











 Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por email Citado por SciELO
Citado por SciELO  Similares em
SciELO
Similares em
SciELO 
 Permalink
Permalink
INTRODUCCIÓN
En la última década se hicieron numerosas prospecciones y excavaciones arqueológicas en el extremo norte de la provincia del Neuquén, Argentina (e.g., Gordón et al., 2017 y trabajos allí citados). El sitio Cueva Huenul 1 (en adelante CH1) se destaca por las excelentes condiciones de preservación de sus depósitos estratificados, la presencia de un componente paleontológico con megafauna extinta del Pleistoceno tardío (~16,6 ka cal. AP) y de tres componentes arqueológicos holocénicos (Barberena et al., 2010, 2015; Pompei et al., 2012; Barberena, 2014, 2017). CH1 aportó datos valiosos en materia de aprovisionamiento y contexto artefactual lítico, arte rupestre, registro arqueobotánico, polen, paleoparasitología, isótopos estables y zooarqueología (Beltrame et al., 2011; Fernández et al., 2011a, 2012; Pompei et al., 2012; Llano y Barberena, 2013; Barberena et al., 2015, 2018; Llano et al., 2019, 2020). Algunas de las investigaciones mencionadas se orientaron hacia el análisis paleoambiental (Fernández et al., 2011a, 2012; Pompei et al., 2012; Llano et al., 2020), principalmente para evaluar si los cambios del ambiente tuvieron incidencias en las ocupaciones humanas (Barberena et al., 2015, 2018; Barberena, 2017; Llano et al., 2020).
Previamente, en CH1 se habían registrado restos bien preservados de micromamíferos provenientes de la cuadrícula A1 (Fernández et al., 2011a, 2012). El estudio de esa muestra permitió establecer un conjunto faunístico conformado por dos taxones de roedores caviomorfos (Ctenomys sp., Microcavia australis), cuatro de roedores sigmodontinos (Akodon iniscatus, Eligmodontia sp., Euneomys chinchilloides, Phyllotis xanthopygus) y un marsupial (Thylamys pallidior). Los restos de estas especies habrían sido acumulados por la lechuza de campanario Tyto furcata (Fernández et al., 2012). La estructura taxonómica general de los conjuntos de micromamíferos se mantuvo estable desde la transición Pleistoceno-Holoceno hasta el Holoceno tardío final. La interpretación paleoambiental efectuada sugirió un paisaje regional de áreas abiertas y rocosas, aunque con mayor heterogeneidad hacia el Holoceno temprano (~9,5 ka cal. AP), reflejada por el registro de A. iniscatus y E. chinchilloides (Fernández et al., 2011a, 2012). En esos trabajos se enfatizó en la necesidad de aumentar la muestra para robustecer la hipótesis paleoambiental.
Afortunadamente, las nuevas excavaciones permitieron recuperar materiales apreciables. En este trabajo se incorporan al análisis los restos de micromamíferos (˂1kg) de tres cuadrículas inéditas, que abarcan la totalidad del material recuperado. El objetivo principal de esta investigación es brindar un panorama refinado de la estructura taxonómica y del valor paleoambiental de los pequeños roedores y marsupiales de CH1. Esto permitió rediscutir hipótesis ya formuladas (Fernández et al., 2011a, 2012) y robustecer las nuevas inferencias con otros proxies publicados para el sitio arqueológico (Llano et al., 2020).
MATERIALES Y MÉTODOS
Sitio arqueológico y marco ambiental
CH1 (36º57’S, 69º49’O, 1000 msnm; Figura 1A) se ubica en el extremo norte de la provincia del Neuquén, sobre la margen sur del río Colorado, en cercanías de su nacimiento. El sitio se emplaza en un área con extensa actividad volcánica en el retroarco Andino de la Payunia sur, concretamente, dentro del complejo del Tromen (e.g., Folguera et al., 2008). En pocas decenas de kilómetros en línea recta desde CH1, los rangos altitudinales oscilan desde 850 a 3900 msnm. Esta heterogeneidad topográfica determina que en distancias cortas se verifiquen cambios abruptos en las comunidades de micromamíferos (Fernández, 2012; Fernández et al., 2012; Bernardis, 2019; López et al., 2021). En este sector de Patagonia, comprendido entre los 30°–40°S, ocurre la transición de los sistemas de circulación atmosférica Subtropical y Templado, lo que se traduce en un clima árido y semiárido, con una variabilidad en la estacionalidad de las precipitaciones, que oscilan entre 200 y 300 mm anuales. La vegetación incluye un mosaico de comunidades fitogeográficas del desierto arbustivo del Monte y de la estepa arbustiva y herbácea Patagónica y Altoandina (e.g., Abraham et al., 2009).
Se realizaron dos campañas de excavación en CH1: en 2009 se abrieron las cuadrículas A1 (2 × 1 m) y B1 (1 × 1 m) y en 2012, las cuadrículas C1 y D1 (2 × 1 m cada una). Esto totaliza una superficie excavada de 7 m. y un volumen de sedimento removido de 7,75 m. (Barberena, 2014, 2017). Las cuadrículas A1, C1 y D1 se ubican en posición adyacente y comparten, en términos generales, la misma secuencia estratigráfica. La cuadrícula B1 se localiza en el sector más externo de la cueva y solo presenta el segmento temporal más reciente (Figura 1B).
La estratigrafía del sitio se organizó en cuatro componentes cronoestratigráficos que se extienden entre ca. 16,6 y 0,4 ka cal. AP (las fechas fueron publicadas en detalle, ver Barberena, 2014, 2017): el Componente 1 abarca el Pleistoceno final (16,6-13,6 ka cal. AP) y carece de evidencias arqueológicas in situ. Su límite superior está marcado por una discordancia erosiva denominada Hiato 1 (Figura 1C). El Componente 2 se extiende entre 11,3-10,2 ka cal. AP (Figura 1C). En los niveles que lo integran se registraron las primeras ocupaciones humanas. El Componente 3 corresponde en su totalidad a la estructura sedimentaria antrópica G, que pertenece a un amplio pozo en cubeta relleno de material vegetal (Senna aphylla) cubierto por ocre rojo y datado en 5,6-5,3 ka cal. AP (Llano et al., 2019). Hasta el momento, representa la única evidencia del Holoceno medio en CH1. Finalmente, el Componente 4 se registra en todas las cuadrículas excavadas con fechados de 1,5- 0,4 ka cal. AP, que lo vinculan al Holoceno tardío final (Figura 1C). Este coincide con un pulso de elevada ocupación regional y, en la escala del sitio, se asocia a las evidencias antrópicas más abundantes y diversas (Barberena et al., 2015; Llano et al., 2019).
En trabajos previos, Fernández y coautores (2011a, 2012) estudiaron la muestra de micromamíferos recuperada de la cuadrícula A1 (Número de Especímenes Identificados por Taxón, NISP = 1426; Número Mínimo de Individuos, MNI = 87). Las nuevas muestras aquí incorporadas corresponden a los restos de micromamíferos (NISP = 1707) de las cuadrículas B1 (NISP = 203), C1 (NISP = 921) y D1 (NISP = 583). En este trabajo se considerará la muestra total de CH1 (cuadrículas A1, B1, C1 y D1), conformada por un NISP = 3135 y un MNI = 324.
Los resultados tafonómicos de las cuadrículas B1, C1 y D1 se expresaron en el trabajo de Fernández y coautores (2017), los cuales coinciden con aquellos presentados por Fernández y coautores (2011a, 2012) para la cuadrícula A1. En síntesis, los bajos valores de corrosión digestiva registrados en todos los componentes –principalmente del tipo ligera, acorde con las distintas morfologías dentarias de los grupos de micromamíferos representados en el sitio– sugieren que los restos fueron acumulados por aves Strigiformes, posiblemente Tyto furcata (Fernández et al., 2017, pp. 45-46). La representación de roedores en los conjuntos con señal antrópica es mínima y el foco habría estado en el consumo de Lama guanicoe y especies vegetales (Maihueniopsis, Prosopis, etc.). En este sentido, no hay una evidencia clara de ampliación de la dieta mediante la incorporación de recursos animales de bajo ranking (Barberena et al., 2015; Llano et al., 2019).

Métodos
Los restos de micromamíferos se examinaron con lupa binocular y las determinaciones taxonómicas se realizaron comparando los restos craneodentarios con ejemplares de referencia de la colección de mamíferos del Grupo de Estudios en Arqueometría de la Facultad de Ingeniería (Universidad de Buenos Aires) y bibliografía (e,g,, Fernández et al., 2011b).
Para el análisis paleoambiental, se evaluaron la presencia/ausencia y los cambios en las frecuencias relativas calculadas a partir del MNI de algunas especies de micromamíferos consideradas como indicadores. La aproximación paleoecológica se basa, principalmente, en los requerimientos ambientales y de distribución conocidos para las distintas especies. Este método está afectado por la historia tafonómica de los agregados y el tamaño de la muestra (e.g., Andrews, 1990; Pardiñas, 1999). Como parámetro actualista, se utilizaron muestras de egagrópilas de T. furcata recuperadas en el sitio y en zonas adyacentes (Tabla 1, Figura 1A).
Se calcularon las curvas de rarificación para las muestras arqueológicas y actuales para evaluar la confiabilidad de la riqueza (NTaxa) y la abundancia taxonómica en términos de MNI. Además, se hicieron curvas de rarificación en base al índice de Shannon (H’) y al MNI, para ajustar las comparaciones entre muestras en materia de diversidad taxonómica. Se realizó un análisis de correspondencias para explorar el ordenamiento de las especies y de las muestras arqueológicas y actuales en forma conjunta, a través de una matriz de valores de abundancia (MNI transformados a logaritmo natural). Los análisis estadísticos se realizaron mediante el uso del programa PAST (PAleontologicalSTatistics), versión 4.03.
Tabla 1 Muestras actuales empleadas como parámetro actualista

NotaMuestras actuales de egagrópilas de Strigiformes empleadas como parámetro actualístico. Las localidades 3 (36º57’S, 69º49’O, 1000 msnm) y 4 (36º59’19,08’’S, 69º59’51,06’’O, 1813 msnm) son novedosas. Las localidades restantes fueron reportadas en Fernández y coautores (2012, p. 24). Localidades: 1= Barrancas; 2= Buta-Có, P.P. Tromen; 3= Cueva Huenul actual; 4= Paso de las Vueltas 4; 5= P.P. Tromen Arroyo; 6= 5,5 km W Buta Ranquil; 7= 12 km NE refugio P.P. Tromen; 8= Cerro Wayle; 9= P.P. Tromen; 10= 2,2 km W Laguna Tromen; 11= Ea. Corcel Negro; 12= 4 km SW Laguna Tromen; 13= 12 km SE Buta Ranquil.
RESULTADOS
A partir del material recuperado de todas las cuadrículas de CH1, la composición y abundancia taxonómica de los micromamíferos por cada componente arqueológico se detalla en la Tabla 2. El roedor sigmodontino Eligmodontia sp. es dominante en todos los componentes, con valores que oscilan entre ~40 y 47% del MNI total. Continúan, en orden de abundancia (~5-20%) y con proporciones relativamente constantes a lo largo de la secuencia, los roedores caviomorfos Ctenomys sp. y Microcavia australis, el sigmodontino Phyllotis xanthopygus y el marsupial Thylamys pallidior. Estos taxones, a su vez, tienen alta representación en las muestras actuales de egagrópilas de Tyto furcata recolectadas en cercanías de CH1 (Figura 2). En líneas generales, se mantienen las mismas tendencias taxonómicas previamente reseñadas para el sitio sobre la base del material de la cuadrícula A1, si bien con la incorporación de los restos de las cuadrículas B1, C1 y D1 algunos componentes se enriquecen con el caviomorfo Galea leucoblephara (Componentes 1, 2 y 4) y los sigmodontinos Abrothrix olivacea (Componente 1) y Oligoryzomys longicaudatus (Componentes 1 y 4). Por otra parte, los sigmodontinos Akodon iniscatus y Euneomys chinchilloides, previamente restringidos al Holoceno, a partir de este estudio se extienden también al Pleistoceno tardío (Tabla 2).

Figura 2 Cambios temporales en la abundancia relativa de algunas especies de micromamíferos en el sitio Cueva Huenul 1. Se excluyó al Componente 3 por tener un MNI = 4. Referencias: El: Eligmodontia sp., Ct: Ctenomys sp., Ma: Microcavia australis, Px: Phyllotis xanthopygus, Tp: Thylamys pallidior.
Tabla 2 Micromamíferos registrados en el sitio arqueológico Cueva Huenul 1 (expresado en NISP y MNI).
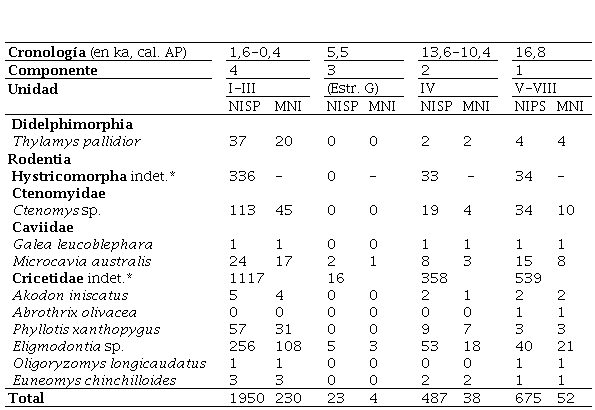
*Únicamente restos poscraneanos que posiblemente correspondan a los taxones identificados
*Únicamente restos poscraneanos que posiblemente correspondan a los taxones identificados.
El análisis de rarificación realizado sobre los valores de riqueza y diversidad taxonómica (índice de Shannon) muestra que el conjunto arqueológico de micromamíferos del Componente 4 (1,5-0,4 ka cal. AP) es el único que alcanza una asíntota. De esta forma, constituye el componente estadísticamente más significativo, pero dificulta su comparación con los otros componentes arqueológicos (Figura 3). La mayor parte de las muestras de micromamíferos actuales de egagrópilas de .. furcata, con un MNI ˃ 50, tienen un comportamiento con tendencia asintótica y constituyen una herramienta confiable como parámetros actualistas, considerando la riqueza (Figura 3A) y la diversidad taxonómica (Figura 3B). En este sentido, la Figura 4A grafica los valores de riqueza y diversidad de los componentes arqueológicos y de las muestras actuales más robustas, y señala además escasas variaciones entre ellas y, principalmente, entre los componentes arqueológicos y las muestras actuales más cercanas al sitio (e.g., Barrancas).
En congruencia con los resultados reseñados, el análisis de correspondencias (Figura 4B) exhibe un agrupamiento de las muestras arqueológicas y las actuales más próximas al sitio, lo que sugiere una estabilidad de las comunidades de micromamíferos a lo largo del tiempo. El Eje 1 (62,1% de la varianza total) agrupa, hacia los valores negativos, a los taxones principalmente asociados a la estepa patagónica y, en la topografía local, a mayores niveles de altitud (Abrothrix spp., Euneomys spp., Paynomys macronyx); hacia los valores positivos se concentran las especies que habitan en el Monte y zonas de menor altitud (Akodon dolores, Calomys musculinus, Graomys griseoflavus, G. leucoblephara) o potencialmente comunes a ambos ambientes (A. iniscatus, Eligmodontia spp., M. australis, O. longicaudatus, P. xanthopygus,Reithrodon auritus, T. pallidior). El Eje 2 (14,6% de la varianza total) reúne hacia los valores positivos a los sigmodontinos típicos del Monte (A. dolores, C. musculinus, G.griseoflavus), ausentes en CH1 y registrados en algunas muestras actuales ubicadas en un rango altitudinal de entre 1000 y 1450 msnm (Figura 1A, Tablas 1 y 2). La posición central y levemente hacia los valores positivos de los componentes arqueológicos se relaciona con la abundancia de especies asociadas a ambientes arbustivos, abiertos y rocosos, que pueden habitar tanto en el Monte como en la estepa patagónica (Eligmodontia spp., M. australis, P. xanthopygus,T. pallidior). Esto resulta coherente con la virtualmente nula ocurrencia de elementos típicamente patagónicos en la historia de CH1 (escasa representación de A. olivacea y .. chinchilloides; ausencia de A. hirta y P. macronyx).

Figura 3 Análisis de rarificación de las muestras de micromamíferos arqueológicas y actuales en base a la riqueza taxonómica (A) y en base al índice de diversidad de Shannon (B). La numeración de las muestras actuales se indicó en la nota de la Tabla 1.
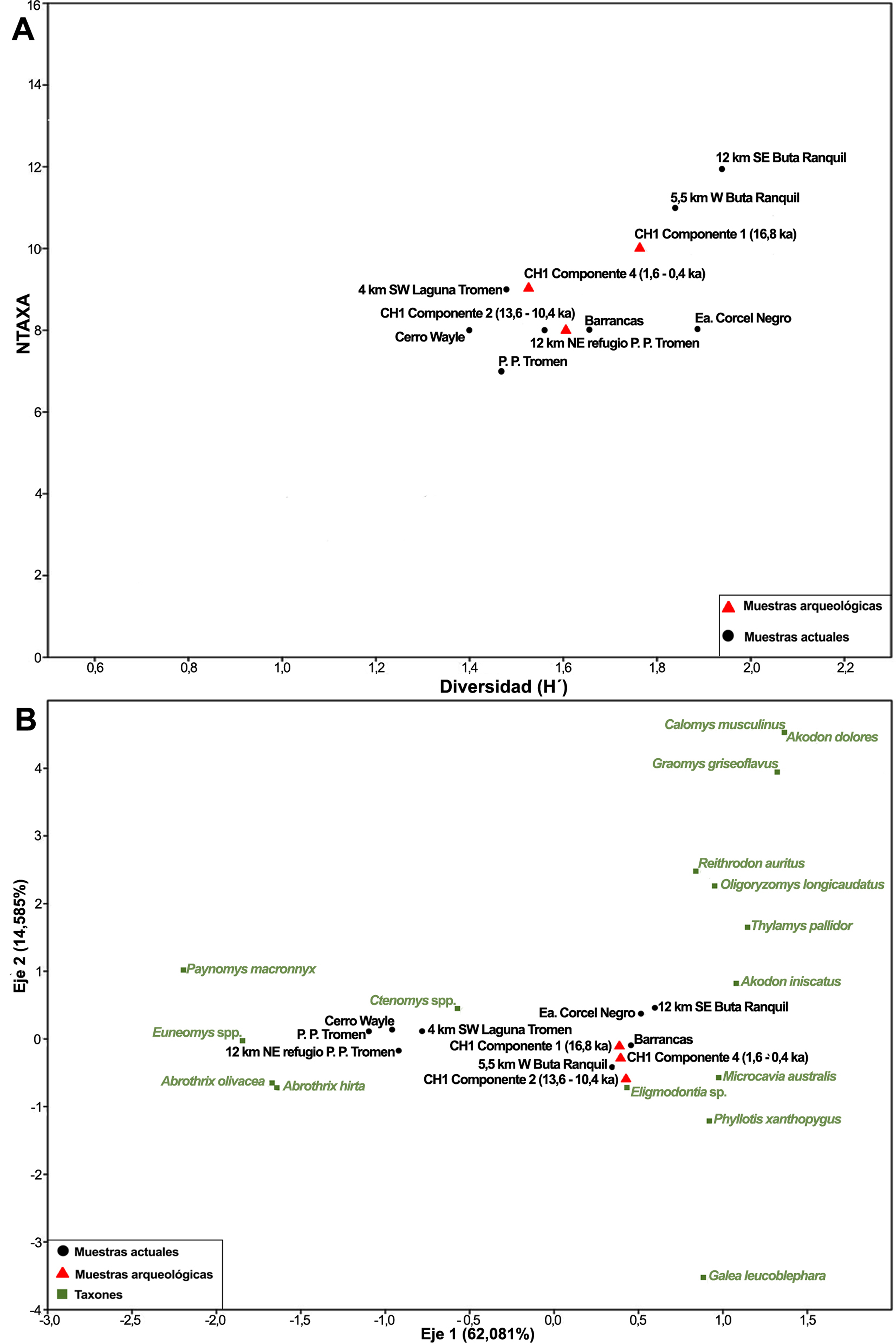
Figura 4 A. Riqueza (NTaxa) e índice de diversidad de Shannon (H’) de las muestras de micromamíferos del sitio Cueva Huenul 1 y de las muestras actuales de Tyto furcata; B. Análisis de correspondencias de las muestras de micromamíferos del sitio Cueva Huenul 1 y de las muestras actuales de egagrópilas de Strigiformes. Ambos ejes muestran los porcentajes de la varianza.
DISCUSIÓN
El aumento significativo en la muestra estudiada permite refinar algunos aspectos de la reconstrucción paleoambiental. En primer lugar, es necesario remarcar que la secuencia cronoestratigráfica de CH1 es discontinua y registra un segmento del Pleistoceno tardío-Holoceno temprano y otro del Holoceno tardío; la evidencia para el Holoceno medio es puntual, y al tratarse de una estructura sedimentaria antrópica, no representaría en forma significativa aportes no humanos. Por lo tanto, el planteo de estabilidad de las comunidades de micromamíferos que aquí se hace debe ser tomado con cautela. Al menos para las ventanas temporales indicadas, la estabilidad de los componentes faunísticos principales, tanto en presencia/ausencia como en abundancias relativas, resulta manifiesta.
El incremento de la muestra estudiada implicó, entre algunas novedades taxonómicas, el registro de Abrothrix olivacea como exclusivo del componente más antiguo. Si bien su presencia está sustentada por un único ejemplar, considerando su ausencia en los demás componentes, merece un tratamiento más pormenorizado. A. olivacea, al igual que el congenérico A. hirta, son abrotriquinos de amplia distribución y notable abundancia tanto en Patagonia continental e insular, andina y extraandina, pero están virtualmente ausentes en las porciones que quedan incluidas en el desierto del Monte (e.g., Lozada et al., 1996; Pardiñas et al., 2003). Esta restricción ambiental resulta elocuente en las muestras de egagrópilas actuales empleadas como parámetro comparativo (Tabla 1). En efecto, A. olivacea es depredado por lechuzas en las partes más altas del gradiente topográfico. Su ocurrencia en el componente finipleistoceno de CH1 podría indicar un descenso altitudinal de elementos faunísticos. Esto, a su vez, podría ser respuesta a condiciones climáticas más frías que las actuales imperantes en el área, vinculadas con un ambiente posglacial. Este tipo de fenómenos, coherente con englazamientos en los máximos orográficos, ha sido conjeturado en la cuenca superior del río Limay para explicar el registro fósil de Euneomys mordax en Cueva Traful 1 y Cueva del Caballo (Pearson, 1987; Pearson y Pearson, 1993; Tammone et al., 2016).
Los análisis realizados apuntan en definitiva a que, aunque los taxones ya estaban presentes en el Pleistoceno tardío, sus abundancias parecen ser un poco diferentes. En el área general de CH1 y en la zona adyacente boreal de la cuenca media del río Grande, la penetración de elementos más típicos del Monte (Akodon dolores, Galea leucoblephara .Graomys griseoflavus) parece verificarse siguiendo los principales cursos de los ríos y sus áreas de influencia (Fernández et al., 2015a). Contra eso, en tierras “intermedias” entre los valles fluviales y las porciones altoandinas, parece que las comunidades están conformadas por estos taxones (Eligmodontia, Microcavia, Phyllotis, Thylamys) y que la clave de su expresión paleoambiental podría intentar buscarse en sus abundancias, antes bien que en sus presencias.
Integración con otros proxies paleoambientales
La información paleoambiental del Pleistoceno tardío-Holoceno sigue siendo pobre en el extremo norte del Neuquén, concentrada en registros polínicos, arqueobotánicos, antracológicos y en paleomadrigueras de roedores (e.g.,, Markgraf et al., 2009; Fernández et al., 2011a, 2012; Pompei et al., 2012; Llano et al., 2019, 2020). Si ampliamos el área al noroeste de Patagonia y a ambas vertientes de los Andes, resulta una abundancia y diversidad mayor de proxies paleoambientales, con la adición de registros glaciológicos, sedimentológicos y dendrocronológicos (e.g.,, Mercer, 1982; Markgraf, 1983; Jenny et al., 2002; Espizua, 2003, 2005; Hajdas et al., 2003; Hofreiter et al., 2003; Gil et al., 2005; Whitlock et al., 2006; Espizua y Pitte, 2009; Bianchi y Ariztegui, 2012; de Jong et al., 2013; Frugone-Álvarez et al., 2020). En este panorama, los análisis que involucran restos de micromamíferos sugieren gran potencial, aunque por ahora la indagación sobre este tipo de materiales ha sido escasa (e.g., Pearson, 1987; Pardiñas, 1999; Teta et al., 2005, 2014; Agnolín y Pérez, 2014; Fernández et al., 2015a, 2015b, 2016; Fernández, 2012; Pardiñas y Teta, 2013; Tammone et al., 2016; Fernández y Pardiñas, 2018; López et al., 2021).
El análisis integral de los restos de micromamíferos de CH1 corrobora la hipótesis paleoambiental planteada a partir de los restos de la cuadrícula A1: un escenario de estabilidad ambiental desde el Pleistoceno tardío hasta la actualidad (Fernández et al., 2011a, 2012). En este contexto y desde los 16,6 ka cal. AP, los conjuntos locales habrían estado dominados por especies de micromamíferos que hoy en día habitan en áreas abiertas, arbustivas y rocosas de baja a media altitud, tanto en el desierto del Monte como en la estepa patagónica. Sin embargo, la discordancia erosiva revelada en CH1 entre 11,8 y 9,5 cal. AP (Barberena et al., 2010, 2015; Barberena, 2014, 2017) pudo ocultar los efectos del aumento de la aridez y del enfriamiento ocurridos a finales del Pleistoceno, en ocasión de la Inversión Climática Huelmo Mascardi (Hajdas et al., 2003). Este cambio climático tuvo una mayor expresión en otros sectores del noroeste de Patagonia, con variaciones en la composición florística local y/o el incremento en los regímenes de incendios forestales detectados en los bosques de Austrocedrus . Nothofagus (Whitlock et al., 2006; Bianchi y Ariztegui, 2012). Debido a la discordancia erosiva, ningún componente de CH1 tiene dataciones sincrónicas con los eventos áridos detectados en la región para el Holoceno medio (e.g., Markgraf, 1983; Gil et al., 2005; Zárate et al., 2010; Frugone-Álvarez et al., 2020; Llano et al., 2020). En un mismo sentido, los depósitos de CH1 tampoco se pueden correlacionar con los neoavances glaciarios registrados en los valles El Azufre y El Peñón del sistema del río Valenzuela de la cuenca superior del río Grande durante el Holoceno medio (5,7-4,4 ka cal. AP) y el Holoceno tardío (2,5-2,2 ka cal. AP), incluso, en tiempos más recientes (0,4-0,15 ka cal. AP) vinculados con la Pequeña Edad de Hielo (Mercer, 1982; Espizua, 2003, 2005; Espizua y Pitte, 2009). Sin embargo, se obtuvieron dataciones radiocarbónicas asociadas al Holoceno temprano y medio a partir de los excrementos del chinchillón Lagidium (Rodentia, Chinchillidae) recuperados de paleomadrigueras halladas en la localidad de CH (Llano et al., 2020). A partir de los macrorrestos vegetales, del polen y de los parásitos contenidos en esas deposiciones, se registró un aumento de la aridez entre 9,2 y 5,5 ka cal. AP. La escasez de restos de micromamíferos en la estructura antrópica G de CH1, con cronología asociada al Holoceno medio (5,6-5,3 ka cal. AP), tal vez sea la razón por la cual no se registraron las especies típicas del Monte (e.g., Akodon dolores, Calomys musculinus, Graomys griseoflavus) que están presentes en algunas muestras actuales recolectadas en cercanías del sitio.
A pesar de estas limitaciones cronoestratigráficas, se plantea un ambiente local estable que incluye un mosaico de estepas arbustivas, áreas abiertas con suelo desnudo y pedregoso, algo menos árido que en la actualidad. En coincidencia, el estudio de los granos de polen recuperados de los componentes estratigráficos de CH1 indicó el desarrollo de una estepa arbustiva y herbácea sin mayores cambios a lo largo de la secuencia (Pompei et al., 2012). Entre 13,8 y 9,5 ka cal. AP, el registro polínico de arbustos (Lycium, Schinus), arbustos en cojín (Ephedra, Nassauvia) y gramíneas que actualmente se encuentran a 1000 m por encima de CH1 indicó la presencia de estepas arbustivas y graminosas típicas de la estepa patagónica. Para este segmento temporal, este proxy paleoambiental expuso condiciones locales más frías que la actualidad, algo no manifiesto en el registro de micromamíferos. Mayor coincidencia entre ambos tipos de indicadores paleoambientales se observa entre 1,6 y 0,4 ka cal. AP, donde domina el polen de arbustos (Lycium, Schinus) y disminuye la proporción de gramíneas, lo que sugiere el desarrollo de una estepa arbustiva-graminosa de ecotono Monte y estepa patagónica similares a la actualidad (Pompei et al., 2012). Otras muestras de polen y de macrorrestos vegetales recuperadas de las heces de Lagidium provenientes de paleomadrigueras de CH cubren algunos períodos cronológicos no detectados en los componentes estratigráficos (Llano et al., 2020). Durante el Holoceno temprano (10,5-9,4 ka cal. AP), dominan los arbustos del Monte tales como Larrea, Lycium y Prosopis, junto con arbustos de la estepa patagónica (e.g., Adesmia, Berberis) y con arbustos del ecotono Monte-Patagonia (Ephedra,Schinus, Senecio). Las condiciones de aridez se intensifican hacia el 9,2 ka cal. AP y durante el Holoceno medio (6,5-5,5 ka cal. AP), cuando la vegetación del Monte (e.g.,, Larrea, Prosopis) prevalece sobre la vegetación patagónica y de transición entre ambos biomas. Hacia el Holoceno tardío, expresado en los intervalos 4,2-3,8 y 3,5-2,5 ka cal. AP, la codominancia de arbustos del Monte (Larrea, Lycium, Prosopis), arbustos y gramíneas patagónicos (e.g., Baccharis, Chuquiraga, Festuca, Jarava, Mulinum, Nassauvia, Poa) y arbustos del ecotono Monte-Patagonia (Ephedra, Schinus. Senecio) sugiere condiciones de humedad ligeramente mayores que en la actualidad (Llano et al., 2020).
Por otra parte, la estabilidad ambiental indicada para los componentes estratigráficos de CH1 coincide con el registro polínico de Mallín Vaca Lauquen para el segmento temporal de 17,8-8,9 ka 14C AP (Markgraf et al., 2009) y con los datos proporcionados por los excrementos de Lagidium recuperados de las paleomadrigueras del sitio Cuchillo Curá (centro del Neuquén) hacia 16,4 ka cal. AP (Hofreiter et al., 2003). Diversos estudios paleoambientales provenientes de Chile central, sur de Mendoza y Neuquén señalan condiciones de estabilidad para el Holoceno tardío, con variaciones menores asociadas a El Niño Oscilación del Sur, semejantes a la actualidad (Markgraf, 1983; Jenny et al., 2002; Whitlock et al., 2006; Le Quesne et al., 2009; Markgraf et al., 2009; Zárate et al., 2010; Bianchi y Ariztegui, 2012; de Jong et al., 2013; Frugone-Álvarez et al., 2020).
En sitios arqueológicos ubicados en ambientes áridos y semiáridos en áreas adyacentes al norte y al sur de CH1, también se detectaron condiciones de cierta estabilidad en los conjuntos de micromamíferos (e.g., Teta et al., 2005; Fernández et al., 2011a, 2012, 2015a, 2015b, 2016; Fernández, 2012; Pardiñas y Teta, 2013; Fernández y Pardiñas, 2018; López et al., 2021). No obstante, la Cueva Parque Diana, ubicada a 400 km al sudoeste de CH1, en un ambiente de bosque cercano a las estepas altoandinas, ofrece un registro de micromamíferos con variaciones en la secuencia temporal (Agnolín y Pérez, 2014). Allí se observan sigmodontinos asociados al bosque (Geoxus valdivianus) o al ecotono bosque-estepa en los niveles más antiguos (2,35-0,85 ka cal. AP) y un empobrecimiento taxonómico, posiblemente relacionado con aumentos de la aridez, hacia 0,66-0,55 ka cal. AP. Otro elemento común en los registros de micromamíferos de algunos de estos sitios resulta en el incremento exponencial de las especies de sigmodontinos oportunistas tales como C. musculinus y Oligoryzomys longicaudatus y, en ocasiones, la retracción regional o extinción de especies (e.g.,, Holochilus spp., Lestodelphys halli) en los conjuntos actuales, que reflejan un deterioro de los ambientes ocasionado, principalmente, por el pisoteo y sobrepastoreo de ovicaprinos ocurridos en las últimas centurias (e.g., Fernández, 2014; Teta et al., 2014; López et al., 2021). Sin embargo, en el área de CH1 no se detectaron cambios tan marcados ni extinciones recientes de especies, aspecto que sugiere que el impacto antrópico no fue tan significativo.
CONCLUSIÓN
A partir de la ampliación de la muestra de los micromamíferos y de la mayor resolución arqueológica de CH1 se alcanzó un conocimiento más detallado sobre la configuración de las comunidades de pequeños marsupiales y de pequeños roedores caviomorfos y sigmodontinos que habitaron en las inmediaciones del sitio desde el Pleistoceno tardío final hasta la actualidad.
Al conjunto ya conocido para el sitio, conformado por los roedores Akodon iniscatus, Ctenomys sp., Eligmodontiasp., Euneomys chinchilloides, Microcavia australis y Phyllotis xanthopygus y el marsupial Thylamys pallidior, se adicionan dos nuevos sigmodontinos (Abrothrix olivacea, Oligoryzomys longicaudatus) y un caviomorfo (Galea leucoblephara). Los nuevos datos no favorecen una mayor heterogeneidad ambiental hacia el Holoceno temprano (~9,5 ka cal. AP), como se había planteado previamente. Por el contrario, permiten inferir una marcada estabilidad del paisaje en general y, por lo tanto, de las condiciones climáticas durante el segmento cronológico representado. Se interpreta un ambiente caracterizado por un mosaico de estepas arbustivas, áreas abiertas de suelo desnudo y pedregoso, posiblemente un poco menos árido que en la actualidad por la ausencia de elementos típicos del Monte. Esta estabilidad contrasta con el registro de micromamíferos de otros yacimientos del Pleistoceno tardío/Holoceno de la cuenca superior del río Limay y sugiere respuestas bióticas diferenciales que podrían estar asociadas con la distancia a zonas de englazamiento.
Finalmente, la integración de líneas de evidencia paleoambiental destaca que los eventos climáticos más importantes, a nivel regional, durante el Pleistoceno tardío final (Inversión Climática Huelmo Mascardi) y el Holoceno (neoavances glaciarios), o bien no fueron de suficiente magnitud como para generar modificaciones significativas en las comunidades locales de micromamíferos donde se emplaza CH1 o no están representados los sedimentos portadores.

