










Servicios Personalizados
Revista

Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkCiencia, docencia y tecnología
versión On-line ISSN 1851-1716
Cienc. docencia tecnol. n.39 Concepción del Uruguay nov. 2009
CIENCIAS EXACTAS Y NATURALES - COMUNICACIÓN
Resistencia a herbicidas. Glifosato *
Resistance to Herbicides. Glyphosate *
Villalba, Andrea **
*) Artículo producido en el marco del Proyecto de Beca Doctoral "Efectos de glifosato y metsulfurón metil en plantas acuáticas y uso posible uso como bioindicadoras". Universidad Nacional del Litoral (UNL), 2008; recibido en marzo 2009, aceptado en setiembre 2009.
**) Área de Diversidad Vegetal, Departamento de Ciencias Naturales, Facultad de Humanidades y Ciencias, UNL, Santa Fe (Argentina). abvdesantafe@yahoo.com.ar
Resumen: Se presenta una revisión actualizada de los mecanismos generadores de resistencia a herbicidas, en plantas, y los factores que favorecen su desarrollo. Se reúnen las especies que actualmente han sido denunciadas como resistentes a glifosato, en Argentina y el mundo. Se explica el mecanismo de acción y de desarrollo de resistencia de este xenobiótico, poniendo énfasis en las conductas preventivas.
Palabras clave: Ciencias Agrarias; Resistencia a herbicidas; Glifosato
Abstract: This paper presents a revision of resistance to herbicides mechanisms and factors of those developments in plants. Glyphosate resistent plants in Argentina and the world are considered. Mechanisms of action and development of glyphosate resistance and preventive behaviors are proposed.
Key words: Agricultural sciences; Herbicide resistance; Glyphosate
l. Introducción
A nivel mundial, la agricultura moderna ha logrado incrementos productivos notables, acompañados de una creciente obtención de alimentos, impulsada por un amplio uso de insumos para el control de malezas, plagas y enfermedades, como así también fertilizantes. Los últimos 50 años del siglo XX y lo que va del siglo XXl se caracterizan por una fuerte dependencia respecto de los agroquímicos como elementos fundamentales en el control de adversidades de los cultivos. El desarrollo de diferentes familias de herbicidas a partir de la década de 1940 ha dado lugar a la idea de que el control químico es una herramienta que posibilita la erradicación de malezas, convirtiéndose, desde entonces, en el método principal utilizado por los productores agropecuarios (11). Según CASAFE (2), en 1991 se emplearon 39 millones de litros de plaguicidas. Durante 2001, se comercializaron unos 127 millones de litros, de los cuales los herbicidas constituyeron entre el 61 y 66 por ciento. Es evidente, entonces, que estos xenobióticos se han constituido en protagonistas de la agricultura y, en Argentina, las ventas más importantes son las de glifosato, el herbicida núcleo de la producción sojera (13).
Dentro de este contexto de control químico, el tema de la tolerancia y resistencia de malezas constituye una de las líneas de investigación a destacar en relación a este xenobiótico: el glifosato (16, 5).
Por resistencia debe entenderse la capacidad poblacional o de un biotipo que le permite a sus individuos sobrevivir y dejar descendencia ante la aplicación de una dosis letal de herbicida para la cual su filial original era susceptible. Una de las primeras consideraciones que deben realizarse es no confundir este fenómeno con el de tolerancia. La tolerancia es una habilidad natural propia de la especie que le permite reproducirse y sobrevivir, y viceversa, ante la aplicación de herbicidas. A diferencia de las tolerantes, las poblaciones resistentes suelen sobrevivir no solo a las dosis de aplicación agrícola del herbicida sino a otras bastante superiores (4).
En este trabajo se reconstruye parte de la teoría en torno al fenómeno de resistencia de los herbicidas en general, haciendo referencia especial a la resistencia a glifosato, entendiendo que de ningún modo se ven agotados en el mismo todos los aportes teóricos existentes sino que esta propuesta es un recorte de la bibliografía consultada.
ll. Resistencia a herbicidas
En términos generales, el desarrollo de resistencia a cualquier herbicida involucra un proceso de selección ligado al de variabilidad intraespecífica. Se asume que cualquier población de malezas puede tener biotipos resistentes en baja frecuencia, debido a mutaciones que ocurren naturalmente. Así, el uso repetido de un mismo herbicida expone a la población a una presión de selección que conduce al aumento del número de individuos resistentes. Los biotipos susceptibles mueren mientras que los resistentes sobreviven produciendo propágulos. Si persiste la aplicación de herbicidas que actúan sobre el mismo sitio de acción, la proporción del biotipo resistente se incrementa en relación al biotipo susceptible. Indiquemos también que además de la resistencia que puede ocurrir de forma natural, existen posibilidades de inducirla por técnicas de ADN recombinante propuestas por la ingeniería genética o selección de variantes resistentes obtenidas por cultivos de tejidos o mutagénesis (4).
Diferentes autores coinciden en afirmar que, naturalmente, existen tres mecanismos que provocan resistencia, a través de la eliminación de la acción fitotóxica del herbicida: la modificación del sitio de acción, la detoxificación por metabolización y la reducción de absorción, transporte, aislamiento o secuestro.
II. 1. Modificación del sitio de acción
O pérdida de afinidad por el sitio de acción. Salvo casos excepcionales, estos sitios (enzimas, generalmente) son específicos, y la acción del herbicida sobre los mismos (efecto primario) suele conducir al desarrollo de efectos secundarios, de naturaleza mucho más general, que normalmente acaban produciendo la muerte de la planta. Cualquier modificación en la estructura de la enzima (una o varias mutaciones en la secuencia de aminoácidos que componen la molécula) puede resultar en una pérdida de afinidad del herbicida por ésta, imposibilitando la unión efectiva de ambos e impidiendo así la inhibición del proceso vital mediado por dicho sitio. En la mayoría de los casos la razón de la resistencia es ésta.
II. 2. Detoxificación por metabolización
Según la cual, los biotipos o poblaciones resistentes degradan el herbicida a metabolitos no fitotóxicos. Los biotipos resistentes son capaces de degradar el herbicida antes que éste cause daños irreversibles. La velocidad de degradación enzimática puede variar por factores endógenos y exógenos, tales como el estadío de crecimiento de la planta y las condiciones meteorológicas.
Los procesos de detoxificación metabólica de herbicidas en tejidos vegetales pueden dividirse en tres fases: conversión, conjugación y deposición. Esta división no es una regla general dado que alguna de las fases puede faltar, la molécula del herbicida puede ser proherbicida inactivo que debe ser enzimáticamente convertido en compuesto activo o porque ciertos procesos de conjugación son de carácter reversible, por lo que afectan parcialmente la cantidad de herbicida libre intracelular.
Fase 1. Conversión: en el caso de los herbicidas que no tienen sustituyentes disponibles (grupos amonio, hidroxilo, etc.) para formar conjugados, deben ser convertidos en metabolitos mediante una reacción química.
Fase 2. Conjugación: los conjugados suelen ser los metabolitos finales en los procesos de detoxificación de herbicidas y su naturaleza puede ser diversa (azúcares, aminoácidos, péptidos y lignina como grupos orgánicos, y enlaces éster, éter, amida o glicosídico).
Fase 3. Deposición: los conjugados glicosídicos son depositados en la vacuola donde quedan almacenados, mientras que los de origen aminoacídico son excretados a la pared celular donde se integran en el componente de lignina de éstas, formando un residuo insoluble. Estos procesos no son irreversibles, pero su conversión a herbicida activo intracelular es muy lenta.
II. 3. Reducción de absorción, transporte, aislamiento o secuestro
Su objetivo es la reducción de herbicida en el sitio de acción. Una condición esencial es que el herbicida llegue al sitio de acción en una concentración suficiente como para que su efecto sea letal. La falta de movimiento del herbicida permitirá reducir la concentración de éste en el sitio de acción, lo que le posibilitará a este último mantenerse funcional. Estas bajas concentraciones pueden lograrse mediante reducción de la penetración, absorción o translocación, o por la existencia de fenómenos de secuestro a organelas metabólicamente inactivas.
a) Absorción del herbicida: En términos generales, el modo de absorción depende, además de las características físicas y químicas de las moléculas del herbicida, de la forma de aplicación del mismo: posemergente (penetración foliar) o preemergente (penetración radicular). La resistencia/sensibilidad a herbicidas con acción residual, en tratamientos de preemergencia, puede venir asociada a factores morfológicos tales como diferencias en la profundidad o estructura anatómica del sistema radicular, como a factores fisiológicos tales como la limitada absorción activa del herbicida en especies sensibles. Con respecto a la vía foliar, las condiciones meteorológicas, la tensión superficial de la solución del herbicida, el volumen de tratamiento y las características foliares tales como el área, la orientación foliar y la cantidad de ceras epicuticulares presentes en las hojas determinarán la cantidad de herbicida que queda pegado a la planta y penetra luego en los tejidos vegetales.
b) Translocación del herbicida: Este proceso también puede clasificarse según el tratamiento de aplicación del herbicida. En tratamientos vía radicular, el movimiento del herbicida va a depender de su naturaleza química, y esto se debe a tres cuestiones, fundamentalmente: la acumulación del herbicida no metabolizado en la raíz (lo que resulta en una falta de translocación de la materia activa a la parte aérea), la metabolización del herbicida en la raíz a una forma no translocable del mismo, generalmente conjugados polares, o la restricción del movimiento del herbicida al sistema vascular, lo que le impide alcanzar su sitio primario de acción en el tejido vascular. En el caso de la vía foliar, el movimiento del herbicida puede realizarse por floema o xilema. Mientras por el xilema el herbicida sigue el flujo de agua libremente hacia los márgenes foliares y los espacios intervasculares, el transporte por floema va a depender de dos procesos diferenciados: gradiente de concentración del herbicida entre las células floemáticas y las mesofílicas, y la capacidad del herbicida de ser retenido por las células floemáticas durante su transporte.
c) Secuestro del herbicida: El aislamiento o secuestro consiste en retirar el herbicida o sus metabolitos de las regiones metabólicamente activas y llevarlos a sitios menos activos, donde son inocuos para el crecimiento, como es el caso de las vacuolas (4, 23).
Si pensamos en los factores que influyen en la determinación de la tasa de selección y, por lo tanto, en la tasa de evolución de la resistencia, es decir la velocidad con que se desarrolla la resistencia, podemos afirmar que:
- A medida que aumenta la frecuencia de alelos resistentes, la tasa de evolución de resistencia es mayor.
- La herencia a través de alelos dominantes es más rápida, puesto que tanto los individuos homocigotos dominantes como heterocigotos resultan resistentes.
- La herencia de la resistencia es generalmente monogénica, pero cuando es poligénica la tasa de evolución es más lenta, pues requiere recombinaciones durante varias generaciones para producir un número de alelos que puedan dar origen a un genotipo resistente.
- En especies alógamas, los alelos de resistencia pueden dispersarse a través de las semillas y polen. En las especies autógamas, el flujo génico es reducido entre individuos, produciéndose la dispersión de alelos casi sólo por semillas.
- La producción de un elevado número de semillas favorece la dispersión de la resistencia.
- En poblaciones de alta densidad, la probabilidad de individuos resistentes es mayor.
- En cada estación de crecimiento sólo una parte de las semillas de un banco estará expuesta a la selección. En campañas sucesivas se irán introduciendo individuos susceptibles, lo que disminuirá la frecuencia de alelos resistentes.
- En caso de especies anemófilas, el viento puede dispersar genotipos resistentes a áreas no infectadas. Lo mismo pasa con animales y máquinas.
- Si las malezas tienen períodos largos de germinación, la probabilidad de que el herbicida afecte a una parte de la población se incrementa. Los individuos que emerjan después de la aplicación disminuirán la frecuencia de alelos resistentes.
- Si ante la aplicación de herbicidas los individuos heterocigotos tienen mayor fitness que los homocigotos susceptibles, la evolución de la resistencia será más rápida.
También hay factores que sólo se relacionan con las características del herbicida, e intervienen en la generación de resistencia:
- La eficiencia del herbicida nos indica la mortalidad que causa en una población de malezas. Los herbicidas más eficaces eliminan más individuos susceptibles y facilitan la evolución de resistencia.
- En el caso de los herbicidas que interfieren en un solo sitio de acción, el cambio en un solo gen puede ser suficiente para afectar la unión de la molécula herbicida al sitio de acción.
- Los herbicidas no residuales actúan sobre las malezas emergidas mientras que los residuales afectarán las cohortes siguientes.
- El empleo repetido de un mismo herbicida en la misma campaña o en campañas sucesivas favorece la evolución de poblaciones resistentes.
- Cuando la resistencia es monogénica, el empleo de dosis elevadas aumenta la presión de selección y favorece la evolución de resistencia.
- Si la resistencia es poligénica deberán ocurrir recombinaciones que generen un número suficiente de alelos que generen un genotipo resistente. El uso de subdosis permite que individuos que expresan mecanismos de resistencia débiles por no poseer la cantidad de alelos suficientes sobrevivan y contribuyan al pool de genes de resistencia. Al aplicar dosis altas, estos individuos son eliminados y disminuye la frecuencia de alelos resistentes en la población.
La Sociedad Española de Malherbología (19) propone un listado de factores que nos indican las opciones que provocan mayor o menor riesgo de inducir resistencia, que pueden observarse en el Cuadro 1. Obviamente, cuando mayor número de factores contribuyan negativamente, más inminente será la aparición de resistencia en malezas.
Cuadro 1: Factores e inducción de riesgo de resistencia

En función también del Cuadro 1, y a modo de árbol de ideas, pueden deducirse las prácticas aconsejables para evitar la aparición de este fenómeno indeseable, las que se mencionan como Métodos de prevención:
. Emplear los herbicidas sólo cuando sea necesario.
. Utilizar la rotación de cultivos.
. Utilizar programas de control integrado de malezas en los que se combine el uso de herbicidas con siegas, laboreo, etc.
. Limpiar muy cuidadosamente los equipos de laboreo y recolección, a fin de no diseminar las semillas de individuos resistentes.
. Vigilar la eficacia obtenida para ver si las fallas se deben a errores de tratamiento y no del herbicida.
. Combinar herbicidas con distintos modos de acción (19).
lll. Resistencia a glifosato
III.1. Antecedentes
El primer caso de resistencia a herbicidas registrado fue para 2,4 D, pero la temática de resistencia de malezas a los herbicidas comenzó a ser conocida en 1970, cuando se publicó el primer caso de resistencia a triazina en Senecio vulgaris, mientras que en la últimas décadas se han reportado varios casos más. En la actualidad, unas 185 especies, con 321 biotipos resistentes, distribuidas en 111 dicotiledóneas y 74 mono-cotiledóneas, presentan resistencia a nivel mundial (8) .
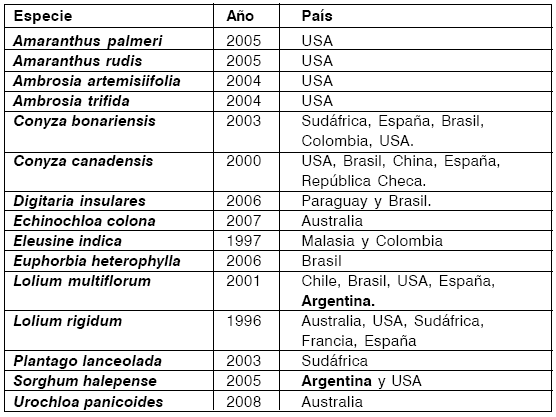
En 1974 se introdujo en el mercado el herbicida Roundup® cuyo ingrediente activo es el glifosato, y 22 años después, en 1996, se informó el primer caso de resistencia a glifosato en Lolium rigidum. En Argentina, la comprobación de especies tolerantes y biotipos resistentes al glifosato (12, 20) estaría indicando no sólo una disminución de biodiversidad, por la susceptibilidad diferencial que existe entre los taxa, sino también el aumento obligado en la frecuencia y dosis de aplicación del herbicida a corto plazo (1, 14, 21). En el Cuadro 2 se pueden observar las especies reconocidas como resistentes a glifosato, a nivel mundial.
Cuadro 2: Listado de especies de plantas resistentes a glifosato
El glifosato actúa como herbicida post-emergente de amplio espectro, no selectivo y "seguro" desde el punto de vista ambiental (baja toxicidad para organismos no blanco, bajo movimiento en el agua subterránea y persistencia limitada). Es un herbicida inhibidor de la síntesis de aminoácidos en plantas, bacterias, algas, hongos y parásitos apicomplejos, a través de la inhibición de la enzima EPSPS (5-enolpiruvil shikimato 3-fosfato sintetasa). La EPSPS es codificada por el núcleo celular y transportada al cloroplasto a través de un péptido de transporte, y es en el cloroplasto donde participa de la ruta metabólica del ácido shikìmico. En esta vía se emplea un 20 por ciento del carbono fijado durante la fotosíntesis. Esta enzima está asociada a la síntesis de tres aminoácidos esenciales cromáticos: fenilalanina, tirosina y triftofano. Además, este trayecto está relacionado a la síntesis de compuestos aromáticos como ligninas, alcaloides, flavonoides, ácidos benzoicos y hormonas vegetales (15), puesto que los aminoácidos sintetizados son precursores de estos compuestos secundarios. Ya en el cloroplasto, la EPSPS enlaza primero una molécula de shikimato-3-fosfato (S3P), inmediatamente después una molécula de PEP se enlaza al sitio activo de la enzima. La EPSPS cataliza entonces una reacción de condensación para producir 5-enolpiruvilshikimato-3-fosfato. Queda claro que PEP no presenta afinidad por EPSPS a menos que una molécula de S3P se enlace primero.
lII.2. Mecanismo de acción de la 5-enolpiruvil shikimato-3-fosfato sintetasa
S3P + enzima -> S3P.enzima +PEP -> S3P.enzima.PEP -> Enzima
EPSP -> PI+enzima+EPSP
Donde:
Enzima: 5-enolpiruvil shikimato-3-fosfato sintetasa
S3P: shikimato-3-fosfato
PEP: fosfoenolpiruvato
EPSP: 5-enolpiruvil shikimato-3 fosfato
La función de la EPSP es unir el ácido shikimico con ácido fosfoe-nolpirúvico para formar la EPSPS. Como la estructura de PEP y del glifosato son muy similares, el glifosato actúa como inhibidor competitivo y se une fuertemente al complejo formado por el shikimato y la EPSPS, resultando una acumulación de shikimato en concentraciones tóxicas. El glifosato se transporta simplásticamente hacia los meristemas de la planta en crecimiento y, al actuar como inhibidor competitivo de la EPSPS, resulta en la acumulación de shikimato y el bloqueo de la síntesis de los aminoácidos aromáticos. Reiteramos: su modo de acción es a través de la inhibición de la enzima 5-enolpiruvilshykimato-3-fosfato sintetasa (EPSPS), impidiendo la biosíntesis de los tres aminoácidos aromáticos nombrados. Ésta es la forma en que comienza a actuar el glifosato. En consecuencia, la presencia de glifosato determina supresión de crecimiento y muerte.
Se han clonado los genes correspondientes a las enzimas de esta vía. Mutaciones en el gen que codifica la EPSPS o su sobreexpresión determinan resistencia a glifosato en cultivos celulares o plantas (por ingeniería genética). Se observó, tanto en bacterias como en cultivos celulares de plantas, que el incremento en el número de copias del gen codificante para la EPSPS confiere resistencia a este herbicida.
Para el desarrollo de plantas transgénicas resistentes a glifosato se aisló el gen que codifica una EPSPS resistente a glifosato de la cepa CP4 de Agrobacterium tumefaciens (cp4 epsps) y se clonó el gen gox, que codifica la enzima glifosato oxidoreductasa (responsable del proceso de degradación del glifosato por la ruta del ácido aminometilfosfónico), a partir de la cepa LBAA de Achromobacter sp. El gen cp4 epsps derivado de la cepa CP4 de Agrobacterium tumefaciens codifica la enzima C4-EPSPS altamente insensible al glifosato. Esta enzima sólo difiere en el aminoácido de la versión de las plantas (alanita por glicina). En presencia de glifosato, las plantas que expresan C4-EPSPS continúan la síntesis de aminoácidos aromáticos.
La región codificante del gen cp4 spsps está bajo la regulación de un promotor constituido del virus del mosaico del coliflor y termina con la secuencia terminador t-nos del gen nos (nopalina sintetasa) derivado de A tumefaciens. Una secuencia de ADN derivada de Petunia x hybrida que codifica para un péptido de tránsito cloroplastídico (CTP4) se clonó en el extremo 5' del gen. La secuencia que codifica CTP4 unido a la región codificante del gen cp4epsps determina la producción de una proteína de fusión que facilita la importación de la enzima recientemente traducida dentro del cloroplasto, donde la ruta del shikimato como el sitio de acción del glifosato está localizada. Una vez importada al cloroplasto, el péptido de tránsito es eliminado y rápidamente degradado por una proteasa específica. La EPSPS es ubicua en la naturaleza y no se espera que sea tóxica o alérgena (22).
También pueden ocurrir mutaciones en la EPSPS, que tornan a la enzima insensible al glifosato. La unión de este herbicida a la enzima nativa bloquea su actividad e impide el transporte del complejo EPSPS-shikimato 3-fosfato al cloroplasto. La enzima producida por el gen mutado (epsps*) tiene una menor afinidad por el glifosato y es catalíticamente activa en presencia del herbicida. Efectivamente, se ha confirmado en varias especies que variaciones puntuales en la secuencia de nucleótidos que codifican para la EPSPS confieren resistencia al glifosato. Así en Eleusine indica y Lollium rigidum se verificó que la sustitución del aminoácido de posición 106 genera resistencia a glifosato, en tanto en L. multiflorum se verificó la misma sustitución en una población proveniente de Chile, pero no ocurrió lo mismo para otra población proveniente de Estados Unidos (15). Obviamente, estamos hablando de procesos que llevan mucho tiempo y varias generaciones, razón por la cual no es fácil ni inmediato detectar un biotipo resistente. De acuerdo a observaciones realizadas, sólo cuando el biotipo resistente supera el veinte por ciento del total de la población se habla de resistencia.
Las poblaciones de malezas se adaptan a la selección intensa ejercida por los herbicidas, cuestión que dificulta un control adecuado. Para que el desarrollo de la resistencia pueda prosperar es indispensable la variabilidad genética dentro de la población de interés. Como sabemos, la diversidad genética es el material o sustrato sobre el cual tienen lugar las mutaciones. La probabilidad de que la población adquiera resistencia dependerá de la frecuencia de mutación, de las ventajas selectivas de los alelos o genes que confieren resistencia y del tamaño de la población.
Por otro lado, la hibridación interespecífica, que promueve la diversidad genética y la evolución del genoma, es un proceso que puede facilitar la evolución y adaptación de genes resistentes y favorecer la diversidad de semillas de las especies resistentes. Entre las especies cultivadas y las malezas hay flujo génico y transferencia de genes que confieren resistencia al herbicida. Considerando que el glifosato tiene un solo modo de acción y su metabolismo es limitado en las plantas, son éstas razones suficientes para determinar una baja frecuencia de fenotipos de resistencia al glifosato, comparado con otros herbicidas (9, 24).
Existe limitado material bibliográfico que recoja el nivel de resistencia a glifosato en especies con información genética transferida autoin-compatibles, polinizadas por el viento, o de transferencia entre interfértiles. Proponemos que la hibridación de nuevas taxa con resistencia a glifosato puede ocurrir con cierta facilidad. La alta compatibilidad genética entre especies está asociada con una igual dotación cromosómica que permite el apareamiento de cromosomas en meiosis, y el intercambio de alelos en el paquiteno. Además la hibridación se ve favorecida por un aporte a la compatibilidad como es la relación filogenética entre especies muy cercanas, y por otro lado, por factores como la polinización anemófila y entomófila, dadas las características de las especies. Especies genéticamente compatibles, capaces de transferencia de alelos y producción de progenies de híbridos interespecíficos, vigorosos y fértiles pueden trasladar resistencia (23).
Estudios realizados (5) sostienen que la resistencia es estrictamente dependiente del estado de crecimiento de la planta.
Si bien se afirma que la desactivación metabólica es un mecanismo generador de resistencia, no es el caso de Conyza canadensis (L) Cronq. En los biotipos estudiados para esta especie, se han encontrado tres mecanismos hipotéticos que sostendrían la generación de resistencia. El primero de ellos tiene que ver con la traslocación del herbicida en la planta, que ha sido considerado por otros autores como el factor primario generador de resistencia. El glifosato usualmente trasloca por floema y xilema. En los biotipos R, este herbicida parece ser transportado principalmente por el xilema, lo cual representa una estrategia básica de supervivencia. Sin importar dónde ha sido absorbido el glifosato, se acumula predominantemente en hojas. La traslocación debilitada, sólo por xilema, protege el meristemo apical en crecimiento, aunque muere el resto de las hojas adultas. La aparición de hojas nuevas emitidas por el meristema apical sustituye el efecto fitotóxico generado por el glifosato en los biotipos R.
Esta translocación debilitada podría ser la consecuencia de otro evento fisiológico. La traslocación vía xilema sugiere secuestro del herbicida en el apoplasto. Por otro lado, distintas hipótesis consideran que el floema transporta los componentes iónicos como el glifosato, favorecido por cambios de pH entre el sector apoplástico (pH ácido o subácido) y el simplático (pH neutral o subalcalino). Una alteración de la bomba iónica de la membrana plasmática reduce la habilidad de los tejidos de acidificar el apoplasto, lo cual inhibe el transporte de glifosato a través del plasmalema (al mismo tiempo se reduce la traslocación). Otra hipótesis para justificar el secuestro de glifosato en el apoplasto es la mutación de algunos carriers específicos que intervienen en el transporte del herbicida a través del plasmalema. Un tercer factor es el aumento del ARNm, que es dos o tres veces myor en biotipos R, mientras que la EPSPS transcripta no es una respuesta inducida por la aplicación de glifosato pero sí con la acumulación de shikimato. En biotipos R, declina shikimato y recobra el crecimiento. El tercer factor es la aparición de ramas nuevas después del tratamiento con glifosato (5).
Considerando las características particulares del glifosato, durante mucho tiempo se pensó que las malezas no podían desarrollar resistencia a este principio activo (21, 9). Hoy no sólo se sabe que esto no es así, sino que Tuesca et. al. (21) sostienen que la resistencia se desarrolla también cuando las aplicaciones del herbicida se hacen en estadios de desarrollo avanzado (de menor susceptibilidad).
Las primeras deficiencias en el control por parte del glifosato se observaron en Salta y Tucumán, en el año 2003, y experiencias en 2004 y 2005 corroboraron la resistencia a ese principio activo. Hoy se asegura que, en la región sojera núcleo, ya hay casos de resistencia de sorgo de alepo. En el INTA Oliveros y la Cátedra de Malezas de la Facultad de Ciencias Agrarias de la Universidad Nacional de Rosario (UNR) se ha logrado determinar un grado relativamente alto de resistencia a glifosato por parte de esta especie y actualmente se está buscando la relación con los casos del NOE argentino.
Por último, el mercado ofrece otros productos con resistencia a glifosato adquirida mediante las estrategias anteriormente mencionadas. Algunos de ellos están disponibles a nivel comercial en varios países:
- Soja: promotor 2x35S / secuencia codificante cp4 epsps (1996).
- Canola: Promotor FMV (Figwort mosaic virus) / secuencia codificante cp4 epsps y promotor FMV / secuencia codificante gox (1996).
- Algodón: promotor FMV / secuencia codificante cp4 epsps (versión sintética con optimización de uso de codones; 1997).
- Maíz: promotor de actina 1 de arroz / secuencia codificante cp4 epsps y promotor 2x35S / secuencia codificante cp4 epsps (2001).
IV. Conclusiones
La revalorización de acciones de monitoreo de los cultivos que permita un seguimiento y evaluación del cambio en la riqueza específica de un campo, con especial referencia a las malezas, más un conocimiento adecuado de los herbicidas en el marco de prácticas agrícolas que tengan en cuenta los métodos de prevención de resistencia, dan el lugar a toma de decisiones convenientes. Para esto último es necesario el intercambio interdisciplinario y la capacitación del productor. En otras palabras, un conocimiento riguroso de la ecología de las malezas, el modo de acción de los herbicidas y los mecanismos de resistencias constituyen un buen trípode para generar buenos controles.
Por otro lado, la historia de los herbicidas nos está enseñando que no debemos depender exclusivamente de ellos, y mucho menos de uno solo de ellos. En poco más de 20 años conocemos 15 especies resistentes a glifosato. Si miramos la procedencia de esta información, observaremos que proviene, mayoritariamente, de países desarrollados con gran impulso y apoyo a la investigación, pero no debemos caer en el error de pensar que sólo en esos sitios se encuentran o desarrollan las especies resistentes, sino que es ahí donde se destinan recursos económicos y humanos para elucidar estas cuestiones y se generan publicaciones científicas al respecto.
Pensemos también en el riesgo latente de cruzamientos entre especies compatibles genéticamente, correspondientes a ambientes agrícolas y naturales, y la diseminación de híbridos en el ambiente. Necesitamos actuar prudentemente ante la aparición espontánea o introducida de especies modificadas.
Finalmente, es necesario poner énfasis en los problemas locales y las condiciones en que éstos se desarrollan. Si el modelo actual no se modifica y se continúa con el mismo nivel de presión de selección sobre las poblaciones y comunidades de malezas, es altamente probable que se manifiesten más casos de resistencia.
1. CARLISLE, S. M.; TREVORS, J.T. (1988). Glyphosate in the environment, en: Water Air Soil Poli., 39 (3): 409-420. [ Links ]
2. CASAFE. (2005). Guía de Productos Fitosanitarios. Buenos Aires: CASAFE. [ Links ]
3. CRISTOFFOLETI, R (2003). Curva de respuesta de resistencia y susceptibilidad de biotipo Bidens pilosa L. al herbicida inhibidor de ALS, en: Sci. Agrie, 59 (3): 513-519. [ Links ]
4. DE PRADO, R.; CRUZ-HIPOLITO, H. 2005. Mecanismos de resistencia de las plantas a los herbicidas. www.inia.org.uy/estaciones/la_estanzuela/webseminariomalezas/articulos/depradorafael.pdf [22 de octubre de 2008] [ Links ].
5. DINELLI, G.; MAROTTI, I.; BONETTI, A.; MINELLI, M; CATIZONE, R; BARNES, J. (2006). Physiological and molecular insight on the mechanisms of resistance to glyphosate in Conyza canadensis (L.) Cronq. Biotypes, en Pest. Bioch. & Physiol., 86: 30-41. [ Links ]
6. FENG P.C.C.; CHIU, T.; SAMMONS, R. D. (2003). Glyphosate efficacy is contributed by its tissue concentration and sensitivity in velvetleaf (Abutilón theophrasti), en: Pest. Bioch. and Physiol., 77(3): 83-91. [ Links ]
7. FENG P.C.C.; CHIU, T. (2005). Distribution of [14C]glyphosate in mature glyphosate-resistant cotton from application to a single leaf or over-the-top spray, en Pest. Bioch. and Physiol., 82(1): 36-45. [ Links ]
8. HEAR I. (2008). Survey of herbicide resistant weeds. En: www.Weedscience.com [18 de octubre de 2008] [ Links ].
9. JASENIUK, M.; BULE-BABEL, A.L.; MORRISON, I, (1996). The evolution and genetics of herbicides resitence in weeds, en: Leed. Science, 44: 176-193. [ Links ]
10. LORRAINE-COLWILL,D.F, POWLES, S.B.; HAWKES, T.R.; HOLLINSHEAD, R H.; WARNER, S. A. J.; PRESTON, C. (2002). Investigations into the mechanism of glyphosate resistance in Lolium rígidum, en Bioch. and Physiol núm. 74(2): 62-72 [ Links ]
11. MÓNACO, C; ACCIARESI, H. (2004). Bioherbicidas: una alternativa en el manejo de malezas en sistemas agrícolas sustentables, en: Revista Ciencia Hoy, 14(79): 30-33. [ Links ]
12. PAPA J.C. (2000). Malezas Tolerantes que Pueden Afectar al Cultivo de Soja. Información Técnica de soja y Maíz de Segunda Campaña 2000, en: Miscelánea 93: 1-24. [ Links ]
13. PAPA, J.C; CASTELLARIN, J.M.; PEDROL, H.M.; SALVAGIOTTI, M.; ROSSO, O. (2005). Efecto del herbicida metsulfuron metil sobre el rendimiento de un cultivo de trigo pan, en: Para mejorar la producción, 28: 56-58. [ Links ]
14. PENGUE, W. (2000). Cultivos Transgénicos ¿Hacia dónde vamos? Algunos efectos sobre el ambiente, la sociedad y la economía de la nueva "recombinación" tecnológica. Programa de Ciencia y Tecnología para América Latina y el Caribe. Buenos Aires: UNESCO. [ Links ]
15. PÉREZ JONES, A.; POLGE, PARK, K. W.; N.; COLQUHOUN, J.; MALLORY-SMITH, C. A. (2007). Investigating the mechanism of glyphosate resistance in Lolium multiflorum, en: Planta, 226(2):395-404. [ Links ]
16. PLINE W.A.; WU, J.; HASTÍOS, K.K. (1999). Effects of Temperature and Chemical Additives on the Response of Transgenic Herbicide-Resistant Soybeans to Glufosinate and GlyphosateApplications, en: Bioch. and Physiol., 65(2): 119-13. [ Links ]
17. PLINE, W.A.; WILCUT, J.W.; DUKE, S.O.; EDMISTEN, K.L.; WELLS, R. (2002). Tolerance and accumulation of shikimic acid in response to glyphosate applications in glyphosate resistant and non-glyphosate resistant cotton (Gossypium hirsutum L), en: J. Agric. Food Chem, 50:506-512. [ Links ]
18. SANDERMANN, H. (2006). Plant biotechnology: ecological case studies on herbicide resistance, en Trends Plant Sci., núm. 3(7): 324-328. [ Links ]
19. SOCIEDAD ESPAÑOLA DE MALHERBOLOGIA. 2008. www.semh.net [ Links ]
20. TUESCA, D.; NISENSOHN, L. (2001). Resistencia de Amaranthus HBK imazetapir y clorimurón etil, en: Pesquiza Agropecuaria Brasileira, 36(4): 601-606. [ Links ]
21. TUESCA, D; NISENSOHN, L; PAPA, J.C. (2007). Para estar alerta: el Sorgo de Alepo (Sorghum halepense) resistente a glifosato, en: Revista SOJA- Para mejorar la producción, 36: 72-75. [ Links ]
22. UNIVERSIDAD DE LA REPÚBLICA. Facultad de Agronomía. Curso: Genética 2005. www.fagro.edu.uy/~genetica/docs/11%BATPract2005.pdf [ Links ]
23. VITTA, J.; FACCINI, D.; LEGUIZAMÓN, E.; NISENSOHN, L; PAPPA, J.; PURICELLI, E.; TUESCA, D. (2005). Herbicidas. Características y fundamentos de su actividad. Rosario: UNR Editora. [ Links ]
24. ZELAYA, I.; OWEN, M.K.; VANGESSEL, M. (2007) Transfer of glyphosate resistance: evidence of hybridization in Conyza (Asteraceae), en: Amer. Jour. Botany, 94(4): 660-673. [ Links ]

