










Servicios Personalizados
Revista

Articulo
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkRevista argentina de cardiología
versión On-line ISSN 1850-3748
Rev. argent. cardiol. v.74 n.3 Buenos Aires mayo/jun. 2006
Conexinas y sistema cardiovascular
Jorge E. Suárez1, 2, Alicia I. Bravo3
1 Servicio de Cardiología, Hospital I. G. A. Eva Perón. San Martín, Buenos Aires
2 Servicio de Cardiología, Hospital San Juan de Dios. Ramos Mejía, Buenos Aires
3 Unidad de Patología Molecular, Hospital I. G. A. Eva Perón. San Martín, Buenos Aires
Dirección para separatas:
Dr. Jorge E. Suárez
Av. Rivadavia 2431 - Entrada 2, Dpto. 1 -
(A1034ACD) Ciudad de Buenos Aires, Argentina.
e-mail: jesuarezar@yahoo.com.ar
RESUMEN
Las comunicaciones intercelulares son eventos biológicos esenciales en los organismos multicelulares, asociadas con el control del crecimiento y la diferenciación celular, la apoptosis, las respuestas adaptativas de células diferenciadas y la sincronización de funciones celulares. Las uniones intercelulares, conocidas como uniones gap, estructuralmente constituidas por conexinas, tienen una participación activa en estos procesos. A nivel cardiovascular, la comunicación célula a célula es indispensable, en condiciones normales, para la embriogénesis cardíaca, la transmisión del impulso eléctrico, la sincronización de la actividad contráctil cardíaca, la transmisión de señales reflejas vasculares, entre otras funciones biológicas, mientras que en condiciones patológicas, a causa de mutaciones genéticas heredadas o adquiridas, participan en el desarrollo de cardiopatías congénitas, arritmogénesis y remodelación eléctrica cardíaca, aterosclerosis e isquemia miocárdica, hipertensión arterial y remodelación miocárdica. La investigación en genética y biología molecular seguramente encontrará recursos para la prevención y el tratamiento de las distintas cardiopatías en las que la comunicación intercelular tiene un papel fisiopatológico.
Palabras clave: Arritmia; Conexinas; Biología molecular; Defectos congénitos; Endotelio; Genética; Miocardio.
Abreviaturas:
AV Auriculoventricular
CIC Comunicaciones intercelulares
Cx Conexina/s
D / kD Dalton / kilodalton
DDPR Dispersión en la duración de los períodos refractarios
DI Discos intercalados
mRNA Ácido ribonucleico mensajero
PAA Péptidos antiarrítmicos
PK Proteincinasa
pS picoSiemens
UG Uniones gap
Glosario:
Expresión: la información de un gen es convertida en un componente molecular para una estructura o una función.
Recambio (turnover): proceso por el cual proteínas u otras sustancias se metabolizan, destruyen y reemplazan por otras nuevas en forma continua.
Knockout: desarrollo de descendientes con carencia específica de un gen o de sus productos.
SUMMARY
Conexins and the Cardiovascular System
Intercellular communication is an essential biologic event of multicellular organisms and is associated with cell growth and differentiation control, apoptosis, adaptive responses of differentiated cells and the synchronization of cellular functions. A kind of intercellular junctions known as gap junctions, actively participate in these processes and are structurally made of conexins. From a cardiovascular viewpoint, cell to cell communication, under normal conditions, is essential in cardiac embryogenesis, electrical impulse transmission, synchronization of cardiac contractile activity, transmission of vascular reflex signals and other biological functions. Under pathological conditions, either by inherited or acquired genetic mutation, intercellular communications participate in the development of congenital cardiopathies, arrhythmogenesis and electrical remodelling, atherosclerosis and myocardial ischemia, arterial hypertension and myocardial remodelling. Genetic and molecular research will probably find the means to prevent and treat the different cardiopathies where intercellular communication plays a physiopathological role.
Key words: Arrhythmia; Connexins; Molecular biology; Congenital defects; Endothelium; Genetics; Myocardium.
INTRODUCCIÓN
El desarrollo productivo de las sociedades humanas o de otro origen requiere comunicación permanente. El entendimiento recíproco, la transferencia de conocimientos y de experiencia, la transferencia de órdenes o la coordinación de actividades conjuntas precisan la transmisión de señales entre los componentes de la sociedad. Las células vivientes se comportan como "microsociedades" que necesitan, precisamente, comunicación permanente. (1) Para conocer cómo se organizan las "sociedades celulares" en condiciones normales o patológicas, se deben "interceptar" esas señales, "traducir" el mensaje y "descifrar" los objetivos de la comunicación intercelular (CIC). Ésta es la gigantesca tarea que hoy enfrenta la biología molecular.
Cada célula mantiene su individualidad funcional, aunque la funcionalidad tisular depende de la coordinación armónica con otras células. Los organismos han desarrollado múltiples estrategias para alcanzar estos propósitos, que incluyen interacciones a distancia mediadas por mecanismos neuronales o endocrinos o interacciones cercanas mediadas por contacto físico directo, célula a célula, por medio de canales especializados. Una forma de CIC involucra a verdaderos poros o canales acuosos de la membrana plasmática entre células contiguas, denominados canales intercelulares, uniones comunicantes, uniones gap o gap junctions.
ESTRUCTURA Y FUNCIÓN DE LAS UNIONES GAP
Las uniones gap (UG) son sistemas de comunicación de la membrana celular que une los citoplasmas de células adyacentes, constituidos por canales por los cuales los organismos multicelulares se proveen de un sencillo método de sincronizar respuestas mediante intercambio directo de iones, pequeñas moléculas de peso molecular no superior a 1.000-1.200 dalton, como Ca++, cAMP, glutatión y macromoléculas como nucleótidos, azúcares y amionoácidos. Estas funciones intercelulares permiten actividades de coordinación rápida, como la contracción del músculo cardíaco o la transmisión de señales eléctricas neuronales. La CIC es esencial en procesos más lentos, relacionados con el crecimiento, el desarrollo y la muerte celular, como desarrollo embrionario, acoplamiento eléctrico, transporte metabólico, apoptosis, diferenciación celular y homeostasis tisular. Las UG están constituidas por proteínas transmembrana denominadas conexinas (Cx), que en grupos de seis rodean un poro de la membrana formando un "hemicanal" o conexón (Figura 1). Dos conexones de células adyacentes se alinean y anclan entre sí, en el espacio extracelular, para formar una UG o canal intercelular. Cantidades variables de UG se agrupan en segmentos de la membrana y conforman placas unionales. Actualmente se encuentran identificadas 20 Cx en el genoma humano y en el ratón. (2) La familia de las Cx puede encontrarse en todos los tipos de células de los vertebrados, en tanto que proteínas similares se han clonado de especies vegetales. En los vertebrados no existen otros canales que provean conductos similares para intercambio por difusión de iones y pequeñas moléculas, con diámetros suficientemente grandes para permitir el pasaje de moléculas de hasta 1.200 D. Los invertebrados utilizan una familia diferente de moléculas de conexión, las inexinas, que comparten similitudes estructurales con las Cx de los vertebrados, pero sus secuencias no son idénticas. (3)

Fig. 1. La conexina agrupada en seis unidades conforma un conexón, que perfora la membrana plasmática. Dos conexones, acoplados en el espacio extracelular, forman una unión gap, la que, rodeando el poro acuoso, conecta los citoplasmas de dos células vecinas.
Las Cx contienen regiones especializadas que les permiten fijarse en la doble capa lipídica de la membrana celular, poseen segmentos extracelulares de fijación al conexón opuesto y regiones intracelulares con funciones reguladoras. Estas regiones son básicamente dos terminales citoplasmáticas (Figura 2), amino una (N) y carboxílica la otra (C), un bucle citoplasmático (CL), dos bucles extracelulares (E1, E2) y cuatro dominios transmembrana (M1 a M4). El tercer dominio transmembrana (M3) de cada Cx forma el límite externo del canal (Figura 3) y es el segmento en mayor contacto con el poro acuoso de la UG. (4) Las Cx difieren entre sí en la secuencia de los bucles intracelulares y los terminales carboxílicos. La permeabilidad de la UG depende de los tipos de Cx constitutivas y de la carga de la molécula que transita el poro. La conductancia de la UG de la célula miocárdica se ubica entre los 20 a 200 pS, dependiente de la Cx. La conductancia es modulada por el voltaje transgap, por iones H+, Ca2+, el estado de fosforilación de la Cx y por la composición de los ácidos grasos extracelulares. Distintos factores modulan la expresión de las diferentes Cx. El tiempo de recambio proteico a nivel de las Cx (degradación y resíntesis -turnover-) es notablemente rápido. En el corazón de rata adulta, la vida media de las Cx es de 1,3 horas. (5)

Fig. 2. Estructura básica de la conexina: el esquema representa la cadena de conexina (en gris) transcurriendo la membrana celular. E1, E2: bucles extracelulares. M1, M2, M3, M4: dominios transmembrana. CL: bucle citoplasmático. N, C: terminales amino y carboxílico citoplasmáticos.
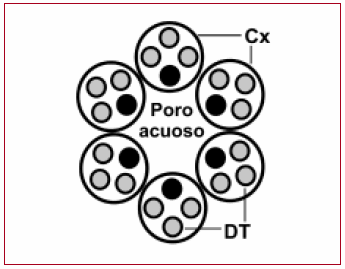
Fig. 3. Diagrama del corte transversal de un conexón: el poro acuoso rodeado por seis conexinas y sus cuatro dominios transmembrana. El dominio M3 (círculo negro lleno) conforma los límites externos del poro. Cx: conexina. DT: dominio transmembrana.
DISTRIBUCIÓN DE LAS DISTINTAS CONEXINAS
Existen Cx en prácticamente todos los tejidos, con excepción de los glóbulos rojos y las células del músculo esquelético. Se usan dos nomenclaturas para clasificarlas: una las divide en dos grupos, alfa y beta, de acuerdo con similitudes en sus secuencias proteicas y la otra las agrupa por su peso molecular, en kilodalton (kD), y es la más usada. En el órgano de Corti del oído interno se expresa principalmente Cx26 (beta-2). Mutaciones espontáneas de su gen se asocian con la sordera no sindrómica hereditaria autosómica recesiva, la forma más común de sordera hereditaria. En la piel, el oído, la placenta y el ojo se expresa de manera predominante Cx31 (beta-3). Sus mutaciones se asocian con la eritroqueratodermia variabilis y una forma autosómica dominante de deterioro auditivo. En el hígado, el páncreas exocrino, el sistema nervioso central y el epitelio del sistema gastrointestinal se expresa Cx32 (beta- 1). En los seres humanos, Cx32 es crítica para las células de Schwann de los nervios periféricos. La mutación de su gen da lugar a una forma de neuropatía hereditaria denominada enfermedad de Charcot-Marie-Tooth ligada al cromosoma X. En el cerebro, el corazón, el útero, el ovario y las células endoteliales de los vasos sanguíneos se expresa Cx37 (alfa-4). Cx46 (alfa 3) y Cx50 (alfa 8) tienen expresión en el cristalino uniendo las células epiteliales en un sincitio funcional (Cx46) y manteniendo su transparencia (Cx50). En ratas, sus mutaciones producen distintas formas de catarata.
DISTRIBUCIÓN CARDÍACA DE LAS CONEXINAS
En el corazón predomina Cx43, con excepción del nodo AV y partes del sistema de conducción. En mamíferos, Cx40 se expresa en las aurículas, en el sistema de conducción intraventricular proximal y en el endotelio vascular coronario. (6) El miocardio ventricular adulto contiene Cx43, localizada principalmente a nivel de los discos intercalados (DI). La disposición geométrica de los DI así como el tamaño y el número de las UG parecen ser determinantes de la conducción anisotrópica que caracteriza al miocardio ventricular. (7) La expresión de Cx45 estaría limitada al tejido nodal y al sistema de conducción proximal. No se han detectado otras conexinas a nivel cardíaco.
PAPEL DE LAS CONEXINAS EN LA EMBRIOGÉNESIS CARDÍACA
Cecilia W. Lo analizó la expresión genética de la UG en el ratón y sugirió un papel significativo de Cx43 en el desarrollo embriogénico. (8) Estudios en ratones transgénicos o con mutaciones provocadas de Cx43 destacan la actividad de las UG en el desarrollo cardíaco. (9) Se observaron defectos cardíacos que afectaban el área conotroncal y el ventrículo derecho en ratones con knockout de Cx43, lo cual implica que su interferencia afecta a las estructuras cardíacas derivadas de la cresta neural. (10) Mediante cultivo de células de la cresta neural, Huang y colaboradores demostraron una tasa incrementada de migración celular en embriones con sobreexpresión de Cx43, aunque reducida en tejidos con knockout de dicha Cx. Se sugiere que las células de la cresta neural cardíaca pueden modular el crecimiento y el desarrollo de tejido cardíaco no originado en esa cresta. Aunque las células de la cresta neural migran como células individuales, hay evidencia de que estas células se comunican entre sí durante el proceso de migración. (11)
Existe actividad de Cx43 en la CIC de otro tipo de células en desarrollo: los mioblastos esqueléticos, trasplantados para transdiferenciarse en cardiomiocitos, requerirían conexiones a cardiomiocitos vecinos por medio de Cx43. La transdiferenciación sería favorecida por la transmisión, a través de las UG, del estímulo eléctrico y la ritmicidad de la fuerza contráctil de los cardiomiocitos vecinos. (12) En la reciente "2005 International Gap Junction Conference", Shasha Liu y colaboradores sugieren que la falta de Cx43 en la cresta neural da por resultado la deslaminación anormal y la migración aberrante de las células del tubo neural, con eventual incorporación de estas células en el tracto de salida cardíaco. (13) En el desarrollo y la conformación de las arterias coronarias intervendría Cx43, papel que probablemente involucra a las células de la cresta neural cardíaca y proepicárdicas. (14, 15)
Durante el desarrollo cardíaco de roedores, en el decimocuarto día embrionario, Cx40 alcanza su máxima expresión. Posteriormente queda restringida a las aurículas y el sistema de conducción ventricular. La única Cx conocida en el primitivo corazón murino antes del noveno día embrionario es Cx45. En ratones, el déficit de Cx45 se asoció con muerte embrionaria en el día 10, con inicio de contracciones cardíacas pero con presentación de bloqueo cardíaco 24 horas después. (16)
CONDUCCIÓN ELÉCTRICA CARDÍACA Y ARRITMOGÉNESIS
La conducción eléctrica cardíaca requiere una propagación eficiente de la carga eléctrica de célula a célula. Las propiedades fisiológicas esenciales que facilitan este proceso son la excitabilidad de membrana y la CIC mediada por uniones gap.
En las células de la musculatura lisa vascular, endotelial y del miocardio de los vertebrados se detectan Cx40, Cx43 y sus mRNA. (17) Durante el desarrollo temprano, en el miocardio ventricular se expresa Cx43 en forma gradual y alcanza su máximo luego del nacimiento. En el nodo AV y en el haz de His colindante (Figura 4) se encuentra Cx45, que forma UG de baja conductancia sensibles al voltaje. Por el contrario, Cx40 forma canales de alta conductancia y se expresa en los tejidos de rápida conducción del sistema His-Purkinje y es encapsulado por expresiones de Cx45. (8) En fibras distales de Purkinje, Cx40 se coexpresa con Cx43. Así, parecería que Cx45, a nivel del nodo AV y el haz de His, provee una vaina aislante del miocardio circundante, Cx40 provee la alta velocidad central y Cx43 facilitaría el acoplamiento al miocito ventricular. Tanto Cx40 como Cx43 tendrían efectos aditivos en la septación cardíaca, demostrado en ratas embrionarias con ausencia homocigota de uno de los genes y heterocigota del otro. (18, 19) El desarrollo de ratones con técnica de knockout de Cx45 demostró que esta Cx participa en la conformación de las almohadillas endocárdicas y del sistema de conducción durante las primeras etapas del desarrollo cardíaco (20) y es, además, la primera Cx detectable durante el desarrollo del septum interauricular y el sistema de conducción proximal en ratones embrionarios. (21) Las modificaciones de la expresión que afecten la conductancia de las UG podría deteriorar la conducción y originar arritmias. (22) Ratones con carencia transgénica de Cx40 desarrollan retardo de la conducción en el nodo AV (23) y en el sistema de conducción intraventricular. (24)
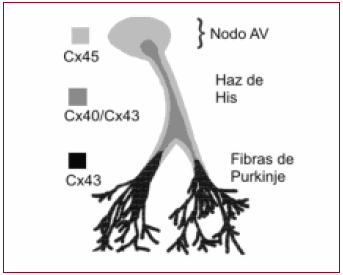
Fig. 4. Distribución de las conexinas del sistema de conducción: Cx45 en la región nodal AV y envainando el haz de His y ramas. La rápida conducción de Cx40, predominante en el sistema de conducción hisiano. Cx43 en la interfase Purkinje-miocito.
El estado de fosforilación de las Cx es determinante de su funcionalidad. La fosforilación de las Cx depende de un balance entre proteincinasas (PK) y proteinfosfatasas. La PK A y la PK C son activadas por receptores ß y α1, respectivamente. La actividad α1 elevada a nivel miocárdico, en pacientes con insuficiencia cardíaca, podría ser un mecanismo de afectación de la funcionalidad de las Cx. Chen y colaboradores (22) publicaron resultados de desfosforilación crónica de las Cx cardíacas, sin pérdida de expresión, en asociación con defectos de la conducción cardíaca, como PR prolongado y/o bloqueo cardíaco de alto grado.
Kizana y colaboradores, en una publicación reciente, señalan la posibilidad de producir genéticamente fibroblastos modificados para conducir el estímulo mediante acoplamiento eléctrico, proceso en el que intervienen UG de Cx43. (25) La transferencia genética focal ha logrado avances en la creación de marcapasos biológicos (26-28) e inducción de retardo en la conducción nodal AV, (29) hechos que marcarían el inicio de una era de "marcapasos y catéteres biológicos".
Las anormalidades de las UG pueden determinar arritmias cardíacas en el corazón enfermo. Gutstein demuestra en ratones la letalidad de la inactivación de Cx43 cardíaca inducida en etapa embrionaria. Todos los ratones con knockout de Cx43 sufrieron muerte súbita por arritmia ventricular a los dos meses de vida. (30) Las referencias al papel de las Cx en la génesis o la participación en las arritmias cardíacas fue incesante en los últimos cinco años y abarca entidades como arritmias durante isquemia cardíaca, (31-33) muerte súbita, (34, 35) arritmias auriculares, (36-38) displasia arritmogénica del ventrículo derecho, (39) bloqueo cardíaco (40-42) y conducción por vías accesorias en el síndrome de Wolff-Parkinson- White. (43)
ANTIARRÍTMICOS ESPECÍFICOS
El descubrimiento de un nuevo grupo de drogas con propiedades antiarrítmicas, denominadas péptidos antiarrítmicos (PAA) por Aonuma y colaboradores, (44) impulsa experiencias basadas en la posibilidad de corregir las anormalidades funcionales de las UG como sustrato fisiopatológico de arritmias auriculares y ventriculares. El péptido descubierto es una cadena endógena (H-Gli-Pro-4Hip-Gli-Ala-Gli-OH), aislado de aurícula bovina. (45) Posteriormente se sintetizaron compuestos peptídicos, de los cuales el más investigado es AAP10. Su capacidad para aumentar la conductancia de las UG cardíacas se propone como mecanismo de una nueva generación de drogas antiarrítmicas. (46) El uso de AAP10 se vio limitado por su baja estabilidad plasmática. Un nuevo compuesto, ZP123, demostró, en ratas y en seres humanos, una estabilidad 1.700 veces más prolongada, una depuración 140 veces más lenta y una eficacia máxima con dosis 10 veces menores que AAP10. En condiciones control, ambas drogas tienen poco efecto sobre la dispersión en la duración de los períodos refractarios (DDPR). En cambio, en presencia de isquemia hipopotasémica, el incremento de DDPR, considerada el principal sustrato proarrítmico, es prevenido por ZP123. (47) Esta droga, denominada genéricamente rotigaptide o GAP-486, previene arritmias ventriculares y provoca, además, reducción del tamaño del infarto en presencia de injuria en la isquemia/reperfusión miocárdica en perros. (48) Los PAA pueden aumentar la conductancia de las UG en otros territorios, como ocurre con el rotigaptide sobre cultivos de osteoblastos humanos, o en vivo, en ratas, al prevenir la pérdida ósea inducida por ovariectomía. (49) En junio de 2005 se inició la fase II del ensayo clínico, doble ciego controlado con placebo, multicéntrico internacional, de la eficacia del rotigaptide en pacientes con enfermedad cardíaca crónica y con infarto agudo de miocardio. Otros dos ensayos doble ciego contra placebo evaluarán las propiedades electrofisiológicas agudas de GAP-486 y su capacidad para prevenir la inducibilidad de taquiarritmia ventricular. La información de estos ensayos está disponible en ClinicalTrials. gov, un servicio de los U.S. National Institutes of Health. (50-52)
CONEXINAS Y PARED VASCULAR
A nivel vascular, en el endotelio y en la musculatura lisa de la pared existen UG y se ha identificado la presencia de Cx37, Cx40, Cx43 y Cx45. (53) Las UG vasculares son una vía de comunicación importante en los ejes longitudinal y transversal de los vasos. Las proteínas de las UG no actúan en forma aislada para mantener las CIC; su funcionalidad es dependiente de las interrelaciones que mantienen con otras proteínas, como cadherinas, caveolinas u otras proteínas mecánicas y de adhesión al citoesqueleto celular. (54, 55) Cx40 tiene funciones en el control del tono vasomotor, como lo demuestra la transmisión, muy rápida y no decremental en el eje axial, de reflejos vasomotores inducidos por estimulación eléctrica o acetilcolina. (56, 57) Un capítulo especial de la función de las UG a nivel de los vasos es su posible participación en la hipertensión arterial. (58) Dos excelentes y recomendables trabajos de 2004, relativos a las características de las UG a nivel vascular, son ofrecidos por Xavier F. Figueroa, de la Universidad de Virginia, (59) y por Cor de Wit, de la Universidad de Lübeck. (60)
En los grandes conductos arteriales, las Cx se expresan de manera diferencial en los distintos tipos de células de la pared vascular. Cx43 es la principal Cx del músculo liso vascular y Cx40 se encuentra principalmente en el endotelio. También Cx37 se expresa en el endotelio, aunque en menor proporción que Cx40. En pequeñas cantidades, en el músculo liso vascular, también se encuentra Cx45. (61)
ATEROSCLEROSIS
En la generación de aterosclerosis, las interacciones celulares paracrinas son eventos fundamentales. Kwak y colaboradores (62) sugieren funciones importantes de las CIC basadas en Cx37, Cx40 y Cx43 durante el desarrollo de placas ateroscleróticas en aortas humanas y de ratón, en las que se expresan diferencialmente en células asociadas con ateromas con lesiones. Una expresión significativa de mRNA para Cx43, componente de las uniones gap de células vasculares, se demostró en macrófagos espumosos (foam cells) de arterias carotídeas ateroscleróticas humanas, lo cual indicaría que condiciones tisulares específicas inducen la expresión de esta conexina. (63) Se ha demostrado una vinculación molecular entre inflamación y CIC mioendotelial de la pared de los vasos sanguíneos. (64) Polimorfismos en los genes de Cx37 podrían ser un marcador genético en la valoración del desarrollo de la placa arteriosclerótica, particularmente en grupos de riesgo cardiovascular. (65)
ISQUEMIA MIOCÁRDICA
La isquemia miocárdica afecta la CIC, así como la distribución y la localización de las Cx. (66) La hipoxia miocárdica determina desfosforilación de Cx43 y su redistribución desde los DI a toda la membrana plasmática y es uno de los primeros signos de injuria miocárdica luego de hipoxia. (67) Durante la reperfusión, la CIC mediaría en la propagación, célula a célula, de hipercontractura y muerte celular, con evidencias que explicarían la continuidad de la necrosis en bandas de contracción e influirían sobre el tamaño final del infarto. (68) El precondicionamiento isquémico como mecanismo para la limitación del tamaño del infarto induciría una reducción en la permeabilidad de las UG durante la isquemia, presumiblemente por forforilación de Cx43 mediada por PK C. (69) De esta forma, las UG tendrían una actividad significativa en la fisiopatología de la injuria y muerte celular durante isquemia-reperfusión miocárdica y serían potenciales objetivos de estrategias terapéuticas cardioprotectoras.
CONEXINAS EN EL CORAZÓN HIPERTRÓFICO E INSUFICIENTE
La remodelación hipertrófica miocárdica en respuesta a una sobrecarga de trabajo, injuria o enfermedad intrínseca, es inicialmente adaptativa. La prolongación de la sobrecarga lleva a diversos cambios deletéreos. Uno de ellos es la remodelación de las UG, básicamente de Cx43, cuya alteración se correlaciona con el grado de hipertrofia. (70-72) En pacientes con miocardiopatía dilatada, isquémica o inflamatoria se observa redistribución de Cx43 desde los DI a las superficies laterales del miocito ventricular izquierdo (lateralización) en las áreas con daño estructural. En el ventrículo izquierdo de todos los corazones insuficientes, la cantidad de Cx43 por miocito fue significativamente más baja que en el corazón normal. (73) La reducción de los niveles de Cx43 es acompañada por una reducción de sus mRNA, hecho que sugiere que la menor expresión de Cx43 en el corazón hipertrófico es regulada a nivel transcripcional. (74)
En la asincronía ventricular izquierda, si bien la expresión total de Cx43 no está alterada, en las células de las áreas miocárdicas con activación retrasada se encuentra lateralización de Cx43. (75) Por otro lado, la remodelación de las Cx en la hipertrofia/dilatación ventricular conllevan el riesgo de arritmias ventriculares graves. (76, 77)
CONCLUSIONES
La comunicación intercelular por intermedio de uniones gap es un proceso biológico cuya importancia en el campo de la medicina se devela día a día, destacándose el papel particular de las moléculas más activas de estos canales, las conexinas. Se están vislumbrando cambios terapéuticos revolucionarios en el campo de la medicina genética y molecular.
Aunque es largo el camino por recorrer, las respuestas necesarias para la aplicabilidad clínica de los resultados observados en las investigaciones de las comunicaciones intercelulares finalmente tendrán su fruto.
1. Perbal B. Communication is the key. Cell Commun Signal 2003;1:3. [ Links ]
2. Willecke K, Eiberger J, Degen J, Eckardt D, Romualdi A, Guldenagel M, et al. Structural and functional diversity of connexin genes in the mouse and human genome. Biol Chem 2002;383:725-37. [ Links ]
3. Phelan P, Bacon JP, Davies JA, Stebbings LA, Todman MG, Avery L, et al. Innexins: a family of invertebrate gap-junction proteins. Trends Genet 1998;14:348-9. [ Links ]
4. Skerrett M, Kasperek E, Cao FL, Shin JH, Aronowitz J, Ahmed S, et al. Application of SCAM (substituted cysteine accessibility method) to gap junction intercellular channels. Cell Commun Adhes 2001; 8:179-85. [ Links ]
5. Beardslee MA, Laing JG, Beyer EC, Saffitz JE. Rapid turnover of connexin43 in the adult rat heart. Circ Res 1998;83:629-35. [ Links ]
6. Gros D, Jarry-Guichard T, Ten Velde I, de Maziere A, van Kempen MJ, Davoust J, et al. Restricted distribution of connexin40, a gap junctional protein, in mammalian heart. Circ Res 1994;74:839-51. [ Links ]
7. Spach MS, Heidlage JF. The stochastic nature of cardiac propagation at a microscopic level. Electrical description of myocardial architecture and its application to conduction. Circ Res 1995;76:366-80. [ Links ]
8. Lo CW. The role of gap junction membrane channels in development. J Bioenerg Biomembr 1996;28:379-85. [ Links ]
9. Waldo KL, Lo CW, Kirby ML. Connexin 43 expression reflects neural crest patterns during cardiovascular development. Dev Biol 1999; 208:307-23. [ Links ]
10. Huang GY, Wessels A, Smith BR, Linask KK, Ewart JL, Lo CW. Alteration in connexin 43 gap junction gene dosage impairs conotruncal heart development. Dev Biol 1998;198:32-44. [ Links ]
11. Huang GY, Cooper ES, Waldo K, Kirby ML, Gilula NB, Lo CW. Gap junction-mediated cell-cell communication modulates mouse neural crest migration. J Cell Biol 1998;143:1725-34. [ Links ]
12. Iijima Y, Nagai T, Mizukami M, Matsuura K, Ogura T, Wada H, et al. Beating is necessary for transdifferentiation of skeletal musclederived cells into cardiomyocytes. FASEB J 2003;17:1361-3. [ Links ]
13. Liu S, Liu F, Shah B, Amand TS, Epstein JA, Gutstein DE. Loss of connexin43 results in abnormal delamination and migration of neural tube cells. 2005 International Gap Junction Conference. August, Whistler, Canada. Session 9. Abstract. [ Links ]
14. Li WE, Waldo K, Linask KL, Chen T, Wessels A, Parmacek MS, et al. An essential role for connexin43 gap junctions in mouse coronary artery development. Development 2002;129:2031-42. [ Links ]
15. Walker DL, Vacha SJ, Kirby ML, Lo CW. Connexin43 deficiency causes dysregulation of coronary vasculogenesis. Dev Biol 2005; 284:479-98. [ Links ]
16. Kumai M, Nishii K, Nakamura K, Takeda N, Suzuki M, Shibata Y. Loss of connexin45 causes a cushion defect in early cardiogenesis. Development 2000;127:3501-12. [ Links ]
17. Little TL, Beyer EC, Duling BR. Connexin 43 and connexin 40 gap junctional proteins are present in arteriolar smooth muscle and endothelium in vivo. Am J Physiol 1995;268:H729-39. [ Links ]
18. Reaume AG, de Sousa PA, Kulkarni S, Langille BL, Zhu D, Davies TC, et al. Cardiac malformation in neonatal mice lacking connexin43. Science 1995;267:1831-4. [ Links ]
19. Van Kempen MJ, Vermeulen JL, Moorman AF, Gros D, Paul DL, Lamers WH. Developmental changes of connexin40 and connexin43 mRNA distribution patterns in the rat heart. Cardiovasc Res 1996; 32:886-900. [ Links ]
20. Kumai M, Nishii K, Nakamura K, Takeda N, Suzuki M, Shibata Y. Loss of connexin45 causes a cushion defect in early cardiogenesis. Development 2000;127:3501-12. [ Links ]
21. Coppen SR, Gourdie RG, Severs NJ. Connexin45 is the first connexin to be expressed in the central conduction system of the mouse heart. Experimental and Clinical Cardiology 2001;6:17-23. [ Links ]
22. Chen HH, Baty CJ, Maeda T, Brooks S, Baker LC, Ueyama T, et al. Transcription enhancer factor-1-related factor-transgenic mice develop cardiac conduction defects associated with altered connexin phosphorylation. Circulation 2004;110:2980-7. [ Links ]
23. Simon AM, Goodenough DA, Paul DL. Mice lacking connexin40 have cardiac conduction abnormalities characteristic of atrioventricular block and bundle branch block. Curr Biol 1998;8:295-8. [ Links ]
24. Tamaddon HS, Vaidya D, Simon AM, Paul DL, Jalife J, Morley GE. High-resolution optical mapping of the right bundle branch in connexin40 knockout mice reveals slow conduction in the specialized conduction system. Circ Res 2000;87:929-36. [ Links ]
25. Kizana E, Ginn SL, Allen DG, Ross DL, Alexander IE. Fibroblasts can be genetically modified to produce excitable cells capable of electrical coupling. Circulation 2005;111:394-8. [ Links ]
26. Qu J, Plotnikov AN, Danilo P Jr, Shlapakova I, Cohen IS, Robinson RB, et al. Expression and function of a biological pacemaker in canine heart. Circulation 2003;107:1106-9. [ Links ]
27. Plotnikov AN, Sosunov EA, Qu J, Shlapakova IN, Anyukhovsky EP, Liu L, et al. Biological pacemaker implanted in canine left bundle branch provides ventricular escape rhythms that have physiologically acceptable rates. Circulation 2004;109:506-12. [ Links ]
28. Lin G, Cai J, Jiang H, Shen H, Jiang X, Yu Q, et al. Biological pacemaker created by fetal cardiomyocyte transplantation. J Biomed Sci 2005;12:513-9. [ Links ]
29. Donahue JK, Heldman AW, Fraser H, McDonald AD, Miller JM, Rade JJ, et al. Focal modification of electrical conduction in the heart by viral gene transfer. Nat Med 2000;6:1395-8. [ Links ]
30. Gutstein DE, Morley GE, Tamaddon H, Vaidya D, Schneider MD, Chen J, et al. Conduction slowing and sudden arrhythmic death in mice with cardiac-restricted inactivation of connexin43. Circ Res 2001;88:333-9. [ Links ]
31. Cascio WE, Yang H, Muller-Borer BJ, Johnson TA. Ischemiainduced arrhythmia: the role of connexins, gap junctions, and attendant changes in impulse propagation. J Electrocardiol 2005;38:55-9. [ Links ]
32. De Groot JR, Coronel R. Acute ischemia-induced gap junctional uncoupling and arrhythmogenesis. Cardiovasc Res 2004;62:323-34. [ Links ]
33. Garcia-Dorado D, Rodriguez-Sinovas A, Ruiz-Meana M. Gap junction-mediated spread of cell injury and death during myocardial ischemia-reperfusion. Cardiovasc Res 2004;61:386-401. [ Links ]
34. Betsuyaku T, Nnebe NS, Sundset R, Patibandla S, Krueger CM, Yamada KA. Overexpression of cardiac connexin45 increases susceptibility to ventricular tachyarrhythmias in vivo. Am J Physiol Heart Circ Physiol 2006;290:H163-71. [ Links ]
35. Poelzing S, Rosenbaum DS. Altered connexin43 expression produces arrhythmia substrate in heart failure. Am J Physiol Heart Circ Physiol 2004;287:H1762-70. [ Links ]
36. Kanagaratnam P, Peters NS. Conduction, gap junctions, and atrial fibrillation: an eternal triangle? Heart Rhythm 2004;1:746-9. [ Links ]
37. Wetzel U, Boldt A, Lauschke J, Weigl J, Schirdewahn P, Dorszewski A, et al. Expression of connexins 40 and 43 in human left atrium in atrial fibrillation of different aetiologies. Heart 2005;91:166-70. [ Links ]
38. Ausma J, van der Velden HM, Lenders MH, van Ankeren EP, Jongsma HJ, Ramaekers FC, et al. Reverse structural and gapjunctional remodeling after prolonged atrial fibrillation in the goat. Circulation 2003;107:2051-8. [ Links ]
39. Kaplan SR, Gard JJ, Protonotarios N, Tsatsopoulou A, Spiliopoulou C, Anastasakis A, et al. Remodeling of myocyte gap junctions in arrhythmogenic right ventricular cardiomyopathy due to a deletion in plakoglobin (Naxos disease). Heart Rhythm 2004;1:3-11. [ Links ]
40. van Rijen HV, van Veen TA, van Kempen MJ, Wilms-Schopman FJ, Potse M, Krueger O, et al. Impaired conduction in the bundle branches of mouse hearts lacking the gap junction protein connexin40. Circulation 2001;103:1591-8. [ Links ]
41. Saffitz JE, Schuessler RB. Connexin-40, bundle-branch block, and propagation at the Purkinje-myocyte junction. Circ Res 2000; 87:835-6. [ Links ]
42. Bevilacqua LM, Simon AM, Maguire CT, Gehrmann J, Wakimoto H, Paul DL, et al. A targeted disruption in connexin40 leads to distinct atrioventricular conduction defects. J Interv Card Electrophysiol 2000;4:459-67. [ Links ]
43. Peters NS, Rowland E, Bennett JG, Green CR, Anderson RH, Severs NJ. The Wolff-Parkinson-White syndrome: the cellular substrate for conduction in the accessory atrioventricular pathway. Eur Heart J 1994;15:981-7. [ Links ]
44. Aonuma S, Kohama Y, Akai K, Komiyama Y, Nakajima S, Wakabayashi M, et al. Studies on heart. XIX. Isolation of an atrial peptide that improves the rhythmicity of cultured myocardial cell clusters. Chem Pharm Bull (Tokyo) 1980;28:3332-9. [ Links ]
45. Aonuma S, Kohama Y, Makino T, Fujisawa Y. Studies of heart. XXI. Amino acid sequence of antiarrhythmic peptide (AAP) isolated from atria. J Pharmacobiodyn 1982;5:40-8. [ Links ]
46. Muller A, Schaefer T, Linke W, Tudyka T, Gottwald M, Klaus W, et al. Actions of the antiarrhythmic peptide AAP10 on intercellular coupling. Naunyn Schmiedebergs Arch Pharmacol 1997; 356:76-82. [ Links ]
47. Kjolbye AL, Knudsen CB, Jepsen T, Larsen BD, Petersen JS. Pharmacological characterization of the new stable antiarrhythmic peptide analog Ac-D-Tyr-D-Pro-D-Hyp-Gly-D-Ala-Gly-NH2 (ZP123): in vivo and in vitro studies. J Pharmacol Exp Ther 2003; 306:1191-9. [ Links ]
48. Hennan JK, Swillo RE, Morgan GA, Keith Jr JC, Schaub RG, Smith RP, et al. Rotigaptide (ZP123) Prevents Spontaneous Ventricular Arrhythmias And Reduces Infarct Size During Myocardial Ischemia/Reperfusion Injury In Open-Chest Dogs. J Pharmacol Exp Ther 2005 Dec 12; [Epub ahead of print]. [ Links ]
49. Jorgensen NR, Teilmann SC, Henriksen Z, Meier E, Hansen SS, Jensen JE, et al. The antiarrhythmic peptide analog rotigaptide (ZP123) stimulates gap junction intercellular communication in human osteoblasts and prevents decrease in femoral trabecular bone strength in ovariectomized rats. Endocrinology 2005;146:4745-54. [ Links ]
50. ClinicalTrials.gov. US National Institutes of Health. National Library of Medicine. Study Evaluating GAP-486 With Non-Sustained Ventricular Tachycardia. Accedido en enero 2006 en http://www.clinicaltrials.gov/show/NCT00124891 [ Links ]
51. ClinicalTrials.gov. US National Institutes of Health. National Library of Medicine. Study to Characterize the Acute Electrophysiologic Properties, Safety and Tolerability of GAP-486. Accedido en enero 2006 en http://www.clinicaltrials.gov/ct/show/NCT00137293?order=1 [ Links ]
52. ClinicalTrials.gov. US National Institutes of Health. National Library of Medicine. A Study of the Effects GAP-486 on Ventricular Tachyarrhythmia Induction. Accedido enero 2006 en http://www.clinicaltrials.gov/ct/show/NCT00137332?order=2 [ Links ]
53. Evans WH, Martin PE. Gap junctions: structure and function (Review). Mol Membr Biol 2002;19:121-36. [ Links ]
54. Li J, Patel VV, Kostetskii I, Xiong Y, Chu AF, Jacobson JT, et al. Cardiac-specific loss of N-cadherin leads to alteration in connexins with conduction slowing and arrhythmogenesis. Circ Res 2005;97:474-81. [ Links ]
55. Ismat FA, Zhang M, Kook H, Huang B, Zhou R, Ferrari VA, et al. Homeobox protein Hop functions in the adult cardiac conduction system. Circ Res 2005;96:898-903. [ Links ]
56. de Wit C, Roos F, Bolz SS, Kirchhoff S, Kruger O, Willecke K, et al. Impaired conduction of vasodilation along arterioles in connexin40- deficient mice. Circ Res 2000;86:649-55. [ Links ]
57. Figueroa XF, Paul DL, Simon AM, Goodenough DA, Day KH, Damon DN, et al. Central role of connexin40 in the propagation of electrically activated vasodilation in mouse cremasteric arterioles in vivo. Circ Res 2003;92:793-800. [ Links ]
58. Rummery NM, McKenzie KU, Whitworth JA, Hill CE. Decreased endothelial size and connexin expression in rat caudal arteries during hypertension. J Hypertens 2002;20:247-53. [ Links ]
59. Figueroa XF, Isakson BE, Duling BR. Connexins: gaps in our knowledge of vascular function. Physiology (Bethesda) 2004;19:277-84. [ Links ]
60. De Wit C. Connexins pave the way for vascular communication. News Physiol Sci 2004;19:148-53. [ Links ]
61. Yeh HI, Rothery S, Dupont E, Coppen SR, Severs NJ. Individual gap junction plaques contain multiple connexins in arterial endothelium. Circ Res 1998;83:1248-63. [ Links ]
62. Kwak BR, Mulhaupt F, Veillard N, Gros DB, Mach F. Altered pattern of vascular connexin expression in atherosclerotic plaques. Arterioscler Thromb Vasc Biol 2002;22:225-30. [ Links ]
63. Polacek D, Lal R, Volin MV, Davies PF. Gap junctional communication between vascular cells. Induction of connexin43 messenger RNA in macrophage foam cells of atherosclerotic lesions. Am J Pathol 1993;142:593-606. [ Links ]
64. Hu J, Cotgreave IA. Differential regulation of gap junctions by proinflammatory mediators in vitro. J Clin Invest 1997;99:2312-6. [ Links ]
65. Boerma M, Forsberg L, Van Zeijl L, Morgenstern R, De Faire U, Lemne C, et al. A genetic polymorphism in connexin 37 as a prognostic marker for atherosclerotic plaque development. J Intern Med 1999;246:211-8. [ Links ]
66. Peters NS. New insights into myocardial arrhythmogenesis: distribution of gap-junctional coupling in normal, ischaemic and hypertrophied human hearts. Clin Sci (Lond) 1996;90:447-52. [ Links ]
67. Matsushita S, Kurihara H, Watanabe M, Okada T, Sakai T, Amano A. Alterations of Phosphorylation State of Connexin 43 during Hypoxia and Reoxygenation Are Associated with Cardiac Function. J Histochem Cytochem 2006;54:343-53. [ Links ]
68. Garcia-Dorado D, Rodriguez-Sinovas A, Ruiz-Meana M. Gap junction-mediated spread of cell injury and death during myocardial ischemia-reperfusion. Cardiovasc Res 2004;61:386-401. [ Links ]
69. Miura T, Ohnuma Y, Kuno A, Tanno M, Ichikawa Y, Nakamura Y, et al. Protective role of gap junctions in preconditioning against myocardial infarction. Am J Physiol Heart Circ Physiol 2004; 286:H214-21. [ Links ]
70. Teunissen BE, Jongsma HJ, Bierhuizen MF. Regulation of myocardial connexins during hypertrophic remodelling. Eur Heart J 2004;25:1979-89. [ Links ]
71. Ritter O, Neyses L. The molecular basis of myocardial hypertrophy and heart failure. Trends Mol Med 2003;9:313-21. [ Links ]
72. Kostin S, Dammer S, Hein S, Klovekorn WP, Bauer EP, Schaper J. Connexin 43 expression and distribution in compensated and decompensated cardiac hypertrophy in patients with aortic stenosis. Cardiovasc Res 2004;62:426-36. [ Links ]
73. Kostin S, Rieger M, Dammer S, Hein S, Richter M, Klovekorn WP, et al. Gap junction remodeling and altered connexin43 expression in the failing human heart. Mol Cell Biochem 2003; 242:135-44. [ Links ]
74. Dupont E, Matsushita T, Kaba RA, Vozzi C, Coppen SR, Khan N, et al. Altered connexin expression in human congestive heart failure. J Mol Cell Cardiol 2001;33:359-71. [ Links ]
75. Spragg DD, Akar FG, Helm RH, Tunin RS, Tomaselli GF, Kass DA. Abnormal conduction and repolarization in late-activated myocardium of dyssynchronously contracting hearts. Cardiovasc Res 2005;67:77-86. [ Links ]
76. Poelzing S, Rosenbaum DS. Altered connexin43 expression produces arrhythmia substrate in heart failure. Am J Physiol Heart Circ Physiol 2004;287:H1762-70. [ Links ]
77. Betsuyaku T, Nnebe NS, Sundset R, Patibandla S, Krueger CM, Yamada KA. Overexpression of cardiac connexin45 increases susceptibility to ventricular tachyarrhythmias in vivo. Am J Physiol Heart Circ Physiol 2006;290:H163-71. [ Links ]

