











 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por email Citado por SciELO
Citado por SciELO  Similares em
SciELO
Similares em
SciELO 
 Permalink
PermalinkIntroducción
Los sitios arqueológicos del valle del río Negro presentan gran cantidad y diversidad de materiales faunísticos. Los estudios realizados han puesto atención en el conocimiento de las especies explotadas por las poblaciones prehispánicas (Mange et al. 2013, 2016; Prates 2008), en las condiciones tafonómicas que incidieron en la formación del registro faunístico (Fernández et al. 2011a; Prates 2008) y en la significancia paleoambiental a partir del registro de micromamíferos (Fernández et al. 2011a). Las investigaciones sistemáticas se realizaron sobre conjuntos recuperados en el valle medio-inferior de la cuenca, principalmente entre Gral. Conesa y Negro Muerto (64°30´ a 65° 30´ O; Figura 1). A partir del análisis de materiales faunísticos de este sector se propuso la existencia de estrategias de caza de tipo generalista entre las poblaciones humanas del Holoceno tardío (Prates 2008: 264-273). De acuerdo a esta propuesta, los cazadores-recolectores aprovecharon especies fluvio-lacustres (roedores anfibios, aves, peces y moluscos) en las cercanías de los ambientes ribereños donde se ubicaban sus campamentos residenciales, vertebrados pequeños y medianos y huevos de ñandúes en el interior del valle, y vertebrados grandes principalmente en las mesetas adyacentes al valle.
En este trabajo se presentan los resultados del análisis de los restos faunísticos recuperados en el sitio arqueológico Pomona, con el objetivo de discutir el aprovechamiento de recursos animales en el sitio y en el valle del río Negro durante el Holoceno tardío. Este sitio se ubica en cercanías de la isla Grande de Choele Choel, en el valle medio del río Negro (39º 31` 30” S y 65º 34` 16” O; Figura 1). Se encuentra emplazado sobre un médano, junto a la orilla derecha de un paleocauce que presenta, en este sector, una laguna de aproximadamente 80 m de largo por 20 m de ancho, seca la mayor parte del año. Los terrenos próximos al cauce y al sitio arqueológico están cubiertos por una formación arbustiva poco densa, con especies típicas de la provincia fitogeográfica del Monte, dominada por Larrea spp., asociadas a Schinus johnstonii, Condalia microphylla, Prosopis alpataco y otras (León et al. 1998).
Tres fechados radiocarbónicos ubican la ocupación humana en el Holoceno tardío final: 1011 ± 38 años 14C AP (AA93602, tibia de Lama guanicoe, nivel 5; 1013-1160 cal d.C.), 1070 ± 43 años 14C AP (AA94709, fémur de L. guanicoe, superficie; 962-1054 cal d.C.) y 983 ± 36 años 14C AP (AA90955, molar de Homo sapiens, colección; 1026-1177 cal d.C.). Los fechados podrían coincidir con una única ocupación, aunque la presencia de restos de ovejas (Ovis aries) y de un raspador de vidrio recuperados a poca profundidad (0-10 cm) permiten afirmar que hubo una segunda ocupación indígena, posiblemente durante el siglo XIX. La talla de vidrio ha sido asignada a los siglos XIX y XX (Casamiquela 1978) y las ovejas se introdujeron en Norpatagonia durante el siglo XVII (Enrique 2015).
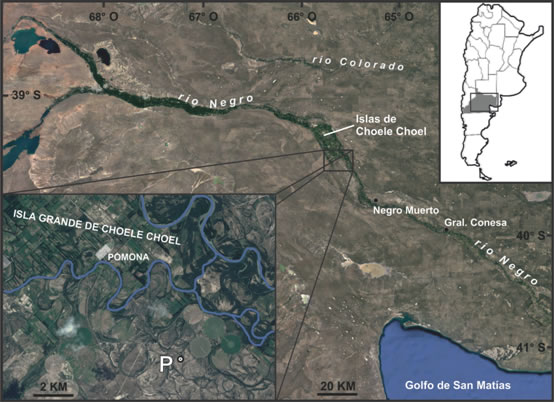
Figura 1: Ubicación del sitio arqueológico Pomona (P) en cercanías de la localidad homónima, y otras localidades mencionadas en el texto.
En Pomona, además de los materiales faunísticos, se hallaron diversos materiales que indican que el sitio habría sido un campamento residencial. Este conjunto incluye numerosos artefactos líticos tallados (entre ellos, puntas de proyectil y raspadores que podrían asociarse con la captura y el procesamiento de recursos faunísticos) y modificados por picado, abrasión y uso. También se registraron fragmentos de cerámica, leños y endocarpos carbonizados, cuentas sobre diversos soportes, pigmentos y sedimentos termoalterados que evidencian el encendido de fuegos (Mange 2019). Coleccionistas locales extrajeron, previamente, restos de al menos un esqueleto humano en el sector más alto del médano. En este trabajo se pondrá énfasis en los restos óseos faunísticos, mientras que otros materiales (exoesqueletos de moluscos y cáscaras de huevos de aves) solo se presentan en forma general. Se evalúa el aprovechamiento de especies y también los posibles sesgos causados por distintos agentes y procesos tafonómicos que actuaron sobre los materiales.
Materiales y métodos
Los trabajos de campo se realizaron en el año 2009. Se excavaron cuatro cuadrículas de 1 m2 en el sector oeste del médano, donde se observaron diversos materiales superficiales. Se siguieron niveles artificiales de cinco centímetros hasta alcanzar una profundidad de 50 cm. El sedimento consiste en arenas eólicas finas, bien seleccionadas, que no presentan bioindicadores hídricos; es decir que el sitio no estuvo expuesto a eventos de desborde por crecidas del río (Mange 2019). Los restos arqueológicos se vuelven menos frecuentes a partir del nivel 8 (35-40 cm) y desaparecen hacia los 45-50 cm de profundidad. Todo el sedimento se tamizó en zarandas metálicas con aberturas de 2,5 mm de diámetro.
La muestra analizada (n = 4059) incluye restos óseos (n = 3057), cáscaras de huevos de aves (n = 423) y exoesqueletos de moluscos fluviales (n = 579). Se excluye aquí el análisis de los elementos perforados elaborados sobre exoesqueletos de moluscos por no estar vinculados con la subsistencia (véase Mange 2019). En los restos de moluscos se cuantificaron aquellos exoesqueletos enteros o semi-enteros y los fragmentos con porciones no repetitivas (charnela o ápice; Prates y Marsans 2007). La determinación de las especies se realizó en base a bibliografía (e.g. Parada et al. 1989) y a muestras comparativas recolectadas en la zona de estudio. Por otro lado, la determinación taxonómica de las cáscaras de huevo de rheidos se realizó siguiendo metodologías propuestas previamente (Apolinaire y Turnes 2010) con modificaciones menores (e.g. se utilizó carbón molido como agente de tinción). Las cáscaras de especies medianas se identificaron mediante el uso de bibliografía (de la Peña 2016) y comparación con ejemplares provenientes del valle del río Negro. Tanto en las cáscaras de huevo como en los exoesqueletos de moluscos se evaluó la presencia de termoalteración, disolución y huellas antrópicas.
Para la identificación anatómica y taxonómica de los restos óseos se utilizaron muestras de la colección de Mastozoología del Museo de La Plata (FCNyM-UNLP) y atlas osteológicos de referencia (Bochenski y Campbell 2006; Fernández et al. 2011b; Sierpe 2015; entre otros). Los restos de mamíferos y de aves que no se pudieron determinar a nivel específico se separaron en categorías generales según su tamaño y peso corporal: micromamíferos (menores a 1 kg), mamíferos medianos (1-20 kg) y mamíferos grandes (mayores a 20 kg); aves pequeñas (menores a 200 g), aves medianas (mayores a 200 g) y aves grandes (i.e. Rhea sp.). Las especies de micromamíferos se agruparon a su vez en categorías de tamaño: pequeños (menores a 50 g), medianos (50-150 g) y grandes (150-1000 g).
Se realizó la cuantificación de los restos óseos mediante el empleo de medidas de abundancia taxonómica y anatómica: número de especímenes identificados por taxón (NISP), número de especímenes no identificados (NUSP), número mínimo de individuos (NMI), número mínimo de elementos (NME) y unidades anatómicas mínimas (MAU y MAU%) (Binford 1984; Lyman 2008; entre otros). Para conocer que agentes intervinieron en la formación del conjunto óseo y la preservación del mismo se relevaron los tipos de fracturas (i.e. estado fresco o seco) y los siguientes efectos tafonómicos: huellas de corte, raspado y machacado, termoalteración, marcas de raíces y de pisoteo, acción de carnívoros y de roedores, y depositaciones químicas (Binford 1981; Blumenschine et al. 1996; Gutiérrez 2004; Lyman 2008). Los valores de MAU% de Lama guanicoe fueron correlacionados con el Índice de utilidad de carne (Borrero 1990), con el objetivo de determinar si el ingreso está relacionado con el rendimiento de cada elemento. El grado de meteorización de los huesos se evaluó siguiendo la metodología de Behrensmeyer (1978) para el caso de los mamíferos medianos/grandes y la propuesta de Andrews (1990) para micromamíferos.
En la muestra de micromamíferos se evaluó el grado de corrosión en los dientes y en las epífisis de húmeros y fémures siguiendo la metodología propuesta por Andrews (1990) y modificada ulteriormente por Fernández et al. (2017a). Además, se evaluaron variables tafonómicas relacionadas con el aprovechamiento humano como huellas de corte, patrones de quemado e índices de Tamaño [IT= (NMI pequeños + NMI medianos / NMI grandes) x 100] y Predictibilidad [IP= (NMI gregarios + NMI coloniales + NMI agrupados) / (NMI crípticos + NMI solitarios) x 100] (Fernández et al. 2017b; Medina et al. 2012; Pardiñas 1999). Valores debajo de 100 en el índice IT indicarían actividad humana, en tanto, para el índice IP la tendencia es contraria (Pardiñas 1999).
Para realizar una comparación de arqueofaunas a nivel regional, se calcularon el índice de Artiodáctilos y el NTaxa de Pomona y otros sitios arqueológicos cercanos. Mediante el NTaxa (Lyman 2008) se evalúa la riqueza taxonómica de especies explotadas; para esto se sumaron los taxones de vertebrados con evidencias de explotación antrópica, evitando solapar categorías taxonómicas para no sobreestimar la muestra. El índice de Artiodáctilos (Broughton 1994) se utilizó para determinar el aporte relativo de los artiodáctilos a la dieta. Se calculó mediante la división de la suma del NISP total de artiodáctilos nativos por el NISP total de taxones con evidencias de explotación antrópica (incluyendo nuevamente artiodáctilos). Dentro del NISP total de taxones explotados no se incluyeron las placas dérmicas y los restos óseos de sigmodontinos (exceptuando a Holochilus vulpinus).
Resultados
1. Abundancia taxonómica y anatómica
De la muestra de especímenes óseos el 56,1 % se determinó al menos hasta nivel de clase (NISP=1713; Tabla 1), mientras que, debido principalmente a la elevada fragmentación, el 43, % de los restos no fue determinado (NUSP=1344). La muestra identificada incluye elementos de al menos 23 taxones, principalmente de mamíferos (94,3 %) y, en menor medida, de peces (2,6 %), aves (2,6 %) y reptiles (0,5 %). Muchos de ellos presentan evidencias de aprovechamiento antrópico (NTaxa= 18).
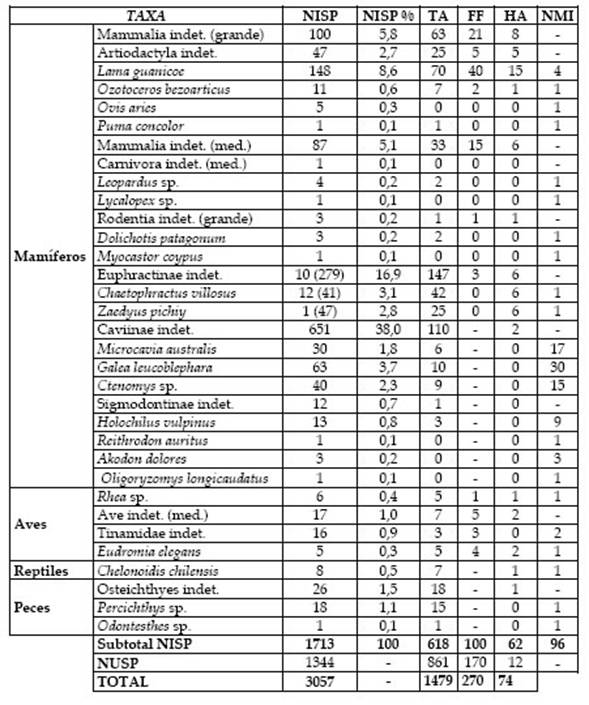
Tabla 1: Estructura taxonómica de los restos óseos (entre paréntesis se muestra la cantidad de placas del esqueleto externo de eufractinos). Se indican los especímenes termoalterados (TA), con fracturas en estado fresco (FF) y con huellas antrópicas (HA).
Entre los restos óseos de mamíferos (NISP = 1616) predominan los de micromamíferos (50,4 %), en menor medida de especies medianas (30,3 %) y grandes (19,3 %). Todos los especímenes de micromamíferos (NISP = 814; NMI = 76) pertenecen a roedores e incluyen restos postcraneales (n = 578; 71 %) y cráneo-mandibulares (n = 236; 29 %). Los elementos mejor representados son húmeros (NME=80), fémures (NME = 73), mandíbulas (NME = 73), tibias (NME = 64) y hemipelvis (NME = 64). Se determinó que el 93 % de los especímenes corresponde a especies de tamaño grande, principalmente a los caviomorfos Caviinae indet. (cuises), Galea leucoblephara (cuis moro), Microcavia australis (cuis chico) y Ctenomys sp. (tuco-tuco); y, en menor medida, al sigmodontino Holochilus vulpinus (rata nutria). Se identificaron dos morfotipos de Ctenomys, en base a la diferencia de tamaño; el mayor (NISP = 1) es notablemente más escaso que el menor (NISP = 39). Casos semejantes se observaron en la cuenca del río Limay (véase Fernández et al. 2016 y referencias allí citadas). Además, se registraron muy pocos elementos de otros sigmodontinos, algunos indeterminados y otros de especies medianas (Reithrodon auritus) y pequeñas (Akodon dolores y Oligoryzomys longicaudatus). Esta disparidad de valores a favor de los roedores grandes y sociales queda manifestada en los índices de tamaño (IT = 7) y de predictibilidad (IP = 442,9).
Los restos de mamíferos medianos (NISP = 490, NMI = 6) incluyen principalmente especímenes de armadillos (NISP = 390) e indeterminados (NISP = 87). Con menor representación, siguen roedores grandes (NISP = 7) y carnívoros (NISP = 6). El conjunto de especímenes de armadillos contiene elementos del esqueleto externo (placas, n = 367) e interno (n = 23). Muchos de ellos únicamente se pudieron determinar a nivel supragenérico (Euphractinae indet.) y otros corresponden a las especies Chaetophractus villosus (peludo) y Zaedyus pichiy (piche). Los restos de roedores grandes pertenecen a Dolichotis patagonum (mara), Myocastor coypus (coipo) y tres elementos indeterminados que podrían ser de esas especies o de Lagostomus maximus (vizcacha), el otro roedor grande que habita en el área. Dentro de los carnívoros medianos se identificaron especímenes de Leopardus sp. (gatos silvestres) y Lycalopex sp. (zorros), que corresponden a elementos del basipodio (calcáneo, astrágalo y otros). Además, se registró un fragmento de ilion de carnívoro mediano indeterminado.
Los especímenes de mamíferos grandes (NISP = 312, NMI = 7) pertenecen principalmente a Lama guanicoe (guanaco), artiodáctilos indeterminados y, en menor medida, a Ozotoceros bezoarticus (venado de las pampas), Ovis aries (oveja) y Puma concolor (puma), aunque muchos restos no pudieron determinarse (Tabla 1). El índice de artiodáctilos indica que aproximadamente uno de cada cinco especímenes es de artiodáctilos (IA = 0,19). Los restos de guanaco (NISP=148, NME=66) corresponden al menos a 4 individuos: dos adultos, identificados de acuerdo al número de vértebras lumbares fusionadas (NME = 10); un juvenil (19-24 meses), reconocido en base a dos hemimandíbulas con el segundo molar en proceso de erupción; y un individuo menor a los seis meses de edad inferido por el hallazgo de un fragmento distal de ulna no fusionado (véase Kaufmann 2009: 90-91). La representación anatómica (Figura 2) muestra abundancia tanto de elementos del esqueleto axial (mandíbula y vértebras lumbares) como apendicular (pelvis, porciones distales de metapodios y astrágalo). La correlación entre el MAU% y el Índice de utilidad de carne (Rs = -0.02, p = 0,94) muestra que el ingreso de partes de la carcasa no está relacionado con su rendimiento. Los restos de venado de las pampas son más escasos e incluyen elementos del esqueleto axial (astas, atlas, fragmentos de vértebras cervicales y torácicas) y apendicular (falanges, epífisis proximales de húmero y de tibia, epífisis distal de metatarso y un elemento del tarso). Los especímenes de oveja son también exiguos y corresponden a cuatro elementos del basipodio (calcáneo, cuboides, cuneiforme y lunar) y a un fragmento de mandíbula, hallados a escasa profundidad (0-10 cm). El espécimen de puma corresponde a una falange.
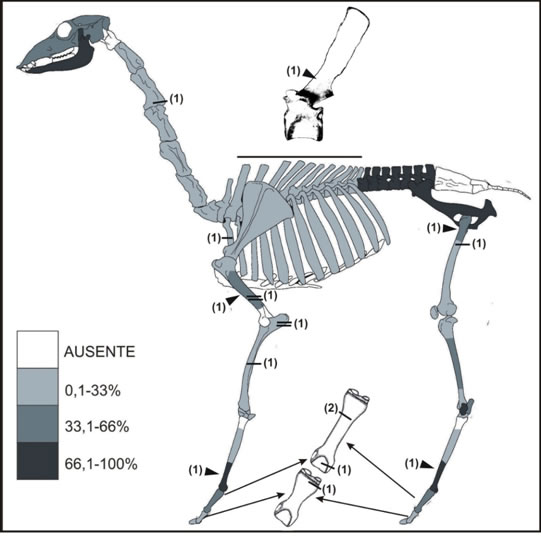
Figura 2 Representación esqueletaria (MAU%) de Lama guanicoe en Pomona y ubicación de las huellas antrópicas (línea= huella de corte; doble línea= raspado; triángulo= impacto/machacado). Entre paréntesis se indica el NME con huellas. Los restos de aves incluyen especímenes óseos (NISP = 44) y cáscaras de huevo (n = 23). Entre los primeros predominan los de especies medianas (86,4 %; Tabla 1), principalmente de tinámidos, mientras que son escasos los de ñandú (13,6 %). El conjunto de especímenes de tinámidos (Tinamidae indet. y Eudromia elegans) incluye solo elementos del esqueleto apendicular (NME = 20): coracoides (n = 8), tarsometatarso (n = 4), falanges (n = 3), carpometacarpo (n = 2), fémur, escápula y húmero (n = 1). Los restos indeterminados pertenecen a aves de porte mediano que podrían asignarse tentativamente a tinámidos, considerando el tamaño, la abundancia en este sitio y su recurrencia en conjuntos faunísticos del valle del río Negro (i Lorenzo et al. 2017; Prates y Acosta Hospitaleche 2010). Los especímenes de ñandúes (Rhea sp.) incluyen cuatro falanges, una carilla articular de vértebra y un fragmento de epífisis distal de tarsometatarso. Por otro lado, los fragmentos de cáscaras de huevo corresponden principalmente a rheidos (NISP = 420) y unas pocas a E. elegans (NISP = 3). Entre las primeras se determinó, en una muestra reducida (n = 38) debido a las condiciones de preservación y de visibilidad de las superficies, la presencia de las dos especies de Rhea que habitan actualmente en simpatría en el área de estudio: R. pennata (n = 17) y R. americana (n = 8).
Por último, se hallaron restos óseos de reptiles y de peces, y exoesqueletos de moluscos. Los especímenes de reptiles (NISP = 8) corresponden a Chelonoidis chilensis (tortuga terrestre), principalmente a fragmentos del espaldar (NISP = 7) y a un fragmento de cráneo. Los restos de peces (NISP = 45) incluyen principalmente vértebras con distinto grado de completitud y un fragmento de angular. Se determinó la presencia de Percichthys trucha (perca) y una vértebra de Odontesthes sp. (pejerrey), mientras que los especímenes restantes (n = 26) no pudieron identificarse debido a su fracturación. Por último, la muestra de moluscos incluye principalmente exoesqueletos de especies fluviales locales: la almeja Diplodon chilensis (NISP=505, NMI=264) y el caracol Chilina gibbosa (NISP = 74, NMI = 39).
2. Aspectos tafonómicos
El análisis muestra una elevada fracturación de los restos óseos (87 %), una tendencia que se mantiene en las distintas sub-muestras: micromamíferos (74,6 %), placas de eufractinos (65 %), aves (84 %), mamíferos medianos y grandes (85,6 %) y en la totalidad de los restos indeterminados (NUSP = 1344). En los exoesqueletos de moluscos fluviales son escasos los individuos enteros o semi-enteros (con más del 80% del exoesqueleto), tanto los de almejas (4,5 %) como los de caracoles (6,8 %). Las fracturas en estado fresco (Tabla 1) se presentan principalmente en restos óseos de mamíferos grandes (48,8 %), mamíferos medianos (45,5 %), guanacos (36,7 %), venados (33,3 %), artiodáctilos (16,1 %), rheidos (33,3 %) y aves medianas (44,4 %). En el caso de los guanacos la fracturación se registró en la totalidad de huesos largos de mayor tamaño (n = 35), incluyendo diez metapodios (Figura 3e), y también en primeras falanges (71,4 % de 14 especímenes) y en segundas falanges (85,7 % de siete especímenes).
Además de las fracturas frescas, en el conjunto de restos óseos se registraron otras evidencias de explotación: huellas antrópicas (4,4 %) y termoalteración (45,8 %). Los especímenes que no presentan evidencias antrópicas de ningún tipo son los de oveja, de zorro, de carnívoros indeterminados, de coipo y de roedores sigmodontinos de tamaño mediano y pequeño (con excepción de un espécimen de Sigmodontinae indet. con evidencia de quemado), aunque todos ellos están sub-representados (Tabla 1). Las huellas antrópicas (corte, raspado y machacado) se observaron en numerosos restos de mamíferos (n = 56) y también en especímenes de aves (n = 5), de reptiles (n = 1) y de peces (n = 1). Otros elementos que muestran modificaciones antrópicas no están relacionados con la subsistencia: un fragmento óseo de vertebrado grande con incisiones paralelas y cruzadas sobre la superficie cortical, un fragmento de cáscara de huevo de ñandú con una incisión y numerosos elementos perforados (n = 31) realizados sobre exoesqueletos de moluscos marinos y en un caso sobre valva de Diplodon chilensis (véase Mange 2019).
En los restos de aves las huellas de corte se ubican en el extremo distal de una primera falange de ñandú, en la porción proximal de la escápula y del coracoides de martineta (Figura 3c) y en una vértebra cervical de ave mediana. Se registró también un negativo de lascado en un hueso largo de ave mediana. En especímenes de tortuga las huellas de corte se encuentran en una placa del caparazón (Figura 3d) y en peces sobre un angular (mandíbula). En mamíferos se observaron huellas antrópicas en numerosos especímenes de artiodáctilos (n = 21) y de eufractinos (en un hueso largo y en la cara ventral de 17 placas de peludo y piche), y también en las diáfisis de ulna (Figura 3a) y de fémur de cuis, en el cráneo de un roedor grande, y en dos costillas, un hueso largo y tres elementos indeterminados de mamífero mediano (dos de estos casos son de raspado). En elementos de artiodáctilos se encontraron huellas de corte tanto en el esqueleto axial (sector medial de costilla y postcigoapófisis de vértebra cervical de guanaco, y dos mandíbulas de artiodáctilo) como en el apendicular, incluyendo elementos del miembro anterior (diáfisis medial de radioulna de guanaco y húmero de artiodáctilo) y posterior (diáfisis de fémur de guanaco). Además, se registraron en dos huesos largos de artiodáctilos, una falange de venado, y en una epífisis distal y dos diáfisis proximales de primera falange y en epífisis proximal de segunda falange de guanaco (Figura 2). Por otro lado, se relevaron negativos de impacto en diáfisis mediales de dos metapodios, un húmero y en apófisis espinosa de vértebra torácica de guanaco, raspado en diáfisis medial de húmero de guanaco y en diáfisis de hueso largo de artiodáctilo, y machacado en diáfisis distal de fémur de guanaco (véase Figura 2). Asimismo, se identificaron huellas antrópicas en elementos de mamíferos grandes no determinados: de corte sobre cuatro diáfisis de huesos largos, dos fragmentos de vértebra y en sector medial de costilla, y de raspado en dos diáfisis de huesos largos (uno de ellos con huella de corte).

Figura 3: Efectos tafonómicos relacionados con el aprovechamiento antrópico en Pomona: huellas de raspado en ulna de Caviinae indet. (a), quemado distal en incisivo de Galea leucoblephara (b) huellas de corte y quemado en coracoides de Eudromia elegans (c), huellas de corte en placa ósea de Chelonoidis chilensis (d) y carbonizado en metapodio de Lama guanicoe con fractura en estado fresco (e). Escalas= 5 mm.
La termoalteración se registró en especímenes óseos de diversos grupos: mamíferos (47,9 %), aves (45,4 %), reptiles (87,5 %) y peces (75,5 %). En mamíferos el quemado es importante en restos de especies medianas (37,9 %) y grandes (63 %), y supera el 50 % tomando a los artiodáctilos en su conjunto; también en placas de eufractinos (60 %). Los micromamíferos grandes presentan un porcentaje de quemado muy similar entre ellos, cercano al 20 %, aunque en algunos casos (e.g. ratas nutrias), los restos son escasos para hacer interpretaciones consistentes. En los tuco-tucos la termoalteración se presenta en elementos del esqueleto axial (ocho incisivos y una mandíbula) aunque no siguen patrones típicos de cocción (e.g. quemado distal de los incisivos). Entre los restos de cuises, los especímenes termoalterados son numerosos (n = 126; 16,9 %) y corresponden a casi todos los elementos óseos, incluyendo partes de cráneos y mandíbulas, vértebras y elementos de los miembros anteriores y posteriores. Se registró quemado distal en un maxilar, en seis incisivos (tres son de cuis moro; Figura 3) y en una tibia. El porcentaje de especímenes quemados de micromamíferos por nivel oscila entre el 7 % y el 28 % y no muestra una tendencia de cambio con la profundidad.
Los especímenes de puma, gato silvestre y mara presentan evidencia de quemado como única alteración que podría indicar acción antrópica. Además, es importante la alteración térmica en restos de ñandúes, aves medianas/tinámidos (incluyendo martinetas, Figura 3c), en los restos del esqueleto externo de tortuga y en vértebras de peces (véase Tabla 1). Considerando la totalidad de la muestra de especímenes termoalterados, los carbonizados alcanzan el 14,7 % y los calcinados el 8,1 %. También se hallaron numerosas cáscaras de huevo termoalteradas (41,2 %). En los moluscos fluviales la termoalteración afectó a escasas conchillas de C. gibbosa (4 %) y a casi la mitad de las valvas de D. chilensis (n = 129; 49 %). Además, se observaron algunas valvas de esta especie (n = 10) con disolución o pérdida de consistencia, pero sin evidencia de cambio de coloración por quemado (véase Mange et al. 2013).

Figura 4: Efectos tafonómicos post-depositacionales sobre restos óseos en Pomona: marcas de raíces en hemimandíbula de Lama guanicoe (a), depositación de carbonato en asta termoalterada de Ozotoceros bezoarticus (b) y meteorización (estadios 2-3) en vértebra cervical de L. guanicoe (c). Escalas= 10 mm.
No se observaron otros efectos (no antrópicos) relacionados con la depositación de restos, como actividad de carnívoros o corrosión digestiva producida por depredadores en la muestra de micromamíferos. Aun así, las marcas de raíces (Figura 4a) se registraron en el 31,2 % de la muestra (25 % en especímenes de micromamíferos). Las depositaciones de óxido de manganeso (1,1 %) y las marcas de roedores (0,18 %) tuvieron muy baja incidencia. La meteorización (Figura 4c) se observó en el 38,3 % de un conjunto de 188 especímenes de mamíferos medianos y grandes, con predominio del estadio 1 (20,1 %) y en menor medida los estadios 2 (11,7 %) y 3 (6,4 %). Esta evidencia junto con el blanqueamiento en el 3,6 % de esta muestra, indicaría cierta exposición superficial de al menos una parte de los materiales. Por otro lado, los restos de micromamíferos presentaron baja incidencia de meteorización (0,6 %), sugiriendo una incorporación más rápida a la matriz sedimentaria que el resto del conjunto. Finalmente, se observó que la depositación de carbonato (Figura 4b) es el agente tafonómico post-depositacional con mayor incidencia (67,7 %), lo cual podría haber ocultado efectos de otros agentes tafonómicos.
Discusión
El análisis faunístico de los materiales del sitio Pomona permite discutir algunas tendencias generales y rangos de movilidad vinculados con la captura y explotación de las presas. El conjunto se caracteriza por una riqueza taxonómica alta y por la presencia de especies que habitan distintas zonas ecológicas de la región (cauces de ríos, sectores ribereños, interior del valle, terrazas elevadas y mesetas), un patrón registrado en otros sitios del valle medio del río Negro (Mange et al. 2016; Prates 2008). Entre los restos óseos con evidencias claras relacionadas con el consumo (i.e. huellas antrópicas y termoalteración distal) se incluyen especímenes de peces, tortugas, martinetas, ñandúes, cuises, ratas nutrias, piches, peludos, grandes roedores, venados y guanacos. Además, deben agregarse huevos de ñandú y moluscos fluviales. El ingreso antrópico de almejas está sustentado por la asociación estratigráfica y horizontal con los materiales arqueológicos y por la elevada incidencia de alteración térmica; también es importante recordar que no habrían ocurrido inundaciones naturales sobre el sitio. Estos argumentos son válidos también para los restos de C. gibbosa, aunque su presencia es siempre menor a nivel regional, en comparación con las valvas de almejas (Mange 2019).
El consumo de peces se infirió principalmente por las evidencias de termoalteración. En otros sitios del valle del río Negro se ha mencionado la presencia de restos óseos de percas y bagres (Mange et al. 2013; Prates 2008); pero no se habían encontrado especímenes de pejerrey. Hasta el momento no se ha citado en la región el hallazgo de artefactos relacionados con tecnologías pesqueras; es probable que la captura de peces se realizara en las lagunas cercanas a los campamentos residenciales luego de que las crecidas estacionales del río las llenaran y dejaran peces atrapados allí (véase un caso similar para Norpatagonia en Martínez et al. 2005). Es importante resaltar que las evidencias de consumo de peces en la región inferidas a partir del registro arqueológico, contrastan con las observaciones realizadas por cronistas y viajeros en tiempos históricos (véase síntesis en Prates 2009).
Entre los micromamíferos predominaron ampliamente los roedores grandes y sociales (cuises, tuco-tucos y rata nutria), aspecto que se puede asociar con la explotación humana (e.g. Fernández et al. 2017b; Pardiñas 1999). Estas y otras especies de roedores son frecuentes en sitios arqueológicos cercanos, y las causas de su ingreso constituyen uno de los temas más discutidos en los estudios arqueofaunísticos del valle del río Negro (Fernández et al. 2011a; Mange et al. 2016; Prates 2008), así como también en otras regiones de Norpatagonia (e.g. Alcaraz 2012; Fernández et al. 2017b). El principal problema es discernir los aportes de muerte natural (causas eto-ecológicas, sensu Pardiñas 1999), predadores y descarte antrópico en la formación de los conjuntos óseos. Los restos de cuises de Pomona muestran quemado distal en incisivos y algunas huellas de corte largas y oblicuas al eje mayor del hueso, que estarían relacionadas con el descarne (Quintana 2005), y permiten afirmar su consumo. En el caso de los tuco-tucos no se han registrado evidencias consistentes que sostengan su consumo en Pomona y en otros sitios del valle del río Negro, a pesar de que es la especie más citada entre los micromamíferos consumidos en la Patagonia (ver síntesis en Fernández et al. 2017b). La rata nutria es una especie anfibia, hoy extinta en el área, que se habría capturado en cercanías de las orillas de cuerpos de agua, en este y otros sitios del área (Fernández et al. 2011b; Prates 2008). Por otro lado, los restos escasos de sigmodontinos pequeños, sin evidencias de explotación antrópica ni de acción de depredadores, podrían asignarse a muerte natural en los médanos (Andrews 1990; Pardiñas 1999). El pisoteo (avalado por la alta fragmentación) y la inestabilidad sedimentaria de los médanos habrían producido con el tiempo, la desarticulación y dispersión de los elementos óseos.
Los restos de mamíferos medianos con evidencias de consumo incluyen principalmente placas de eufractinos (Z. pichiy y C. villosus), que presentan quemado y huellas de corte en las caras internas probablemente relacionadas con la separación del escudo óseo. Son un recurso típico de la subsistencia prehispánica en la región (Mange et al. 2013, 2016). Por otro lado, son pocos los restos de roedores grandes y muestran escasa evidencia de aprovechamiento, principalmente quemado, de manera similar a la observada en otros conjuntos faunísticos de la cuenca del río Negro (Mange et al. 2016; Prates 2008). Por último, los restos de carnívoros presentan alteración térmica, aunque es más difícil probar su aprovechamiento antrópico: los especímenes son escasos y la mayoría corresponde a elementos del autopodio, lo cual podría indicar el uso de los cueros, quedando estos elementos adheridos al extremo de los mismos. La evidencia etnohistórica muestra el uso de pieles de estas especies (Prates 2009), aunque también se menciona el consumo de pumas (Musters [1869-70] 1997:73).
Otro de los recursos que podrían haber sido capturados en el interior del valle son las aves de porte mediano, principalmente tinámidos. La ubicación de las huellas antrópicas permite establecer como pauta de procesamiento de las carcasas la separación del miembro anterior, la porción anatómica con mayor rendimiento cárnico (Marani y Borella 2014), mediante la desarticulación de la cintura escapular. Los restos de martineta son comunes entre los conjuntos arqueofaunísticos del valle del río Negro (Di Lorenzo et al. 2017; Prates y Acosta Hospitaleche 2010). Es necesario considerar que la baja proporción de huellas de corte se deba a que el tamaño de las presas admite que pueda prescindirse de instrumentos líticos para su trozamiento y consumo (Blasco et al. 2014). Esto último también debe considerarse para micromamíferos (Fernández et al. 2017b) y peces (Stoessel 2015). Por otro lado, los restos de tortuga muestran el consumo de esta especie en el valle del río Negro, aunque suelen presentarse en escasa cantidad (Mange et al. 2016; Prates 2008).
Los especímenes óseos de ñandúes, aunque tienen evidencia de manipulación antrópica, son escasos, al igual que en otros conjuntos del valle medio del río Negro (Mange et al. 2016; Prates 2008). Esto contrasta con las numerosas menciones sobre su consumo en las fuentes escritas por viajeros y naturalistas (Prates 2009; Salemme y Frontini 2011, y referencias allí citadas). Los restos presentan termoalteración y se observaron huellas de corte en una primera falange que podrían indicar la extracción de los tendones de las extremidades para su uso como cuerdas (Prates 2009; Salemme y Frontini 2011). Las dos especies presentes en el este de Norpatagonia, R. americana y R. pennata, se han registrado en sitios arqueológicos del valle del río Negro a través de cáscaras de huevo (Apolinaire y Turnes 2010), pero en los registros óseos se determinó solo el hallazgo de R. americana (Mange et al. 2013; Prates 2008). Aunque los restos óseos de R. pennata podrían estar incluidos entre los no determinados (Rheidae indet.), podría ser también que la especie tuviera una distribución más austral y no habitara la zona del centro-norte de la provincia de Río Negro. En este caso la aparición de cáscaras de huevo de esta especie podría estar relacionada con el uso y traslado de huevos como contenedores, tal vez para transportar agua en las travesías por las mesetas que unían los cursos de agua (Prates y Mange 2016). Esto es congruente con la aparición de cáscaras grabadas, considerando que se trataría de artefactos conservados sobre los que se imprimiría cierto contenido comunicacional o simbólico (véanse Carden y Martínez 2014; Fiore y Borella 2010).
Por último, los artiodáctilos muestran abundante evidencia de explotación en el sitio. Especialmente el guanaco habría sido una fuente importante de carne y grasa, considerando su abundancia en el conjunto y su tamaño (hasta 100-120 kg, De Nigris y Mengoni Goñalons 2004). Sin embargo, el bajo índice de artiodáctilos (0,19) indica una dieta no orientada a este recurso debido a la riqueza de especies explotadas. Valores similares se obtuvieron en sitios cercanos de acuerdo a los datos publicados por Prates (2008): Angostura 1 (0,18) y Negro Muerto (0,13) (véase Tabla 2). Otros sitios del valle presentaron valores más elevados: Loma de los Muertos (0,33) y Negro Muerto 3 (0,62), lo cual podría deberse a un sesgo de muestreo considerando que en ambos sitios se realizaron recolecciones superficiales, luego del paso de maquinaria de nivelación del terreno (Mange et al. 2013, 2016). Los restos de algunos taxones medianos y pequeños podrían haberse perdido y de esta manera aumenta la importancia relativa de los vertebrados grandes. En el valle medio del río Negro, los especímenes de venados aparecen siempre en menor cantidad que los de guanaco y corresponden a diversas partes anatómicas (Mange et al. 2013, 2016; Prates 2008), lo cual indicaría que las carcasas habrían sido ingresadas enteras. Se trata de una especie que habita los sectores de pastizales de los fondos de valle (Jackson 1987).
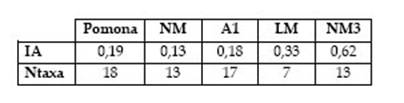
Tabla 2: Comparación entre sitios del valle medio del río Negro de acuerdo al Índice de artiodáctilos (IA) y la riqueza de especies explotadas (Ntaxa). Referencias: NM=Negro Muerto; A1=Angostura 1 (Prates 2008); LM=Loma de los Muertos (Mange et al. 2013); NM3=Negro Muerto 3 (Mange et al. 2016).
La tendencia regional indica el ingreso de pocos individuos de guanaco y un procesamiento intensivo, con fracturación de los huesos largos para acceder a la cavidad medular (Mange et al. 2016). En Pomona fueron ingresados individuos enteros y de acuerdo a las huellas antrópicas pueden inferirse diversas etapas de trozamiento (Binford 1981; Mengoni Goñalons 1999). Por ejemplo, las huellas de corte en postcigoapófisis de vértebras cervicales podrían indicar el seccionamiento del cuello durante el trozamiento primario y las huellas en radioulna proximal sugieren la desarticulación del codo durante el trozamiento secundario (Mengoni Goñalons 1999: 124-125). Las evidencias de trozamiento terciario o consumo incluyen huellas de corte y raspado en las diáfisis que indicarían descarne de los huesos largos y, los negativos de impacto en diáfisis mediales de metapodios y de húmero que señalarían el acceso a la cavidad medular para consumir la grasa interna. La totalidad de los huesos largos y la mayoría de las falanges se encuentran fracturadas, lo que indicaría cierta intensidad en el aprovechamiento de las grasas medulares (Wolverton 2002). La fracturación de los metapodios sugiere el consumo de grasas no saturadas, líquidas (Miotti y Marchionni 2014); además pudieron haberse utilizado para la fabricación de instrumentos (véase Mange et al. 2016).
Los guanacos pudieron cazarse tanto en la meseta como en el interior del valle (véanse discusiones en Mange et al. 2016; Prates 2008). Aunque probablemente vivían principalmente en las mesetas (Prates 2008: 266), bajarían ocasionalmente o estacionalmente en busca de agua en periodos de sequía, considerando la ausencia de fuentes permanentes fuera del valle (Prates y Mange 2016), momentos en los que podrían ser cazados. Esto explicaría los perfiles de representación anatómica presentes en el sitio, que muestran el ingreso de todas las partes anatómicas, situación que no es esperable cuando el área de captura y los campamentos base están distantes y las presas son de gran tamaño (Binford 1978; Mengoni Goñalons 1999; Perkins y Daly 1968). Además, explicaría la baja frecuencia de individuos de guanacos en Pomona y en otros sitios excavados en el valle del río Negro. El hallazgo de un individuo menor a seis meses en Pomona indicaría su captura en verano/otoño considerando la época de parición (Kaufmann 2009). Entre diciembre y febrero es el periodo del año con menos precipitaciones (Rodríguez y Muñoz 2006), lo cual podría haber causado que las manadas bajen al valle.
La muestra analizada se destaca por la abundante termoalteración y fragmentación. La primera podría deberse a la cocción, al uso de los elementos óseos como combustible o al descarte de los huesos en fogones. Por otra parte, también podría haber ocurrido la alteración no intencional en sucesivas re-ocupaciones de los mismos médanos; el encendido de nuevos fogones habría causado la termoalteración de los especímenes que se encontraban ya depositados en el sedimento circundante (quemado accidental sensu Costamagno et al. 2005). La fragmentación también podría haber sido causada por distintos procesos, y en muchos casos se observó que podría haber ocurrido durante el procesamiento antrópico, con los huesos en estado fresco. Luego de la depositación de los especímenes en el sedimento, la fragmentación podría haber ocurrido por pisoteo y por la dinámica propia de los médanos (Kocurek y Lancaster 1999). Debe tenerse en cuenta que la termoalteración causa fragmentación (Stiner et al. 1995), y que los médanos son espacios relativamente acotados que eran re-ocupados, y por lo tanto se espera en ellos una incidencia importante de pisoteo y alteración térmica sobre los especímenes faunísticos.
El análisis muestra que las depositaciones de carbonato, la meteorización y las marcas de raíces son las principales alteraciones tafonómicas que sufrieron los materiales luego del abandono del sitio. El carbonato podría haberse depositado sobre los distintos materiales del sitio mediante lixiviación producida por el agua de lluvia, considerando que los sedimentos loessoides del valle del río Negro poseen carbonato en su composición (Muhs et al. 2014: 431); también puede formarse a partir de precipitaciones biogénicas o incluso elevarse desde las napas (Imbellone et al. 2012). La meteorización habría ocurrido por la exposición ambiental luego del abandono del sitio o por re-exposición debida a movimientos de los sedimentos. Aunque es variable en cada ambiente particular y no se cuenta con estudios actualísticos cercanos, puede afirmarse que los elementos que presentan meteorización en estadios 2 y 3 podrían indicar exposición entre seis meses y un año (Borrero 2007; Massigoge et al. 2010).
Consideraciones finales
El conjunto faunístico analizado posee una buena integridad (sensu Binford 1981), considerando que las actividades de un grupo de cazadores recolectores hace aproximadamente 1000-800 años habrían sido el principal agente acumulador. Los especímenes constituyen la basura resultante del procesamiento y consumo de animales en bases residenciales, es decir, lugares donde un grupo de cazadores-recolectores se instaló y desde donde organizó distintas actividades relacionadas con la búsqueda de recursos. Algunas evidencias, como los restos de un individuo de guanaco menor a seis meses y las cáscaras de huevo de rheidos, sugieren que el sitio fue ocupado en verano, lo cual no invalida una ocupación continua a lo largo del año.
El sitio se localiza, como muchos otros de la región (Mange 2019; Prates et al. 2010), sobre un médano en cercanías de lagunas y cauces de agua, desde donde se explotarían diversos recursos disponibles en zonas acuáticas, ribereñas y en sectores bajos del valle. La presencia de restos de moluscos fluviales, peces y rata nutria indica la utilización de los cursos de agua; en el caso de los peces, la laguna cercana al sitio podría haber sido una trampa natural luego de las grandes crecidas de fines de otoño y primavera (Soldano 1947). Otros recursos de tamaño pequeño (principalmente roedores caviomorfos, eufractinos, tinámidos y otras aves medianas) podrían haber sido capturados principalmente en los sectores bajos del valle. Por último, las cacerías de grandes vertebrados (artiodáctilos y ñandúes) y quizás de carnívoros, pudieron realizarse en sectores alejados de los campamentos, tanto a lo largo del valle como fuera del mismo (Mange et al. 2016; Prates 2008: 264).
En síntesis, el registro faunístico de Pomona muestra una gran variedad de recursos explotados, abarcando mamíferos grandes, medianos y pequeños, reptiles, aves grandes y medianas, huevos de ñandúes y aves medianas, peces y moluscos. La amplitud de la dieta, vinculada con la disponibilidad de recursos que ofrece el valle, se relaciona también con los valores bajos del índice de artiodáctilos. Esta tendencia general se observa en otros sitios del valle del río Negro hacia el Holoceno tardío, aunque es contraria a otros sectores de Patagonia, donde el registro faunístico se concentra en pocas especies y entre ellas el guanaco tuvo un rol fundamental en la subsistencia (Mengoni Goñalons 1999; Miotti 2012). La ocupación humana que aportó los restos de ovejas habría ocurrido durante el S. XIX, cuando las sociedades de cazadores norpatagónicos habrían sufrido cambios importantes en su estructura social, relacionados con la incorporación del caballo y la movilidad, y con diversos recursos de uso cotidiano y de consumo (Nacuzzi 1998).
Desde su introducción en el siglo XVII (Cabrera 2000 [1620-1621], en Fernández 2006) el caballo habría facilitado la caza de grandes animales, como el ñandú, una especie veloz y de difícil captura, y también el guanaco (Musters [1869-1970] 1997:130-131; Salemme y Frontini 2011). Estos cambios explicarían dos situaciones regionales, aparentemente contradictorias, relacionadas con los recursos faunísticos: el aumento en tiempos post-hispánicos de la explotación del ñandú (véase discusión en Salemme y Frontini 2011) y una disminución del consumo de peces, que aparecen en el registro prehispánico regional (Mange 2019; Mange et al. 2016; Prates 2008), y son muy poco mencionados en las crónicas post-hispánicas (véase Prates 2009). En los últimos siglos las sociedades cazadoras-recolectoras habrían virado al aprovechamiento de animales de mayor retorno energético, tanto locales como introducidos. En el caso del sitio Pomona, aunque no puede precisarse si en la ocupación tardía del sitio se incorporaron otros materiales faunísticos al sitio, por el momento se ha propuesto que no: los especímenes de oveja se encuentran en posición sub-superficial y el resto de la muestra presenta en general efectos tafonómicos similares (Mange 2019).
Aunque el sitio se encuentra ubicado en un médano dentro de un valle fluvial, y por lo tanto es esperable cierta dinámica sedimentaria al menos a largo plazo (Waters 1992), la presencia de una cobertura vegetal (la actual y la del pasado según marcas de raíces) habría dado cierta estabilidad al depósito eólico. En este sentido, los estadios de meteorización bajos y la escasa incidencia de blanqueamiento indican buena preservación y condiciones de enterramiento. En concordancia, la elevada incidencia de depositación de carbonato de calcio en la muestra, sumada a la presencia de marcas de raíces en las superficies corticales de los huesos permiten inferir un enterramiento rápido de los materiales y cierta estabilidad a lo largo del tiempo, al menos para gran parte del conjunto.

