










Servicios Personalizados
Revista

Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkRevista argentina de cardiología
versión On-line ISSN 1850-3748
Rev. argent. cardiol. v.77 n.6 Ciudad Autónoma de Buenos Aires nov./dic. 2009
INVESTIGACIÓN BÁSICA
Diferenciación en cardiomiocitos a partir de células madre embrionarias humanas
María Élida Scassa1, Darío Fernández Espinosa1, María Questa1, Guillermo Videla Richardson1, Noelia Losino2, Carlos Luzzani2, Alejandra S. Guberman2, 3, Hugo O. Grancellimtsac, 4, Gustavo Sevlever1, Santiago G. Miriukamtsac, 1, 3, 4
MTSAC Miembro Titular de la Sociedad Argentina de Cardiología
1 Laboratorio de Biología del Desarrollo Celular, FLENI
2 Departamento de Química Biológica, Facultad de Ciencias Exactas y Naturales, Universidad de Buenos Aires
3 Consejo Nacional de Investigaciones Científicas y Técnicas (CONICET)
4 Departamento de Cardiología, FLENI
Recibido: 12/02/2009
Aceptado: 07/08/2009
Dirección para separatas: Dr. Santiago Miriuka Instituto FLENI Departamento de Cardiología Montañeses 2325 (C1428AQK) Ciudad Autónoma de Buenos Aires
RESUMEN
Introducción
Las células madre son motivo de intensa investigación debido a la posibilidad de su utilización en el tratamiento de numerosas enfermedades, en particular las cardiovasculares. La diferenciación de células madre embrionarias humanas en cardiomiocitos se ha realizado exitosamente in vitro. Se han establecido métodos de cultivo y diferenciación, señales involucradas en la cardiogénesis y los cardiomiocitos generados se han utilizado en modelos de regeneración miocárdica. Sin embargo, aún quedan muchos interrogantes que se están investigando activamente.
Objetivo
Desarrollar una metodología que permita el cultivo de células embrionarias y su diferenciación en cardiomiocitos.
Material y métodos
Se utilizaron cuatro líneas de células madre embrionarias humanas. Se cultivaron y diferenciaron a través de los métodos publicados previamente en la bibliografía. El estado indiferenciado y la diferenciación en cardiomiocitos se verificaron por medio de inmunomarcación fluorescente y RT-PCR.
Resultados
La metodología utilizada permitió cultivar las células y mantenerlas en estado indiferenciado. Aunque con eficacia dispar, se logró la diferenciación en cardiomiocitos de las cuatro líneas celulares utilizadas. La confirmación se realizó por medio de la expresión de factores de transcripción miocárdicos y proteínas estructurales cardíacas.
Conclusiones
El cultivo y la diferenciación de células madre embrionarias humanas fue posible en nuestro sistema. Estos resultados preliminares nos impulsan a continuar y a desarrollar nuestros métodos con células pluripotentes inducidas.
Palabras clave: Células madre embrionarias; Miogénesis; Cardiomiocito
Abreviaturas:
CME Células madre embrionarias
FERi Fibroblastos embrionarios de ratón inactivados
PNA Péptido natriurético auricular
RT-PCR Reacción en cadena de la polimerasa con transcripción inversa
SUMMARY
Differentiation of Human Embryonic Stem Cells into Cardiomyocytes
Background
The role of stem cells in the treatment of several conditions, especially heart diseases, is under permanent investigation. Human embryonic stem cells have been successfully differentiated in vitro into cardiomyocytes. Methods of cell culture and cardiomyocyte differentiation are well established; signals regulating cardiogenesis have been identified and the cardiomyocytes generated have been used in models of myocardial regeneration. However, several questions still remain and are currently under active investigation.
Objective
To develop a culture system that is suitable for the induction of embryonic stem cells to cardiomyocyte differentiation.
Material and Methods
Four human embryonic stem cell lines were used. The cells were cultured and differentiation was induced using methods previously described. The presence of cells in an undifferentiated state and cardiomyocyte differentiation was detected by immunohistochemical studies (fluorescent staining) and RT-PCR.
Results
The methodology used allowed stem cells growth in the culture, and maintained them in an undifferentiated state. Cardiomyocyte differentiation was achieved in the four cell lines used, yet with uneven efficacy. This was confirmed by the expression of myocardial transcription factors and heart structural proteins.
Conclusions
Our system allowed human embryonic stem cell growth and differentiation in the culture. These preliminary results encourage us to continue developing our methods with induced pluripotent stem cells.
Key words: Embryonic Stem Cells; Myogenesis; Cardiomyocyte
INTRODUCCIÓN
Las células madre son motivo de activa investigación debido a la posibilidad de su utilización en el tratamiento de numerosas enfermedades, en particular las cardiovasculares. La definición actual de célula madre se basa en la presencia de tres características: en primer lugar, su capacidad de perpetuarse y renovar su linaje. Segundo, su capacidad de diferenciación en uno o varios tipos celulares. Por último, la capacidad de regularse y así limitar su potencial de propagación y crecimiento. En mayor o menor grado, estas tres características están presentes en las poblaciones celulares que conocemos como células madre. Sin embargo, estas células comprenden un grupo heterogéneo. Existen diferencias pronunciadas entre distintas células madre, que incluyen su capacidad de diferenciación, proliferación, localización y ciclo celular. La división más general establece las células madre adultas y las células madre embrionarias (CME). Entre las primeras se encuentran las células madre hematopoyéticas y mesenquimáticas (presentes mayormente en la médula ósea), las células progenitoras endoteliales y las células madre presentes en los diferentes tejidos especializados, como las cardíacas. Todas las células madre adultas presentan un gran potencial de diferenciación en linajes adultos, aunque su espectro de diferenciación es relativamente limitado (multipotencialidad). Así, las células madre hematopoyéticas presentan la capacidad de diferenciarse en las diferentes líneas sanguíneas, pero su potencial para diferenciarse en células no hematopoyéticas se ha discutido ampliamente (transdiferenciación) y, en caso de que exista, parece ser un fenómeno raro y de dudosa relevancia biológica y clínica.
Las células madre embrionarias presentan un potencial de diferenciación mucho mayor (pluripotencialidad). Estas células son capaces de diferenciarse en todas las formas adultas. Además, las CME presentan una capacidad proliferativa prácticamente ilimitada y senescencia nula o mínima. Estas características se deben a su origen, ya que se obtienen de la masa celular interna del blastocisto. Las primeras células embrionarias fueron derivadas de un blastocisto de ratón en 1981 por Evans y colaboradores. (1) La originalidad del trabajo se basó en establecer las condiciones de cultivo necesarias para mantener las células embrionarias en un estado indiferenciado por tiempo indefinido. Debe aclararse que in vivo las CME mantienen sus propiedades por períodos muy breves durante la formación embrionaria. Posteriormente, se diferencian para dar origen a distintos tipos celulares. Las CME humanas no fueron derivadas sino hasta el año 1998 por Thompson y colaboradores.(2) Nuevamente, su generación fue un hito y, una vez más, las condiciones de cultivo fueron complicadas y difíciles. Estos trabajos mostraban que una característica de estas células era la capacidad de diferenciarse en todos los tejidos adultos y que, al ser inyectadas en ratones inmunosuprimidos, generaban teratomas. En el año 2001, el grupo de Lior Gepstein, en Israel, estableció las bases para diferenciar las CME en cardiomiocitos. (3) Posteriormente han aparecido numerosos trabajos que ampliaron los conocimientos acerca de los mecanismos involucrados en el proceso de diferenciación miocárdica. Se han establecido métodos de cultivo y diferenciación, señales involucradas en la cardiogénesis y los cardiomiocitos generados se han utilizado en modelos de regeneración miocárdica. (4, 5) Sin embargo, aún quedan muchos interrogantes que se están investigando activamente.
El objetivo de nuestro trabajo fue el de reproducir las condiciones para mantener en estado indiferenciado las células embrionarias humanas y, posteriormente, diferenciarlas en cardiomiocitos en nuestro laboratorio. El cumplimiento de este objetivo tiene el propósito desarrollar un laboratorio con la capacidad para realizar estudios básicos y preclínicos en el área de las células pluripotentes.
MATERIAL Y MÉTODOS
Células y método de cultivo
Todas las células embrionarias humanas utilizadas se obtuvieron de los laboratorios que las derivaron. En los experimentos realizados se utilizaron las líneas celulares originadas en los Estados Unidos H9, H5, H16 y una línea originada en Suecia (S351). Las CME fueron cocultivadas con fibroblastos embrionarios de ratón inactivados (FERi) por irradiación en medio knockout-DMEM (Invitrogen) suplementado con 10% de suero sustituto (knock-out serum replacement, KSR, Invitrogen), bFGF 4 ng/ml (Calbiochem), penicilina 100 U/ml, estreptomicina 100 mg/ml, L-glutamina 2 mM, aminoácidos no esenciales 100 mM y b-mercaptoetanol 55 mM. Alternativamente, las células se cultivaron en placas cubiertas con Matrigel![]() (BD Bioscience) diluido 1:40 en PBS y en medio condicionado con FERi. Los FER se obtuvieron en nuestro laboratorio luego de ser aislados y expandidos a partir de embriones de ratones CF-1 en el día 13,5 de desarrollo.
(BD Bioscience) diluido 1:40 en PBS y en medio condicionado con FERi. Los FER se obtuvieron en nuestro laboratorio luego de ser aislados y expandidos a partir de embriones de ratones CF-1 en el día 13,5 de desarrollo.
Diferenciación en cardiomiocitos: las células embrionarias se cultivaron por los métodos previamente descriptos. Las colonias de CEH se incubaron con colagenasa (0,5 mg/ ml) (Sigma) y dispasa (0,25 mg/ml) (Sigma) por aproximadamente 1-2 minutos y luego se disgregaron mecánicamente con un cell-scraper (raspador de células). Luego, las colonias se suspendieron en placas no adherentes y en medio DMEM suplementado con suero fetal bovino 20% (Invitrogen), penicilina 100 U/ml, estreptomicina 100 mg/ml, Lglutamina 2 mM y aminoácidos no esenciales100 mM. Este método lleva a la formación de conglomerados celulares llamados cuerpos embrioides. Luego de 7 días de crecimiento en suspensión, estos cuerpos se traspasaron a placas adherentes previamente cubiertas con gelatina bovina 0,1% (Sigma). El cultivo se continuó en las mismas placas y el medio se cambió cada 2 o 3 días.
Inmunomarcación
Para la inmunomarcación, las células se fijaron con paraformaldehído al 4% en PBS; tras tres lavados con 0,1% de seroalbúmina bovina (BSA) en PBS, las células se trataron con solución de permeabilización y bloqueo (0,1% BSA, 0,1% Triton X-100 y 10% suero normal de cabra en PBS) por 45 minutos. Posteriormente, las células se incubaron con los respectivos anticuerpos primarios diluidos en solución de bloqueo (0,1% BSA y 10% de suero normal de cabra en PBS) por 2 horas a temperatura ambiente. Finalmente, luego de tres lavados con 0,1% BSA en PBS, se agregaron los correspondientes anticuerpos secundarios. La tinción nuclear se realizó con el colorante fluorescente 4,6-diamidino-2-fenilindol (DAPI) (5 mg/ml). Las imágenes se tomaron con un microscopio Nikon Eclipse TE2000-S.
Se utilizaron los siguientes anticuerpos primarios: monoclonal de ratón anti-SSEA4 (clon 813-70), monoclonal de ratón anti-Tra-1-81, monoclonal de ratón anti-Tra-1-60, monoclonal de ratón anti-OCT3/4 (clon C-10), policlonal de conejo anti-NANOG (clone H-155), policlonal de conejo anti- ANP (clon FL-153), policlonal de conejo anti-Nkx2.5 (clon H-114), monoclonal de ratón anti-GATA-4 (clon G-4), policlonal de conejo anti-GATA-6 (clon H-92), monoclonal de ratón anti-troponina T cardíaca (clon CT3), monoclonal de ratón anti-SCN5A (clon H-170), policlonal de cabra anti- Islet-1 (clon K-20). Todos los anticuerpos son de Santa Cruz Biotechnology, USA. Se emplearon los siguientes anticuerpos secundarios conjugados con fluoróforos: anti-IgG-FITC de ratón, anti-IgG-Texas Red-X de conejo, y anti-IgG-Alexa594 de cabra.
RT-PCR
El cDNA se sintetizó a partir de 0,5 mg de RNA total obtenido con el kit de aislamiento y purificación de RNA total "PureLink![]() Micro-to-Midi" de Invitrogen por retrotranscripción con la enzima SuperScript
Micro-to-Midi" de Invitrogen por retrotranscripción con la enzima SuperScript![]() III y hexámeros de secuencia al azar, de acuerdo con las instrucciones del proveedor (Invitrogen). La amplificación de los cDNA se realizó por PCR con empleo de la enzima Platinum
III y hexámeros de secuencia al azar, de acuerdo con las instrucciones del proveedor (Invitrogen). La amplificación de los cDNA se realizó por PCR con empleo de la enzima Platinum![]() Taq DNA polimerasa (Invitrogen) y oligonucleótidos específicos cuyas secuencias se detallan en Tabla 1.
Taq DNA polimerasa (Invitrogen) y oligonucleótidos específicos cuyas secuencias se detallan en Tabla 1.
Tabla 1. Primers (cebadores) utilizados en RT-PCR
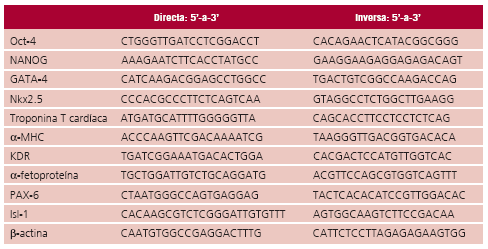
Los productos de amplificación se sometieron a electroforesis en gel de agarosa al 1,5% en buffer TAE 1x (Tris 40 mM Tris, ácido acético glacial 20 mM y EDTA 1 mM). Los fragmentos se visualizaron por tinción con bromuro de etidio (0,5 mg/ml) en un analizador de imágenes.
RESULTADOS
Las cuatro líneas celulares se cultivaron como se ha publicado por un largo número de pasajes sin que presentaran modificaciones en su morfología o aspecto. A través de los sucesivos pasajes se observan colonias en cocultivo con FERi. Estas colonias adoptan la típica forma descripta, con bordes netos y compactos. Las células embrionarias presentan una relación núcleo/ citoplasma alta, con nucléolos prominentes. La inmunomarcación mostró que estas colonias expresan marcadores de indiferenciación, incluidos OCT-4, SSEA- 4, Tra-1-60, Tra-1-81 y NANOG. La expresión de estos marcadores es homogénea. Las colonias fueron negativas para otros marcadores de indiferenciación, como Nkx2.5 y Pax-6 (dato no mostrado). Posteriormente, las CME se cultivaron por varios pasajes sobre Matrigel y con medio condicionado de FERi y se observó que las colonias indiferenciadas se mantenían como tales, expresando en la inmunomarcación los marcadores de indiferenciación, sin presentar signos morfológicos de diferenciación.
Por último, la expresión de marcadores de indiferenciación se confirmó por medio de RT-PCR para OCT-4 y NANOG (datos no mostrados).
Las células se diferenciaron por medio de la formación de cuerpos embrioides. Estos cuerpos en suspensión adoptan una forma esférica con bordes regulares y definidos. Luego de ser pasados a placas adherentes, los cuerpos embrioides se adhieren y sus células se expanden y adoptan diferentes morfologías pertenecientes a diferentes linajes celulares. A partir del día 8 a 9 desde el inicio de la diferenciación se observaron zonas con actividad contráctil espontánea en un porcentaje menor de los cuerpos adheridos (véase video en material suplementario on line). Estas zonas presentan un ritmo y una contractilidad similares a los de las células cardíacas. Las zonas corresponden a una porción menor del total de las células y publicaciones previas establecen que el porcentaje de las células con actividad contráctil es menor del 10%. (3) Las diferentes zonas observadas difieren en la frecuencia de contracción, que varía en un rango de aproximadamente 5 a 10 veces por minuto hasta 60- 80 veces por minuto. Esta diferencia fue estudiada previamente y se corresponde con la diferenciación de cardiomiocitos con fenotipos diferentes y correspondientes a tejido de conducción, auricular o ventricular. (6) Las zonas contráctiles mantienen su actividad por largos períodos y algunas zonas se mantuvieron por 60 días sin que se observaran cambios. Además, la frecuencia de contracción se mantiene por días sucesivos sin alteraciones. Por último, se encontraron diferencias en la propensión a formar zonas contráctiles entre las distintas líneas celulares. Así, los cuerpos embrioides de las células H5 presentaron zonas contráctiles en el 10% de los casos, las H9 en el 5% y las H16 y las S351 en el 0,5%. Estas diferencias se informaron recientemente y podrían deberse a diferencias epigenéticas entre las líneas utilizadas. (7)
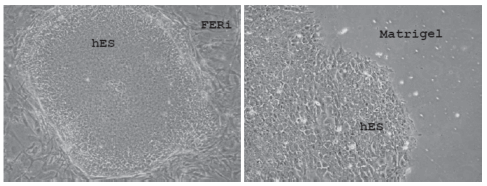
Fig. 1. Colonias de células embrionarias humanas en estado indiferenciado cultivadas tanto sobre FERi como sobre Matrigel. Se observa una relación núcleo/citoplasma alta, agrupadas densamente y formando una colonia de bordes netos.
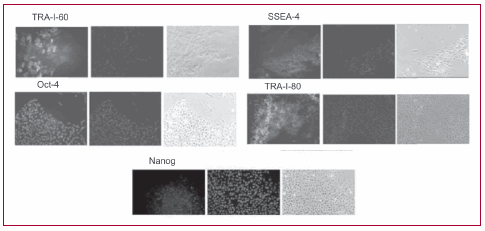
Fig. 2. Inmunomarcación fluorescente para indiferenciación. Se detecta una marcación homogénea de las colonias para OCT-4, SSEA-4, Tra-1-60, Tra-1- 81 y NANOG, todos ellos marcadores de pluripotencialidad.
Se utilizó inmunofluorescencia para distintos marcadores de diferenciación miocárdica a fin de determinar el fenotipo de las zonas contráctiles. Se observó que estas zonas eran positivas para los factores de transcripción cardíaco-específicos Nkx2.5 y GATA-4. Además, las zonas fueron positivas para marcadores estructurales cardíacos como troponina C cardíaca, péptido natriurético auricular (PNA) y el canal del sodio SCN5A. La inmunomarcación fue positiva en las cuatro líneas celulares estudiadas (Figura 3). Estas mismas zonas no presentaron marcación para OCT- 4, lo que evidencia desaparición de marcadores de indiferenciación (dato no mostrado). La ampliación de algunas células contráctiles demuestra la arquitectura de las miofibrillas cuando son teñidas para troponina C cardíaca (Figura 4).

Fig. 3. Inmunomarcación fluorescente de zonas contráctiles. Las cuatro líneas celulares embrionarias fueron diferenciadas y se observaron zonas con actividad contráctil espontánea, las cuales fueron positivas para marcadores cardíacos específicos.
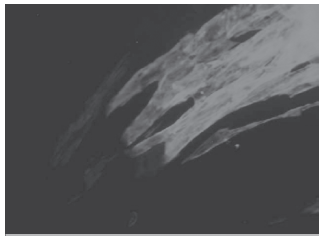
Fig. 4. Ampliación de una zona contráctil con marcación para troponina T cardíaca. Se observa la distribución fibrilar.
Posteriormente, por medio de RT-PCR se analizó la expresión de marcadores de diferenciación miocárdica. Se extrajo RNA de muestras secuenciales tomadas a lo largo de los primeros días de diferenciación y se observó que la expresión de GATA-4 y troponina C cardíaca se incrementaba (Figura 5 A). Posteriormente se realizó una escisión mecánica de las zonas contráctiles y se efectuó RT-PCR sobre el RNA total obtenido de esas secciones. Se halló que estas zonas eran positivas para troponina C cardíaca y negativas para el marcador de ectodermo Pax-6 (Figura 5 B). La expresión del marcador endodérmico fue positiva en ambas muestras, probablemente por contaminación con células pertenecientes a zonas no contráctiles durante la disección.

Fig. 5. A. Evaluación por RT-PCR de la aparición de marcadores de diferenciación cardíaca durante la diferenciación. B. Evaluación por RT-PCR de la especificidad de los marcadores cardíacos en las zonas disecadas con actividad contráctil.
A continuación, las zonas contráctiles se incubaron con dosis crecientes de epinefrina. Se observó que la activación adrenérgica generó un incremento de la frecuencia de contracción de hasta el 67% respecto al valor basal (Figura 6). Esta respuesta demuestra un desarrollo funcional del sistema adrenérgico en las células diferenciadas. Por último, seis zonas con actividad contráctil se sometieron a un estímulo de hipoxia gaseosa durante 18 horas (concentración de oxígeno 1%). Luego de 15 minutos de reoxigenación en condiciones atmosféricas, se observó ausencia de actividad contráctil en dos de las zonas y, en una de ellas, con clara actividad desorganizada del tipo fibrilatorio.
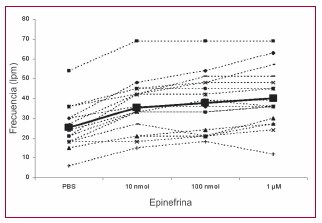
Fig. 6. Respuesta adrenérgica positiva de las zonas contráctiles. Al agregar concentraciones crecientes de epinefrina se observa una respuesta cronotrópica positiva.
DISCUSIÓN
La diferenciación en cardiomiocitos a partir de células embrionarias indiferenciadas es una técnica en investigación con el fin de aplicarla en medicina regenerativa cardiovascular. Este modelo permite además investigar en células humanas las distintas señales involucradas en la cardiogénesis. Por último, se postula que los cardiomiocitos así obtenidos se podrían utilizar en experimentos de toxicidad farmacológica. El presente trabajo demuestra la factibilidad del desarrollo de cardiomiocitos a partir de CME humanas en un laboratorio local. Las células derivadas presentan actividad contráctil espontánea, inmunomarcación positiva para proteínas cardíacas, expresión génica específicamente cardíaca y respuesta adrenérgica positiva. En nuestro conocimiento, ésta es la primera comunicación sobre derivación de cardiomiocitos a partir de células madre embrionarias humanas desarrolladas en nuestro país. La ausencia de trabajos en esta materia en el ámbito local posiblemente se deba a la complejidad de la metodología y sus altos costos. Sin embargo, en la actualidad, las células pluripotenciales son herramientas poderosas para la investigación básica y aplicada. Particularmente, los recientes avances en reprogramación celular generan enormes expectativas acerca del futuro cercano de estas líneas experimentales.
Los avances en reprogramación celular son un hecho que merece ser destacado. La reprogramación celular consiste en inducir la desdiferenciación de fibroblastos u otros tipos celulares adultos hacia células similares a las embrionarias, tanto con las mismas particularidades morfológicas y funcionales como con la misma capacidad proliferativa y de diferenciación que las células embrionarias. Es decir, el método permite conjugar una fuente individual ilimitada de células madre totipotenciales para ser diferenciadas en cualquier tipo de células adultas. Los trabajos de reprogramación celular representan un avance significativo para la medicina regenerativa. En el año 2006, el grupo de Shinya Yamanaka, en Japón, comunicó la reprogramación de fibroblastos de ratón por medio de la introducción de factores de transcripción específicos que regulan el estado indiferenciado de las células embrionarias. (8) En su trabajo demuestran la diferenciación de las células obtenidas, llamadas iPS, por induced-pluripotent stem cells, en distintos tejidos adultos como neuronal y miocárdico, entre otros. Un año más tarde, el mismo grupo y el grupo de Thomson, en Chicago, presentaron simultáneamente la misma metodología aplicada en fibroblastos humanos. (9, 10) Este hallazgo ya vislumbra nuevas oportunidades terapéuticas. Por ejemplo, Hanna y colaboradores crearon iPS a partir de fibroblastos de ratón portadores de un tipo de anemia de células falciformes. En este trabajo, introdujeron modificaciones genéticas ex vivo con el propósito de revertir la anomalía genética. Posteriormente desarrollaron células madre hematopoyéticas, las cuales fueron trasplantadas en la médula ósea de los ratones enfermos. El resultado fue una notable mejoría en la enfermedad de los ratones. (11) Más recientemente, se han logrado iPS a partir de pacientes con diferentes patologías, como diabetes, enfermedad de Alzheimer o distrofias musculares. (12) Potencialmente, estas células iPS generadas a partir de los mismos pacientes se podrán modificar y utilizar para el tratamiento de su propia enfermedad. Además, se podrían emplear en modelos celulares de las distintas enfermedades, por lo que podrían surgir nuevos conocimientos a partir de ellas. En nuestro laboratorio hemos comenzado a trabajar en la generación de iPS. Actualmente, nuestro grupo se encuentra desarrollando células pluripotenciales a partir de fibroblastos humanos. Esperamos poder aplicar en los próximos meses las técnicas de diferenciación miocárdica en iPS desarrolladas localmente.
Las células madre embrionarias o sus derivadas presentan el riesgo de formar teratomas una vez implantadas. Este riesgo se ha observado en modelos en animales luego de la implantación de células indiferenciadas. El implante de células diferenciadas también conlleva el riesgo implícito del desarrollo de teratomas. Sin embargo, no se ha aclarado debidamente cuál es la capacidad de formación de teratomas una vez que las células pluripotenciales son diferenciadas terminalmente. Específicamente en el tejido cardíaco, en las comunicaciones realizadas se afirma que las células diferenciadas in vitro, seleccionadas por su fenotipo cardíaco e implantadas en el miocardio, no forman teratomas. Estos modelos en animales, sin embargo, deberán estudiarse extensamente y validarse antes de avanzar con estudios clínicos.
En conclusión, en nuestro país es posible desarrollar la diferenciación en cardiomiocitos a partir de células madre. Lejos de representar un avance para su aplicación clínica inmediata, estos hallazgos deben fomentar la investigación básica y aplicada para posibles terapias celulares en el futuro.
1. Evans MJ, Kaufman MH. Establishment in culture of pluripotential cells from mouse embryos. Nature 1981;292:154-6. [ Links ]
2. Thomson JA, Itskovitz-Eldor J, Shapiro SS,Waknitz MA, Swiergiel JJ, Marshall VS, et al. Embryonic stem cell lines derived from human blastocysts. Science 1998;282:1145-7. [ Links ]
3. Kehat I, Kenyagin-Karsenti D, Snir M, Segev H, Amit M, Gepstein A, et al. Human embryonic stem cells can differentiate into myocytes with structural and functional properties of cardiomyocytes. J Clin Invest 2001;108:407-14. [ Links ]
4. Leor J, et al. Human embryonic stem cell transplantation to repair the infarcted myocardium. Heart 2007;93(10):1278-84. [ Links ]
5. Laflamme MA, Murry CE. Regenerating the heart. Nat Biotechnol 2005;23:845-56. [ Links ]
6. Kolossov E, Lu Z, Drobinskaya I, Gassanov N, Duan Y, Sauer H, et al. Identification and characterization of embryonic stem cell-derived pacemaker and atrial cardiomyocytes. FASEB J 2005;19:577-9. [ Links ]
7. International Stem Cell Initiative. Adewumi O, Aflatoonian B, Ahrlund-Richter L, Amit M, Andrews PW, Beighton, et al. Characterization of human embryonic stem cell lines by the International Stem Cell Initiative. Nat Biotechnol 2007;25:803-16. [ Links ]
8. Takahashi K, Yamanaka S. Induction of pluripotent stem cells from mouse embryonic and adult fibroblast cultures by defined factors. Cell 2006;126(4):663-76. [ Links ]
9. Takahashi K, Yamanaka S. Induction of pluripotent stem cells from mouse embryonic and adult human fibroblasts by defined factors. Cell 2006;131:663-76. [ Links ]
10. Yu J, Vodyanik MA, Smuga-Otto K, Antosiewicz-Bourget J, Frane JL, Tian S, et al. Induced pluripotent stem cell lines derived from human somatic cells. Science 2007;318:1917-20. [ Links ]
11. Hanna J,Wernig M, Markoulaki S, Sun CW, Meissner A, Cassady JP, et al. Treatment of sickle cell anemia mouse model with iPS cells generated from autologous skin. Science 2007;318:1920-3. [ Links ]
12. Park IH, Arora N, Huo H, Maherali N, Ahfeldt T, Shimamura A, et al. Disease-specific induced pluripotent stem cells. Cell 2008; 134:877-86. [ Links ]

