











 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
PermalinkINTRODUCCIÓN
El registro arqueobotánico ofrece evidencia sólida de que la recolección de plantas cumplió un papel importante en la economía de las poblaciones cazadoras-recolectoras de Patagonia (Pérez de Micou 1991; Crivelli et al. 1996; Pérez y Erra 2011; Caruso 2012; Lema et al. 2012; Capparelli y Prates 2015; Belmar et al. 2017; Ciampagna y Mange 2017). Estos antecedentes en escala macrorregional suministran un valioso aporte para la interpretación local del registro de Cueva Huenul 1 (CH1 de aquí en más).
El presente artículo muestra los hallazgos arqueobotánicos del sitio CH1, localizado en el norte de la provincia del Neuquén (Figura 1). Las primeras menciones sobre su existencia tuvieron lugar en relación con el arte rupestre en la década de 1970 (Fernández 1974-1975; Schobinger 1985; ver también Cordero et al. 2002). Posteriormente, se retomaron estos trabajos iniciales en el marco de un proyecto regional dirigido a caracterizar el uso humano del espacio en un paisaje heterogéneo a nivel topográfico y ecológico. En esta línea, este proyecto macro incluyó el análisis de conjuntos líticos, faunísticos, arqueobotánicos, de arte rupestre y paleoecología (Barberena et. al. 2015).

Este trabajo expande el análisis de la cuadrícula A1, excavada en el año 2009 (Llano y Barberena 2013), y presenta los resultados de las cuadrículas C1 y D1, excavadas en el año 2012. El objetivo general consiste en evaluar las prácticas de subsistencia desde el registro de macrorrestos vegetales, considerando sus implicaciones en términos de estacionalidad ocupacional del emplazamiento. Se tiene en cuenta la amplitud de dieta humana producto de la selección de taxones, por lo cual se evalúan su frecuencia, densidad y riqueza específica. Se enfoca particularmente en evaluar las tendencias temporales en las pautas de procesamiento y descarte antrópico de plantas. Para este análisis se cuenta con un robusto marco cronoestratigráfico para las cuadrículas A1-C1-D1, que incluye numerosas fechas taxón para las principales especies consumidas (ver abajo).
Las excelentes condiciones de preservación de los macrorrestos en toda la secuencia permiten reconocer indicadores tafonómicos de manejo y descarte antrópico. El análisis pone de relevancia la existencia de similitudes y diferencias en la composición taxonómica de los conjuntos, lo cual conlleva a una interpretación más compleja de las estrategias de aprovisionamiento, procesamiento y descarte de vegetales y, sobre esta base, de una reconstrucción de la historia ocupacional del sitio mediante la integración con múltiples líneas de evidencia.
CH1 en su contexto ecológico regional
En la región sur de los Andes patagónicos (30-40°S) ocurre la transición de los sistemas de circulación atmosférica Subtropical y Templado, lo que se traduce en un clima árido y semiárido (Páez et al. 2004). La cordillera occidental recibe precipitaciones promedio de 2000 mm anuales, mientras que las estepas patagónicas localizadas al este reciben menos de 300 mm anuales (Roig 1998). Estas condiciones climáticas crean como unidad fisiográfica un gradiente bioecológico pronunciado que genera ecosistemas variados en distancias cortas a partir del cual es posible distinguir tres regiones principales. De oeste a este, se localiza inicialmente la provincia fitogeográfica Altoandina (2600-2800 msnm), caracterizada por suelos rocosos muy pobres en materia orgánica, excepto en las turberas, que actúan como reservorios de agua donde la vegetación característica es una estepa de gramíneas y plantas en cojín. Los principales taxones son Azorella, Nassauvia, Maihueniopsis y Senecio. En segundo lugar se encuentra la provincia fitogeográfica Patagónica de llanuras, mesetas y montañas bajas dominantes con vegetación arbustiva y herbácea (Pappostipa, Mulinum, Chuquiraga), con una elevada capacidad de carga de herbívoros en la estación estival (Mendía 2006). Por último, la provincia fitogeográfica del Monte, compuesta por llanuras y bolsones con suelos en general arenosos con predominio de vegetación arbustiva xerófila (Larrea, Prosopis y Prosopidastrum) (Roig et al. 2009). Algunas de estas especies son clave desde el punto de vista de su potencial alimenticio para el consumo humano (Llano 2014).
Historia paleoambiental
El escenario paleoambiental del norte de Patagonia incluye el análisis de múltiples indicadores con diferencias en sensibilidad y resolución temporal (Markgraf et al. 2009; Fernández et al. 2012). Estos estudios se focalizan en el área de bosque y el ecotono estepa-monte (de Porras 2017).
En el tardiglacial (~17.000 años cal. AP), las condiciones frías y secas se reflejan en el desarrollo de un bosque abierto de Nothofagus. Durante el Holoceno temprano (11.000-8000 años cal. AP), un aumento en la diversidad en los elementos de la estepa señalaría un incremento en la precipitación y temperatura, pero sin alcanzar los valores actuales. Para el Holoceno medio (8000-3000 años cal. AP), el registro demuestra condiciones húmedas, visualizado en la presencia de Cyperaceae. Finalmente, en el Holoceno tardío (últimos 3000 años cal. AP) se evidencian condiciones climáticas similares al presente, con precipitaciones invernales y veranos secos. Estas condiciones indican que el paisaje fitogeográfico se mantuvo sin alteraciones significativas en los últimos milenios (de Porras 2017).
En escala local, estudios preliminares del polen fósil de CH1 sugieren que las comunidades de arbustos y gramíneas estuvieron presentes en el entorno del sitio desde ca. 13.800 años cal. AP. Si bien los conjuntos polínicos no evidencian cambios sustantivos, hay diferencias en la composición. Entre ca. 13.800-9500, el polen indica la presencia de estepas graminosas típicas de la provincia Patagónica, integrada con arbustos ( Lycium, Schinus). Estos resultados sugieren condiciones locales más frías que en el presente, consistentes con los resultados de microfauna de CH1 (Fernández et al. 2012). Para los últimos 1500 años, el polen indica la presencia de la transición Monte-Patagonia, con dominio de Lycium, Schinus y gramíneas en menor proporción, lo que refleja condiciones similares a la vegetación moderna ( Pompei et al. 2012).
MATERIALES Y MÉTODOS
El sitio CH1 está ubicado en la provincia del Monte, 8 km al este del ecotono con la provincia Patagónica (Figura 1). Consiste en una cueva originada por procesos erosivos cuyo espacio bajo reparo es de 30 × 20 m. El gran tamaño del espacio, sumado a la baja circulación de agua en un amplio sector del sitio, redundaron en excelentes condiciones de acumulación sedimentaria y preservación de materiales orgánicos. En total se excavaron cuatro cuadrículas. En este trabajo se presenta nueva información para las cuadrículas C1 y D1, que se integra con los datos ya publicados para la cuadrícula A1 ( Llano y Barberena 2013). Cada una de estas tres tiene una dimensión de 2 × 1 m, lo que totaliza 6 m2 de superficie de excavación. No se presenta el análisis de la última cuadrícula excavada (B1) porque presenta procesos significativos de migración de materiales por remoción antrópica reciente. Considerando que la composición taxonómica de la cuadrícula B1 es semejante a las aquí estudiadas, se optó por excluirla del análisis detallado.
Para C1 y D1 se alcanzó una profundidad máxima de 1 m (4 m3 de volumen excavado). La excavación se realizó empleando niveles artificiales de extracción de 10 cm de potencia, numerados a partir de 1 desde la superficie hacia la base, y luego se efectuó la localización tridimensional de los hallazgos. La numeración de los componentes se estableció desde el más antiguo hasta el más reciente. Los sedimentos fueron tamizados en seco utilizando zarandas con malla de 1 mm de apertura. Todas las fechas mencionadas en el texto se presentan en años calibrados AP obtenidos con dos sigmas mediante la curva del hemisferio sur SHCAL 13 (Hogg et al. 2013). La secuencia cronoestratigráfica describe la sucesión en las tres cuadrículas analizadas (Tabla 1, detalles en Barberena 2014).
Tabla 1Fechados radiocarbónicos del sitio CH1. Se encuentran resaltados los fechados puntuales sobre material vegetal (publicados en Barberena 2014)
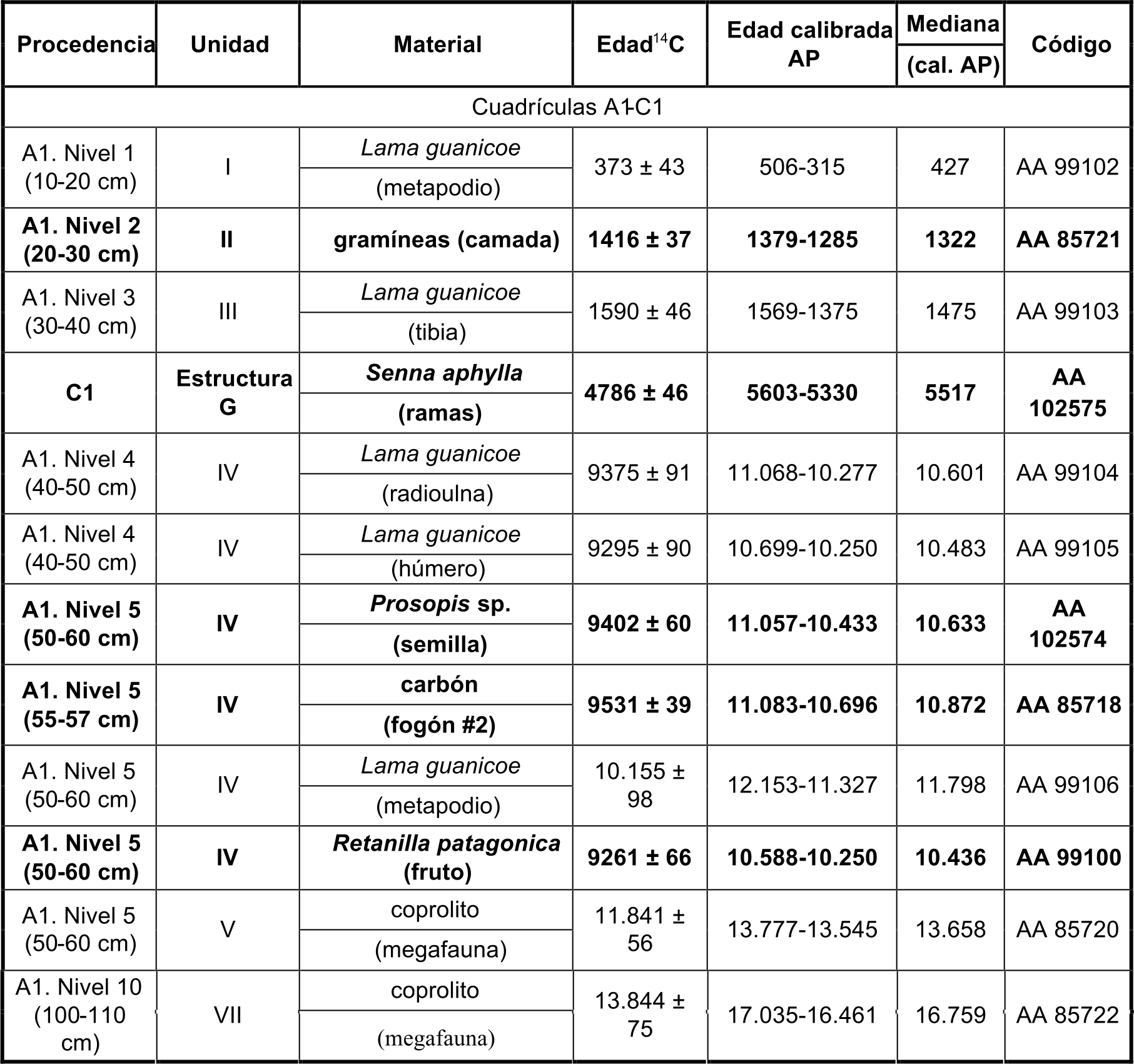
Componente 1: extracciones artificiales 10 a 7. Las fechas disponibles abarcan un rango temporal de entre 16.600-13.600 años cal. AP. No se registraron evidencias arqueológicas in situ, aunque se destaca la notable abundancia de excrementos de carnívoros pequeños con una excelente preservación (Tietze et al. 2019). Se fecharon en forma directa dos coprolitos que corresponderían a perezoso. El límite superior de este componente está dado por una discordancia erosiva denominada Hiato 1.
Componente 2: extracciones artificiales 6 a 4 ubicadas inmediatamente por encima del Hiato 1. Incluye las primeras ocupaciones humanas en CH1 fechadas entre 11.300-10.200 años cal. AP.
Componente 3: fue excavado como una unidad de extracción en sí misma. Corresponde a una estructura antrópica denominada G, un pozo relleno de material vegetal datado en 5600-5300 años cal. AP.
Componente 4: extracciones artificiales 3 a 1, fechadas entre 1500-300 años cal. AP. Coincide con un pulso de elevada ocupación regional.
Métodos
El planteo metodológico se basó en el análisis de macrorrestos (hojas, areolas, frutos, espinas, semillas) provenientes de muestras estratigráficas que reflejan el material disperso contenido en la matriz. La clasificación del material botánico se realizó a ojo desnudo y con microscopio estereoscópico (Zeiss Modelo Stemi DV4) con un aumento de 10x a 20x. Los materiales se encontraron secos en su totalidad. La identificación taxonómica se llevó a cabo por medio de anatomía comparada y observación de la morfología externa de los especímenes con la colección de referencia de frutos y semillas colectadas en las inmediaciones del sitio. Para ello se realizaron tres muestreos tomando puntos de intercepción (Mostacedo y Fredericksen 2000), en los que se recolectaron de manera sistemática los especímenes potencialmente explotables (resultados en Barberena et al. 2018). Además, se integraron plantas de importancia económica de la región, referenciadas en diferentes fuentes escritas (Azar 2008). Se evaluó la presencia de señales tafonómicas como cortes, decorados y marcas de fricción (Llano et al. 2012).
La metodología utilizada para la identificación de la muestra conformada por el cariopse de maíz fue tanto cuali como cuantitativa (Oliszewski y Olivera 2009). Para las variables cuantitativas se relevaron: longitud, latitud y espesor (mm). Se utilizó un calibre HL (150 × 0,02 mm). Las variables cualitativas que se tuvieron en cuenta fueron: forma, color, dureza y composición del endosperma.
Para realizar el análisis general se han utilizado tres parámetros: a) número de individuos, que corresponde a la suma total de restos de un taxón, excluyéndose los fragmentos; b) densidad, utilizado para comparar muestras de volúmenes diferentes (densidad = conteo absoluto/volumen); y c) Índice de Abundancia Relativa (IAR), que permite comparar cambios y similitudes sin sobrevalorar ciertas concentraciones puntuales de individuos (Popper 1988).
Los patrones de procesamiento son clave para evaluar la incorporación antrópica de los macrorestos a la secuencia del sitio. Los resultados son comparados con diseños experimentales (Capparelli y Lema 2011; Llano et al. 2012) y con datos etnohistóricos (Capparelli y Prates 2015). Se prestó especial atención a las partes del fruto de algarrobo, dada su abundancia. Para la evaluación de las características de los fragmentos botánicos se descartaron las posibilidades de tostado y fragmentación poscarbonización (sensu Capparelli y Prates 2015), ya que los macrorrestos de algarrobo se encontraban en su totalidad secos. Las variables evaluadas fueron el remojo, hervido, molienda para harina y consumo directo.
RESULTADOS
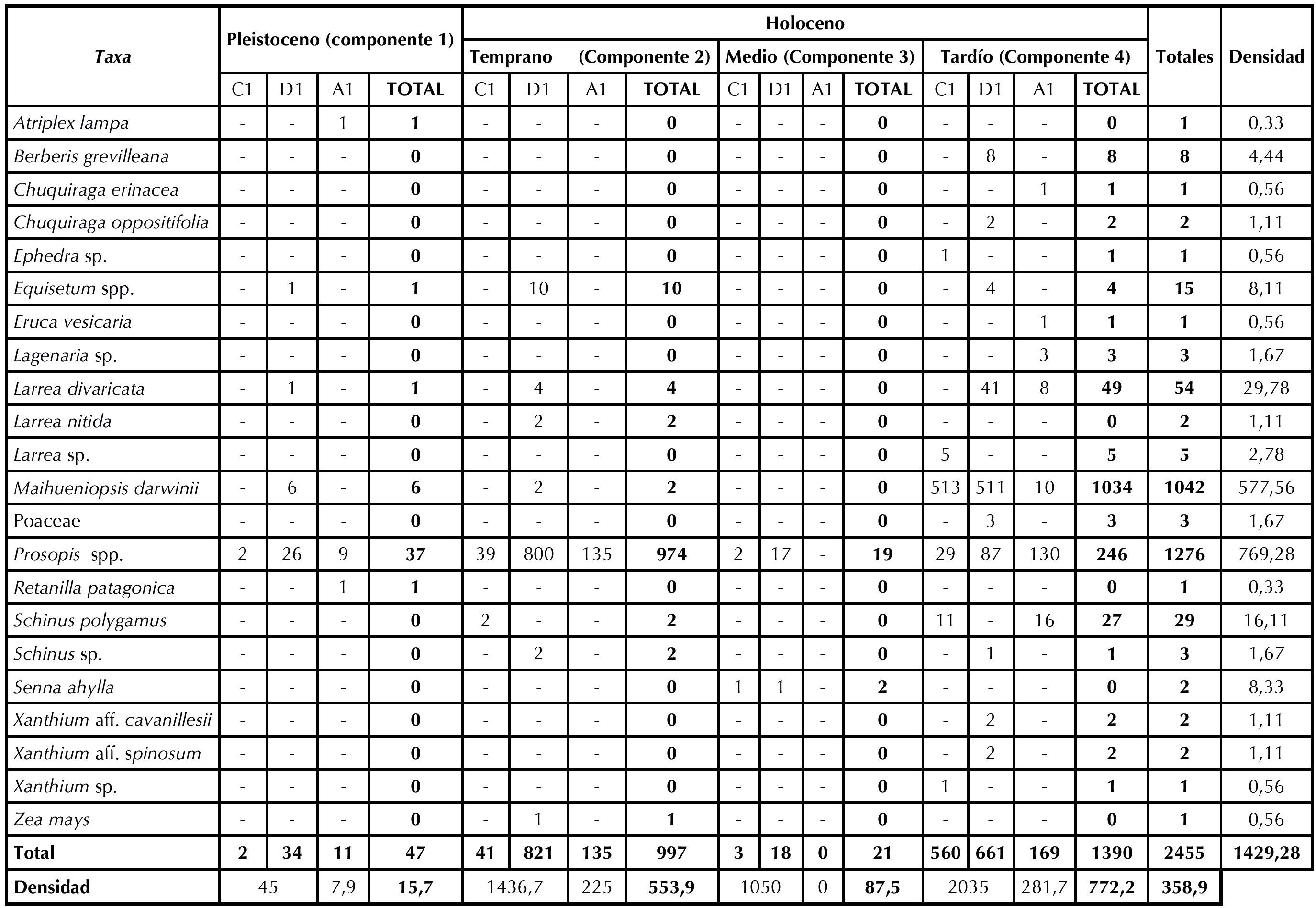
Se recuperaron 2140 macrorrestos vegetales de las cuadrículas C1 y D1, los cuales han sido identificados en diferentes grados de especificidad (Figura 2). En la Tabla 2 se presentan las frecuencias absolutas y relativas por nivel estratigráfico de cada componente, donde se visualiza que de la cuadrícula C1 se recuperaron 606 macrorrestos (28% del total), mientras que de D1 se recuperaron 1534 especímenes (71%). Al tratar ambas cuadrículas en forma conjunta por componente cronoestratigráfico, se observa que en el componente 1, que corresponde al Pleistoceno final, se recuperó el 1,6% (n = 36) del total del material arqueobotánico (Tabla 3), entre el que se destaca Prosopis spp. (n = 28), acompañado por Maihueniopsis darwinii (n = 6), Larrea divaricata (n = 1) y Equisetum spp. (n = 1).

Figura 2Registro fotográfico de los macrorrestos vegetales recuperados en CH1. A. Espinas de Berberis grevillana; B. tallos de Equisetum spp. y madera indeterminada; C. Endocarpo de Schinus sp.; D. semillas y epicarpio de Prosopis spp.; E. artejos, espinas y areolas de Maihueniopsis darwinii; F. estructura G de Senna aphylla.
Tabla 2Frecuencias absolutas y relativas por nivel estratigráfico de cada componente de los macrorrestos vegetales recuperados del sitio CH1-Cuadrículas C1 y D1. S/V sin vegetales.
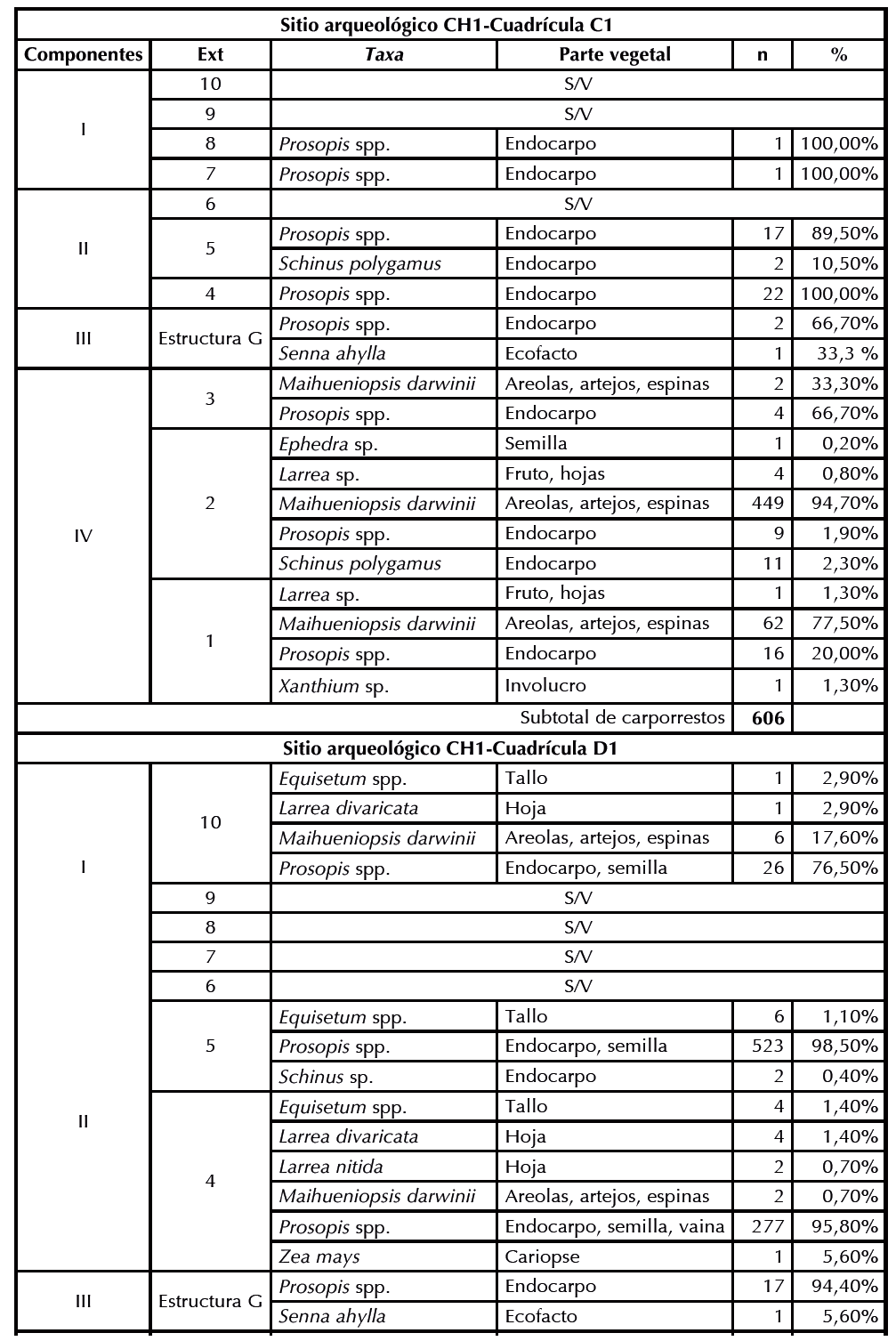
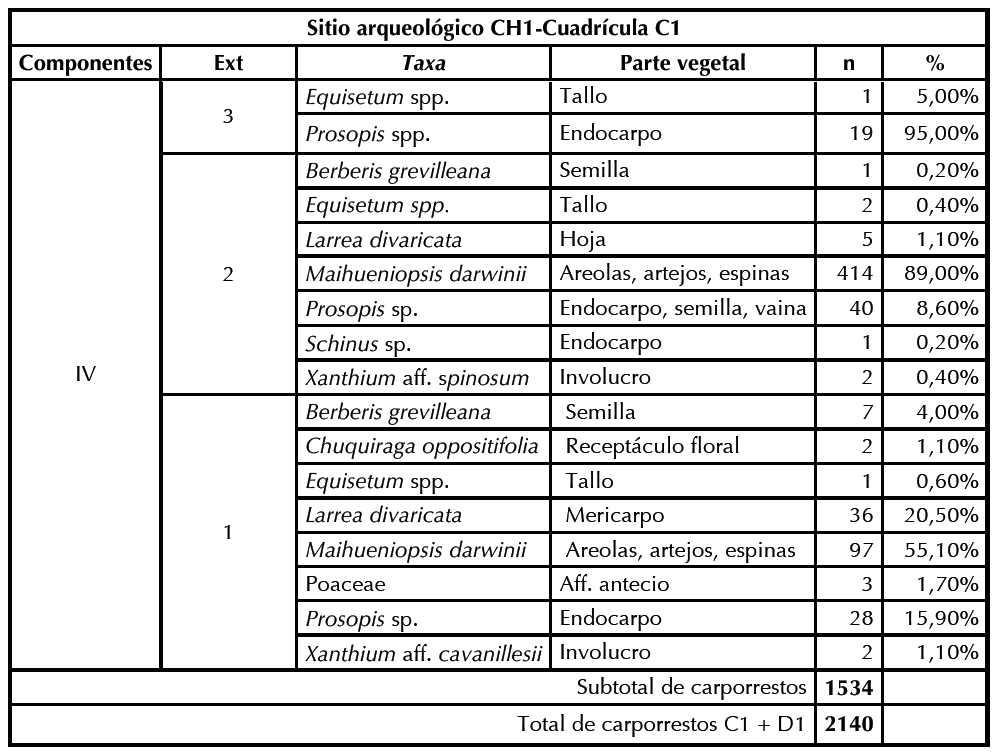
En el componente 2 (Holoceno temprano) se registra el 40,2% (n = 862) de los especímenes vegetales. En la cuadrícula C1 se registraron dos taxa: Prosopis spp. (n = 39) y Schinus polygamus (n = 2), mientras que en D1 la abundancia de Prosopis spp. (n = 800) supera la de todos los periodos evaluados. Un conjunto de taxones se repite en este lapso comparado con el anterior, aunque en baja frecuencia: Maihueniopsis darwinii (n = 2), Larrea divaricata (n = 4), Equisetum spp. (n = 10), Schinus sp. (n = 2) y Larrea nitida (n = 2). El componente 3 (Holoceno medio) solo se encuentra representado por una estructura antrópica denominada G, que es un pozo relleno por Senna aphylla, con contenido de endocarpos de Prosopis spp. (n = 19).
En el componente 4 (Holoceno tardío) se evidencia una mayor abundancia en términos de cantidad absoluta de recursos vegetales (57,08%, n = 1221) y una diversificación taxonómica. La densidad de este componente refleja que el Holoceno tardío es superior a la del resto de los componentes.
Se destaca que la densidad de los macrorrestos de Maihueniopsis en este componente es significativamente superior a la de los componentes previos (Pleistoceno 7,5 ind/m3; Holoceno temprano 3,3 ind/m3; Holoceno medio 0 ind/m3; Holoceno tardío 1706,7 ind/m3). Otro caso llamativo es el de Prosopis, que, en términos de densidad, es superior en el Holoceno temprano (1398,3 ind/m3) que en el resto de los componentes (Pleistoceno 35,0 ind/m3; Holoceno medio 950 ind/m3; Holoceno tardío 193,3 ind/m3). Considerando la densidad en conjunto (C1 + D1) se observan valores superiores en el Holoceno tardío (2035 ind/m3) con respecto al Holoceno temprano (1436,7 ind/m3), Holoceno medio (1050 ind/m3) y el Pleistoceno (45,0 ind/m3) (Tabla 3).
Se registró un cariopse completo de Zea mays (Figura 3) en el nivel 4 de la cuadrícula D1, que estratigráficamente corresponde al componente 2 del Holoceno temprano. Este se encontraba en estado de preservación seco, tiene forma obovada y el color del pericarpio es marrón. Las dimensiones son 7 mm de longitud por 9 mm de latitud y 6 mm de espesor. Se identificó la variedad (sensu Parodi 1959): Zea mays var. amylacea. Los caracteres cualitativos se describen como endosperma totalmente harinoso, blando y blanco, con una porción córnea periférica muy delgada.

Dentro del conjunto de restos de Prosopis predominan los fragmentos de vaina, como artejos enteros y medios artejos (sensu Capparelli y Prates 2015). Se observaron marcas de masticación en los endocarpos enteros y, en el caso de endocarpos terminales, se encontraban acompañados de restos de epicarpo en forma de finas hebras entrelazadas, lo que evidencia el consumo fresco sin procesamiento (Figura 2). También se recuperaron epicarpios de consistencia gruesa y fibrosa, signo observado en la experimentación de molienda con vainas secas (Llano et al. 2012).
Por otra parte, algunos de los macrorrestos de Maihueniopsis, madera y la totalidad del material que conforma la estructura G (Senna aphylla) presentaban signos de tinción antrópica por ocre en tonalidades pardo-rojizo.
DISCUSIÓN
Las escasas reconstrucciones paleoambientales disponibles para la región no registran la existencia de modificaciones sustanciales en la distribución de la vegetación durante el Holoceno (Markgraf et al. 2009; de Porras 2017). Las principales variaciones se relacionan con las fluctuaciones de los límites de las zonas de transición, debido a cambios en la humedad efectiva. Aunque es probable que esto se deba al carácter acotado y fragmentario de las reconstrucciones disponibles, se puede afirmar que la posición del ecotono Monte-Patagonia no se habría corrido lateralmente como para generar un impacto en las comunidades de vegetación que rodean a CH1, que habrían sido similares a las actuales.
Los restos hallados en el componente 1 –fechado en el Pleistoceno final– se habrían depositado como producto de la ocupación de la cueva por parte de herbívoros. La matriz de estos estratos está conformada en gran parte por material orgánico producto de la desagregación de coprolitos de megafauna. No se cuenta con indicadores contextuales fehacientes que respalden el carácter antrópico de estos materiales (Barberena et al. 2015). En este sentido, se los considera un indicador de las condiciones paleoecológicas en forma previa a la llegada de los primeros grupos humanos a la región.
La secuencia ocupacional de CH1 indica prácticas de subsistencia de caza y recolección de acuerdo con las cuales el aprovechamiento de los recursos vegetales silvestres, tales como Prosopis, Schinus y Maihueniopsis, se reconocen desde el inicio de las ocupaciones humanas en el Holoceno temprano (componente 2) de A1, C1 y D1. El registro de tallos y semillas, representado por una riqueza de cinco taxones en este lapso, indica la presencia de plantas arbustivas típicas de monte (i.e., Prosopis) y plantas indicadoras de zonas húmedas (i.e., Equisetum), que fructifican en época estival, lo que sugiere el uso de laderas cerca de fuentes de agua y ambientes húmedos, los cuales habrían estado presentes en forma discontinua en el entorno de CH1. El registro del componente 2 indica el consumo redundante de algarrobo (Prosopis spp.) con fecha taxón directa para una muestra de frutos del nivel 5 de la cuadrícula A1 con una mediana calibrada de 10.633 años AP (Tabla 1).
Los componentes 2, 3 y 4 evidencian señales de procesamiento y depositación antrópica de estos restos, ya que los endocarpos se encuentran fragmentados y con señales de fricción (Figura 2). Esto sugiere que, al menos, una parte de estas plantas fueron procesadas antes de ser utilizadas. Además del uso alimentario, los macrorrestos presentaron impregnaciones con pigmento colorante en Senna aphylla (componente 3), Maihueniopsis y Equisetum (componente 4). Los macrorrestos asignados a Ephedra, Larrea y Xanthiun no presentaban marcas de procesamiento.
Si bien los usos potenciales de las plantas están limitados por factores biológicos, el comportamiento humano juega un papel importante en la definición del uso de un recurso ( Ford 1979). Las evidencias obtenidas en referencia al procesamiento de Prosopis permiten marcar la existencia de diferentes estrategias en el Holoceno temprano (componente 2), comparado con el Holoceno tardío (componente 4). En el componente 2 se observa que los restos de algarrobo son producto del consumo directo mediante mascado de vainas sin procesamiento previo bajo la forma de tostado o hervido. Por el contrario, en el componente 4 del Holoceno tardío se observan marcas en las vainas compatibles con un consumo que implicó procesamiento como la molienda. Se señala que, aunque en baja frecuencia, en el sitio CH1 y en la localidad Huenul en general se registró la presencia de artefactos líticos de molienda.
En cuanto a la estacionalidad de este tipo de recurso, en general se establecieron en períodos de producción de frutos y semillas, los cuales coincidieron con el verano. Al analizar estas tendencias desde el rendimiento nutricional y la fenología vegetal, es esperable que en el pasado las poblaciones hayan consumido mayor cantidad de algarrobo, ya que no solo está ranqueado en primer lugar por su rendimiento calórico (Llano y Ugan 2010), sino que tiene una amplia época de fructificación, disponibilidad para el consumo y posibilidad de almacenamiento. En el caso de Maihueniopsis, la época de fructificación está acotada a los meses de noviembre a enero (Roig 1998), que sería un indicador de uso estival. En tanto que el uso tecnológico para el acondicionamiento del lugar (Senna aphylla) es potencialmente anual.
La interpretación del consumo de especies que ofrece el entorno se corresponde con los estudios de ecología isotópica regional para el extremo norte de la provincia del Neuquén (Gordón et al. 2017), ya que todos los ecosistemas tendrían una señal dominante de base C3. Este dato se sustenta en la señal isotópica de plantas de esa vía fotosintética registrada en el muestreo vegetacional en la localidad Huenul, así como en los valores de 13C/12C registrados para muestras de Lama guanicoe de CH1 (Barberena et al. 2018).
En base al pequeño conjunto de CH1 que es asignado a esta instancia temprana del poblamiento humano regional, se registra el consumo de algarrobo y guanaco (Barberena et al. 2015), que son respectivamente los taxones de más alto ranking dentro de los recursos vegetales y animales en la región (Llano 2015). El único ejemplar de Zea mays se recuperó en el nivel 4 asignado al componente 2. Se considera altamente probable su migración vertical, sobre la base de los registros disponibles para este taxón exclusivamente para el Holoceno tardío en zonas cercanas a la región de estudio (Pérez y Erra 2011; Lema et al. 2012), como así también en la vertiente occidental de la cordillera (Roa et al. 2018). Se está preparando un informe que permita posicionarlo a nivel cronológico y morfológico (Llano en preparación).
El componente 3 del Holoceno medio no resulta comparable a los otros, ya que corresponde exclusivamente a la estructura G, que es un pozo relleno de material vegetal teñido con pigmento rojo (fecha taxón para Senna aphylla con una mediana calibrada de 5517 años AP). Esta es la única evidencia registrada en estratigrafía para este período de tiempo escasamente representado a nivel arqueológico macrorregional (Neme y Gil 2009; Barberena et al. 2015). Esta estructura contiene también escasos especímenes de algarrobo. El registro en el Holoceno tardío reflejó una mayor densidad y riqueza especifica de especies recuperadas, con un total de 14 taxones identificados, entre los que es dominante Maihueniopsis darwinii. Este conjunto arqueobotánico no solo es abundante, sino que también indica el procesamiento de la planta in situ con trazas de pigmento rojo en ciertos casos.
La disminución en la densidad de algarrobo en el Holoceno tardío comparado con el Holoceno temprano refleja una conducta que tiene correlato con los relevamientos actuales en torno al sitio ( Sosa 2016) y con los estudios macrobotánicos y palinológicos realizados en paleomadrigueras de roedores de la localidad Huenul (Llano et al. 2019) ya que se interpreta que, entre 12.000 y 8000 AP, el entorno ofrecía una mayor abundancia de algarrobo. La disponibilidad hídrica también podría condicionar el patrón de distribución espacial de los algarrobales ( Villagra et al. 2004), que se localizan en sitios donde se disponga de agua freática anual. Actualmente se ha observado la amplia disponibilidad de este taxón en el entorno del arroyo Buta Có, ubicado a ca. 4 km de CH1, donde está situada la localidad arqueológica Puesto Cuello-Cueva Yagui.
Por último, este registro posiblemente evidencie la recolección cuando los ocupantes del sitio realizaban movimientos con objetivos diversos. En este sentido, por su disponibilidad dispersa en la región, una posibilidad es que la recolección de plantas ocurriera en forma complementaria en el marco de la realización de otras actividades ( Binford 1979), tales como búsqueda de agua, que no está disponible en forma permanente en el entorno de CH1, recursos líticos, o presas de caza como el guanaco. Los frutos y semillas de las plantas fueron probablemente consumidos siempre que estuvieron disponibles, lo que evidencia así la especificidad en las zonas de recolección, pero también que tenían suficiente importancia para ser llevados al sitio, donde se habrían realizado las actividades de procesamiento y, muy posiblemente, de consumo.
CONCLUSIÓN
El registro arqueobotánico presenta variaciones en la densidad y riqueza específica de plantas entre las cuadrículas analizadas. Los restos estudiados sugieren una mayor intensidad de descarte de vegetales en el componente 4, que representa entre 1500-300 años AP. El consumo y uso de plantas habría aumentado con el tiempo, representado por una gran riqueza del registro de semillas, especialmente cuando se comparan las etapas iniciales de ocupación con las tardías. Sin embargo, cabe señalar que, dado el carácter breve de las ocupaciones iniciales en CH1, la muestra puede ser muy poco representativa de la dieta humana. Sí se destaca el registro de formas de procesamiento más intensivas en el Holoceno tardío.
La abundancia de algarrobo sustenta su rol como alimento, basado principalmente en la ingesta de proteínas y lípidos (Llano y Ugan 2010), lo que se corresponde con la categorización de alimento suplementario propuesto por Tanaka (1998). Las marcas de procesamiento en los macrorrestos indican un consumo en fresco en momentos tempranos, que puede ser caracterizado como “expeditivo”, mientras que, hacia los períodos tardíos, el registro muestra un aprovechamiento más intensivo del recurso, en asociación a una mayor inversión de trabajo, que permite aumentar el rendimiento calórico.
Se debe destacar que la presencia del cariopse de maíz constituye hasta el momento el único macrorresto del taxón en el noroeste de Patagonia, si bien se cuenta con antecedentes de esta especie representada por microrrestos (Pérez y Erra 2011; Lema et al. 2012). El marco regional de evidencias no sugiere la producción local de esta especie doméstica, sino el ingreso a la región y al sitio bajo mecanismos indirectos tales como el intercambio (Gil 2006). En este sentido, esta evidencia puede ser agrupada con los especímenes de Lagenaria sp. recuperados en CH1, algunos de los cuales presentan evidencias de decoración mediante la técnica de pirograbado ( Llano y Barberena 2013).
El registro arqueobotánico ofrece un marco para situar los análisis de amplitud de dieta y pautas de movilidad humana para la recolección y explotación de recursos en diferentes épocas del año. Los resultados presentados reflejan la conducta de trasladar estas plantas al sitio, posiblemente en forma complementaria al consumo, pero también puede ser que su aprovisionamiento se haya realizado para cubrir otras necesidades; es factible el ingreso accidental de la planta recolectada y que la parte realmente usada corresponda a otras, como las cactáceas pigmentadas o las maderas. Este punto es un tema pendiente en la agenda de análisis botánico del sitio, puesto que se recuperaron maderas con pirograbados similares a los dibujos en el epicarpio de Lagenaria sp. (Llano y Barberena 2013). Actualmente, la representación de motivos en soportes de material leñoso está siendo analizada como parte de un trabajo doctoral que involucra el estudio sobre la comunicación visual mediante múltiples tipos de imágenes presentes en el registro arqueológico del área y en diferentes tipos de soporte ( Romero Villanueva y Barberena 2017). La arqueobotánica permite generar una imagen más completa de las sociedades humanas que habitaron el noroeste de Patagonia.

