










Serviços Personalizados
Journal

Artigo
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares em
SciELO
Similares em
SciELO
Compartilhar

 Permalink
PermalinkComechingonia
versão On-line ISSN 1851-0027
Comechingonia vol.24 no.1 Córdoba abr. 2020
articulos
¿QUÉ HACÍAN LOS PASTORES CON LOS HUESOS?
WHAT DID HERDERS DO WITH THE BONES?
Hugo Yacobaccio1 y Celina Medero1
1Instituto de Arqueologia, Facultad de Filosofía y Letras, Universidad de Buenos Aires.
25 de Mayo 217, tercer piso, (1002) Buenos Aires, Argentina, hdyacobaccio@gmail.com
Introducción
El registro arqueológico atribuible a pastores andinos está pobremente definido. Además de los problemas generales a este tipo de contexto en el resto del mundo (Chang y Koster 1986; Robertshaw y Collett 1983; Voigt 1986), en los Andes se suma una dificultad adicional: la difícil distinción osteológica entre las diferentes especies de camélidos domesticados de silvestres. Sin embargo, últimamente se han dado pasos promisorios en este sentido (Kent 1982).
A pesar de esta ambigüedad intentaremos plantear aquí algunas cuestiones preliminares en torno al análisis faunístico de sociedades pastoriles: caracterizar mínimamente las variables intervinientes y formular un modelo de cadena logística de explotación animal en tal contexto.
Para esto nos basaremos en la formulación de modelos de prácticas de trozamiento por parte de pastores actuales: evaluaremos dos sitios arqueológicos a partir de tales modelos y generaremos expectativas en relación a la explotación y aprovechamiento animal de acuerdo a necesidades logísticas. El análisis de representación de partes esqueletarias y el uso de índices económicos medidos en términos de rendimiento de carne adherida a los huesos, será la herramienta metodológica empleada de manera esencial. Estos estudios están fundamentados teóricamente en otros trabajos que no comentaremos aquí por razones de espacio (Binford 1978, 1984; Borrero 1986; Thomas y Meyer 1983).
Para nuestro interés es importante señalar que 1) el sistema pastoril andino no es totalmente sedentario ya que está organizado en función de dar cuenta de la disponibilidad estacional diferencial de pasturas (Flores Ochoa 1968; Inamura 1981) y 2) los tiempos de ocupación de los asentamientos son variables y desiguales durante el ciclo anual. La movilidad, entonces, es un factor organizativo importante a tener en cuenta al tratar de definir la actividad humana hacia el rebaño.
En la Puna de Jujuy, Rabey (1986) registró dos sistemas de movilidad; sin embargo, ambos coinciden en algunos aspectos estructurales. Ocupan la base residencial ("la casa") por 150 días, y dos asentamientos temporarios o "puestos" por 90 días cada uno, cubriendo en los traslados un promedio de 121.9 km2 anualmente. Estos movimientos están relacionados con la disponibilidad de pasturas localizadas a diferentes altitudes. Permanecen en la base residencial en la estación húmeda, pero retornan a ella por 15-20 días en julio/agosto para sacrificar llamas jóvenes y manufacturar charqui. Aunque reconocemos la variabilidad existente en el sistema de asentamiento de los pastores (hemos registrado otros dos sistemas distintos en el norte de la Quebrada de Humahuaca), este será nuestro modelo de referencia ya que está basado en un grupo pastor de llamas. Obviamente, la presencia de ovicápridos altera los patrones de movilidad tradicional ligados a la explotación de camélidos.
La actividad económica pastoril es planteada como base de subsistencia para algunos desarrollos Formativos del área Circumpuneña (Nuñez 1981; Olivera 1986; Raffino 1977). Nosotros tomaremos dos sitios pertenecientes a tal período para ejemplificar nuestro análisis. Ellos son el Montículo Chávez (Chávez de ahora en adelante) y ChiuChiu 200 (ChiuChiu en adelante). El primero está localizado en ambiente de planicie de Puna, en la cuenca sedimentara de Antofagasta de la Sierra (Prov. de Catamarca) a 3450 msnm. El montículo tiene 280 m2 y se estima el comienzo de la ocupación en ca. 1800 años AP (Olivera y Nasti 1988). ChiuChiu se encuentra en la terraza más alta de la ribera este del río Loa (II Región, Chile) y cubre una superficie total de 28.693 m2. Se trata de un poblado conformado por estructuras habitacionales de planta circular (Benavente 1982). Posee tres fechas de termoluminiscencia cuyo promedio es de 2890 ± 290 AP. La lista de fauna base de nuestro análisis está tomada de Benavente (1982: 85). Las muestras empleadas por nosotros provienen de estructuras de basural en ambos casos.
Modelos actuales de trozamiento
Proponemos aquí dos modelos. El Modelo I (Tabla 1) está derivado de los datos de Miller (1979) de eventos de descuartizamiento en la Puna meridional de Perú. El Modelo II fue construido sobre observaciones efectuadas por el primer autor en la zona de llanco (Puna de Catamarca) en noviembre de 1984 (Tabla 2). Los índices de utilidad empleados son los del guanaco (Lama guanicoe) elaborados por Borrero (1986).
En la lectura de las Tablas 1 y 2 se podrá observar que los dos modelos incluyen el mismo número de unidades. Están representados cortes muy similares pero ordenados de manera un tanto diferente. El modelo I enfatiza un rendimiento óptimo de carne en algunas unidades de trozamiento. El Modelo II produce más unidades con rendimiento relativamente equivalentes.
Esta diferencia puede ser referida tanto a diversas necesidades de procesamiento y transporte o a factores de tradición cultural (ver Miller 1979). No podemos decidir en este caso, pero nos parece que el Modelo II es más adecuado allí donde las necesidades de transporte son más imperiosas.
Los huesos no son fracturados durante el proceso de trozamiento; si lo son, posteriormente, para preparar las unidades de consumo. Sin embargo, a pesar de esto, los valores de las unidades se conservan ya que cada corte es procesado por separado conservando su valor de utilidad general.
Tabla 1. Modelo I. (*) Índice de utilidad del guanaco.
| Corte | Huesos | IUG (*) | ||
| Mak´in | Costilla (-1) | 96.15 | ||
| Escápula | 38.4 | |||
| Húmero | 23.8 | |||
| Radiocúbito | 7.8 | |||
| Ch´akin | Fémur | 83.2 | ||
| Tibia | 21.3 | |||
| Sacro | - | |||
| Kunka | V. Cervicales | 51.3 | ||
| Atlas | 8.8 | |||
| P´altawasan | V. Torácicas | 22.1 | ||
| 1 costilla | - | |||
| V. Lumbares | 44.9 | |||
| Pelvis | 40.2 | |||
| Chuqchuku | Metapodios | 1.5 | ||
| Carpo | 1.6 | |||
| Tarso | 1.6 | |||
| - | Falanges | 2.1 | ||
| - | Cráneo | 10 | ||
| Mandíbula | 5.7 | |||
Análisis de las muestras
Las partes esqueletarias presentes en los sitios, más las unidades anatómicas mínimas (MAU) pueden verse en la Tabla 3. El control tafonómico aplicado a las muestras implica la evaluación de la destrucción diferencial causada por la diferencia de densidad de los huesos. Ya Binford y Bertram (1977) habían resaltado el factor de densidad diferencial de los huesos como causa principal de su destrucción (ver también Brain 1981). En este trabajo se empleó el índice de densidad global (Lyman 1985). (Para un análisis más detallado de Chávez ver olivera y Nastri 1988).
Se correlacionó el valor de cada MAU con su respectivo valor de densidad empleando el coeficiente de correlación r de Pearson. Los resultados obtenidosfueron:
Chávez: N: 13 r= 0.139 p<0.001
ChiuChiu: N: 25 r=0.330 p<0.05
De acuerdo a estos resultados ningún de los contextos está severamente afectado por destrucción diferencial causada por el factor de densidad.
Tabla 2. Modelo II. *Índice de utilidad de guanaco
| Corte | IUG (*) | |
| 1 | Escápula | 38.4 |
| Húmero | 23.8 | |
| Radiocúbito | 7.8 | |
| 2 | Costillas | 100 |
| 3 | V. Cervicales | 51.3 |
| Atlas-Axis | 8.8 | |
| 4 | V. Torácica | 22.1 |
| V. Lumbares | 44.9 | |
| Sacro | ||
| 5 | Fémur | 83.2 |
| Tibia | 21.3 | |
| Pelvis | 40.2 | |
| 6 | Metapodios | 1.5 |
| Falanges | 2.1 | |
| 7 | Cráneo | 10 |
| Mandíbula | 5.7 | |
Las partes esqueletarias presentes en Chávez sugieren que fueron desarrolladas actividades de matanza selectiva. Metapodios, cráneos y mandíbulas están altamente representadas; esto estaría relacionado con matanza, cuereado y trozamiento primario de los animales en el sitio. Hay, además, proporciones bajas de unidades de trozamiento que encajan bien con los cortes 1,2 y 5 del Modelo II, de alto rendimiento.
Como se ve en la tabla 3, en ChiuChiu, hay una baja representación de esqueleto axial en general -como en Chávez- y altamente selectivas en cuanto a las del esqueleto apendicular: tibias distales, articulaciones tarsianas y metatarsos. Hay una alta representación de escápulas y pelvis. También hay una moderada representación de mandíbulas. Esto sugiere consumo de partes de alto rendimiento -escápulas y pelvis- descartadas luego en el basural y la entrada de patas traseras. La presencia de mandíbulas es más enigmática aunque podría estar vinculada a técnicas particulares de cuereo. Hay huellas de corte que afectan el cuerpo de las mismas.
Es interesante hacer algunas comparaciones entre ambos sitios. Aunque presentan rayos de similitud general hay algunas diferencias: entre éstas la distribución de huesos de mediano rendimiento (Tabla 4).
La diferencia está dada en la presencia de escápulas y pelvis como corte aparentemente independientes en ChiuChiu. Mientras en Chávez se ajusta a los patrones de trozamiento sugeridos por el Modelo II, ChiuChiu está más lejos de estos.
Pero, cómo se explican los conjuntos óseos en general? Pensamos que las partes esqueletarias presentes en los sitios responde principalmente al remanente de actividades relacionadas con la manufactura de charqui. Miller (1979) llamó la atención sobre el significado del transporte de carne en forma de charqui razonando que esta práctica determinará en alguna medida representación de prácticas esqueletarias en los sitios arqueológicos. Propuso detectar este "efecto" mediante la relación entre la parte baja de las patas (pie) y las partes altas (pata). Una mayor proporción de pie indicaría manufactura de charqui mientras que el caso contrario indicaría consumo de carne desecada. Para prepararcharqui según Miller: All parts of the carcass are utilized with the exception of the lower legs and the heads which are never used (Miller 1979: 99).
Dentro del término "parte baja de las patas" incluye: metapodios, falanges y tal vez, huesos del carpo y tarso, dependiendo de la técnica de trozamiento.
Browman (1986) señaló que la manufactura de charqui no es uniforme y que por lo tanto deben esperarse variaciones al índice propuesto por Miller. Browman observó que cabezas y pies eran vendidos de forma de charque en mercados bolivianos.
Tabla 3. Partes esqueletarias en los sitios. (+) Los cráneos no se emplearon para calcular el porcentaje de MAU por carecer de datos sobre fragmentación.
| Montículo Chávez | Chiu-Chiu 200 | |||||
| Q | MAU | % | Q | MAU | % | |
| Cráneo | 12 | 12 | -(+) | 10 | 10 | -(+) |
| Maxilla | 3 | 3 | 24 | - | - | - |
| Mandíbula | 21 | 10.5 | 84 | 9 | 4.5 | 50 |
| Atlas | - | - | - | 1 | 1 | 11.11 |
| Axis | - | - | - | - | - | - |
| V. cervical | - | - | - | 6 | 0.8 | 8.88 |
| V. torácica | 36 | 1 | 8 | 3 | 0.25 | 2.77 |
| | - | - | - | 5 | 0.71 | 7.88 |
| Sacro | - | - | - | - | - | - |
| Costillas | 36 | 1.5 | 12 | 12 | 0.5 | 5.55 |
| Escápula | 8 | 4 | 32 | 18 | 9 | 100 |
| Húmero proximal | 2 | 1 | 8 | 3 | 1.5 | 16.66 |
| Húmero distal | 5 | 2.5 | 20 | 6 | 3 | 33.33 |
| Radiocúbitoprox. | 2 | 1 | 8 | 5 | 2.5 | 27.77 |
| Radiocúbito distal | 7 | 3.5 | 28 | 7 | 3.5 | 38.88 |
| Carpo | - | - | - | 17 | 1,21 | 13.44 |
| Metacarpo prox. | - | - | - | 3 | 1.5 | 16.66 |
| Metacarpo distal | - | - | - | 1 | 0.5 | 5.55 |
| Pelvis | 9 | 4.5 | 36 | 12 | 6 | 66.66 |
| Fémur proximal | 3 | 1.5 | 12 | 2 | 1 | 11.11 |
| Fémur distal | 2 | 1 | 8 | 4 | 2 | 22.22 |
| Tibia proximal | 3 | 1.5 | 12 | 2 | 1 | 11.11 |
| Tibia distal | 5 | 2.5 | 20 | 11 | 5.5 | 61.11 |
| Tarso | 22 | 4.5 | 36 | 9 | 0.9 | 10 |
| Metatarso prox. | - | - | - | 5 | 2.5 | 27.77 |
| Metatarso distal | - | - | - | 11 | 5.5 | 61.11 |
| Astrágalo | - | - | - | 16 | 8 | 88.88 |
| Calcáreo | - | - | - | 11 | 5.5 | 61.11 |
| Falange 1 | - | - | - | 28 | 3.5 | 38.88 |
| Falange 2 | 40 | 2.6 | 12.8 | 14 | 1.75 | 19.44 |
| Falange 3 | - | - | - | - | - | - |
| Metapodios | 50 | 12.5 | 100 | - | - | - |
Tabla 4
| Chávez | Chiu-Chiu 200 | |||
| Bajo rendimiento (0-20) | 56.8 | 73% | 50.86 | 61.93% |
| Medio rendimiento /21-40) | 11.5 | 14.21% | 20 | 24.75% |
| Ato rendimiento (+40) | 9.5 | 12.21% | 11.26 | 13.71% |
| Total | 77.8 | 82.12 | ||
De esta breve presentación puede extraerse dos elementos importantes: 1) que las formas de manufactura de charqui son variables regionalmente y 2) si bien el índice pie:pata no puede emplearse como indicador absoluto, puede utilizarse para monitorear el procesamiento de miembros, descartados otros elementos recurrentes.
Como se ve, el "efecto charqui" está pobremente reconocido en cuanto a las consecuencias arqueológicas que genera. Preliminarmente, entonces planteamos un modelo que propone -en orden de importancia- las partes más adecuadas para desecar:
1) Pata delantera posterior
2) Para trasera superior
3) Escápula
4) Pelvis
5) Costillas
6) Cabeza (Browman 1986)
Este modelo permite explicar la baja presencia de huesos de alto rendimiento en las bases residenciales de grupos pastores.
En el caso de Chávez la manufactura de charqui estaría dada por el descarte de metapodios que no pudieron entrar con el cuero dada la disparidad de representación con las falanges (Figura 1), y la coincidencia del modelo anterior con los cortes 1, 2 y 5 del Modelo II. La manufactura de charqui con el fin de transportar carne a otro sitio se ve sostenida por la afirmación de Haber (1988) en el sentido de que la hoyada de Antofagasta de la Sierra, donde está localizado el sitio, tiene un potencial forrajero limitado. Esto determinaría un traslado de rebaños hacia las vegas puneñas y los pajonales al menos durante una parte del año. Por otra parte, el modelo de asentamiento de pastores aquí adoptado también favorecería esta opción.
El caso de ChiuChiu presenta una situación en la cual se explicaría la baja representación de tibias proximales y fémures. El procesamiento en el lugar de patas traseras, que habrían ingresado originalmente completas al sitio, puede ser apoyado por la alta representación de rótulas (50 % de MAU), correspondientes a la articulación entre la tibia y el fémur. Este procesamiento de patas estaría reforzado por la presencia de huellas de corte en el fémur proximal, escápula y radiocúbito proximal (Figura 2).
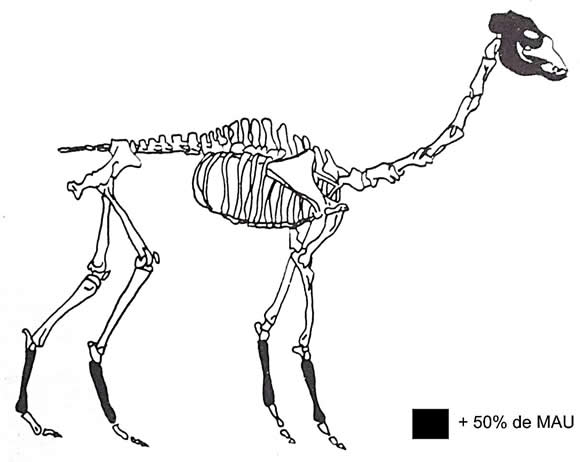
Figura 1. Montículo Chávez.
En este sentido, la relación parte baja-parte alta de las patas traseras es relevante: 2.2:1para ChiuChiu y 3.8:1 para Chávez. ChiuChiu presenta un patrón de consumo algo distinto con descarte de escápulas y pelvis, aunque en Chávez se puede visualizar algo similar en menor proporción (32% y 36% de MAU respectivamente). Análisis recientes de Olivera muestras la presencia de escápulas como corte independiente.
En síntesis, pensamos que las partes de alto rendimiento fueron transportadas en charqui y llevadas de los sitios. Los cortes de Chávez estarían más de acuerdo con el Modelo II, mientras que las técnicas de trozamiento de ChiuChiu diferían algo.
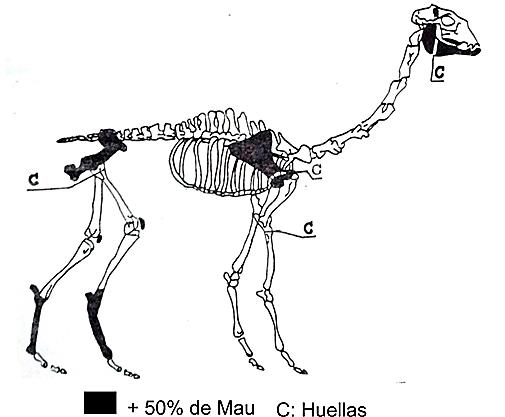
Figura 2. Chiu Chiu-200.
¿Eran domesticados todos los camélidos consumidos?
La pregunta, aunque sorprenda, es pertinente. En ChiuChiu, a través de la fibra, se identificó la presencia no sólo de llama, sino también de guanaco y vicuña (Benavente 1982:86). Por lo tanto, los contextos óseos de sitios de pastores poseen animales silvestres cazados. Esto está de acuerdo con el empleo de técnicas de caza colectivas durante momentos prehistóricos tardíos.
De los documentos españoles se deduce que fueron empleados dos métodos diferentes: el chaco y el caycu. El primero era una técnica de encierro en el que el grupo de hombres formaba un gran círculo en un territorio plano capturando así diversos animales. Luego se realizaba una matanza selectiva de camélidos adultos para la obtención de carne para secado y almacenamiento (charqui). El caycuera una técnica que dirigía a los animales a corrales o lugares cercados naturalmente.
Aún no están claras las consecuencias arqueológicas de ambos métodos, pero se pueden establecer algunas expectativas. El chaco fue una caza ritual en tiempos incaicos. En los sitios de habitación pueden haberse descartado las partes esqueletarias transportadas para el secado de carne. Así pues este tipo de sitio debería presentar partes de alto rendimiento de camélidos adultos silvestres. Por el contrario, el caycupuede ser identificado en sitios al aire libre cercanos a desfiladeros o gargantas. La principal especie cazada era la vicuña. El caycu era realizado por pastores para asegurarse abastecimiento de carne si necesidad de matar a sus rebaños. En los sitios de matanza deberían haberse descartado huesos de bajo rendimiento, mientras que las partes de mayor rinde deben haber sido transportadas a los campamentos temporarios o sitios de habitación.
Junto a las prácticas de caza colectiva también existieron cazas individuales llevadas a cabo por unos pocos hombres. La caza individual debe ser relacionada con ocupaciones en sitios temporarios tales como cuevas y aleros rocosos, que habrían actuado como localidades de procesamiento primario. En estos sitios se esperarán partes esqueletarias de bajo rendimiento aunque la situación puede alterarse debido a elecciones logísticas relacionadas con el transporte de las unidades procesadas.
Tanto la caza individual como colectiva parecen haber actuado como importantes fuentes de abastecimiento de camélidos para las comunidades pastoriles. Matienzo (1964:276), relata la existencia de grupos de cazadores en el altiplano boliviano en una época tan tardía como el siglo XVI.
Esto plantea alternativas interesantes para desarrollar, no sólo en relación al contexto óseo, sino también acerca de los distintos tipos de sitios presentes en el sistema de asentamiento. Veremos este punto en el próximo acápite.
Discusión y Conclusiones
Para la obtención de carne los pastores implementaron dos estrategias: 1) explotación de camélidos silvestres mediante la caza y 2) matanza selectiva de animales de rebaño. Es de esperar que ambas produzcan variabilidad en el registro arqueológico.
La actividad de caza estaría marcada por la existencia de una cadena logística más larga e involucraría mayor cantidad de sitios en su desarrollo. En la Tabla 5 se sintetiza este problema tomando en cuenta el factos distancia como la variable singular que más afecta el proceso.
Un rápido análisis de la tabla en cuestión muestra que la distancia no incide sobre la explotación de animales de rebaño, mientras que puede ser un factor de peso cuando se toman decisiones con respecto al transporte de productos de caza. En este caso, la dispersión de partes esqueletarias debería ser mayor. En el caso de los animales de rebaño la dispersión de partes fuera del asentamiento original ocurriría luego de su transformación en charqui y, según las categorías empleadas en la Tabla 5, luego del primer consumo. Obviamente, no debe esperarse lo mismo para el primer caso.
Binford (1980) ha enfatizado que cuanto mayor son los componentes logísticos del sistema, la variabilidad intersitio tenderá a incrementarse. Esto es verdad si el ritmo de movilidad es mantenido constantemente. Pero si hay una movilidad decreciente, entonces el número de actividades en cada sitio del sistema tenderá a aumentar disminuyendo, de esta manera, la variabilidad intersitio. Los sitios pastores son ocupados redundantemente con estadías largas, por lo tanto, se espera que una variabilidad intersitio menor que en el caso de los grupos cazadores-recolectores acopiadores (collectors), aunque esta variabilidad se verá complicada por los factores de explotación faunística diferencial.
Tabla 5. Referencias. =: Mismo sitio (distinto locus); C: cercano; L: lejano.
| Matanza | P. Procesamiento | P. Consumo | ||||
| Caza | Rebaño | Caza | Rebaño | Caza | Rebaño | |
| Matanza | - | - | =/C | = | C/L | = |
| Primer Procesamiento | =/C | = | - | - | C/L | = |
| Primer consumo | C/L | = | C/L | = | = | - |
¿Cuál es la moraleja que podemos extraer?: 1) que debemos esforzarnos por distinguir en el contexto faunístico animales silvestres de domesticados; 2) que el relevamiento estricto de las unidades de proveniencia espaciales (basural, habitación, patrio) de los sitios arqueológicos de pastores pueden llegar a ser la única pista para distinguir las distintas actividades de procesamiento y consumo efectuadas en las bases residenciales; 3) que debemos considerar la mayor variedad posible de tipos funcionales de sitios dentro del análisis.
Expresamos recién que la cadena logística de explotación de animales de rebaño tiene particularidades propias. Seguidamente proponemos un modelo de cadena logística de explotación ganadera. Esta se detalla en la figura 3.
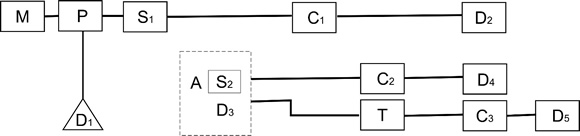
Figura 3. M: Matanza de individuo camélido. P: Procesamiento o trozamiento inicial del animal. D1: Descarte. Partes esqueletarias descartadas en el procesamiento. S1: Selección de partes destinadas a: C1: Consumo inmediato; D2: partes esqueletarias descartadas luego del consumo inmediato; S2: partes seleccionadas para la manufactura de charqui; D3: descarte de partes luego de la manufactura de charqui; C2: consumo diferido sin sujeción a trasporte (en el mismo asentamiento de manufactura); D4: descarte de partes esqueletarias producto de C2. A: Almacenamiento. T: Transporte. C3: consumo diferido sujeto a transporte (en asentamiento diferente al lugar de manufactura de charqui). D5: descarte de partes esqueletarias producto de C3
Cada evento de descarte es potencialmente informativo acerca de los comportamientos involucrados. Así, D nos informa acerca de la presencia de actividad de procesamiento inicial; D2acerca de los eventos de consumo inmediato al procesamiento de las partes seleccionadas. D3 es informativo respecto de la manufactura de charqui; D4 es indicador de consumo de charqui en el locus de manufactura del mismo. D5 es indicador del consumo de charqui luego de su transporte a otro asentamiento. Como nada es tan claro en el registro arqueológico, este modelo deberá monitorearse etnoarqueológicamente para determinar su funcionamiento antes de ser útil para su aplicación arqueológica. Pero pensamos que es un buen punto de partida.
Lo propuesto en este trabajo tiene como objetivo plantear algunos de los problemas que enfrentamos en la arqueología de sociedades pastoriles andinas. Esta contribución espera aportar una vía de investigación adecuada para enfrentar tales problemas. Recién estamos en el comienzo.
Agradecimientos:
queremos agradecer a Daniel Olivera por permitirnos tan gentilmente emplear sus datos de Montículo Chávez. También a todos los amigos que discutieron el tema con nosotros y quienes nos beneficiaron con sus ideas.
Bibliografía citada
Benavente, A.
1982 ChiuChiu. Una comunidad pastora temprana en la Provincia del Loa (II Región). Actas del XI Congreso Nacional de Arqueología Chilena, pp. 75-94. La Serena.
Binford, L.
1978 NunamiutEthnoarchaelogyAcademic Press, New York.
1980 Willow smoke and dog´s tails: hunter-gatherer settlement systems and archaeological site formation. American Antiquity 47: 4-20.
1984 Faunal Remains form Klasis River Mouth. Academic Press, New York.
Binford, L. y J. Bertram
1977 Bone frequencies end attritional processes. En For Theory Building in Archaeology, L.Binford (ed.), pp. 77-153. Academic Press, New York.
Borrero, L.
1986 Bone assemblages and butchering variability in Fuego-Patagonia: the case of guanaco.World Archaeology Congress, precirculated papers. Allen y Unwin, London.
Brain, C.
1981 The Hunters or the Hunted. University of Chicago Press, Chicago.
Browman, D.
1986 Origins and development of Andean pastoralism: an overview of the last 6.000 years. World Archaeology Congress, precirculated papers. Allen y Unwin, London.
Chang, C. y H. Koster
1986 Beyond bones: toward an archaeology of pastoralism. Advances in Archaeological Method and Theory 9: 97-148.
Flores Ochoa, J.
1968 Los Pastores de Paratía. Instituto Indigenista Americano, México.
Haber, A.
1992 Pastores y pasturas. Recursos forrajeros en Antofagasta de la Sierra (Catamarca), en relación a la ocupación formativa. Shincal2: 15-23.
Inamura, T.
1981 Adaptación ambiental de los pastores altoandinos en el Sur del Perú. En Estudios Etnográficos del Perú Meridional, S. Masuna (ed.), pp. 65-83. Universidad de Tokio, Tokio.
Kent, J.
1982 The Domestication and Exploitation of the South American Camelids: Methods of Analysis and their Application to Circumlacustrine Archaeology Sites in Bolivia and Peru. PhD dissertation. Washington University, St. Louis.
Matienzo, J.
1967 [1567] Gobierno del Perú. Travaux del Institut Francais d´ Etudes Andines, Tome XI. Paris-Lima.
Miller, G.
1979 An Introduction to the Ethnoarchaeology of the Andean Camelids. PhD dissertation. University of California, Berkeley.
Nuñez, L.
1981 Asentamiento de cazadores-recolectores tardíos en la Puna de Atacama: hacia el sedentarismo. Chungara8: 137-168.
Olivera, D.
1986 El Formativo de Antofagasta de la Sierra (Agroalfarero Temprano). Informe de avance de investigación. CONICET. Ms.
Olivera, D. y A. Nasti
1988 Procesos de formación de sitios en la Puna del Noroeste argentino: investigaciones tafonómicas sobre la preservación del registro arqueofaunístico. Trabajo presentado en el 46º Congreso Internacional de Americanistas. Amsterdam.
Rabey, M.
1986 Are llamas herders in the South Central Andes true pastoralists? En World Archaeological Congress,precirculated papers.Allen y Unwin, London.
Raffino, R.
1977 Las aldeas del Formativo inferior de la Quebrada del Toro. Obra del Centenario del Museo de La Plata II: 253-299.
Robertshaw, P. y D. Collett
1983 The identification of pastoral peoples in the archaeological record: an example from East Africa. World Archaeology 15: 67-78.
Thomas, D. y D. Mayer
1983 Behavioral faunal analysis of selected horizons. En The Archaeology of Monitor Valley. 2. GatecliffRockshelter, D. Thomas (ed.), pp. 353-391. Anthropological Papers nº 59. American Museum of Natural History, New York
Voigt, E.
1986 Iron Age herding: archaeological and ethnoarchaeological approaches to pastoral problems. South African Archaeological Society Goodwin Series 5: 13-21.

