










Services on Demand
Journal

Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO
Related links
-
 Similars in
SciELO
Similars in
SciELO
Share

 Permalink
PermalinkComechingonia
On-line version ISSN 1851-0027
Comechingonia vol.20 no.1 Córdoba June 2016
ARTÍCULOS ORIGINALES
El registro faunístico del sitio Negro Muerto 3 (provincia de Río Negro, Argentina): tafonomía y patrones de explotación
The faunal record from Negro Muerto 3 site (Río Negro province, Argentina): taphonomy and exploitation patterns
Emiliano Mange1, Luciano Prates2, Lucio González Venanzi3 y Maitén Di Lorenzo4
1 INCUAPA-CONICET. Av. Del Valle 5737, (7400) Olavarría, Argentina, emilianomange@gmail.com;
2 CONICET. Facultad de Ciencias Naturales y Museo, Universidad Nacional de La Plata. Paseo del Bosque s/n, (1900) La Plata, Argentina, lprates@fcnym.unlp.edu.ar;
3 Facultad de Ciencias Naturales y Museo, Universidad Nacional de La Plata. Paseo del Bosque s/n, (1900) La Plata, Argentina, luciogonzalezvenanzi@gmail.com;
4 Facultad de Ciencias Naturales y Museo, Universidad Nacional de La Plata. Paseo del Bosque s/n, (1900) La Plata, Argentina, maiten.dilorenzo@gmail.com
Presentado: 06/03/2016
Aceptado: 10/06/2016
Resumen
En el sitio arqueológico Negro Muerto 3, ubicado en el valle medio del río Negro, se recuperaron entierros humanos y diversos materiales (artefactos líticos, fragmentos de cerámica y restos faunísticos). El sitio fue interpretado como un área de uso residencial y mortuorio, ocupada durante el Holoceno tardío. En este trabajo se presentan los resultados del análisis de los materiales faunísticos, que incluyen valvas de moluscos, cáscaras de huevos y elementos óseos (de peces, reptiles, aves y mamíferos). El análisis muestra que el conjunto es de origen principalmente antrópico y que una parte importante de los restos fue ingresada para su consumo. Las evidencias de dinámica postdepositacional muestran escasa exposición superficial de los materiales, y una incidencia importante de precipitación de carbonato y raíces. Si bien las características generales del sitio sugieren un patrón de explotación de la fauna similar al de otros sitios de la región con cronologías cercanas (alta diversidad de especies de pequeño porte, propias del ambiente de valle), se señalan algunas diferencias. La nueva información generada en el trabajo es integrada a los modelos desarrollados previamente y, a partir de esto, se plantean nuevos interrogantes e hipótesis sobre los patrones de explotación de los recursos faunísticos.
Palabras clave: Restos faunísticos; Cazadores-recolectores; Valle del río Negro; Holoceno tardío final.
Abstract
Negro Muerto 3 archaeological site is located in the middle Negro river valley and consists of human burials and various archaeological cultural evidence (such as lithic, pottery sherds, and faunal remains). The site was interpreted as an area used both as residential base camp and as mortuary area, and was dated to the late Holocene. In this paper the results of the analysis of fauna (fresh water shells, eggshells and vertebrate bones) are presented. The vertebrate bone assemblage includes specimens of fishes, reptiles, birds and small, medium and large sized mammals. Several lines of evidence (presence of burning, cut marks and crushed bones) have been identified. The analysis of weathering indicates a short-term superficial exposure of the ensemble, and a high incidence of carbonate precipitation and root marks. Though main results of faunal analysis of NM3 show a similar pattern than emerged from other sites of the same region (e.g. high diversity of low-sized prey, and predominance of riverine species), large-sized mammals seems to have been relatively more significant. The new information generated in this paper is integrated with previous archaeological models and, on this basis, new questions and hypotheses regarding to the human exploitation of fauna are raised.
Keywords: Faunal remains; Hunter-gatherers; Negro river valley; Final late Holocene.
Introducción
En los sectores medio e inferior del valle del río Negro (Pcia. de Río Negro) se han hallado numerosos sitios arqueológicos desde mediados del siglo XIX, muchos de ellos con presencia de restos faunísticos (Fernández et al. 2011; Mange et al. 2014; Prates 2008; Prates y Acosta Hospitaleche 2010; Prates et al. 2010). Como tendencia general, estos sitios están localizados en la planicie aluvial moderna del río, cerca de paleocauces del interior del valle y cronológicamente corresponden al Holoceno tardío. Algunos sectores, como en la ex-laguna del Juncal y el valle de Negro Muerto, presentan una densidad significativamente alta de evidencia arqueológica. Esta última es una microrregión ubicada en el sector medio y en la ribera norte del río Negro, uno de los sectores de valle más amplios de esta cuenca, en la cual se han registrado numerosos sitios arqueológicos, la mayoría superficiales y algunos de ellos en estratigrafía (Luchsinger 2006; Prates 2008). En todos estos sitios se hallaron materiales faunísticos y a partir de ellos, de los materiales asociados, y de la distribución de los sitios, se propuso un modelo de aprovechamiento de los recursos animales de distintos nichos o sectores del área: de los ambientes fluviales del valle (río, brazos y lagunas), de sectores del interior del valle no conectados con ambientes fluviales (pastizales y montes), y de las mesetas adyacentes ubicadas fuera del valle (Prates 2008). Los dos primeros se habrían explotado dentro del área diaria de captación de recursos alrededor de las bases residenciales (daily foraging trips) establecidas en la planicie aluvial, mientras que para el tercero se habrían realizado ocasionalmente partidas logísticas de más de un día de duración (Prates 2008).
El sitio Negro Muerto 3 (en adelante NM3) se ubica en el extremo oeste del valle de Negro Muerto (39° 48.8' S. y 65° 16.8' W.), a unos 70 km de la ciudad de Choele Choel y en cercanías de los sitios Negro Muerto (NM) y Negro Muerto 2 (NM2) (Figura 1). En el lugar se recuperaron entierros humanos y diversos restos asociados a una ocupación humana, como artefactos líticos, sedimentos termoalterados, fragmentos de cerámica y restos faunísticos. El sitio se encontraba sobre la planicie aluvial del río, en una pequeña elevación de arena fina (textura areno-fangosa sensu Folk 1951) que apoya sobre el manto de loess que recubre la mayor parte del valle (Luchsinger 2006: Figura 31) y que fue removida por maquinaria agrícola cuando se realizaban tareas de desmonte y nivelación. Esta elevación se encontraba al oeste y adyacente a una pequeña laguna formada en el interior de un canal de inundación del río. El área de exposición de los materiales faunísticos en superficie era de unos 3250 m2 (65 x 50 m), aunque la mayor concentración se encontraba en el sector central de unos 400 m2 (20 x 20 m). Se obtuvieron fechados radiocarbónicos sobre restos de Lama guanicoe (1430 ± 70 14C años AP; LP3219, tibia) y de Ozotoceros bezoarticus (1110 ± 33 14C años AP, AA106437, falange). Esta cronología (Holoceno tardío final) es similar a la obtenida para otras ocupaciones residenciales del mismo valle (Prates 2008; Prates et al. 2010). En este trabajo se presentan los primeros resultados del análisis de los materiales faunísticos del sitio NM3 y, sobre esta base, se discute el modelo general planteado con anterioridad para interpretar el registro faunístico del área. Para esto se definen las características de la muestra desde el punto de vista taxonómico y anatómico, se identifican los agentes y acciones que intervinieron en la formación del depósito, y se evalúa la importancia relativa y el área de captación de los distintos taxones.

Figura 1. Ubicación del sitio NM3 y sitios cercanos mencionados en el trabajo. Referencias: PO: Pomona, NM: Negro Muerto, NM2: Negro Muerto 2; LM: Loma de los Muertos, A1: Angostura 1.
Materiales y métodos
La muestra analizada en este trabajo (n=529) fue recuperada en el año 2013, luego de que el médano que contenía al sitio fuera nivelado. Se realizó una recolección superficial exhaustiva de toda la fauna visible en superficie. Para el análisis se utilizó una lupa binocular, y la metodología empleada fue distinta en cada tipo de material (elementos óseos, valvas y cáscaras de huevo). En los restos óseos se realizó la identificación anatómica, taxonómica y de lateralidad por medio de atlas osteológicos (Barone 1990; Giardina 2010; Pacheco et al. 1979, entre otros) y colecciones de referencia propias. Los restos de mamíferos sin determinación taxonómica más precisa fueron separados, cuando fue posible, en categorías según el peso corporal (Redford y Eisenberg 1992): micromamíferos (<1 kg), mamíferos medianos (1-20 kg) y mamíferos grandes (mayores a 20 kg). Para la cuantificación de los materiales se tuvieron en cuenta medidas de abundancia taxonómica y anatómica (NISP, NMI, MNE, MAU y MAU %) (Binford 1981; Grayson 1991; Mengoni Goñalons 1999).
Para el estudio tafonómico del conjunto óseo se consideraron las siguientes variables: meteorización, tipo de fractura, marcas de raíces, actividad de carnívoros y roedores, depositaciones químicas y marcas de procesamiento (termoalteraciones, huellas de corte, marcas de impacto), siguiendo las propuestas de varios autores (Behrensmeyer 1978; Gutiérrez 2004; Lyman 1994; Mengoni Goñalons 1999, entre otros). Únicamente la alteración térmica y las huellas antrópicas (de corte u otras) fueron evaluadas en toda la muestra. Las otras variables fueron relevadas en el conjunto de especímenes óseos de vertebrados medianos y grandes, excluyendo placas óseas de dasipódidos. Debido a su pequeño tamaño, no se consideró a estas últimas en los cálculos de incidencia de agentes tafonómicos como raíces, carnívoros, roedores y depositaciones químicas. La meteorización (siguiendo a Behrensmeyer 1978), se consideró únicamente en especímenes óseos no termoalterados de más de 1 cm de largo de mamíferos medianos y grandes, excluyendo de nuevo las placas dérmicas de armadillos. En el caso de los mamíferos chicos se registraron, además de termoalteraciones y huellas de corte, marcas de corrosión en las epífisis de fémures y húmeros (Andrews 1990; Fernández-Jalvo y Andrews 1992; Medina et al. 2012). El cálculo de NMI de este conjunto se realizó en base a las mandíbulas y cráneos, siguiendo los criterios de Pearson (1995) y Fernández et al. (2011).
En las cáscaras de huevo se registraron termoalteraciones y modificaciones antrópicas, y se calculó la densidad de poros a fin de determinar la presencia de distintas especies del género Rhea (Apolinaire y Turnes 2010). Por último, en el caso de los restos de moluscos de río, se determinó la lateralidad de las valvas de almejas, se midieron los tamaños de las enteras, se estimaron los de las fracturadas, y se cuantificaron los individuos ingresados (NMI) considerando el número máximo de valvas de la misma lateralidad que poseen charnela (Prates y Marsans 2007). Se registraron modificaciones antrópicas como termoalteración y formatización en la totalidad de la muestra de valvas de moluscos.
Resultados
1. Aspectos cuantitativos
Una parte importante del conjunto faunístico de NM3 se determinó hasta algún nivel taxonómico (NISP=512, NMI=118). La muestra incluye restos óseos y dentales (68.3%), valvas de moluscos (28.6%) y cáscaras de huevos (3.1%) (Tabla 1). Del total de individuos ingresados (NMI), más de la mitad (62%) corresponde a Diplodon chilensis, una almeja típica del río Negro con aparición recurrente en sitios arqueológicos del área y de la cuenca del río Limay (Caviglia y Borrero 1981; Crivelli Montero et al. 1996; Mange et al. 2013; Prates y Marsans 2007; Prates et al. 2011). Entre los materiales óseos y dentales se registraron principalmente elementos de mamíferos (NISP=319, NMI=26), y en menor medida de aves (NISP=28, NMI=4), reptiles (NISP=1, NMI=1) y peces (NISP=2, NMI=1).
Tabla 1. Representación taxonómica de los restos faunísticos de NM3.

Entre los restos de mamíferos predominan los de especies grandes (58%), seguidos por los micromamíferos (27%) y mamíferos medianos (15%). Los restos de micromamíferos pertenecen al orden Rodentia, y en conjunto incluyen tanto restos postcraneales (n=44) como cráneo-mandibulares (n=42). Considerando el tamaño (Fernández y De Santis 2013; Pardiñas 1999), corresponden principalmente a la categoría micromamíferos grandes (más de 200 g); muy pocos restos postcraneales pertenecerían a micromamíferos pequeños o medianos. Se registraron principalmente especímenes de caviomorfos (Microcavia australis, Galea leucoblephara, Ctenomys sp.) y cricétidos (Holochilus brasiliensis). Entre los restos de mamíferos medianos predominan los dasipódidos, tanto peludo (Chaetophractus villosus) como piche (Zaedyus pichiy), principalmente placas dérmicas y escasos restos óseos de otras especies (Dolichotis patagonum -mara-, Myocastor coypus -coypo-, Lycalopex sp.). Los restos de mamíferos grandes pertenecen a Lama guanicoe (guanaco) y Ozotoceros bezoarticus (venado de las pampas), numerosos restos no determinados y un fragmento vertebral de Mylodontinae (Figura 4e). Este último espécimen, correspondiente a una especie de megafauna extinguida en el Holoceno temprano (Brandoni et al. 2010), posiblemente Mylodon sp. (E. Tonni com. pers. 2014), se diferencia marcadamente del resto del conjunto óseo por el estado avanzado de mineralización, el color, textura y peso.
Se determinó un número mínimo de tres individuos de guanaco en base a las escápulas y fémures izquierdos; dos de ellos habrían sido menores a 19 meses de edad de acuerdo al estadio de osificación de las escápulas (Kaufmann 2009). La representación anatómica (Figura 2) muestra que las partes más abundantes son las pelvis, escápula (100%) y diáfisis de fémur (75%). Luego la mandíbula, axis, cráneo, húmero distal, diáfisis de tibia, falanges primeras y segundas (50%) y vértebras lumbares (35%). Los especímenes de venado son bastante más escasos (NISP=9, NMI=1, adulto) e incluyen también elementos del esqueleto axial (mandíbula, costillas) y del apendicular (falanges, metapodio, tibia, escápula). A partir de los restos de artiodáctilos (puntualmente de las escápulas izquierdas), que no pudieron determinarse a nivel específico (NISP=14), se agrega un individuo más, de guanaco pequeño o venado, llevando a cinco el total de artiodáctilos ingresados al sitio.
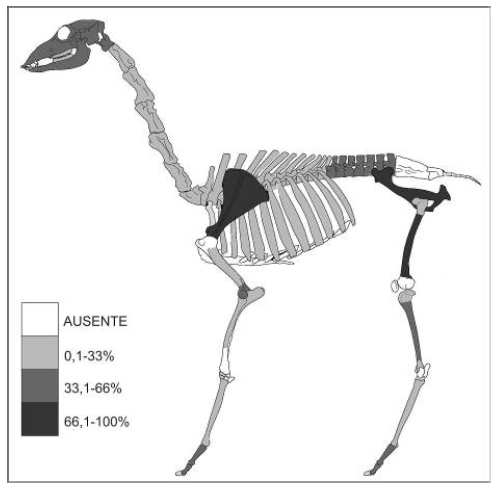
Figura 2. Representación esqueletaria (MAU%) de Lama guanicoe.
Con respecto a las aves, se identificaron restos óseos y cáscaras de huevo de rheidosjunto con especímenes óseos de aves de porte pequeño-mediano. Se determinó la presencia de las dos especies de Rhea sp. (ñandú) a partir de unas pocas cáscaras de huevos recuperadas (n=16), de las cuales pudieron determinarse sólo cinco (dos de R. americana, dos de R. pennata y una con valores intermedios). En base a los fémures se determinó la presencia de al menos tres individuos de aves pequeñas, una de ellas de menor tamaño (posiblemente un Passeriforme). Los elementos óseos presentes son de los miembros posteriores, con excepción de dos sinsacros. Los restos óseos de Rhea sp., aunque escasos (NISP=7), corresponden al esqueleto apendicular posterior. El conjunto incluye tres falanges, un fragmento de fémur, uno de tarsometatarso, uno de tibiatarso y un fragmento de hueso largo indeterminado.
Por último, también se registraron una escápula de tortuga terrestre (Chelonoidis chilensis) y dos vértebras de peces (Osteichthyes indet.). Entre los restos malacológicos predominan las valvas del bivalvo Diplodon chilensis (almeja común de río) y en menor cantidad del gasterópodo Chilina sp., ambas con hábitat en ambientes ribereños y lagunares, y también fragmentos de valvas de mitílidos (Aulacomya atra, Figura 4C), y volútidos de origen marino. En base a los especímenes izquierdos se calculó un número mínimo de 73 individuos de D. chilensis, con un promedio estimado de valva completa de 6.1 cm.
2. Aspectos tafonómicos
El análisis mostró que los agentes tafonómicos naturales que afectaron en mayor medida a los restos arqueofaunísticos fueron la acción de raíces (57.9%) y la precipitación de carbonato de calcio (56%), en forma coincidente con lo observado en otros conjuntos faunísticos de la región (Mange et al. 2013; Prates 2008). Ambos procesos se evaluaron en una muestra de 241 especímenes. Se observó meteorización incipiente en pocos especímenes (19 de una muestra de 128 especímenes evaluados), la mayoría de ellos (90%) en estadio 1 y unos pocos en estadio 2. Sólo dos especímenes tienen marcas de roedores; las marcas por actividad de carnívoros y pisoteo están ausentes, y se observaron restos de periostraco en algunas valvas de D. chilensis (12%). El registro de 14 fragmentos óseos con blanqueamiento (en el sentido de Mengoni Goñalons 1999) indica cierto tiempo de exposición de algunos elementos óseos. Se trata de un espécimen de C. chilensis (escápula) y de mamíferos grandes (algunos de guanaco: una vértebra y tres falanges y por otro lado, astillas y fragmentos de huesos largos). Por último se observa que el conjunto está altamente fragmentado: sólo el 11.2% de los especímenes óseos son elementos enteros. Entre los fracturados, el 52.4 % presenta fracturas con superficies y bordes rugosos, características de las ocurridas en estado seco (Gutiérrez 2004). Debido a que en la mayoría de estos especímenes las superficies de fractura no presentan indicadores (color y textura) de haber ocurrido recientemente, se asume que se produjeron con anterioridad a la acción mecánica durante el desmonte. El análisis de corrosión en restos de roedores pudo ser evaluado en escasos fémures y húmeros (n=12), que presentan hoyuelos de disolución en ocho elementos (cinco de ellos de micromamíferos grandes y tres de micromamíferos medianos/pequeños -i.e. sigmodontinos-).
Las evidencias directas de acción antrópica (huellas de corte, raspado y negativos de impacto) se registraron en 54 especímenes óseos (10.6%), de los cuales 27 pertenecen a L. guanicoe. De esta especie se hallaron varias huellas de corte en huesos largos, especialmente del miembro posterior (cuatro especímenes de fémur y tibia) y en menor medida del anterior (tres de húmeros y uno de radioulna), vértebras torácicas (dos) y en axis, vértebra lumbar, hemimandíbula, cráneo y hemipelvis. Son menos abundantes las evidencias de procesamiento en restos de otros vertebrados grandes, como mamíferos (seis fragmentos de diáfisis de huesos largos), venado (dos falanges, costilla, escápula y fragmento de metapodio) y Rhea sp. (primera falange). También se registraron en seis especímenes de roedores caviomorfos (huellas de corte en una mandíbula y tres tibias de G. leucoblephara -Figura 3b-, un fémur de sigmodontino que por su tamaño podría asignarse a Holochilus brasiliensis, y una hemipelvis indeterminada), cuatro huesos largos de aves medianas, dos placas de peludo, dos restos de mamíferos medianos y un calcáneo de mara (Figura 3c).

Figura 3. Evidencias de consumo de roedores en NM3: a) quemado en mandíbula de H. brasiliensis; b) huellas de corte en tibia de G. leucoblephara; c) huellas de corte en calcáneo de D. patagonum.
Se registraron numerosos restos quemados (29.4%) y fracturados en fresco (29%), que apoyarían también el aprovechamiento antrópico. La termoalteración afectó a restos óseos (30.7%), valvas (25.3%) y cáscaras de huevo (37.5%). Entre los primeros predominan los especímenes de artiodáctilos, principalmente guanaco (36.5%), dasipódidos (36,7%; entre los que se destacan las placas), y mamíferos grandes (23%). La evidencia de quemado es más escasa en restos de micromamíferos (20.7%), aunque se destaca el quemado en tres mandíbulas de tuco-tuco (Ctenomys sp.) y en una de Holochilus brasiliensis (Figura 3a). También se hallaron restos quemados de Mylodontinae, ñandú, mara y aves pequeñas. En el caso de las valvas de Diplodon chilensis la termoalteración se observa en el debilitamiento de las valvas, y en muy pocos ejemplares en cambios de coloración. Las fracturas en estado fresco se registraron principalmente en mamíferos grandes: guanaco (40.6%, la mayoría en el esqueleto apendicular), artiodáctilos (40%), indeterminados (32.8%) y venado (22.2%). Aunque escasas en mamíferos medianos, son importantes en aves pequeñas (35%) y rheidos (28.6%). Considerando los huesos largos de las extremidades, se observaron fracturas en guanaco (52%, predominan tibias, fémures y falanges) que podrían indicar el consumo de médula (Mengoni Goñalons 1999: 83), pero no en venado. Las fracturas en estado fresco en huesos de aves se encuentran todas en huesos largos. En Rhea americana (33.3%) se encuentran en el fémur y tarsometatarso.
Por último, se registraron dos instrumentos óseos sobre huesos largos de artiodáctilo(metapodio; Figura 4a) y de guanaco (radioulna; Figura 4b). Este último mide 13.6 cm y presenta huellas de corte laterales y aserrado distal. El primero mide 9.5 cm; ambos presentan pulido superficial y puntas. También se registró un pendiente realizado sobre valva de D. chilensis, con una perforación cónica realizada por rotación (Fig. 3c). Los taxa sin evidencia cierta de acción humana (huellas de corte, machacado, raspado o manufactura), o con evidencia ambigua (termoalteración o fracturas en fresco), son los de carnívoros, M. australis, Chelonoidis chilensis y moluscos marinos. En este último caso puede asignarse de todas maneras a un uso humano, considerando su traslado desde el litoral atlántico (ca. 150 km).

Figura 4. Materiales faunísticos de NM3 no asociados directamente al consumo de presas: a) retocador sobre metapodio de mamífero grande; b) instrumento sobre radioulna de Lama guanicoe; c) pendiente sobre valva de Diplodon chilensis; d) fragmento de valva de Aulacomya atra; e) fragmento de vértebra de Mylodontinae.
Discusión
El sitio NM3 se encuentra sobre un depósito eólico contiguo a una pequeña paleo-laguna, posiblemente activa durante su ocupación (discusión en Luchsinger 2006), en el que además de actividades de procesamiento y consumo de fauna se realizó el entierro de varios cuerpos humanos. Los dos fechados obtenidos sobre restos faunísticos (1430 y 1110 años AP) sugieren que corresponden a distintos eventos de ocupación del sitio, y debido a que no se dispone de fechas sobre los restos humanos no es posible determinar la contemporaneidad entre las ocupaciones residenciales y los eventos de entierro. Como también fue observado en otros sitios de similares características ubicados en el mismo valle, no es posible diferenciar los materiales correspondientes a las distintas ocupaciones debido a las condiciones favorables para la migración de los mismos por la alta dinámica del depósito. Los análisis tafonómicos, sobre todo el estado de meteorización, indican una buena preservación del conjunto, una transición rápida entre su depositación y entierro, y un período breve de re-exposición. Estos mismos análisis y las condiciones de los materiales al momento del hallazgo sugieren que la exposición de la mayoría de ellos se inició con las tareas modernas de desmonte y la acción eólica facilitada por la quita de la cobertura vegetal que fijaba los sedimentos. Aunque el perfil tafonómico muestra buena preservación (estadios de meteorización bajos y pocas marcas de roedores y carnívoros), la ausencia de compactación en la matriz sedimentaria (arena fina), la acción de raíces y la actividad de mamíferos cavadores sugieren condiciones de poca estabilidad espacial del conjunto.
A partir del análisis de la muestra se registraron evidencias de trozamiento de carcasas y consumo de carne y médula ósea de varias especies de fauna fluvial y terrestre. También se reconocieron algunas tendencias generales vinculadas con la composición faunística del conjunto y la importancia relativa de los distintos taxa en la subsistencia en el sitio, y con aspectos ligados a la movilidad para su captura. Aunque en este sitio el guanaco (NMI=3) constituyó el principal recurso en términos de biomasa, el rasgo principal del conjunto es la alta diversidad taxonómica y de hábitat de los animales ingresados al lugar, siguiendo un patrón similar al observado en otros sitios arqueológicos del mismo valle (Mange et al. 2013; Prates 2008; Prates y Acosta Hospitaleche 2010; Prates et al. 2010).
En el caso del guanaco, las huellas registradas sugieren actividades de trozamiento y consumo: por ejemplo la presencia de primeras falanges (cinco de un total de 10) con marcas profundas de corte en el sector medio de la diáfisis, estarían asociadas a la desarticulación y seccionamiento de los tendones extensores y flexores digitales (Mengoni Goñalons 1999: 185). Las marcas de corte sobre dos escápulas, en el sector proximal de la espina, indicarían la extracción de los músculos supraespinosos, durante el trozamiento secundario o consumo (Mengoni Goñalons 1999: 144). En el caso del venado, si bien el bajo número de elementos y marcas no permite realizar inferencias precisas sobre el trozamiento y consumo, el registro de distintas partes del esqueleto sugiere el ingreso al sitio de carcasas enteras. En cuanto al ñandú, teniendo en cuenta que el perfil tafonómico del sitio no sugiere un sesgo marcado a favor de los huesos con mayor densidad mineral ósea (Cruz y Elkin 2003), el predominio de elementos del esqueleto apendicular posterior podría indicar el traslado hacia el sitio de las partes de mayor rendimiento cárnico (Giardina 2006). Por otro lado, es interesante el registro de marcas de corte en una primera falange, lo que podría estar vinculado con la extracción de los nervios de las extremidades, cuyo uso ha sido destacado y recurrentemente registrado en la literatura etnográfica (síntesis en Salemme y Frontini 2011; Prates 2009).
En el sitio, además de mamíferos de tamaño grande (guanacos y venados), fueron aprovechados mamíferos medianos (maras y peludos) y pequeños (cuises y rata nutria), así como aves grandes (ñandúes) y pequeñas, todos ellos con huellas de corte producidas por instrumentos líticos. Podemos sumar a este conjunto algunos taxa que presentan sólo fracturas en estado fresco (M. coypus) o evidencia de termoalteración: piche, tuco-tuco, huevos de ñandú, peces y moluscos de agua dulce (gasterópodos y bivalvos). En el caso de peces y moluscos puede asumirse el transporte y la acumulación humana para todo el conjunto, teniendo en cuenta que no se detectó la presencia de proxies (e.g. diatomeas, espículas) plausibles de ser correlacionados con ambientes hídricos (Castiñeira et al. 2016).
En los conjuntos arqueofaunísticos del valle del río Negro es frecuente la presencia de numerosos restos de pequeños roedores. El hallazgo de huellas de corte en restos óseos de cuises, ratas nutrias e incluso tuco-tucos, muestra que los micromamíferos de mayor tamaño corporal (más de 200 g) fueron incorporados a la dieta (Fernández et al. 2011; Mange et al. 2014; Prates 2008). En el caso de NM3 y de otros sitios ubicados en la misma cuenca, si bien la presencia de elementos óseos con corrosión digestiva ligera podría indicar la acción de aves Strigiformes o de carnívoros (Fernández et al. 2011), hay varios elementos que añaden mayor complejidad al problema. Primero, la baja diversidad taxonómica del conjunto y la marcada selección de micromamíferos grandes es un correlato esperable de depósitos generados por seres humanos (Pardiñas 1999). Segundo, teniendo en cuenta que varias de las alteraciones corticales identificadas son puntuales y que algunos de los especímenes presentan evidencias de termoalteración, es posible que en ciertos casos la corrosión sea el producto de la acción postdepositacional de raíces (Gutiérrez 2004) sobre restos acumulados previamente por seres humanos. Además, la hipótesis de la acción de las aves Strigiformes de mayor tamaño (Tyto alba y Bubo virginianus) se ve debilitada porque los lugares donde regurgitan las egagrópilas (posaderos) no se encuentran usualmente sobre médanos (Fernández 2012). Más probable sería la participación de Strigiformes pequeñas como Athene cunicularia, que deposita los restos óseos alrededor de las cuevas de mamíferos medianos en que habita. Esta especie consume principalmente pequeños roedores (i.e. sigmodontinos), pero también en forma secundaria roedores de mayor tamaño como cuises y tuco-tucos (Andrade et al. 2004; Tommaso et al. 2009), y produce corrosión ligera a moderada sobre los restos óseos de las especies predadas (Gómez 2007; Montalvo y Tejerina 2009). Todos estos factores en conjunto hacen pensar que en la acumulación de restos de roedores en este, y posiblemente en otros sitios arqueológicos del valle del río Negro, habrían intervenido varios agentes: la caza humana, la acción de predadores y eventualmente la muerte natural de roedores fosoriales.
Aun teniendo en cuenta que los materiales analizados corresponden a, por lo menos, dos ocupaciones humanas diferentes, hay varias tendencias generales de todo el conjunto relevantes para la discusión del uso de distintos sectores del paisaje. Puntualmente, sobre la base de la evaluación del hábitat (y lugar de captura) de las especies registradas, pueden definirse las áreas o microambientes explotados por los grupos humanos durante las ocupaciónes del sitio. El conjunto analizado incluye: especies netamente acuáticas (que sólo habitan el cauce del río o los cuerpos lagunares del valle, e.g. peces y moluscos); especies ribereñas (que aunque incursionan en los cuerpos de agua lénticos y de escasa escorrentía, también ocupan una angosta franja ribereña, e.g. coipo, rata nutria); especies de pastizales abiertos del interior del valle (que raramente se aventuran fuera de la cuenca, e.g. venado); y especies que ocupan principalmente los sectores de terrazas más altas o mesetas fuera del valle, y que sólo ocasionalmente descienden a los sectores bajos de la cuenca (L. guanicoe). Esto implica un uso durante el Holoceno tardío del variado mosaico ambiental ofrecido por el área. Si ambas ocupaciones presentaran un patrón de variabilidad equivalente, lo que no puede definirse con el grado de resolución de la información disponible, se trataría de grupos con una economía de amplio espectro, al menos en cuanto a los recursos faunísticos (casos similares en Mange et al. 2013; Prates 2008).
Por otro lado, se registraron restos faunísticos que no se relacionan directamente con la explotación alimenticia de los animales. Algunos materiales habrían sido empleados como instrumento sin manufactura previa, otros fueron usados como materia prima para la fabricación de instrumentos, y otros fueron ingresados por motivos desvinculados de la esfera tecno-económica. Para el primer caso pueden mencionarse las valvas de moluscos marinos, cuyo motivo de incorporación al sitio no ha sido clarificado, pero existen algunos casos arqueológicos y etnográficos que plantean su utilización frecuente como contenedores para líquido o colorantes (Prates 2009). En análisis de arqueofaunas realizados en los últimos años (Mange et al. 2013) se han identificado contenedores, restringidos al valle del río Negro y al cercano Golfo San Matías (Deodat 1967), realizados por modificación de valvas de volútidos marinos. Si bien en otras regiones es común su depósito como acompañamiento en entierros humanos (Politis et al. 2003; Zubimendi 2010), esto no ha sido aún registrado en el área. Para el segundo caso pueden mencionarse los instrumentos óseos, elaborados sobre huesos largos de mamífero. La presencia de piqueteado (pitting) en el extremo apical (un tipo de marca que fue identificado experimentalmente como resultado de la presión del hueso contra la roca, Maigrot 2003) permite proponerlos como parte del instrumental de talla lítica (retocador). Aunque los instrumentos óseos son abundantes en el registro arqueológico de áreas vecinas, sobre todo la costa norpatagónica (Borella y Buc 2009), son escasos en el área de estudio. Hasta el momento sólo se había publicado un caso para el valle del río Negro (Prates 2008).
Algunas valvas de moluscos fluviales también fueron usadas como materia prima, aunque con un uso más ligado a lo simbólico que a lo utilitario. Con ellas se confeccionaron placas perforadas, posiblemente pendientes (Suárez Diez 2002), diferentes a las halladas en otros sitios de la misma cuenca (Prates 2008: 217-218). Una situación similar podría ser la de los restos de milodontino (Fig. 4e), ya que varias evidencias sugieren que el espécimen fue ingresado en el sitio por acción antrópica pero no para consumo. Primero, porque todo el paisaje correspondiente a la planicie aluvial del río, donde se ubica el sitio, se formó durante el Holoceno tardío. Es decir que los restos de esta especie pleistocénica no pudieron depositarse naturalmente en un ambiente tan tardío (Luchsinger 2006). Segundo, no existen en el sitio ni en sus alrededores depósitos clásticos mayores que la fracción arena y, por lo tanto, los agentes que intervinieron en su acumulación fueron de baja energía, principalmente el viento (Luchsinger 2006). Es decir que no hay evidencias que sugieran el transporte natural del resto desde contextos más antiguos. Y tercero, que las condiciones físicas y composicionales del espécimen óseo (en proceso de mineralización y con abundante carbonatación) son diferentes a las de los otros registrados en el sitio. Por lo señalado puede inferirse que el espécimen habría sido recolectado en otro lugar, ya en estado fósil, y transportado intencionalmente al sitio. Aunque las causas concretas del transporte son difíciles de precisar, pueden plantearse las motivaciones simbólicas como posibles determinantes. No sólo porque para este fragmento de vértebra no se infieren atributos de uso funcional, sino porque varias referencias sobre la recolección, uso y resignificación de los restos fósiles por parte de indígenas en el norte de la Patagonia (especialmente del orden Pilosa, al cual pertenecen los milodontes) han sido observadas en tiempos históricos (Casamiquela 1988: 2, 13). En la mayoría de los casos se otorga a los restos fósiles propiedades curativas o incluso mágicas (Casamiquela 1988: 18-22).
Consideraciones finales
Si se retira el foco de análisis del conjunto faunístico de NM3 y se lo fija en una escala espacial mayor pueden integrarse los resultados alcanzados en este trabajo con el esquema de conocimiento arqueológico disponible hasta ahora para la misma región. En términos generales, el guanaco fue la principal fuente de alimento de los cazadores-recolectores prehispánicos del interior patagónico. Sin embargo, los conjuntos del valle del río Negro (Fernández et al. 2011; Mange et al. 2013; Prates 2008; Prates y Acosta Hospitaleche 2010), y en buena medida los del valle inferior del río Colorado también (síntesis en Stoessel y Martínez 2014), sugieren una marcada diversidad de especies explotadas, con un componente importante de presas de menor porte, al menos para el Holoceno tardío. Esta tendencia general es congruente con los resultados del presente trabajo.
En una escala espacial circunscripta al río Negro, se observa que si bien la composición taxonómica de la muestra arqueofaunística de NM3 y las áreas de procedencia y captura de las especies aprovechadas son similares a las de otros sitios del valle medio del río Negro (Mange et al. 2013; Prates 2008), hay algunas diferencias que merecen una discusión más profunda. En primer lugar, en trabajos previos (Prates 2008, 2009) se había planteado como tendencia “anómala” la ausencia de evidencias de explotación de algunos mamíferos con cierto potencial para el aprovechamiento humano (por su hábitat, rendimiento cárnico y hábitos de comportamiento), como la mara y el coipo. Para explicar esas anomalías se planteó la necesidad de evaluar la existencia de algún tipo de restricción o tabú para la caza y/o consumo de estas especies teniendo en cuenta la recurrencia con que se han observado comportamientos similares en sociedades cazadoras recolectoras (discusión en Politis y Saunders 2002). Si bien los resultados aquí presentados son todavía insuficientes para descartar esa idea definitivamente porque los restos de mara y coipo son escasos en NM3 (n=5) y además las referencias sobre su aprovechamiento en tiempos históricos son también pocas (Prates 2009), la presencia de marcas de procesamiento no serían congruentes con prácticas restrictivas. Es necesario disponer de información de un número mayor de sitios para discutir esto sobre bases más firmes y, al mismo tiempo, evaluar posibles cambios en la densidad poblacional de estas especies en el área durante el Holoceno tardío que hayan podido afectar la disponibilidad para su explotación.
En segundo lugar, también se había propuesto que la captura de guanacos desde las bases residenciales ubicadas en sectores alejados de las mesetas (como ocurre en NM3), se habría realizado, principalmente, mediante estrategias organizadas logísticamente, ejecutadas por grupos pequeños de cazadores que incursionaban fuera del valle por más de una jornada (Prates 2008: 266). Esta propuesta está basada en que las planicies altas habrían sido el ámbito principal donde habita esta especie. Sin embargo, el análisis zooarqueológico realizado en NM3 no parece del todo compatible con el ingreso de presas obtenidas mediante partidas logísticas realizadas lejos de las bases residenciales, cuya principal expectativa sería la presencia de las partes anatómicas de mayor rinde debido al transporte diferencial de partes desde las áreas de captura (Binford 1981; Mengoni Goñalons 1999; Perkins y Daly 1968, entre otros). La evidencia de este y otros sitios del valle medio del río Negro, Angostura 1 y Negro Muerto 1 (Prates 2008), Loma de los Muertos (Mange et al. 2013) y Pomona (Mange y Ramos van Raap 2010) muestra el ingreso de elementos óseos de todo el esqueleto de individuos adultos y juveniles. Una hipótesis alternativa, para cuyo testeo se requiere incorporar un volumen mayor de información arqueológica de la región y etológica del guanaco, podría ser que estos fueran cazados en el interior del valle y transportados enteros a los sitios.
Para finalizar, a lo largo de este trabajo hemos planteado los aspectos taxonómicos y tafonómicos principales del sitio. Sobre algunos temas se plantearon algunas explicaciones alternativas a las propuestas con anterioridad para sitios de la misma zona pero quedan todavía algunos interrogantes, como la presencia de evidencias algo ambiguas (de acción antrópica y de corrosión) en numerosos restos de micromamíferos y el ingreso de esqueletos enteros de guanacos a sectores supuestamente poco frecuentados por la especie. La incorporación de un mayor volumen de información (sólo se han analizado conjuntos faunísticos de cinco sitios arqueológicos del valle del río Negro), permitirá responder estos interrogantes y reformular los modelos propuestos con anterioridad sobre la ocupación humana del área durante el Holoceno tardío.
Agradecimientos: queremos expresar nuestro agradecimiento a Alejandro Serna por su colaboración en los trabajos de campo. A Gustavo Carus por su buena predisposición para trabajar dentro de su propiedad, y a Daniel Cabaza por la oportuna denuncia de los hallazgos. A Alfredo Carlini y Eduardo Tonni, que realizaron la determinación de restos de megafauna. A Carola Castiñeira, Marco Álvarez y Adriana Blasi por el análisis de las muestras sedimentarias. Este trabajo fue realizado con financiamiento de la ANPCyT (PICT-2012-0242) y en convenio con la Secretaría de Cultura de la provincia de Río Negro.
Bibliografía citada
1. Andrade, A.; Udrizar Sauthier, D. y U. Pardiñas 2004 Vertebrados depredados por la lechucita vizcachera (Athene cunicularia) en la Meseta de Somuncurá (Río Negro, Argentina). El Hornero 19 (2): 91-93. [ Links ]
2. Andrews, P. 1990 Owls, Caves and Fossils. University of Chicago Press, Chicago. [ Links ]
3. Apolinaire, E. y L. Turnes 2010 Diferenciación específica de rheidos a partir de fragmentos de cáscaras de huevo. Su aplicación en sitios arqueológicos del Holoceno Tardío. En Mamül Mapu: Pasado y Presente desde la Arqueología Pampeana, M. Berón, L. Luna, M. Bonomo, C. Montalvo, C. Aranda y M. Carrera Aizpitarte (eds.), pp. 215-222. Libros del Espinillo, Buenos Aires. [ Links ]
4. Barone, R. 1990 Anatomía Comparada de los Mamíferos Domésticos. Tomo 1: Osteología. Hemisferio Sur, Buenos Aires. [ Links ]
5. Behrensmeyer, A. 1978 Taphonomic and ecologic information from bone weathering. Paleobiology 4(2): 150-162. [ Links ]
6. Binford, L. 1981 Bones. Ancient Men and Modern Myths. Academic Press, New York. [ Links ]
7. Borella, F. y N. Buc 2009 Ópticas y ópticos. Una aproximación a la tecnología ósea en la Bahía de San Antonio Oeste (Río Negro). En Arqueología de Patagonia: Una Mirada desde el Último Confín, M. Salemme, F. Santiago, M. Álvarez, E. Piana, M. Vázquez y M. Mansur (eds.), pp. 421-432. Utopías, Ushuaia. [ Links ]
8. Brandoni, D.; Ferrero, B. y E. Brunetto 2010 Mylodon darwini Owen(Xenarthra, Mylodontinae) from the Late Miocene of Entre Ríos Province, Argentina, with remarks on individual variability, paleobiology, paleobiogeography, and paleoenvironment. Journal of Vertebrate Paleontology 30(5): 1547-1558. [ Links ]
9. Casamiquela, R. 1988 En Pos del Gualicho. Fondo Editorial Rionegrino, Viedma. [ Links ]
10. Castiñeira, C.; Álvarez, M. y A. Blasi 2016 Informe sedimentológico de muestras del sitio Negro Muerto 3. División Mineralogía, Petrología y Sedimentología, Museo de La Plata. Ms. [ Links ]
11. Caviglia, S. y L. Borrero 1981 Consumo de moluscos en el Alero de los Sauces (Neuquén, Argentina). Trabajos de Prehistoria 1: 77- 87. [ Links ]
12. Crivelli Montero, E.; Pardiñas, U.; Fernández, M.; Bogazzi, M.; Chauvin, A.; Fernández, V. y M. Lezcano 1996 La Cueva Epullán Grande (provincia del Neuquén, Argentina). Informe de avance. Præhistoria 2: 185-265. [ Links ]
13. Cruz, I. y D. Elkin 2003 Structural bone density of the Lesser Rhea (Pterocnemia pennata) (Aves: Rheidae). Taphonomic and archaeological implications. Journal of Archaeological Science 30: 37-44. [ Links ]
14. Deodat, L. 1967 Una antigua manufactura valvácea en el golfo San Matías (Argentina). Runa X: 319-353. [ Links ]
15. Fernández, F. 2012 Microvertebrados del Holoceno de Sitios Arqueológicos en el Sur de Mendoza (República Argentina): Aspectos Tafonómicos y sus Implicancias en la Subsistencia Humana. Tesis de Doctorado. Facultad de Ciencias Naturales y Museo, Universidad Nacional de La Plata. [ Links ]
16. Fernández, F. y L. De Santis 2013 Tafonomía de restos de micromamíferos recuperados del sitio arqueológico Agua de la Mula (Mendoza, Argentina) y su implicancia en la subsistencia humana. Cuadernos del INAPL, Series Especiales 1: 181-195. [ Links ]
17. Fernández, F.; Del Papa, L.; Moreira, G.; Prates, L. y L. De Santis 2011 Small mammal remains recovered from two archaeological sites in the middle and lower Negro River valley (Late Holocene, Argentina): taphonomic issues and paleoenvironmental implications. Quaternary International 245: 135-147. [ Links ]
18. Fernández-Jalvo, Y. y P. Andrews 1992 Small mammal taphonomy of Gran Dolina, Atapuerca (Burgos, Spain). Journal of Archaeological Science 19: 407-428. [ Links ]
19. Folk, R. 1951 Stages of textural maturity in sedimentary rocks. Journal of Sedimentary Petrology 21: 127-130. [ Links ]
20. Giardina, M. 2006 Anatomía económica de Rheidae. Intersecciones en Antropología 7: 263-276. [ Links ]
21. Giardina, M. 2010 El Aprovechamiento de la Avifauna entre las Sociedades Cazadoras-recolectoras del Sur de Mendoza, Un Enfoque Arqueozoológico. Tesis de Doctorado. Facultad de Ciencias Naturales y Museo, Universidad Nacional de La Plata. [ Links ]
22. Gómez, G. 2007 Predators categorizations based on taphonomic analysis of micromammals bones: a comparison to proposed models. En Taphonomy and Zooarchaeology in Argentina, M. Gutiérrez, L. Miotti, G. Barrientos, G. Mengoni Goñalons y M. Salemme (eds.), pp. 1-16. BAR International Series 1601, Oxford. [ Links ]
23. Grayson, D. 1991 Alpine faunas from the White mountains, California: adaptive change in the Late Prehistoric Great Basin? Journal of Archaeological Science 18: 483-506. [ Links ]
24. Gutiérrez, M. 2004 Análisis Tafonómicos en el Área Interserrana (Provincia de Buenos Aires). Tesis de Doctorado. Facultad de Ciencias Naturales y Museo, Universidad Nacional de La Plata. [ Links ]
25. Kaufmann, C. 2009 Estructura de Edad y Sexo en Guanaco. Estudios Actualísticos y Tafonómicos en Pampa y Patagonia. Sociedad Argentina de Antropología, Buenos Aires. [ Links ]
26. Luchsinger, H. 2006 The Late Quaternary Landscape History of the Middle Rio Negro Valley, Northern Patagonia, Argentina: Its Impact on Preservation of the Archaeological Record and Influence on Late Holocene Human Settlements Patterns. Tesis de Doctorado. Texas A&M University. [ Links ]
27. Lyman, R. 1994 Vertebrate Taphonomy. Cambridge University Press, Cambridge. [ Links ]
28. Maigrot, Y. 2003 Etude Technologique et Fonctionnelle de l'Outillage en Matières Dures Animales: La Station 4 de Chalain (Néolithique Final, Jura, France). Tesis de Doctorado. Université de Paris. [ Links ]
29. Mange, E. y A. Ramos Van Raap 2010 Recursos faunísticos explotados en el valle del río Negro: el caso del sitio Pomona (Provincia de Río Negro). Trabajo presentado en el XVII Congreso Nacional de Arqueología, Mendoza. Ms. [ Links ]
30. Mange, E.; Ramos Van Raap, A. y D. León 2013 La arqueofauna del sitio Loma de los Muertos (departamento de General Conesa, Río Negro). Intersecciones en Antropología 14(2): 301-314. [ Links ]
31. Mange, E.; Fernández, F. e I. Moyano 2014 Taphonomical evidences of micromammals exploitation at Pomona archaeological site (Late Holocene, North Patagonia, Negro River). Does it constitute an intensification process? Trabajo presentado en el XII ICAZ, San Rafael, Mendoza. Ms. [ Links ]
32. Medina, M.; Teta, P. y D. Rivero 2012 Burning damage and small-mammal human consumption in Quebrada del Real 1 (Córdoba, Argentina): an experimental approach. Journal of Archaeological Science 39: 737-743. [ Links ]
33. Mengoni Goñalons, G. 1999 Cazadores de Guanaco en la Estepa Patagónica. Sociedad Argentina de Antropología, Buenos Aires. [ Links ]
34. Montalvo, C. y P. Tejerina 2009 Análisis tafonómico de los huesos de anfibios y roedores depredados por Athene cunicularia (Strigiformes, Strigidae) en La Pampa, Argentina. En Mamül Mapu: Pasado y Presente desde la Arqueología Pampeana, M. Berón, L. Luna, M. Bonomo, C. Montalvo, C. Aranda y M. Carrera Aizpitarte (eds.), pp. 323-334. Libros Del Espinillo, Buenos Aires. [ Links ]
35. Pacheco, V.; Altamirano, A. y E. Guerra 1979 Guía Osteológica de Camélidos Sudamericanos. Universidad Nacional Mayor de San Marcos, Lima. [ Links ]
36. Pardiñas, U. 1999 Tafonomía de microvertebrados en yacimientos arqueológicos de Patagonia (Argentina). Arqueología 9: 265-340. [ Links ]
37. Pearson, O. 1995 Annotated keys for identifiyng small mammals living in or near Nahuel Huapi National Park or Lanin National Park southern Argentina. Mastozoología Neotropical 2: 99-148. [ Links ]
38. Perkins, D. y P. Daly 1968 A hunter's village in Neolithic Turkey. Scientific American 219(5): 96-106. [ Links ]
39. Politis, G. y N. Saunders 2002 Archaeological correlates of ideological activity: food taboos and spirit-animals in an Amazonian hunter-gatherer society. En Consuming Passions: Archaeological Studies of Material Culture, P. Miracle (ed.), pp. 113-130. Mc Donald Institute, Cambridge. [ Links ]
40. Politis, G.; Bonomo, M. y L. Prates 2003 Territorio y movilidad entre la costa y el interior de la región pampeana (Argentina). Estudios Ibero-Americanos 29(1): 10-35. [ Links ]
41. Prates, L. 2008 Los Indígenas del Río Negro. Un Enfoque Arqueológico. Sociedad Argentina de Antropología, Buenos Aires. [ Links ]
42. Prates, L. 2009 El uso de los recursos por los cazadores-recolectores posthispánicos de Patagonia continental y su importancia arqueológica. Relaciones de la Sociedad Argentina de Antropología XXXIV: 201-229. [ Links ]
43. Prates, L. y C. Acosta Hospitaleche 2010 Las aves de sitios arqueológicos del Holoceno Tardío en Norpatagonia, Argentina. Los sitios Negro Muerto y Angostura 1 (Río Negro). Archaeofauna 19: 7-18. [ Links ]
44. Prates, L. y N. Marsans 2007 El uso de moluscos de agua dulce (Diplodon chilensis patagonicus) en el sitio Angostura 1 (departamento de General Conesa, Río Negro).Intersecciones en Antropología 8: 355-359. [ Links ]
45. Prates, L. ; Di Prado, V. ; Mange, E. y A. Serna 2010 Sitio Loma de los Muertos. Múltiples ocupaciones sobre un médano del este de Norpatagonia (Argentina). Magallania 38(1): 165-181. [ Links ]
46. Prates, L. ; Luchsinger, H.; Scabuzzo, C. y D. Mansegosa 2011 Investigaciones arqueológicas en el sitio La Victoria 5 (departamento de General Conesa, Río Negro). Intersecciones en Antropología 12: 155-167. [ Links ]
47. Redford, K. y J. Eisenberg 1992 Mammals of the Neotropics. The Southern Cone. The University of Chicago Press, Chicago. [ Links ]
48. Salemme M. y R. Frontini 2011 The exploitation of Rheidae in Pampa and Patagonia (Argentina) as recorded by chroniclers, naturalists and voyagers. Journal of Anthropological Archaeology 30: 473-483. [ Links ]
49. Stoessel, L. y G. Martínez 2014 El proceso de intensificación en la transición pampeano-patagónica oriental. Discusión y perspectivas comparativas con regiones aledañas. Comechingonia 18(2): 65-94. [ Links ]
50. Suárez Diez, L. 2002 Tipología de los Objetos Prehispánicos de Concha. Instituto Nacional de Antropología e Historia, México. [ Links ]
51. Tommaso, D. de; Callico Fortunato, R.; Teta, P. y J. Pereira 2009 Dieta de la lechucita vizcachera (Athene cunicularia) en dos áreas con diferente uso de la tierra en el centro-sur de la provincia de La Pampa, Argentina. El Hornero 24(2): 87-93. [ Links ]
52. Zubimendi, M. 2010 Malacological artifacts in Argentine Patagonia. Munibe. Suplemento-Gehigarria 31: 262-270. [ Links ]

