










Services on Demand
Journal

Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO
Related links
-
 Similars in
SciELO
Similars in
SciELO
Share

 Permalink
PermalinkAnuario de investigaciones
On-line version ISSN 1851-1686
Anu. investig. vol.20 no.2 Ciudad Autónoma de Buenos Aires Dec. 2013
PSICOLOGÍA EXPERIMENTAL
Correlatos electrofisiológicos del aprendizaje de relaciones de equivalencia y estructuras gramaticales: un estudio de potenciales cerebrales
Electrophysiological correlates of equivalence relations and grammar structure learning: a brain potentials study
Tabullo, Angel J.1; Yorio, Alberto2; Wainselboim, Alejandro3
1Licenciado en Psicología de la Universidad de Buenos Aires. Becario de investigación de CONICET, previamente en el laboratorio de Biología del comportamiento del IBYME, actualmente en la Unidad de lingüística del INCIHUSA - CCT Mendoza. Ex ayudante de trabajos prácticos de las cátedras de Neurofisiología y Métodos Psicofisológicos, en la Facultad de Psicología de la UBA. E-mail: angeltabullo@yahoo.com
2Doctor de la Universidad de Buenos Aires. Prof. Titular de Neurofisiología II. Director del Proyecto de Investigación UBACyT, Programación 2011 - 2014, "Teorías y modelos para la investigación en Neurociencias: registros conductuales y fisiológicos asociados a tareas de aprendizaje de categorías, relaciones de analogía y seguimiento de reglas gramaticales".
3Doctor en Biología de la Universidad de Buenos Aires. Investigador independiente de CONICET, previamente en el laboratorio de Biología del comportamiento del IBYME, actualmente en la Unidad de lingüística del INCIHUSA - CCT Mendoza. Ex docente titular de la práctica de investigación Métodos Psicofisiológicos en la Facultad de Psicología de la UBA. Ex JTP de la cátedra de Biología del comportamiento en la Facultad de Psicología de la UBA.
Resumen:
El objetivo del estudio fue comparar el rendimiento y potenciales cerebrales registrados durante el procesamiento de relaciones de equivalencia y gramáticas artificiales. El experimento se realizó en una muestra de adultos diestros sanos de entre 20 y 30 años. Se entrenó a los sujetos en dos tareas, una de aprendizaje de relaciones de equivalencia y otra de gramáticas artificiales, y se comparó el porcentaje de aciertos y los potenciales cerebrales registrados durante la fase de test de ambas. No se encontraron correlaciones en el desempeño en ambas tareas, pero sí negatividades similares en el rango de 400 a 600 ms, que fueron interpretadas como instancias del componente N400 (asociado al procesamiento lexical y semántico). Adicionalmente, se observó en la gramática artificial un componente P600 (vinculado al procesamiento sintáctico). Los resultados sugieren un solapamiento parcial en los mecanismos neurales del procesamiento de gramáticas artificiales y relaciones de equivalencia..
Palabras clave:
Relaciones de equivalencia; Gramática artificial; Aprendizaje; Potenciales cerebrales
Abstract:
The aim of the study was to compare performance and brain potentials recorded during processing of equivalence relations and artificial grammars. The experiment was condcuted on a sample of healthy right-handed adults whitin 20-30 years old. Subjects were trained in two tasks: equivalence relations learning and artificial grammar learning, percentage of correct responses and brain potentials obtained during the test stage were compared. No correlations were found in task performance, but a similar negativity was observed within the range of 400-600 ms. This negativities were interpreted as intances of the N400 component (associated with lexical and semantic processing). Additionally, a P600 component (associated with syntax processing) was observed in the artificial grammar. Results suggest a partial overlap between neural mechanisms of artificial grammar and equivalence relations.
Key words:
Equivalence relations; Artificial grammar; Learning; Brain Potentials
Introducción
El paradigma de relaciones de equivalencia (Sidman & Tailby, 1982) ha sido ampliamente utilizado en el campo del análisis experimental del comportamiento para el estudio de la formación de categorías y el comportamiento simbólico. Una relación de equivalencia se establece cuando, luego de entrenar una serie de asociaciones entre estímulos (típicamente por procedimientos de emparejamiento con la muestra o "matching to sample" - MTS), por ejemplo: A1-B1, B1-C1 y A2-B2, B2-C2, los sujetos son capaces de resolver relaciones nuevas, derivadas de las entrenadas, sin entrenamiento adicional y sin que estén basadas en características perceptuales de los estímulos (tales como forma, color, etc). Estas relaciones cumplen las propiedades de la equivalencia tal como se defne en matemática y lógica (reflexividad: si A1 => A1, simetría, si A1-B1 => B1-A1, transitividad: si A1-B1 y B1-C1 => A1-C1 y simetría-transitividad combinadas: si A1-B1 y B1-C1 => C1-A1). Una vez que se verifican estas propiedades, se considera que los estímulos constituyen una clase de equivalencia. Se ha propuesto que la capacidad de establecer relaciones de equivalencia sería la base para el procesamiento semántico y simbólico en el lenguaje natural (Sidman, 1986, 1984). La incapacidad de otras especies de formar relaciones de equivalencia entre estímulos, y el bajo rendimiento de sujetos con trastornos en la adquisición del lenguaje han sido citadas como evidencia a favor del vínculo entre el lenguaje y las relaciones de equivalencia (Dugdale & Lowe, 1990). Adicionalmente, otros autores han señalado la similitud entre diversos modelos teóricos en red de la memoria semántica (Anderson, 1976, 1983; Collins & Loftus, 1975; McCIelland & Rumelhart, 1988) y la estructura de las clases de equivalencia, que puede ser descrita como una red de estímulos relacionados entre sí a través de nodos (por ejemplo, en la clase A1-B1-C1, B1 sería el estímulo nodal que conecta a A1 con C1) (Fields & Verhave, 1987; Fields, Adams, Verhave, & Newman, 1990). Esta similitud establecida a nivel formal ha podido observarse también a nivel comportamental y neurobiológico mediante la aplicación del paradigma de priming semántico al estudio de las clases de equivalencia. En estos experimentos, se presentan pares de estímulos sucesivos, de manera tal que la aparición del primero, denominado "prime", afecta al procesamiento del segundo, denominado "target" al cual el sujeto debe responder. Cuando el estímulo prime está semánticamente relacionado con el target, (por ejemplo, el par "león" - "tigre"), el procesamiento de este último se ve facilitado, lo cual se traduce generalmente en una disminución de los tiempos de respuesta con respecto a ensayos donde el prime y el target no están relacionados ("camión" - "tigre"), o están indirectamente relacionados ("león" - "rayas", relacionados entre sí a través de la palabra "tigre"). A nivel neurobiológico, este fenómeno de facilitación o "priming" se ve reflejado en la amplitud de un potencial cerebral típicamente asociado al costo de procesamiento semántico, el N400 (Kutas & Hillard, 1980, véase Kutas & Federmeier, 2011, para una revisión). La magnitud de N400 disminuye en forma proporcional al grado de asociación entre el prime y el target, siendo máxima cuando éstos no están relacionados, e intermedia cuando están indirectamente relacionados (Kreher, Holcomb & Kuperberg, 2006). Esta misma modulación del N400 se ha observado al comparar pares de estímulos relacionados por entrenamiento, relacionados indirectamente por equivalencia y no relacionados, luego de haberse entrenado a los sujetos en una tarea de aprendizaje de relaciones de equivalencia (Barnes-Holmes, Staunton, Whelan, Barnes-Holmes, Commins, Wal-sh, Stewart, Smeets y Dymond, 2005). Estos resultados han sido interpretados como evidencia en favor de la existencia de mecanismos neurales y funcionales comunes al procesamiento de relaciones semánticas en el lenguaje y las clases de equivalencia.
Además del vínculo con aspectos semánticos del lenguaje, se ha propuesto que las relaciones de equivalencia podrían estar vinculadas a la adquisición de relaciones sintácticas (es decir, las posibles combinaciones estructurales de las diferentes categorías de palabras que hacen posible la construcción de oraciones en el lenguaje). Hasta el momento, la evidencia disponible consiste en la transferencia de funciones sintácticas simples entre los miembros de una clase de equivalencia: luego de entrenar a los sujetos para construir secuencias con estímulos que pertenecen a una clase de equivalencia, estos estímulos pueden ser sustituidos en la secuencia por otros miembros de la misma clase sin entrenamiento adicional (Chase, Ellenwood & Gregory Madden, 2008, Ellenwood & Chase, 1990; Wulfert & Hayes, 1988). Una posibilidad que aún no se ha evaluado es la posible relación entre la formación de relaciones de equivalencia y la adquisición de regularidades estructurales en secuencias de estímulos, estudiada mediante gramáticas artificiales.
Las gramáticas artificiales consisten en un conjunto de reglas complejas (por ejemplo, una gramática de estados finitos) que se utilizan para construir secuencias de estímulos. Luego de ser expuestos a un número de secuencias gramaticales (es decir, construidas de acuerdo con las reglas de la gramática), los sujetos entrenados son capaces de discriminar entre secuencias nuevas construidas correctamente y otras que contienen violaciones (transiciones inválidas entre elementos), aunque generalmente carecen de un conocimiento explícito de las reglas de la gramática (Reber, 1967). La interpretación típica del fenómeno es que se trata de un aprendizaje implícito en el que se detectan y extraen regularidades estructurales en las secuencias de estímulos presentadas. El paradigma de gramáticas artificiales ha sido utilizado para estudiar la participación de mecanismos de aprendizaje estadístico en la adquisición de aspectos sintácticos del lenguaje (Gómez y Gerken, 2000; Saffran, 2003), aplicándose a un rango de fenómenos que van desde la segmentación de palabras y fonemas (Maye, Werker, & Gerken, 2002; Saffran, Aslin, & Newport, 1996) hasta la adquisición de estructura de frase y categorías sintácticas (Gómez & Gerken, 1999; Saffran, 2002; Thompson & Newport, 2007). La interpretación de que los mecanismos de aprendizaje estadístico implicados en las tareas de gramática artificial son relevantes para la gramática del lenguaje natural es apoyada por estudios de neuroimagen que sugieren un solapamiento de las regiones cerebrales implicadas en el procesamiento de ambas gramáticas, en particular a nivel del área de Broca (Bahlmann, Schubotz, & Friederici, 2008; Petersson, Forkstam, & Ingvar, 2004, Petersson, Folia, & Hagoort, 2012; Folia, Forkstam, Ingvar, Hagoort, & Petersson, 2011). Adicionalmente, un estudio de electrofisiología que comparó los potenciales cerebrales registrados durante el procesamiento de oraciones del lenguaje y secuencias generadas por una gramática artificial mostró que las violaciones sintácticas de ambas gramáticas generaron un componente P600, una positividad posterior tardía considerada el correlato neurofisiológico de procesos sintácticos que tienen lugar en la comprensión del lenguaje (Hagoort, Brown, & Groothusen, 1993; Osterhout & Holcomb, 1992).
Dado que se ha propuesto un potencial vínculo funcional entre las relaciones de equivalencia y aspectos semánticos y sintácticos del lenguaje, resulta de interés comparar los correlatos neurobiológicos de la adquisición de relaciones de equivalencia con los del aprendizaje de una gramática artificial, cuyo procesamiento implica mecanismos estadísticos relevantes para la adquisición y el procesamiento de la sintaxis del lenguaje. Por lo tanto, el presente trabajo tuvo los siguientes objetivos: 1) comparar el desempeño de uno grupo de sujetos adultos sanos en tareas de aprendizaje de relaciones de equivalencia y gramáticas artificiales, a fin de determinar posibles correlaciones entre ambas, 2) Registrar y comparar la actividad EEG de los sujetos durante el procesamiento de relaciones de equivalencia y gramáticas artificiales, analizando el potencial solapamiento de los potenciales cerebrales observados en ambas tareas. La presencia de correlaciones en el desempeño de los sujetos en ambos tipos de aprendizaje y/o de potenciales cerebrales de latencia, topografía y polaridad similares, lo que podría implicar la existencia de procesos neurocognitivos comunes.
Metodología
Participantes
La muestra estuvo constituida por 16 sujetos adultos diestros (7 de ellos mujeres) de entre 20 y 30 años de edad, sin antecedentes de enfermedades psiquiátricas o neurológías. Todos los sujetos eran estudiantes de la Práctica de Investigación "Métodos Psicofsiológicos" y participaron voluntariamente del experimento. El protocolo experimental fue aprobado por el comité de ética del Instituto de Biología y Medicina Experimental (IBYME), y se siguieron estrictamente las normas internacionales para la investigación en humanos.
Procedimiento
Los participantes realizaron en dos sesiones diferentes la tarea de relaciones de equivalencia y gramática artificial. Todas las tareas fueron realizadas en una pe de escritorio y programadas en la plataforma Python. El orden de realización de las tareas fue contrabalanceado entre los sujetos.
Tarea de relaciones de equivalencia: La tarea consistió en una fase de entrenamiento, en donde se entrenaron una serie de asociaciones entre estímulos por medio de procedimientos de emparejamiento con la muestra, y una fase de test. Para el entrenamiento se utilizó una estructura de tipo "muestra como nodo", en el cual el mismo estímulo de muestra es asociado en bloques diferentes a distintos estímulos de comparación. Los estímulos consistieron en pseudopalabras presentadas en forma escrita (estímulos A1 y A2: "perno", "laco"; B1 y B2: "bune", "kime"; C1 y C2: "nufa", "roña"). En un primer bloque de ensayos se entrenaron relaciones de tipo A1-B1 y A2-B2, y en un segundo bloque de ensayos se entrenaron A1-C1 y A2-C2. Cada uno de estos bloques tuvo una duración de 16 ensayos. En un tercer bloque, de 64 ensayos, se entrenaron en forma conjuntas las relaciones AB y AC. Cada ensayo consistió en la presentación de un estímulo de muestra en la parte superior de la pantalla y dos estímulos de comparación equidistantes a ambos lados de la parte inferior. Se le dio al sujeto la instrucción de elegir cuál de los estímulos de comparación correspondía al estimulo de muestra presionando una tecla, y se le explicó que recibiría feedback luego de su elección. Luego del entrenamiento los sujetos pasaron a la fase de test, en la cual se les explicó que se les presentarían pares de estímulos (de a un estímulo por vez) y ellos tendrían que responder presionando una tecla si estaban o no relacionados de acuerdo al entrenamiento que habían recibido, luego de la aparición del segundo estímulo. También se les explicó que durante esta etapa no se informarían aciertos ni errores. Se presentaron en total 160 pares de estímulos, 80 relacionados por equivalencia (ya sea por simetría - B1-A1, C2-A2 - o por simetría y transitividad combinadas: B1-C1, C2-B2) y 80 no relacionados (por ejemplo, B1-A2 ó C2-B1). Cada estímulo tuvo una duración de 250 ms, con un intervalo entre estímulos de 100 ms, con lo cual la asincronía de inicio de estímulos ("stimulus onset asynchrony" - SOA) fue de 350 ms. En esta fase de test se realizó el registro de actividad de EEG, sincronizado con la aparición del segundo estímulo del par (el estímulo "target", al cual los sujetos tenían que responder). La variable dependiente comportamental a lo largo de toda la tarea fue el porcentaje de aciertos.
Fig. 1 - Diagrama de la gramática artifcial de estados fnitos. Cada nodo representa un Ítem o categoría de estímulos y las líneas indican transiciones válidas entre ellos. Las secuencias gramaticales se generaron conectando los nodos de izquierda a derecha.
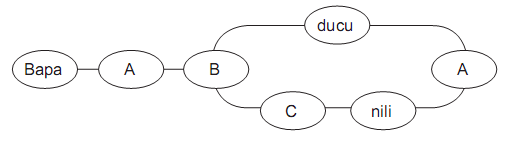
Category A: bañe, cañe, fadi, pefa, leda
Category B: siru, soti, nomi, revu, roli
Category C: pere, mene, tese, sele
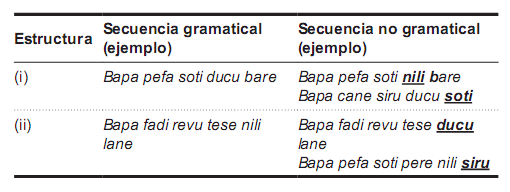
Tarea de gramática artificial: Se empleó una gramática de estados finitos con un léxico de 17 pseudopalabras (todas bisilábicas y fonotácticamente correctas), que admitía dos estructuras posibles (ver figura 1). En el entrenamiento, se les dio a los sujetos la instrucción de prestar atención a las secuencias y recordar las palabras artificiales que aparecían, ya que de tanto en tanto el programa les preguntaría si una palabra determinada apareció en la secuencia anterior. Se presentaron 140 secuencias construidas de acuerdo con las reglas de la gramática, de a un estímulo por vez, con una duración de 250 ms por estímulo y 200 ms de intervalo entre estímulos. Concluida la fase de entrenamiento, se le informó a los sujetos que las secuencias que acaban de observar eran oraciones de un lenguaje artificial, y que a continuación tenían que determinar si las nuevas oraciones que iban a ver habían sido construidas o no de acuerdo con las reglas gramaticales del lenguaje, presionando una tecla según el caso. De acuerdo con el protocolo típico de aprendizaje de gramáticas artificiales (Reber, 1967), se les explicó que en caso de no estar seguros de qué responder, lo hicieran de acuerdo a su intuición. En la fase de test se presentaron 140 secuencias nuevas, 70 construidas de acuerdo a las reglas de la gramática y 70 violaciones, en las cuales uno de los elementos fue presentado en una posición incorrecta (véase tabla 1). El formato de presentación de las secuencias fue idéntico al de la fase de entrenamiento. En ningún momento de esta tarea los sujetos recibieron retroalimentación. Durante la fase de test, se realizó el registro de la actividad EEG, sincronizado con la aparición del estímulo incorrecto en las secuencias que contenían violaciones y con los estímulos correspondientes en las secuencias gramaticales. La variable dependiente comportamental durante esta etapa fue el porcentaje de aciertos de los sujetos.
Tabla 1. Estímulos de la gramática artificial
Registro EEG
El registro de actividad EEG de los sujetos se llevó a cabo mediante un equipo AKONIC BIOPC de 19 canales, con referencia biauricular, digitalizando la señal a una tasa de muestre de 256 Hz. El análisis de los potenciales cerebrales se realizó offline mediante el software EEGLAB. El trazado se dividió en épocas de dos segundos de duración y 200 ms de línea de base, sincronizadas con la aparición de los estímulos críticos mencionados en cada una de las tareas.
Análisis estadístico
Para cada sujeto se calculó el porcentaje de aciertos por condición experimental, y en la tarea de gramática artificial se utilizó un test de T de una muestra para verificar que el desempeño del grupo fuera significativamente superior al azar (50%). Para determinar posibles asociaciones en el desempeño de ambas tareas se calculó el coeficiente r de Pearson entre los porcentajes de aciertos. Con respecto a los potenciales cerebrales, la variable dependiente fue el voltaje promedio dentro de una ventana de interés determinada a partir de la inspección visual del promedio general de los trazados, y se analizó mediante ANOVA de medidas repetidas. En los casos de violación del supuesto de esfericidad se aplicó la corrección Greenhouse-Geiser, y para las comparaciones pareadas se aplicó el ajuste de Bonferroni.
Resultados
Resultados comportamentales
En la tarea de relaciones de equivalencia, todos los participantes completaron exitosamente el entrenamiento (% Aciertos: 91,46 ± 7,11) y el test de relaciones derivadas (% Aciertos: 80,50 ± 11,89). En la tarea de gramática artificial, cuatro de los 16 sujetos tuvieron un desempeño inferior al 50 % en la fase de test, y fueron excluidos del análisis. Los 12 sujetos restantes tuvieron un desempeño significativamente superior al azar en la fase de test (% Aciertos: 61,75 ± 8,9; T(11) = 4,57, p = 0,001), demostrando adquisición de las regularidades estructurales de la gramática. No se encontraron correlaciones significativas en el desempeño de los sujetos en ambas tareas.
Análisis de EEG
En ambas tareas se calculó el promedio general de los potenciales cerebrales por sujeto y por condición experimental. Solamente se analizaron los datos de EEG de los sujetos que tuvieron un desempeño superior al azar en la fase de test de cada tarea, y sólo aquellos que superaron exitosamente ambas tareas (12) fueron incluidos en la comparación entre tareas.
Fig.2 - A la izquierda se observa el promedio general en un canal representativo (Pz) de los potenciales cerebrales en la tarea de relaciones de equivalencia (arriba) y gramática artificial (abajo). A la derecha se observa la topografía del potencial para las diferentes condiciones experimentales, en las correspondientes ventanas de interés.
Tarea de relaciones de equivalencia.
Los pares de estímulos no relacionados por equivalencia generaron un negatividad con distribución centroparietal entre los 400 y 600 ms posteriores a la aparición del target. (ver figura 2). El voltaje promedio en esta ventana de interés se analizó mediante un ANOVA de medidas repetidas 3 x 5 x 2
con los siguientes factores intrasujeto: Región del electrodo (anterior, central, posterior), Posición lateral del electrodo (numeradas del 1 al 5, de izquierda a derecha) y Tipo de estímulo (relacionado por equivalencia, no relacionado por equivalencia). Se encontró un efecto principal de Tipo de estímulo (F(1,15) = 5,154; p = 0.038).
Tarea de gramática artificial
Las secuencias que contenían violaciones a las reglas de la gramática generaron una negatividad ampliamente distribuida entre los 450 y 560 ms, y una positividad centroparietal entre los 630 y los 730 ms (ver figura 2). El voltaje promedio en cada ventana de interés se analizó con un ANOVA de medidas repetidas 3x5x2 con los siguientes factores intrasujeto: Región del electrodo (anterior, central, posterior), Posición lateral del electrodo (numeradas del 1 al 5, de izquierda a derecha), Tipo de estímulo (secuencia gramatical, secuencia no gramatical). En la primer ventana se encontró un efecto significativo de Tipo de estímulo (F(1,12) = 4,750; p = 0,047), mientras que en la segunda ventana se encontró una interacción Posición lateral * Tipo de estímulo significativa (F(4,44) = 3,412; p = 0,036). Las violaciones a las reglas de la gramática generaron una potencia significativamente más positivo que las secuencias gramaticales en los canales de la derecha (p < 0.022) y en los electrodos centrales, aunque en estos últimos el efecto no alcanzó el nivel de significación (p = 0.067).
Comparación de las negatividades observadas en las tareas de relaciones de equivalencia y gramática artificial.
A fin de comparar las negatividades observadas en ambas tareas dentro del rango de los 400 a los 600 ms, se realizó un ANOVA 3x5x2x2 con los siguientes factores intrasujeto: Región del electrodo (anterior, central, posterior), Posición lateral del electrodo (numeradas del 1 al 5, de izquierda a derecha), Tarea (relaciones de equivalencia, gramática artificial) Tipo de estímulo (estímulo relacionado - secuencia gramatical, estímulo no relacionado - secuencia no gramatical). Se encontró un efecto principal de tarea (F(1,1) = 7,143; p = 0,022) y una interacción Región* Tarea (F(2,22) = 20,308; p < 0,001). Los potenciales observados en la tarea de relaciones de equivalencia (para ambos tipos de estímulo) fueron más negativos que los de la tarea de gramática artificial en los canales centrales (p = 0.024) y parietales (p = 0.002). No se observaron interacciones signifcativas entre Tarea y Tipo de estímulo, por lo tanto, los efectos de la violación de una relación de equivalencia y la estructura de una gramática artifcial (comparados con los estímulos y secuencias correctas) no resultaron signifcativamente diferentes.
Discusión
El desempeño de los sujetos fue superior en la tarea de relaciones de equivalencia que en la de gramática artifcial, y la ausencia de correlaciones en el rendimiento de ambas tareas no apoya la hipótesis de que existan procesos cognitivos comunes entre ellas. Por otro lado, el análisis de los potenciales cerebrales señaló la presencia de un componente negativo de topografía y curso temporal similar en ambas tareas, que podría estar indicando un solapamiento parcial a nivel de los correlatos neurofsio-lógicos del procesamiento de relaciones de equivalencia y gramáticas artificiales. La interpretación de estos resultados y sus implicaciones se desarrolla a continuación.
Tarea de relaciones de equivalencia
La negatividad centroparietal observada entre los 400 y 600 ms puede considerarse de acuerdo a su latencia y distribución como una instancia del componente N400. Efectos de tipo N400 se han observado en otros estudios que compararon pares de estímulos relacionados por equivalencia y no relacionados (Barnes-Holmes et al., 2005; Haimson et al., 2009, Yorio et al., 2008), y han sido interpretados como evidencia a favor del vínculo entre las relaciones de equivalencia y el procesamiento semántico del lenguaje. El componente N400 se observa típicamente ante palabras inesperadas o semánticamente incongruentes en el contexto de oraciones, o ante pares de palabras no relacionadas, o indirectamente relacionadas en tareas de priming semántico (para una revisión, véase Kutas & Federmeier, 2011; Lau, Collins & Poeppel, 2008). Adicionalmente, se ha propuesto que la hipótesis del N400 como refejo de activación automática distribuida a través de los nodos de la memoria semántica en tareas de priming (Kreher, Holcomb & Kuperberg, 2006) podría aplicarse para explicar la diferencia entre los potenciales observados ante estímulos asociados directamente por entrenamiento e indirectamente por equivalencia (Barnes-Holmes et al., 2005). En síntesis, la presencia de un componente N400 ante estímulos que no están relacionados por equivalencia es congruente con la literatura previa, y apoya la hipótesis de mecanismos neurocognitivos comunes al procesamiento de relaciones de equivalencia y el lenguaje.
Tarea de gramática artificial:
Las violaciones estructurales en las secuencias de la gramática generaron, con respecto a las secuencias gramaticales, una negatividad más temprana y una positividad más tardía que pueden interpretarse como componentes N400 y P600 respectivamente. La presencia del potencial P600 es esperada, dado que un estudio previo que comparó el procesamiento de violaciones sintácticas en oraciones del lenguaje nativo y secuencias de una gramática artificial encontró efectos P600 análogos en ambos casos (Christiansen, Conway & Onnis, 2012). Adicionalmente, otros estudios observaron efectos P600 ante anomalías sintácticas en lenguajes artificiales (Friederici, Steinhauer & Pfeifer, 2002) y gramáticas artificiales desprovistas de contenido semántico (Bahlmann, Gunter & Friederici, 2006; Tabullo et a., 2011). El componente P600 es un correlato neural del procesamiento de la sintaxis del lenguaje, y se encuentra en una variedad de contextos que incluyen violaciones de estructura de frase y morfología, oraciones de tipo garden path y dependencias de larga distancia, entre otros (Hagoort, Brown, & Groothusen, 1993; Osterhout & Holcomb, 1992; para una descripción detallada véase Gouvea, Phillips, Kazanina and Poeppel, 2010). Se ha propuesto que es un reflejo del costo cognitivo de integración de una palabra con el contexto sintáctico previo (Kaan, Harris, Gibson and Holcomb, 2000), o bien el correlato de procesos de predicción e integración durante el procesamiento sintáctico (Hagoort 2003, 2009). En el contexto de nuestros resultados, el efecto P600 observado constituye evidencia adicional de que el procesamiento de secuencias estructuradas de estímulos generadas a partir de una gramática artificial involucra mecanismos neurales compartidos (al menos en parte) con el procesamiento de la gramática del lenguaje. Cabe destacar, sin embargo, que la aparente lateralización derecha que observamos en el análisis estadístico no ha sido reportada previamente en otros experimentos.
El componente N400, por otro lado, es menos esperable en experimentos con gramáticas artificiales desprovistas de contenido semántico. No obstante, en un estudio previo que utilizó una gramática artificial basada en la sintaxis del japonés, entrenada en sujetos sin conocimiento del idioma, violaciones de caso generaron un componente N400 seguido de un P600 (Mueller, Girsdies & Friederici, 2008). En otro estudio, violaciones de estructura de frase en una gramática artificial también generaron un componente similar al N400, y una positividad más tardía compatible con el P600 (Tabullo et al., 2011). La presencia del N400 en estos contextos ha sido interpretada como el producto de violaciones de expectativa a nivel lexical (Mueller, 2008), una explicación que se basa en evidencia previa de que el N400 también podría ser generado por procesos anteriores y más básicos que la integración léxico-semántica (Deacon, Dynowska, Ritter, & Grose-Fifer, 2004; Me Doyle, Russ & Wells, 1996), y en la modulación del componente N400 observada en tareas de aprendizaje de lenguajes artifciales, en ausencia de semántica (Sanders, Newport & Neville, 2002). Por lo tanto, es posible que las asociaciones probabilísticas entre los Ítems que componen las secuencias en nuestra gramática artifcial le hayan permitido a los sujetos generar predicciones sobre los elementos siguientes en la secuencia (por ejemplo, acerca de su morfología), y el incumplimiento de dichas predicciones en las violaciones sintácticas habría dado lugar a un componente N400.
Comparación entre la tarea de equivalencia y la tarea de gramática artificial
La comparación de las negatividades observadas en las tareas de gramática artificial y de relaciones de equivalencia indicó que, si bien la magnitud de los potenciales fue mayor en canales centrales y posteriores para la tarea de equivalencia, la magnitud del efecto N400 (la diferencia entre estímulos relacionados y no relacionados, y entre secuencias gramaticales y no gramaticales) no fue significativamente diferente entre ambas. Por otro lado, la duración del efecto fue mayor en el caso de las relaciones de equivalencia, abarcando la ventana completa de los 400 a 600 ms, mientras que en la gramática artificial sólo se observo de 450 a 560 ms. Estas diferencias podrían ser atribuidas a las diferencias de desempeño de los sujetos en cada una de las tareas, y asociarse al diferente nivel de aprendizaje de las relaciones de equivalencia (que involucran menos estímulos, y un entrenamiento guiado por realimentación) y las reglas estructurales de la gramática artificial (que son de naturaleza probabilística, e involucran un aprendizaje implícito por exposición pasiva a las secuencias gramaticales). A pesar de las diferencias, el análisis de los efectos sugiere un solapamiento al menos parcial entre los correlatos neurales del procesamiento de "violaciones" de una relación de equivalencia (presentación de estímulos que no están relacionados en forma directa ni derivada) y violaciones en la estructura de las gramáticas artificiales. Adicionalmente, los resultados podrían ser interpretados en ambos casos como el resultado de una violación de expectativa a nivel lexical sobre el próximo estímulo a ser percibido. En el caso de las relaciones de equivalencia, la presentación del prime permitiría preactivar el resto de los miembros de la clase de equivalencia que están relacionados directa o indirectamente con el estímulo, mientras que la aparición de un estímulo perteneciente a otra clase requeriría un costo adicional de procesamiento que se vería reflejada en la magnitud del N400. En el caso de la gramática artificial, las probabilidades de transición entre los elementos de las secuencias gramaticales permitirían preactivar los próximos elementos posibles en la secuencia, lo cual produciría un N400 cuando el elemento presentado no corresponde con las predicciones (es decir, en las violaciones gramaticales). Esta interpretación sería congruente con resultados previos que indican que el N400 podría ser el resultado de predicciones online realizadas por los oyentes durante la comprensión del lenguaje, basadas en asociaciones probabilísticas entre las palabras de una oración y sus continuaciones posibles (Delong, Urbach & Kutas, 2005).
En síntesis, los potenciales cerebrales observados en la tarea de equivalencia y de gramática artificial son congruentes con resultados reportados previamente, y señalan la posibilidad de un solapamiento parcial en el procesamiento de ambas. Este solapamiento podría ser el resultado de un proceso común de predicción, basado en un caso en la asociación indirecta entre los estímulos de la clase de equivalencia (conectados a través de estímulos nodales), y en el otro en la asociación probabilística de los elementos dentro de las secuencias de la gramática. Estos resultados constituyen evidencia a favor del uso de relaciones de equivalencia y gramáticas artificiales para el estudio controlado de procesos relacionados con el lenguaje. Asimismo, representan un antecedente para estudios futuros que indaguen los correlatos neurales de la transferencia de funciones sintácticas dentro de los miembros de una clase de equivalencia, utilizando la metodología de las gramáticas artificiales.
Referencias
1- Anderson, J.R. (1976). Language, memory, and thought. Hillsdale, NJ: Erlbaum. [ Links ]
2- Collins, A.M. & Loftus, E.F. (1975). A spreading activation theory of semantic processing. Psychological Review, 82, 407-428. [ Links ]
3- Bahlmann, J., Schubotz, R.l. & Friederici, A.D. (2008). Hierarchical artifcial grammar processing engages Broca's área. Neurolmage, 42, 525-534. [ Links ]
4- Barnes-Holmes, D., Staunton, C, Whelan, R., Barnes-Holmes, Y., Commins, S., Walsh, D., et al. (2005). Derived stimulus re-lations, semantic priming, and event-related potentials: testing a behavioral theory of semantic networks. Journal of the Experimental Analysis of Behavior, 84, 417-433. [ Links ]
5- Christiansen, M.H., Conway, C.M. & Onnis, L. (2012). Similar neu-ral correlates for language and sequential learning: evidence from event-related brain potentials. Language and Cognitive Processes. 27, 231-256. [ Links ]
6- Chase, P.N., Ellenwood, D.W., Madden, G. (2008). A behavior analytic analogue of learning to use synonyms, syntax, and parts of speech. The Analysis of Verbal Behavior. 24, 31 -54. [ Links ]
7- Deacon, D., Dynowska, A., Ritter, W. & Grose-Fifer, J. (2004). Re-petition and semantic priming of nonwords: implications for theories of N400 and word recognition. Psychophysiology, 41 (1), 60-74. [ Links ]
8- Doyle, M.C., Rugg, M.D., Wells, T. (1996). A comparison of the electrophysiological effects of formal and repetition priming. Psychophysiology 33 (2), 132-147. [ Links ]
9- Dugdale, N.A. & Lowe, C.F. (1990). Naming and stimulus equiva-lence. In D.E. Blackman & H. Lejeune (Eds.), Behaviour analy-sis in theory and practice (pp. 115-138). Hove: Lawrence Erlbaum Associates Ltd. [ Links ]
10- Ellenwood, D.W. & Chase, P.N. (1990). Combining semantic and syntactic classes through paired associate and matching to-sample procedures. Unpublished honor's thesis, West Virginia University, Morgantown. [ Links ]
11- DeLong, K.A., Urbach, T.P., Kutas, M. (2005). Probabilistic word pre-activation during language comprehension inferred from electrical brain activity. Nature Neuroscience, 8, 1117-21. [ Links ]
12- Folia, V., Uddén, J., de Vries, M., Forkstam, C, Petersson, K.M. (2010). Artifcial language learning in adults and children. Language Learning 60 (2), 188-220. [ Links ]
13- Fields, L, Adams, B.J., Verhave, T. & Newman, S. (1990). The effects of nodality on the formation of equivalence sets. Journal of the Experimental Analysis of Behavior, 53, 345-358. [ Links ]
14- Fields, L. & Verhave, T. (1987). The structure of equivalence classes. Journal of the Experimental Analysis of Behavior, 48, 317-332. [ Links ]
15- Friederici, A.D., Steinhauer, K. & Frisch, S. (1999). Lexical integraron: Sequential effects of syntactic and semantic informa-tion. Memory and Cognition, 27, 438-453. [ Links ]
16- Gómez, R.L. & Gerken, L.A. (2000). Infant artifcial language learning and language acquisition. Trends in Cognitive Sciences, 4, 178-186. [ Links ]
17- Gouvea, A., Phillips, C, Kazanina, N., Poeppel, D. (2010). The linguistic processes underlying the P600. Language and Cognitive Processes 25 (2), 149-188. [ Links ]
18- Hagoort, P. (2003). How the brain solves the binding problem for language: A neurocomputational model of syntactic processing. Neurolmage, 20, S18-S29. [ Links ]
19- Hagoort, P. (2009). Refections on the neurobiology of syntax. In D. Bickerton, & E. Szathmáry (Eds.), Biológical foundations and orígin of syntax (pp. 279-296). Cambridge, MA: MIT Press. [ Links ]
20- Hagoort, P., Brown, C. M. & Groothusen, J., (1993). The Syntactic Positive Shift (SPS) as an ERP-measure of syntactic processing. Language and Cognitive Processes 8, 439-483. [ Links ]
21- Kaan, E. & Harris, A. & Gibson, E. & Holcomb, PJ. (2000)The P600 as an Índex of syntactic integration diffculty. Language and Cognitive Processes, 15, 159-201. [ Links ]
22- Kreher D.A., Holcomb, P.J., Kuperberg, G.R. (2006) An electrophysiological investigation of indirect semantic priming. Psychophysiology. ; 43(6):550-63. [ Links ]
23- Kutas, M., Federmeier, K.D. (2011). Thirty years and counting: Finding meaning in the N400 component of the event related brain potential (ERP), Annual Review of Psychology, 62, 621-647 [ Links ]
24- Kutas, M. & Hillyard, S.A. (1980). Brain potentials during reading refect word expectancy and semantic association. Nature, 307, 1161-1163. [ Links ]
25- Lau, E., Phillips, C, Poeppel, D. (2008). A cortical network for semantics: (de)constructing the N400. Nature Reviews Neuroscience, 9, 920-933. [ Links ]
26- Maye, J., Werker, J. & Gerken, L.A. (2002). Infant sensitivity to distributional information can affect phonetic discrimination. Cognition, 82, 101-111. [ Links ]
27- McCIelland, J.L. & Rumelhart, D.E. (1988). Explorations in parallel distríbuted processing: A handbook of models, programs, and exercises. Cambridge, MA: MIT Press. [ Links ]
28- Mueller, J.L., Girgsdies, S., Friederici, A.D. (2008). The impact of semantic-free second- language training on ERPs during case processing. Neuroscience Letters, 443(2):77-81. [ Links ]
29- Osterhout, L., Holcomb, PJ. (1992). Event-related brain potentials elicited by syntactic anomaly. Journal of Memory and Language 31 (6), 785-806. [ Links ]
30- Petersson, K.M., Forkstam, C. & Ingvar, M. (2004). Artifcial syntactic violations actívate Broca's región. Cognitive Science, 28, 383-407
31- Petersson, K.M., Folia, V & Hagoort, P. (2012). What artifcial grammar learning reveáis about the neurobiology of syntax. Brain and Language, 120 (2), 83-95. [ Links ]
32- Reber, A.S. (1967). Implicit learning of artificial grammars. Journal of Verbal Learning and Behavior, 6, 855-863. [ Links ]
33- Saffran, J.R., Aslin, R.N. & Newport, E.L. (1996). Statistical learning by 8-month-old infants. Science, 274, 1926-1928. [ Links ]
34- Saffran, J.R. (2002). Constraints on statistical language learning. Journal of Memory and Language, 47, 172-196. [ Links ]
35- Saffran, J.R. (2003). Statistical learning: Mechanisms and constraints. Current Directions in Psychological Science, 12, 110-114. [ Links ]
36- Sanders, L.D., Newport, E.L., Neville, H.J. (2002). Segmenting nonsense: an event-related potential Índex of perceived on-sets in continuous speech. Nature Neuroscience, 5, 700-703. [ Links ]
37- Sidman, M. (1986). Functional analysis of emergent verbal classes. In T. Thompson & M.D. Zeiler (Eds.), Analysis and integration of behavioral units (pp. 213-245). Hillsdale, NJ: Erlbaum. [ Links ]
38- Sidman, M. (1994). Stimulus equivalence: A research story. Boston: Authors Cooperative. [ Links ]
39- Sidman, M. y Tailby, W. (1982). Conditional discrimination vs. matching to sample. An expansión of the testing paradigm. Journal of the Experimental Analysis of Behavior, 37, 5-22. [ Links ]
40- Tabullo, A., Sevilla, Y., Pasqualetti, G., Vernis, S., Segura, E., Yo-rio, A., Zanutto, B.S. & Wainselboim, A.J. (2011). Expectancy modulates a Late Positive ERP in an artifcial grammar task". Brain Research, 1373, 131-143. [ Links ]
41- Thompson, S. & Newport, E. (2007). Statistical learning of syntax: The role of transitional probability. Language Learning and De-velopment, 3, 1-42. [ Links ]
42- Wulfert, E. & Hayes, S.C. (1988). Transfer of conditional ordering response through conditional equivalence classes. Journal of the Experimental Analysis of Behavior, 50, 125-144. [ Links ]
Fecha de recepción: 20 de marzo de 2013
Fecha de aceptación: 17 de octubre de 2013
