajo es caracterizar tafonómicamente al conjunto óseo formado por restos de presa que fueron depositados acrecionalmente por Lycalopex gymnocercus y otros pequeños carnívoros de la Región Pampeana. Se realizó la determinación taxonómica de la totalidad de los restos y se profundizó en el análisis tafonómico de liebres y armadillos. Las variables evaluadas para describir el procesamiento de los carnívoros fueron la representación anatómica, el grado de completitud, la fragmentación ósea y la diversidad e intensidad de improntas de origen carnívoro. Se recuperó un total de 1266 restos óseos no ingeridos en los cuales dominaban los restos de armadillos (Chaetophractus villosus y Dasypus hybridus). También fueron identificados restos de Lepus europaeus, Lagostomus maximus, Lycalopex gymnocercus y Bos taurus. La información del presente trabajo aporta nuevo conocimiento para establecer el origen de mamíferos medianos en los sitios arqueológicos y da la posibilidad de brindar mayor solidez a los modelos de subsistencia humana.
Palabras clave: Tafonomía actualística; Región Pampeana; Armadillos; Zorro pampeano; Liebre.
ABSTRACT
Taphonomic analysis of a faunal assemblage generated by small carnivores of the Pampas Region
The objective of this study is to taphonomically characterize an assemblage of prey remains that were deposited by Pampas fox and other small carnivores of the Pampas region. Taxonomic identification was preformed for all the remains, and detailed taphonomic analysis was realized on hares and armadillos. The variables evaluated to describe the processing of carnivores were the anatomical representation, the degree of completeness, bone fragmentation and diversity and intensity of carnivore marks. A total of 1266 uneaten skeletal remains were recovered, the majority of which correspond to armadillos (Chaetophractus villosus and Dasypus hybridus). Other species identified were Lepus europaeus, Lagostomus maximus, Lycalopex gymnocercus and Bos taurus. The information in this work provides new knowledge to help establish the origin of medium sized mammals in archaeological sites and the possibility of providing more robust models of human subsistence.
Keywords: Actualistic taphonomy; Pampas region; Armadillos; Pampas fox; Hare.
INTRODUCCIÓN
Esta investigación se enmarca en un proyecto cuyo objetivo es la construcción de modelos tafonómicos que permitan identificar y dimensionar el accionar de predadores sobre el registro arqueológico de la Región Pampeana. Hasta el momento se han realizado experimentos en ambientes controlados con diversos carnívoros (Álvarez et al. 2012; Massigoge et al. 2014; Rafuse et al. 2014) y resulta necesario complementar estos resultados con estudios naturalistas. En el presente trabajo se caracteriza tafonómicamente un conjunto óseo formado por restos de presa no ingeridos que fueron depositados en forma acrecional por zorros pampeanos (Lycalopex gymnocercus) y otros pequeños carnívoros en el contexto de una madriguera localizada entre los restos de un antiguo puente a orillas del arroyo Salado, partido de General Lamadrid, provincia de Buenos Aires, Argentina (Figura 1A). El zorro es un pequeño cánido que posee una dieta generalista que incluye pequeños roedores, aves, reptiles, insectos, frutos y carroña (Redford y Eisenberg 1992). En el ámbito rural es habitual que los zorros se acerquen a los establecimientos humanos para carroñar sobre desechos descartados en basureros y fogones. Esta característica etológica hace de estos cánidos agentes tafonómicos destacados en el proceso de formación de los conjuntos óseos de los sitios arqueológicos. Consecuentemente, numerosos trabajos actualísticos realizados en la Argentina han caracterizado los patrones de depósito, sustracción y modificación ósea que producen estos carnívoros sobre mamíferos de variados tamaños (Mondini 1995, 2012; Borrero y Martin 1996; Nasti 1996; Martin 1998; Mammeli y Estévez 1999-2001; Elkin y Mondini 2001; Kaufmann y Messineo 2002; Gómez y Kaufmann 2007; Fernández et al. 2010; Massigoge et al. 2014; Rafuse et al. 2014).

Figura 1. A) La flecha indica el lugar donde se encontraba la madriguera en cercanías del arroyo Salado; B) fecas de felidae depositadas en una letrina ubicada en una oquedad del suelo; C) fecas de felidae y canidae halladas sobre la madriguera.
En el presente trabajo se estudia la acción del zorro promediada con la de otros predadores, como felinos y, probablemente, mustélidos. Sectores protegidos del paisaje como abrigos rocosos, matorrales, vizcacheras abandonadas o huecos en las barrancas de ríos y lagunas reúnen condiciones bajo las cuales se pueden esperar este tipo de palimpsestos tafonómicos. Estos emplazamientos en ocasiones también fueron ocupados por poblaciones humanas, lo que daría lugar a una mezcla de materiales de origen antrópico y restos de presas depositadas por diversos predadores.
MATERIALES Y METODOLOGÍA
Los restos de presa estudiados fueron recolectados en seis trabajos de campo realizados entre los años 2010 y 2013 en una madriguera situada entre los desechos materiales de un puente demolido sobre la margen izquierda del arroyo Salado, partido de General Lamadrid. Este refugio fue ocupado temporalmente por una pareja de zorros y posiblemente funcionó como madriguera reproductiva. En el sector también fueron observadas áreas de letrinas (Figuras 1B y 1C) que, por el estado y características de las fecas, corresponderían a otros carnívoros que las utilizaron de manera alterna a los zorros. Con el fin de obtener mayor información respecto de los probables predadores involucrados en la acumulación ósea, la Dra. Estela Luengos Vidal llevó a cabo el examen visual de las fecas, y las asignó a pequeños carnívoros de la familia canidae y felidae. En suma, si se tiene en cuenta los predadores que habitan el área y la información contextual disponible, se propone que la acumulación ósea que se analiza fue generada acrecionalmente por zorro pampeano y otros pequeños carnívoros, como gato montés (Leopardus geoffroyi) y mustélidos.
Los restos óseos recolectados fueron categorizados como "huesos desarticulados" (cuando un elemento óseo no está relacionado a otro por tejido blando o por su posición anatómica), "restos articulados" (dos o más huesos unidos por tejido blando o en relación anatómica, involucrando a menos del 75% del esqueleto de un animal) y "carcasa" (más del 75% de los elementos esqueletarios de un animal unidos por tejido blando o en relación anatómica) (Massigoge et al. 2015). Con el objeto de estimar la abundancia relativa de los distintos taxones que integran el conjunto, se realizó la determinación anatómica y taxonómica de los restos y se calculó el número de especímenes óseos identificados por taxón (NISP) y el número mínimo de individuos (NMI). Esta última medida da cuenta de la totalidad de huesos identificados para un determinado taxón y se obtuvo a través de la frecuencia que le corresponde al elemento anatómico más abundante en la muestra y su lateralidad (Andrews 1990). Posteriormente, se caracterizó el procesamiento que los carnívoros realizaron sobre restos no ingeridos de lagomorfos y armadillos, las especies que estaban mejor representadas en el sitio. Las variables tafonómicas consideradas fueron la representación anatómica, el grado de completitud, la fragmentación ósea y la diversidad e intensidad de marcas de origen carnívoro. La diversidad de partes esqueletarias de ambos taxones se estimó a partir del número mínimo de elementos (NME) y de la abundancia relativa (Ri) (Andrews 1990). Para obtener el Ri, se consideró la relación proporcional entre el número de veces que un elemento determinado está representado en la muestra estudiada y el número esperado de este elemento (Ei) en relación con el número mínimo de individuos (NMI): Ri = (Ni/NMIxEi) (Andrews 1990: 45). Con el fin de caracterizar los patrones de procesamiento empleados por los predadores, se estimaron varios índices que permiten evaluar la variación de las proporciones de sobrevivencia de los distintos segmentos esqueletarios. Durante la limpieza de los restos se observaron varios casos en que los dientes se desprendieron de sus alveolos por la remoción del tejido blando; por este motivo, se decidió no utilizar índices que evaluaran y/o consideraran la pérdida de molares e incisivos.
Los índices utilizados fueron los siguientes:
a) PCRT/CR: proporción de elementos craneales con relación a los elementos poscraneales. En armadillos y liebres: [(fémur + húmero) / (mandíbula + maxila)] x 100 (Andrews 1990).
b) C/E: proporción de elementos distales con relación a elementos proximales. En armadillos y liebres: [(tibia + ulna) / (fémur + húmero)] x 100 (Andrews 1990).
c) AN/PO: proporción entre la extremidad anterior y posterior. Liebres: [(húmero + radio + metacarpo) / (fémur + tibia + metatarso)] x 100 (modificado de Lloveras et al. [2008]). Armadillos: [(húmero + ulna) / (fémur + tibia)] x 100.
d) AX/AP: relación entre el esqueleto poscraneal axial y apendicular. Liebres: [(Vértebras + costillas) x 101 / (húmero + ulna + radio + fémur + tibia) x 662] x 100 (Álvarez et al. 2012). Armadillos: [(Vértebras + costillas) x 103 / (húmero + ulna + radio + fémur + tibia) x 594] x 100.
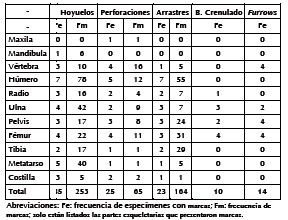
La integridad de los especímenes fue relevada teniendo en cuenta las categorías de fragmentación establecidas por Lloveras et al. (2008). También se estimó el porcentaje de elementos que sobrevivieron completos (C%) para cada unidad anatómica del esqueleto. Este porcentaje se calculó tomando en consideración el número de especímenes completos que se registró para cada unidad anatómica en relación con el NISP total que presentaba el elemento. Por último, el análisis de las marcas se realizó mediante una lupa binocular (Motic ST-39) con aumentos de X20 y X40. Las marcas fueron caracterizadas a partir de los tipos definidos por Binford (1981): perforaciones, hoyuelos, arrastres, bordes crenulados y furrows. Los hoyuelos, perforaciones y arrastres fueron contabilizados para obtener la frecuencia con que se hacen presentes en los dos taxones analizados.
Las proporciones de estas marcas fueron ploteadas en gráficos tripolares, para lo cual se empleó el programa Triangle modified elaborado por Steele y Weaver (2002), que permite representar los porcentajes de las tres categorías de marcas en un punto que se encuentra rodeado por un área circular que representa un nivel de confiabilidad del 95%, calculado en función del tamaño de la muestra analizada.
RESULTADOS
La acumulación ósea estaba compuesta por 1266 restos óseos no ingeridos de diversas especies, conformados por una carcasa (NR: 80), 43 conjuntos de restos articulados (NR: 907) y 279 restos desarticulados. De las porciones articuladas, 36,6% corresponden a segmentos de la columna vertebral, 26,8% son extremidades delanteras, 21,9% extremidades traseras, 9,7% son elementos del autopodio y el 4,9% corresponde a cráneo y mandíbula. Los resultados del análisis taxonómico de los restos informan que las categorías mejor representadas son los armadillos, conformada por 33 especímenes de dasypodidae, 286 especímenes de Dasypus hybridus y 167 de Chaetophractus villosus. También fueron identificados restos de Lepus europaeus (NISP = 281), Lagostomus maximus (NISP = 103), Lycalopex gymnocercus (NISP = 175) y Bos taurus (NISP = 99). Asimismo, se recuperó un fragmento aislado del neurocráneo de Rhamdia sapo., una hemimandíbula de un carnívoro nonato y 120 restos indeterminados (Tabla 1).
Tabla 1. Taxones recuperados en la madriguera con referencia de NISP, NISP% y NMI.

Los armadillos
Se recuperó un total de 486 especímenes óseos que conforman 436 elementos asignados a dasipódidos (Tabla 2). A partir de las corazas, que en su mayoría se encontraban completas, se calculó un NMI de 9. El promedio de abundancia relativa que generaron los carnívoros sobre esta familia fue de 23,3%, lo que muestra que los predadores involucrados produjeron una significativa destrucción ósea de los armadillos.
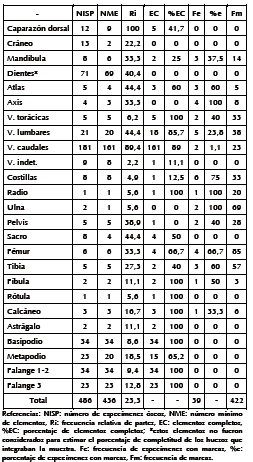
Tabla 2. Medidas de abundancia relativa y daños generados por carnívoro en los restos de armadillos.
La frecuencia relativa de partes esqueletarias
Las vértebras caudales, junto con el escudo dorsal, son los elementos con frecuencias relativas más elevadas en los armadillos (Figura 2). Por otro lado, se observó una muy baja frecuencia de las extremidades delanteras. La escápula y el húmero no están presentes, y el radio y la ulna poseen baja supervivencia. Los miembros traseros, fémur y tibia exhiben una frecuencia media, en tanto que las partes bajas de las patas (autopodio) se encuentran subrepresentadas. El cráneo, mandíbula, axis y atlas poseen frecuencias medias-bajas, y las cervicales, torácicas y costillas poseen una representación baja. Las vértebras lumbares, sacro y pelvis tienen una representación media.

Figura 2. Corazas articuladas a tubos caudales de armadillos recuperados en el contexto de la madriguera. Las dos corazas de la izquierda poseen la pelvis y algunas vértebras articuladas en el interior.
Proporción de los segmentos anatómicos
Al evaluar la proporción de los segmentos anatómicos a través de algunos índices (Tabla 3) se observa que existe una leve sobrerrepresentación de los elementos craneales con respecto al apendicular (fémur y húmero). El índice que considera la proporción de elementos del estilopodio (fémur-húmero) en relación con el cigopodio (tibia y ulna) indica que ambos segmentos se encuentran representados de manera proporcional. Por otro lado, se observa que las extremidades posteriores están mejor representadas que las anteriores. El índice que evalúa el esqueleto axial poscraneal y el apendicular (AX/AP) indica una mejor representación del esqueleto axial poscraneal, resultado que se encuentra influenciado por la alta supervivencia de las vértebras caudales. El porcentaje de elementos completos general fue de 66,2% y la completitud porcentual (C%) para los huesos largos (excluyendo los metacarpos y metatarsos), del 56,25%. Considerando la C% de cada elemento, se observa que todos los huesos del autopodio, falanges y rótula están completos. De manera contraria, no existe ningún elemento completo de cráneo, vértebra axis y pelvis. Las vértebras que lograron sobrevivir poseen una completitud alta; y los huesos largos, una completitud media y alta (Tabla 2). Se registraron fracturas óseas sobre 49 especímenes (10%); la mayor frecuencia corresponde a aquellas de tipo irregular (n = 43), seguidas por las de tipo longitudinal y transversa (n = 2 en ambos casos) y, por último, un único caso de fractura oblicua (n = 1) y helicoidal. Un 63,2% (n = 31) de los huesos fracturados poseen asociadas marcas de carnívoros.
Tabla 3. Resultados de los índices de representación esqueletaria de liebres y armadillos.

Frecuencia y tipo de marcas
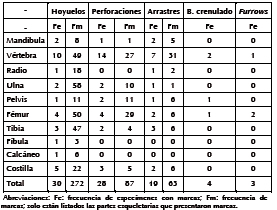
Un 8% (n = 39) de los especímenes analizados presentan modificaciones asignadas a carnívoro. De estos, 30 (35,7%) evidencian hoyuelos; 28 (33,3%), perforaciones (Figuras 3A y 3B); 9 (22,6%), arrastres; 4 (4,8%), bordes crenulados; y 3 (2,4%), furrows. El total de marcas asciende a 422, y las más frecuentes fueron los hoyuelos (n = 272; 63%) y las perforaciones (n = 87; 20%), seguidas por los arrastres (n = 63; 15%) (Tabla 4). Los escudos dorsales, cráneo, sacro, rótula, metapodios, basipodio y falanges no exhibieron marcas, y sólo el 1% de las vértebras caudales se encontraban afectadas. El porcentaje de especímenes dañados correspondientes a mandíbula, vértebras torácicas, lumbares, pelvis, y calcáneo fue moderado. En tanto que los huesos que en su mayor porcentaje presentaron marcas de carnívoro fueron el atlas, axis, costillas, radio, ulna, fémur, tibia y fíbula. De estos huesos, la ulna, radio, fémur y tibia fueron los que exhibieron mayor frecuencia de daños (Tabla 2).
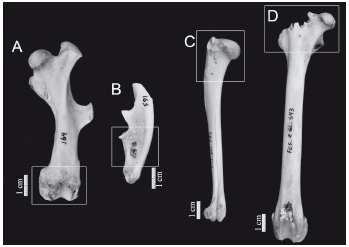
Figura 3. A) Perforaciones en epífisis distal de fémur de Chaetophractus villosus; B) perforaciones en diáfisis medial de ulna de Chaetophractus villosus; C) perforaciones en epífisis proximal de húmero de Lepus europaeus; y D) perforaciones en epífisis proximal de fémur de Lepus europaeus.
Tabla 4. Tipo y frecuencia de marcas que presentan los huesos de armadillos.
Las liebres
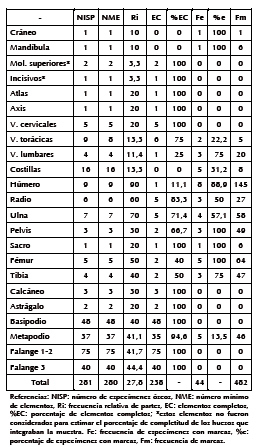
Se recuperó un total de 281 especímenes óseos, que conforman 280 elementos (Tabla 5). A partir del húmero, se calculó un número mínimo de cinco individuos. El promedio de abundancia relativa generado por los carnívoros para este taxón fue de 27,8%, lo cual indica que predadores como zorro pampeano y otros pequeños carnívoros de la región pampeana ocasionan una destrucción esqueletaria significativa de las liebres.
Tabla 5. Medidas de abundancia relativa y daños generados por carnívoro en los restos de liebres.
La frecuencia relativa de partes esqueletarias
Las porciones del esqueleto mejor representadas corresponden a las extremidades delanteras y traseras. La cintura pélvica está medianamente representada, mientras que la escápula está ausente. El esqueleto axial se encuentra, en general, pobremente representado (Tabla 5). La proporción de los distintos segmentos anatómicos evidencia que existe una mayor representación de los elementos apendiculares (fémur y húmero) con respecto a la maxila y mandíbula (Tabla 3). El índice que considera la frecuencia del estilopodio (fémurhúmero) en relación con el cigopodio (tibia y ulna) indica una proporción pareja con una leve sobrerrepresentación del estilopodio. Por otro lado, las extremidades anteriores están mejor representadas que las posteriores. El índice que evalúa el esqueleto axial poscraneal y el apendicular (AX/AP) indica una mejor representación de las extremidades. El porcentaje de elementos completos general fue de 84,7% (n = 238) y la completitud porcentual (C%) para los huesos largos (excluyendo los metacarpos y metatarsos), de 48,4%. Se observa que todos los huesos del autopodio, falanges, vértebras sacras y cervicales poseen una completitud del 100%. Contrariamente, no existe ningún elemento completo de cráneo, mandíbula, costillas y huesos largos (Tabla 5). La mayor frecuencia de fracturas corresponde a aquellas de tipo irregular (n = 39), seguidas por las de tipo helicoidal (n = 3) y, por último, un único caso de fractura oblicua (n = 1). El 67,4% (n = 29) de los huesos fracturados posee modificaciones generadas por carnívoros.
Frecuencia y tipo de marcas
De los 281 especímenes analizados, 44 (15,6%) presentan daños asignados a carnívoro. De estos, 35 (34,6%) evidencian hoyuelos; 25 (24,7%), perforaciones (Figuras 3C y 3D); 23 (22,8%), arrastres; 10 (9,9%), bordes crenulados; y 14 (13,9%), furrows (Tabla 6). Se contabilizaron 482 marcas; las más frecuentes son los hoyuelos (n = 253; 52,4%) y arrastres (n = 164; 34,1%), seguidos por las perforaciones (n = 65; 13,5%) (Tabla 6). Los dientes, las vértebras cervicales (atlas y axis incluidos), huesos del basipodio y falanges no presentan marcas, en tanto que sí lo hacen en un moderado porcentaje las costillas, vértebras torácicas y metapodios. Por último, los huesos que fueron dañados en mayor porcentaje son los cráneos, mandíbulas, vértebras lumbares, pelvis, sacro, húmero, ulna, radio, fémur y tibia. De estos últimos, húmero, ulna, pelvis, fémur y tibia también muestran una mayor frecuencia de marcas (Tabla 5).
Tabla 6. Tipo y frecuencia de marcas que presentan los huesos de liebres.
DISCUSIÓN
Supervivencia ósea de armadillos y liebres El cálculo de las medidas de abundancia taxonómica (NMI y NISP) informa que los armadillos y liebres son las presas más abundantes entre los restos no ingeridos recuperados en el contexto de la madriguera. La supervivencia del esqueleto de estos taxones fue media-baja. Las unidades óseas ausentes o poco representadas, como la escápula, costillas y vértebras, habrían sido en gran medida destruidas o ingeridas por los predadores. Experimentaciones realizadas con gato montés y zorro pampeano evidenciaron que estos pequeños carnívoros tienen la capacidad de masticar y destruir vértebras y costillas de presas medianas (1 a 10 kg) (Álvarez et al. 2012; Rafuse et al. 2014). Los estudios de lince ibérico muestran cierta complementariedad entre la frecuencia de elementos que aparecen en muestras no ingeridas y fecas (Rodríguez-Hidalgo et al. 2013). Sin embargo, estos autores mencionan que algunos restos, como los dientes, tienen la particularidad de aparecer en ambos tipos de registro. La supervivencia de estos elementos estaría relacionada con la alta densidad mineral (Pavao y Stahl 1999), que los hace resistentes a procesos atricionales como la digestión. Otros elementos, como vértebras y costillas, serían consumidos en gran medida por los predadores, pero la sobrevivencia ante procesos como la masticación y digestión gástrica sería baja (Rodríguez-Hidalgo et al. 2013).
El estudio de los restos muestra que existen algunas diferencias en la representación esqueletaria de liebres y armadillos que merecen ser señaladas. El conjunto asignado a liebre está dominado por el esqueleto poscraneal, con una subrepresentación de vértebras y un leve dominio del miembro anterior, en tanto que el cigopodio se encuentra levemente subrepresentado con respecto a los elementos proximales. La abundancia de extremidades sobre el resto del esqueleto también ha sido observada en muestras provenientes de una madriguera de zorro colorado (Lycalopex culpaeus) en el norte de la Patagonia Andina (Fernández et al. 2010) y en muestras experimentales realizadas con zorro pampeano y gato montés (Álvarez et al. 2012; Massigoge et al. 2014; Rafuse et al. 2014). La supervivencia del esqueleto en la muestra ósea analizada, en términos generales, es similar a las muestras experimentales, con una mayor similitud a la generada por zorro pampeano. Esta evidencia por el momento no es suficiente para otorgar un papel preponderante a este cánido con respecto a otros carnívoros, ya que resulta necesario contar con mayor número de muestras experimentales y naturalistas para evaluar la variabilidad intrarreferencial, que podría manifestarse bajo diversas condiciones ecológicas (e.g., Saavedra y Simonetti 1998; Cochard 2008). Los armadillos se caracterizan por poseer un escudo dorsal, uno encefálico y una armadura caudal que protege a sus cuerpos de los predadores (Soibelzon et al. 2013). El estudio realizado sobre los huesos de armadillos informa que la protección que brindan estas estructuras óseas a los xenartros también condicionaría la modalidad e intensidad de procesamiento que los predadores realizan sobre las carcasas de estos animales. El elemento más representado en los armadillos corresponde a los escudos dorsales, los cuales en muchas ocasiones contenían en su interior otros elementos que forman parte del esqueleto de estos mamíferos. Hay que considerar que las corazas recuperadas no fueron modificadas por los carnívoros y permanecieron por un corto tiempo expuestas ante las condiciones atmosféricas, situación que favoreció su integridad. Se estima que, transcurrido cierto tiempo, estas corazas perderían cohesión y se desarticularían en cientos de placas dérmicas. Otros elementos que poseen alta frecuencia relativa son las vértebras caudales, las cuales ingresaron a la madriguera articuladas y protegidas por las placas dérmicas que integran el tubo caudal. Esta porción posee asociado muy poco tejido blando y no resulta atractiva a los predadores, aspecto que se manifiesta en una alta supervivencia y una baja frecuencia de daños.
En suma, se puede sostener que las liebres y armadillos depositados en el contexto de la madriguera representan modelos de supervivencia ósea característicos de la etapa final de la trayectoria generada por el zorro pampeano y otros pequeños carnívoros, como gato montés y mustélidos. Las mulitas, con una masa corporal promedio de 1500 gramos, constituirían el umbral de tamaño cuyos restos esqueletarios sobrevivirían, de modo variable, a la ingesta que realizan estos predadores sobre sus presas. Sólo se recuperaron tres especímenes no ingeridos de vertebrados inferiores a Dasypus hybridus, lo que sugiere que las presas pequeñas serían consumidas completas. Del mismo modo, Mondini (2012: 74-76), en su modelo sobre la acción tafonómica de los mamíferos carnívoros en la puna argentina, plantea que los taxones pequeños habitualmente son ingeridos completos e ingresarían a los sitios casi exclusivamente a través de excrementos. Daño y completitud de los especímenes óseos La extensión del daño medido a través del porcentaje de especímenes afectados por los carnívoros fue menor en armadillos (8%) que en las liebres (15,6%), aspecto quizá relacionado con el acceso más restringido que tienen los carnívoros a determinadas porciones del esqueleto axial de los armadillos. Las porciones esqueletarias modificadas con mayor frecuencia en liebre fueron la cintura escapular y pélvica; y en armadillos, las vértebras atlas, axis, la tibia, el fémur, las costillas, el radio y la ulna. El autopodio presentó invariablemente muy baja frecuencia de modificaciones para los dos taxones analizados. Los huesos que presentaron mayor frecuencia de marcas en liebre son aquellos que están asociados con abundante carne, como pelvis, húmero y fémur. En los xenartros, en cambio, la porción más afectada fue el cigopodio, luego continúa en orden de importancia el estilopodio, la cintura pélvica, el esqueleto axial y, por último, con muy baja frecuencia, las vértebras caudales. Estos datos evidencian, como se propuso anteriormente, que la intensidad de daño estaría influenciada en cierta medida por la estructura ósea particular de los armadillos, en la cual la pelvis, las vértebras lumbares y vértebras caudales se encuentran menos accesibles al mordisqueo que realizan los carnívoros. El menor daño relativo que presenta la cintura pélvica parecería ser una condición característica de los armadillos; así lo sugieren estudios realizados con diversos predadores y presas, que informan que esta porción esqueletaria se encuentra generalmente entre las más afectadas (Borrero et al. 2005; Muñoz et al. 2008; Álvarez et al. 2012; Rodríguez-Hidalgo et al. 2013; Kaufmann et al. 2016).
El tipo de daño más frecuente corresponde a hoyuelos, perforaciones y arrastres, en tanto que la presencia de bordes crenulados y furrows es baja. La proporción de marcas representadas en el gráfico ternario (Figura 4) muestra que existe un agrupamiento de las muestras de campo de liebre y experimentales de conejos procesados por gato montés y zorro pampeano. La muestra de armadillos se diferencia sutilmente del resto. El agrupamiento se caracteriza por una proporción alta de hoyuelos con porcentajes que van entre 54 y 62%. Luego continúan en orden de importancia los arrastres, con porcentajes entre 26 y 37%, y por último, las perforaciones, con porcentajes entre 6 y 15%. La muestra de armadillos difiere de las anteriores al presentar en segundo orden de importancia a las perforaciones, con un 21%, y los arrastres, con 15%. Los elementos óseos analizados en ambos taxones presentan porcentajes de completitud altos (entre 66,2% y 84,7%). Los especímenes que generalmente permanecieron completos son huesos asociados a bajo contenido cárnico, como vértebras cervicales, caudales, carpo/tarso y falanges. Por el contrario, los especímenes incompletos son huesos que poseen porciones poco densas, morfología plana, asociados con alto contenido proteico, como cráneo, costillas y húmero. La completitud de los huesos largos varió entre 48,4% y 57,1%. Este resultado informa que los carnívoros involucrados en el presente estudio tuvieron un interés limitado en la grasa intraósea y medular de las presas medianas, aspecto que deberá ser evaluado en otros escenarios ecológicos.
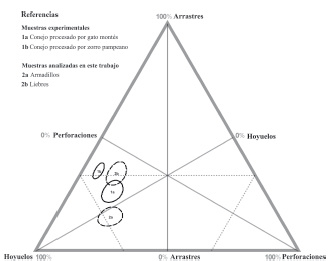
Figura 4. Gráfico tripolar en el cual se observa la proporción de hoyuelos, perforaciones y arrastres en muestras experimentales y de campo.
CONCLUSIONES
Como se evidencia en el presente trabajo, los pequeños carnívoros reutilizan en forma frecuente las letrinas y madrigueras ocupadas anteriormente por otros predadores (Soler et al. 2009). Es esperable que algunos sitios de la Región Pampeana –principalmente aquellos protegidos y con disponibilidad de recursos– hayan sido ocupados de manera alternativa por distintos carnívoros. En muchas ocasiones, estos conjuntos se superpondrían con ocupaciones humanas, lo que conformaría, en definitiva, interesantes ensambles óseos con historias tafonómicas complejas y una rica información paleoecológica.
Las presas más abundantes en la muestra estudiada fueron Chaetophractus villosus y Dasipus hybridus. Estos mamíferos cavadores son registrados sistemáticamente en sitios arqueológicos de la Región Pampeana (Quintana y Mazzanti 2001; Frontini y Deschamps 2007; Álvarez 2009; Messineo et al. 2014; Alcaraz 2015, entre otros) y la naturaleza de su origen en muchas oportunidades es incierta. A pesar de la relevancia de esta problemática, sólo en los últimos años se han abordado desde un enfoque actualístico aspectos tafonómicos que involucran restos de dasipódidos (Frontini y Deschamps 2007; Frontini y Escoteguy 2012; Muñoz 2015). Cuando no existen evidencias claras de aprovechamiento humano de los armadillos, las muertes naturales en sus cuevas o la predación por carnívoros surgen como hipótesis potenciales de su presencia. Este trabajo aporta información novedosa sobre la representación esqueletaria y las modificaciones que pequeños carnívoros generan sobre carcasas de armadillos, aspecto que contribuirá a discutir el origen de sus restos en los sitios arqueológicos y brindará mayor solidez a los modelos de subsistencia humana.
Agradecimientos
Este estudio fue desarrollado dentro del programa de investigaciones INCUAPA-CONICET (Facultad de Ciencias Sociales, UNCPBA) y fue financiado por ANPCyT (PICT 08-814 y PICT 13-0199). Deseo agradecer a la Lic. Ana Alcaraz, a la Dra. Eugenia Conforti, a la Dra. Clara Álvarez y a la Dra. María Gutiérrez por la lectura crítica del manuscrito; a la Dra. Estela Luengo-Vidal por su aporte en la identificación de las fecas, a la Dra. Luciana Stoessel por su colaboración en la determinación de peces y al Dr. Daniel Rafuse por la traducción del resumen.
1 Este número corresponde a la frecuencia de húmeros, ulnas, radios, fémures y tibias que posee un esqueleto completo de liebre.
2 Este número corresponde a la frecuencia de costillas y vértebras que posee un esqueleto completo de liebre.
3 Este número corresponde a la frecuencia de húmeros, ulnas, radios, fémures y tibias que posee un esqueleto completo de armadillo.
4 Este número corresponde a la frecuencia de costillas y vértebras que posee un esqueleto completo de armadillo
REFERENCIAS CITADAS
1. Alcaraz, A. P. 2015 Primeros resultados de los análisis del conjunto de fauna menor del sitio arqueológico Zoko Andi 1 (Transición Pampeano-Patagónica Oriental, Provincia de Buenos Aires, Argentina). Archaeofauna 24: 119-133. [ Links ]
2. Álvarez, M. C. 2009 Análisis de los restos faunísticos del sitio Calera (Sierras Bayas, partido de Olavarría). Un aporte a su funcionalidad a través del estudio de los mamíferos. Relaciones de la Sociedad Argentina de Antropología XXXIV: 29-51. [ Links ]
3. Álvarez, M. C., C. A. Kaufmann, A. Massigoge, M. A. Gutiérrez, D. J. Rafuse, N. A. Scheifler y M. E. González 2012 Bone modification and destruction patterns of leporid carcasses by Geoffroy's cat (Leopardus geoffroyi): An experimental study. Quaternary International 278: 71-80. [ Links ]
4. Andrews, P. 1990 Owls, Caves and Fossils. University of Chicago Press, Chicago. [ Links ]
5. Binford, L. R. 1981 Bones. Ancient Men and Modern Myths. Academic Press, Nueva York. [ Links ]
6. Borrero, L. A. y F. M. Martín 1996 Tafonomía de carnívoros: un enfoque regional. En Arqueología solo Patagonia, editado por J. Gómez Otero, pp. 189-198. Centro Nacional Patagónico (CENPAT)-CONICET, Puerto Madryn. [ Links ]
7. Borrero, L. A., F. M. Martín y J. Vargas 2005 Tafonomía de la interacción entre pumas y guanacos en el Parque Nacional Torres del Paine, Chile. Magallania 33 (1): 95-114. [ Links ]
8. Cochard, D. 2008 Discussion sur la variabilité intraréférentiel d'accumulations osseuses de petits prédateurs. Annales de Paléontologie 94: 89-101. [ Links ]
9. Elkin, D. y M. Mondini 2001 Human and small carnivore gnawing damage on bones. An exploratory study and its archaeological implications. En Ethnoarchaeology of Andean South America: Contributions to Archaeological Method and Theory, editado por L. A. Kuznar, pp. 255-265. International Monographs in Prehistory, Ann Arbor. [ Links ]
10. Fernández, P. M., I. Cruz y A. Forlano 2010 Sitio 37: una madriguera de carnívoro en el norte de la Patagonia Andina (Cholila, Provincia de Chubut, Argentina). En Zooarqueología a principios del siglo XXI: aportes teóricos, metodológicos y casos de estudio, editado por M. A. Gutiérrez, M. De Nigris, P. M. Fernández, M. Giardina, A. F. Gil, A. Izeta, G. Neme y H. D. Yacobaccio, pp. 409-417. Libros del Espinillo, Buenos Aires. [ Links ]
11. Frontini, R. y C. Deschamps 2007 La actividad de Chaetophractus villosus en sitios arqueológicos. El Guanaco como caso de estudio. En Arqueología en las Pampas, vol. 1, editado por C. Bayón, A. Pupio, M. I. González, N. Flegenheimer y M. M. Frère, pp. 439-451. Sociedad Argentina de Antropología, Buenos Aires. [ Links ]
12. Frontini, R. y P. Escoteguy 2012 Chaetophractus villosus: a disturbing agent for archaeological contexts. International Journal of Osteoarchaeology 22 (5): 603-615. [ Links ]
13. Gómez, G. N. y C. A. Kaufmann 2007 Taphonomic analysis of Pseudalopex griseus (Gray, 1837) scat assemblages and their archaeological implications. Journal of Taphonomy 5 (2): 59-70. [ Links ]
14. Kaufmann, C. A. y P. Messineo 2002 Los desplazamientos óseos en carcasas de guanaco afectadas por la acción de zorros: un caso actual y sus implicancias arqueológicas. En Del mar a los salitrales, editado por D. Mazzanti, M. Berón y F. Oliva, pp. 419-429. Universidad Nacional de Mar del Plata, Mar del Plata. [ Links ]
15. Kaufmann, C. A., D. J. Rafuse, M. E. González, M. C. Álvarez, A. Massigoge, N. A. Scheifler y M. A. Gutiérrez 2016. Carcass utilization and bone modifications on guanaco killed by puma in northern Patagonia, Argentina. Quaternary International, Gary Haynes Commemorative. En prensa. [ Links ]
16. Lloveras, L., Moreno-García, M. y J. Nadal 2008 Taphonomic analysis of leporid remains obtained from modern Iberian Lynx (Lynx pardinus) scats. Journal of Archaeological Science 35: 1-13. [ Links ]
17. Mammeli, L. y J. Estévez 1999-2001 Seguimiento tafonómico de carcasas de camélidos, ovinos y bovinos en Tierra del Fuego (Argentina) e implicaciones arqueológicas. Xama 12-14: 87-106. [ Links ]
18. Martin, F. 1998 Madrigueras, dormideros y letrinas: Aproximación a la tafonomía de zorros. En Arqueología de la Patagonia Meridional. Proyecto Magallania, editado por L. A. Borrero, pp. 73-96. Búsqueda de Ayllu, Concepción del Uruguay. [ Links ]
19. Massigoge, A., M. A. Gutiérrez, M. C. Álvarez, C. A. Kaufmann, D. J. Rafuse y M. E. González 2014 Estudio comparativo de las marcas de dientes producidas por dos pequeños carnívoros sudamericanos. Revista Chilena de Antropología 30 (2): 42-49. [ Links ]
20. Massigoge, A., J. D. Rafuse, M. C. Álvarez, M. E. González, M. A. Gutiérrez, C. A. Kaufmann y N. Scheifler 2015 Beached penguins on the Atlantic Coast in the Pampas region of Argentina: Taphonomic analysis and implications for the archaeological record. Palaeogeography, Palaeoclimatology, Palaeoecology 436: 85-95. [ Links ]
21. Messineo, P. G., C. A. Kaufmann, P. G. Steffan, C. F. Dubois y N. Pal 2014 Ocupaciones humanas en un valle intraserrano del sector noroccidental de Tandilia: sitio El Puente (partido de Olavarría, Buenos Aires). Relaciones de la Sociedad Argentina de Antropología XXXIX (2): 435-462. [ Links ]
22. Mondini, M. 1995 Artiodactyl prey transport by foxes in Puna rock shelters. Current Anthropology 36: 520-524. [ Links ]
23. Mondini, M. 2012 Tafonomía de carnívoros en los Andes Centro- Sur. Madrigueras actuales y sus implicaciones para el registro arqueofaunístico. En Temas de arqueología. Estudios tafonómicos y zooarqueológicas (II), editado por A. Acosta, D. Loponte y L. Mucciolo, pp. 67-105. Instituto Nacional de Antropología y Pensamiento Latinoamericano, Buenos Aires. [ Links ]
24. Muñoz, S. A. 2015 Observaciones tafonómicas naturalistas sobre restos de armadillos (Dasypodidae) en Patagonia meridional. Archaeofauna (24): 239-251. [ Links ]
25. Muñoz, A. S., M. Mondini, V. Durán y A, Gasco 2008 Los pumas (Puma concolor) como agentes tafonómicos. Análisis actualístico de un sitio de matanza en los Andes de Mendoza, Argentina. Geobios 41: 123-131. [ Links ]
26. Nasti, A. 1996 Predadores, carroñeros y huesos: la acción del puma y el zorro como agentes modificadores de esqueletos de ungulados en la Puna meridional, Argentina. En II Reunión de Tafonomía y Fosilización, editado por G. Meléndez Hevia, M. F. Blasco Sancho e I. Pérez Urresti, pp. 265-269. Institución Fernando el Católico, Zaragoza. [ Links ]
27. Pavao, B. y P. W. Stahl 1999 Structural density assays of leporid skeletal elements with implications for taphonomic, actualistic and archaeological research. Journal of Archaeological Science 26: 53-66. [ Links ]
28. Quintana, C. y D. Mazzanti 2001 Selección y aprovechamiento de recursos faunísticos. En Cueva Tixi: cazadores y recolectores de las sierras de Tandilia Oriental. 1 Geología, paleontología y zooarqueología, editado por D. Mazzanti y C. Quintana, pp. 181-209. Laboratorio de Arqueología, Facultad de Humanidades, Universidad Nacional de Mar del Plata, Mar del Plata. [ Links ]
29. Rafuse, D. J., M. E. González, C. A. Kaufmann, M. C. Álvarez, M. A. Gutiérrez y A. Massigoge 2014 Análisis comparativo de los patrones de modificaciones óseas de dos carnívoros sudamericanos: el gato montés (Leopardus geoffroyi) y el zorro pampeano (Lycalopex gymnocercus). Aportes para la identificación de la acción de pequeños carnívoros en el registro arqueológico. Magallania 42 (1): 167-186. [ Links ]
30. Redford K. H y J. F. Eisenberg 1992 Mammals of the Neotropics. The Southern Cone, vol. 2. University of Chicago Press, Chicago y Londres. [ Links ]
31. Rodríguez-Hidalgo, A., L. Lloveras, M. Moreno-García, P. Saladié, A. Canals y J. Nadal 2013 Feeding behaviour and taphonomic characterization of non-ingested rabbit remains produced by the Iberian lynx (Lynx pardinus). Journal of Archaeological Science 40: 3031-3045. [ Links ]
32. Saavedra, B. y J. Simonetti 1998 Small Mammal Taphonomy: Intraspecific Bone Assemblage Comparison between South and North American Barn Owl, Tyto alba, Populations. Journal of Archaeological Science 25: 165-170. [ Links ]
33. Soibelzon, E., M. Medina y A. M. Abba 2013 Late Holocene armadillos (Mammalia, Dasypodidae) of the Sierras of Córdoba, Argentina: Zooarchaeology, diagnostic characters and their paleozoological relevance Quaternary International 299: 72-79. [ Links ]
34. Soler, L., M. Lucherini, C. Manfredi, M., Ciuccio y E. M. Casanave 2009 Characteristics of defecation sites of the Geoffroy's cat Leopardus geoffroyi. Mastozoología Neotropical 16 (2): 485-489. [ Links ]
35. Steele, T. E. y T. D. Weaver 2002 The modified triangular graph: A refined method for comparing mortality profiles in archaeological samples. Journal of Archaeological Science 29: 317-322. [ Links ]

