










Servicios Personalizados
Revista

Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO  uBio
uBio
Compartir

 Permalink
PermalinkBoletín de la Sociedad Argentina de Botánica
versión On-line ISSN 1851-2372
Bol. Soc. Argent. Bot. v.40 n.1-2 Córdoba ene./jul. 2005
Variaciones Intra-Específicas en el crecimiento primario de Nothofagus dombeyi (Nothofagaceae)
Javier Puntieri1
1 Departamento de Botánica, Centro Regional Universitario Bariloche, Universidad Nacional del Comahue, Quintral 1250, 8400 Bariloche, y Carrera del Investigador Científico y Tecnológico, CONICET. jpuntier@crub.uncoma.edu.ar
Resumen: Se evaluó el patrón de crecimiento bajo condiciones controladas de los ejes foliados de 16 plántulas de Nothofagus dombeyi. Los individuos fueron ubicados en una cámara de crecimiento y se evaluó el crecimiento del tronco en dos eventos de crecimiento sucesivos (julio 2002 - marzo 2004). Se identificaron tres patrones de crecimiento del primer brote medido: (1) crecimiento ininterrumpido por unos 8 meses, (2) crecimiento interrumpido por un período de reposo no menor de 7 semanas, con rebrote posterior y (3) crecimiento ininterrumpido por 6 a 7 meses. En los individuos con los patrones de crecimiento 1 y 2, la tasa máxima de producción de hojas del primer brote producido fue mayor que en los individuos con el tercer patrón de crecimiento. La tasa media de crecimiento fue mayor para los brotes del primer grupo que para los de los otros dos grupos. El segundo de los brotes medidos se alargó en un único pulso y alcanzó menor tamaño que el primero de los brotes. Se discute el posible rol de gradientes ontogenéticos o de variaciones genéticas entre individuos de coihue como explicación de los diferentes patrones de crecimiento registrados.
Palabras clave: Brote; Policiclismo; Crecimiento; Yema; Dormancia
Summary: Intra-specific variations in the primary growth of Nothofagus dombeyi (Nothofagaceae). The growth pattern under controlled conditions was evaluated for foliated axes of 16 Nothofagus dombeyi seedlings. The seedlings were placed in a growth chamber and the growth of their trunk was assessed weekly throughout two successive extension events (July 2002 - March 2004). Three growth patterns were identified for the first measured shoot: (1) uninterrupted growth for about 8 months, (2) growth interrupted by a resting period of at least 7 weeks with later re-sprout (3) uninterrupted growth for about 6-7 months. The maximum rate of leaf production for the first shoot was higher for those seedlings with the former two growth patterns than for those with growth pattern 3. The mean rate of shoot growth was higher for the seedlings with growth pattern 1 than for those with growth pattern 2 or 3. The second shoot produced extended in a single flush and was smaller than the first shoot for each seedling. The possible role of ontogenetic and genetic factors on the intra-specific variations in shoot growth pattern is discussed.
Key words: Shoot; Polycyclism; Growth; Bud; Dormancy
INTRODUCCIÓN
El crecimiento longitudinal, o primario, de los ejes foliados que componen la estructura aérea de las plantas consiste en la diferenciación de nuevos órganos (nudos, entrenudos y hojas) a partir de un meristema y en su alargamiento. En las plantas leñosas se han identificado dos modos básicos de crecimiento. Se considera que una especie presenta crecimiento continuo cuando sus ejes crecen siempre que las condiciones ambientales son propicias. En estas plantas, el crecimiento puede interrumpirse debido a factores exógenos, como bajas temperaturas, fotoperíodo corto o baja disponibilidad de agua (Hallé & Martin, 1968; Caraglio & Barthélémy, 1997; Grosfeld, 2002; Grosfeld & Barthélémy, 2004). Por otro lado, en las plantas con crecimiento rítmico, los ejes foliados exhiben alternancia de fases de crecimiento y fases de reposo aun bajo condiciones ambientales constantes y favorables (Champagnat et al., 1986; Caraglio & Barthélémy, 1997). En este último caso, el reposo es inducido por factores endógenos (inhibiciones generadas por otros órganos de la misma planta o por el estado fisiológico del meristema terminal del eje; Champagnat, 1989). En las especies con crecimiento rítmico, la porción de eje desarrollada en un año se suele denominar brote anual; ésta puede desarrollarse en un único evento ininterrumpido de creci miento (brote monocíclico) o derivar de varios eventos de crecimiento separados por fases de reposo (brote policíclico; Caraglio & Barthélémy, 1997).
En cada región climática conviven especies con crecimiento rítmico y especies con crecimiento continuo, de manera que la determinación del patrón de crecimiento de una especie no puede inferirse a partir del conocimiento de su área de distribución geográfica (Lavarenne-Allary, 1965; Hallé & Martin, 1968; Gill, 1971; Hallé et al., 1978; Caraglio & Barthélémy, 1997). Las características morfológicas de los ejes de una especie pueden contribuir a identificar ese patrón. En ejes con crecimiento rítmico es habitual la presencia de hojas modificadas como catafilos (hojas carentes de clorofila y frecuentemente escuamiformes), separadas por entrenudos muy cortos, o de marcas dejadas por este tipo de hojas sobre el tallo. Contrariamente, los ejes con crecimiento continuo suelen no presentar catafilos ni exhibir una alternancia destacada de porciones de entrenudos cortos y de entrenudos largos (Caraglio & Barthélémy, 1997; Grosfeld, 2002; Grosfeld & Barthélémy, 2004). Existen, sin embargo, especies de Cupressaceae con crecimiento continuo en las cuales pueden desarrollarse entrenudos cortos bajo condiciones de crecimiento sub-óptimas (Grosfeld, 2002; Grosfeld & Barthélémy, 2004). La prueba más concluyente para determinar el tipo de crecimiento de los ejes de una planta es la medición reiterada de individuos en diferentes ambientes naturales o creciendo bajo condiciones controladas y favorables para el crecimiento (Champagnat et al., 1986).
El género Nothofagus incluye especies de Sudamérica y Oceanía consideradas, tradicionalmente, miembros de la familia Fagaceae (Correa, 1978). Estudios moleculares y morfológicos sugieren que este género se encontraría tan emparentado con la familia Betulaceae como con la familia Fagaceae y sustentan la formación de la familia monotípica Nothofagaceae (Hill, 1992; Hill & Dettmann, 1996; Manos, 1997). Entre las características morfológicas que diferencian a las especies sudamericanas de Nothofagus de las especies de Fagaceae y Betulaceae mejor conocidas se encuentra la estructura de la yema terminal, la cual presenta varios catafilos (a veces más de diez) en especies de estas familias (Collin et al., 1996; Nitta & Ohsawa, 1998), y puede describirse como desnuda (desprovista de catafilos) en todas las especies sudamericanas de Nothofagus estudiadas hasta el momento (Barthélémy et al., 1999; Puntieri et al., 2002). La asociación entre la presencia de catafilos en la yema terminal y el crecimiento rítmico se ha corroborado para especies de Fagaceae y Betulaceae (Collin et al., 1996; Nicolini, 1998; Nitta & Ohsawa, 1998). Por el contrario, no se ha demostrado hasta el momento que la existencia de yemas terminales desnudas en las especies sudamericanas de Nothofagus se vincule con la capacidad de crecimiento continuo de sus ejes. Si bien estudios a campo demuestran una marcada estacionalidad en el alargamiento de los ejes de coihue, Nothofagus dombeyi (Mirb.) Oerst. (Puntieri et al., 1998, 2002), se desconoce en qué medida dicha estacionalidad estaría determinada por factores exógenos. A fin de evaluar la hipótesis sobre la potencialidad de crecimiento continuo de los ejes de N. dombeyi, se colectaron plántulas de esta especie de una población natural, y se midió semanalmente el crecimiento de las mismas en una cámara de crecimiento con fotoperíodo constante y disponibilidad de agua y temperaturas controladas.
MATERIALES Y MÉTODOS
El 27 de junio de 2002, se recolectaron plántulas de coihue de dos años de edad de un área de alrededor de 100 m2 cubierta por bosque de la misma especie, al borde de la ruta nacional 258, en cercanías de la localidad de Villa Mascardi (Parque Nacional Nahuel Huapi, Argentina). Se tomaron varios bloques de suelo incluyendo, en total, 16 plántulas de entre 3,0 y 6,3 cm de altura (promedio = 4,8 cm, error estándar = 0,2 cm); todas ellas constaban de un único eje vertical no ramificado y no mostraban signos de daño por herbívoros. Al momento del muestreo, las plántulas tenían un diámetro basal de entre 0,9 y 1,9 mm (promedio = 1,3 mm, error estándar = 0,1 mm). Los bloques de suelo se cortaron a no menos de 10 cm de distancia del tallo de las plántulas, de manera de minimizar el daño a las raíces. De inmediato, los bloques de suelo fueron dispuestos, evitando su destrucción, en una bandeja de 28 x 43 cm de lado y 9 cm de profundidad, provista de doce perforaciones de drenaje de 5 mm de diámetro. Se completaron los espacios de la bandeja entre los bloques con suelo tomado de la misma localidad. De cada plántula se midieron la altura total desde el nivel de los cotiledones y la longitud (con calibre) y el número de hojas del brote producido en el segundo año de desarrollo.
La bandeja con las plántulas se ubicó sobre otra bandeja algo mayor colocada en el piso de una cámara de crecimiento de 50 x 45 cm de base x 130 cm de altura, iluminada con tubos Osram Fluora (cuatro tubos de 36 w y cuatro tubos de 20 w; 70 µmoles de fotones . m-2 . seg-1, medido con un sensor de PAR LiCor 6200) y 16 hs. de luz diarias. El riego fue provisto por goteo sobre la bandeja inferior, el que fue controlado diariamente a fin de evitar la saturación o la falta de agua. La cámara se encontraba refrigerada por un ventilador eléctrico de funcionamiento permanente. Al momento de realizado el muestreo, las plantas se encontraban en fase de dormancia invernal. A partir del momento en que se observaron cambios indicativos del inicio de la brotación de las plantas (8 de julio de 2002) y en los diez meses siguientes, se registraron semanalmente la longitud (con regla, con una precisión de 1 mm) y el número de hojas del nuevo brote vertical producido por cada planta, así como las temperaturas máxima y mínima (± 0,5°C, con un termómetro digital) dentro de la cámara de crecimiento. El criterio seguido para incluir una hoja en el conteo de hojas de un brote fue la separación total de la lámina de la hoja en crecimiento del ápice del brote que la produjo. El 8 de agosto de 2003, luego de transcurridos por lo menos tres meses desde la detención del crecimiento de todas las plantas, la bandeja fue transferida a condiciones frías (< 10°C de temperatura máxima, < 5°C de temperatura mínima) y de fotoperíodo más corto (11 hs. de luz diarias), bajo las cuales se las mantuvo por alrededor de tres meses. En ese período se midió cada entrenudo componente del brote desarrollado en la cámara de crecimiento con un calibre digital (precisión 0,1 mm).
El 10 de noviembre de 2003, la bandeja fue colocada nuevamente en la cámara de crecimiento bajo las mismas condiciones que al inicio del experimento. Se registraron semanalmente la longitud y el número de hojas del nuevo brote vertical producido a partir del ápice del tronco de cada planta. En las cuatro plantas en las que se produjo la muerte del ápice del eje principal al final del primer período de alargamiento, se procedió a medir el alargamiento de un brote vertical cercano al ápice de la planta, que se supuso, de acuerdo con conocimientos previos acerca de esta especie, constituiría un brote relevo del tronco. Se continuó el registro del tamaño de los nuevos brotes hasta por lo menos tres meses después de la detención del alargamiento de todos los brotes medidos. Por la información de que se dispone sobre especies con crecimiento continuo (Caraglio & Barthélémy, 1997), y el bajo nivel de variación en las condiciones ambientales en la cámara de crecimiento, se consideró que una interrupción de tres meses en el alargamiento correspondía a un estado de dormancia de origen endógeno.
Para cada período de cuatro semanas en el que se registraron incrementos en el número de hojas o en la longitud del brote, se calcularon la tasa de producción de hojas y la tasa de alargamiento, respectivamente (de aquí en adelante denominadas tasas mensuales). Promediando estas tasas para todo el período de alargamiento de cada brote, excluyendo aquellos meses con tasa nula, se obtuvieron la tasa mensual media de producción de hojas (TH) y la tasa mensual media de alargamiento (TA). Se determinaron, asimismo, la tasa mensual máxima de producción de hojas (THM) y la tasa mensual máxima de alargamiento (TAM) por brote.
Estudios anteriores sobre plantas de esta especie desarrolladas en condiciones naturales, indican que una de las características que evidencia el gradiente de diferenciación morfogenética de los ejes es la dinámica de crecimiento de los brotes. Ejes en las primeras fases del gradiente de diferenciación desarrollan brotes con dos o más períodos de elevada tasa de crecimiento separados por períodos en que dicha tasa es baja, mientras que ejes en fases más avanzadas de ese gradiente desarrollan brotes con un solo pico de crecimiento (Puntieri et al., 1998, 2002). Por esta razón, se consideró adecuado clasificar las plantas según el patrón de crecimiento del primero de los brotes medidos (que presentó mayor variación de tamaño entre individuos que el segundo brote). Dicho patrón fue definido según el número de picos y la duración del crecimiento.
Análisis estadísticos
Los primeros y segundos brotes medidos fueron comparados separadamente entre grupos de plantas en cuanto a su longitud, número de hojas y tasas de alargamiento y de producción de hojas. Para ello se utilizó análisis de variancia de un factor y pruebas F de Fisher (Sokal & Rohlf, 1981). Se adoptó un nivel de significación del 5% en todas las comparaciones.
Se buscó una función que explicara la variación en la longitud de cada entrenudo medido a partir de las siguientes variables independientes: (a) el patrón de crecimiento del brote, (b) la posición del entrenudo (contada comenzando en la base del brote) relativa al número total de entrenudos del brote, (c) la posición relativa del entrenudo elevada al cuadrado, (d) el número de hojas del brote correspondiente, (e) la semana de desarrollo del entrenudo, (f) la semana de desarrollo del entrenudo elevada al cuadrado, (g) la temperatura máxima en la semana de desarrollo del entrenudo y (h) la temperatura mínima en la semana de desarrollo del entrenudo. En la selección de estas variables se tuvieron en cuenta los posibles factores que podrían afectar la longitud de los entrenudos. Al incorporar la semana de desarrollo de un entrenudo en el análisis, se evaluó la posible influencia de factores ambientales (diferentes de las temperaturas extremas) o endógenos con efecto simultáneo sobre todos los individuos. Las variables c y f fueron incorporadas luego de analizar gráficamente las relaciones entre la posición relativa de un entrenudo y la semana de desarrollo del entrenudo con la longitud del entrenudo. Se empleó un análisis de regresión por pasos (stepwise, forward selection; Sokal & Rohlf, 1981), adoptándose como criterio de incorporación de cada variable en el modelo su grado de significación en la regresión (evaluado mediante el test F de Fisher).
Se calculó el índice de correlación de rangos de Spearman (Sokal & Rohlf, 1981) entre las tasas de producción de hojas y de alargamiento medias y máximas de ambos brotes medidos para cada planta.
Todos los análisis estadísticos fueron realizados mediante el programa estadístico Minitab 7.1 (1989).
RESULTADOS
Las temperaturas máximas y mínimas semanales registradas en la cámara de crecimiento variaron a lo largo del período de estudio (Fig. 1). La diferencia entre temperatura máxima y temperatura mínima se mantuvo aproximadamente constante a lo largo del estudio. Los valores máximos se registraron en los períodos diciembre 2002 - febrero 2003 y enero - febrero de 2004 y fueron semejantes a los que pueden medirse durante el período crecimiento del coihue en las áreas donde éste se desarrolla. Las temperaturas mínimas fueron algo mayores que el promedio que puede registrarse a campo en ese período (Conti, 1998).

Fig. 1. Temperaturas máximas y mínimas semanales a lo largo del desarrollo de las plantas de N. dombeyi en la cámara de crecimiento.
Todas las plantas respondieron positivamente a las condiciones impuestas. El brote terminal del eje principal de cada planta continuó la dirección de desarrollo vertical, y los ejes laterales presentaron desarrollo mucho más limitado, de manera que la interferencia aérea entre las plantas fue relativamente baja. El despliegue de las hojas y el alargamiento de los entrenudos fueron, en general, simultáneos; en muy pocos se produjo el despliegue de una hoja sin alargamiento de entrenudos próximos. El número de hojas desplegadas para el primer brote medido de cada planta siguió una tendencia aproximadamente linear al inicio del alargamiento. El alargamiento de los brotes fue relativamente lento en las primeras dos o tres semanas de ese período y se fue incrementando posteriormente, siguiendo una tendencia linear por varias semanas o meses, según el caso (Apéndice 1). Se diferenciaron tres grupos según el patrón de crecimiento del primer brote medido. Grupo 1 (Fig. 2 A, B): en siete de las plantas, la fase de crecimiento linear fue seguida por una fase de producción de hojas y de alargamiento más lentos, completándose un período de crecimiento total de más de ocho meses. Grupo 2 (Fig. 2 C, D): en otras siete plantas se detuvo la producción de nuevas hojas por un período no menor de siete semanas, al cabo del cual se reinició la producción de hojas. Tanto el momento de detención de la producción de hojas como la duración de los períodos de producción de hojas y de reposo dependieron de la planta considerada. En algunas de estas plantas el rebrote formado fue muy corto y contó con un número bajo de hojas con relación al número total de hojas del brote. Grupo 3 (Fig. 2 E, F): el brote medido de las restantes dos plantas se alargó en forma continua por no más de siete meses. En promedio, las plantas de estos grupos no difirieron en altura ni en el número de hojas o la longitud del brote producido previamente al desarrollo en la cámara de crecimiento (Tabla 1). El número de hojas y la longitud del primer brote producido en la cámara de crecimiento fueron, en promedio, mayores para las plantas del grupo 1 que para las de los grupos 2 y 3, entre los cuales hubo diferencias menos marcadas en ambos parámetros (Tabla 2 A). Los brotes de los grupos 1 y 2 presentaron mayores THM que los del grupo 3 y los brotes del grupo 1 tuvieron mayor TA que los de los otros dos grupos (Tabla 3). No se detectaron diferencias entre grupos en cuanto a la TH y la TAM (Tabla 2 B, C). Durante el desarrollo de los brotes no se registraron muertes de ápice, pero el ápice de cuatro de las plantas (dos de ellas del grupo 1, una del grupo 2 y una del grupo 3) se secó luego de finalizado el alargamiento del brote. El brote relevo de esas plantas derivó de la yema axilar más cercana al ápice muerto. Cabe destacar que la muerte espontánea del ápice de un brote luego del alargamiento del brote es un evento frecuente en esta y otras especies de Nothofagus sudamericanas (Barthélémy et al., 1999).

Fig. 2. Número de entrenudos y longitud (mm) de dos brotes sucesivos del eje vertical principal de plántulas de N. dombeyi en cámara de crecimiento, agrupadas según el patrón de alargamiento del primero de los brotes. Grupo 1 (A, B): alargamiento prolongado no interrumpido; Grupo 2 (C, D): alargamiento interrumpido por lo menos durante cuatro semanas; Grupo 3 (E, F): alargamiento breve no interrumpido. Para cada grupo, se indican en trazo grueso las curvas de la planta que desarrolló el primer brote con mayor número de hojas. X: muerte del ápice al finalizar el alargamiento del primer brote.
Tabla 1. Media y error estándar (EE) de la altura total y del número de hojas y la longitud del último brote producido en condiciones naturales por plantas de N. dombeyi agrupadas según su patrón de crecimiento. F: estadístico de Fisher correspondiente a las comparaciones de ANOVA entre grupos. p: nivel de significación de F (ns: p > 0,05).

Tabla 2. Tamaño alcanzado y tasas de alargamiento y de producción de hojas del primero (1) y segundo (2) brotes desarrollados por plantas de N. dombeyi en la cámara de crecimiento, agrupados según el patrón de crecimiento del primer brote (grupos 1-3). A. Media y error estándar (EE) del número de hojas y la longitud del brote. B. Media y error estándar (EE) de la tasa mensual de producción de hojas promedio (TH) y máxima (THM). C. Media y error estándar (EE) de la tasa mensual de alargamiento promedio (TA) y máxima (TAM). F: estadístico de Fisher correspondiente a las comparaciones de ANOVA entre grupos. p: nivel de significación de F (**: p < 0,01; *: p < 0,05; ns: p > 0,05).

Tabla 3. Tasas de producción de hojas y de alargamiento del primero y segundo brotes medidos de individuos de Nothofagus dombeyi desarrollados en cámara de crecimiento, calculadas para períodos de cuatro semanas.

El segundo brote inició su desarrollo entre 1 y 4 semanas después de la re-instalación de las plantas en la cámara de crecimiento (Fig. 2). En todos los casos, el segundo brote resultó más corto y presentó menor número de hojas que el primer brote medido de la misma planta; no se registraron diferencias entre grupos en el número de hojas y la longitud de este brote (Tabla 2 A). La THM y la TA del segundo brote fueron mayores para los grupos 1 y 2 que para el grupo 3 (Tabla 2 B, C). Las TH, THM, TA y TAM fueron semejantes entre el primero y el segundo brotes dentro de cada grupo. Se obtuvieron correlaciones significativas entre la TH del primer brote y la TH y la THM del segundo brote, y entre la THM del primer brote y la THM del segundo brote (Tabla 4). Las TA y TAM de los brotes sucesivos no se correlacionaron entre sí.
Tabla 4. Matriz de correlación entre las tasas medias de producción de hojas (TH), tasas máximas de producción de hojas (THM), tasas medias de alargamiento (TA) y tasas máximas de alargamiento (TAM) entre el primero y segundo brotes producidos por cada individuo de N. dombeyi en la cámara de crecimiento. Se indican el coeficiente de correlación de Spearman y su nivel de significación (**: p < 0,005, *: p < 0,05, ns: p > 0,05).

En todas las plantas, la totalidad de las hojas de cada brote del tronco, incluyendo aquellas correspondientes a los l ímites entre brotes, desarrollaron una lámina verde. Los entrenudos correspondientes a los extremos proximal y distal del primer brote fueron levemente más cortos que los entrenudos alargados en la fase de mayor crecimiento longitudinal de cada brote (Fig. 3). El primer brote de algunas plantas del grupo 1 presentó entrenudos cortos en la porción intermedia (Fig. 3 A). En las plantas del grupo 2 se registraron entrenudos cortos en las porciones correspondientes a la detención del alargamiento entre dos pulsos (Fig. 3 B).
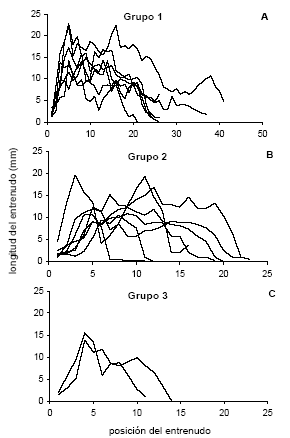
Fig. 3. Longitud de los entrenudos del primer brote desarrollado en la cámara de crecimiento (según la posición de los entrenudos contada desde el extremo proximal del brote) para plantas de N. dombeyi de los grupos 1 (A), 2 (B) y 3 (C).
Alrededor del 42% de la variaci ón en la longitud de los entrenudos del primer brote fue explica da por las variables incluidas en el análisis de regresión aplicado (Tabla 5). De estas variables, la posición relativa del entrenudo fue la que explicó, en sus términos cuadrático y linear, una mayor proporción de esa variación (Tabla 5). El número de hojas del brote y el cuadrado de la semana de desarrollo del entrenudo contribuyen en menor medida a mejorar el ajuste. Las temperaturas máxima y mínima en la semana de alargamiento del entrenudo y el grupo en el que fue incluido el brote no contribuyeron significativamente en los modelos de regresión obtenidos.
Tabla 5. Características de las funciones obtenidas en cada paso de una regresión por pasos (stepwise) para la descripción de la longitud de los entrenudos del primer brote producido por plantas de N. dombeyi en la cámara de crecimiento. Para la regresión resultante de cada paso del análisis se indican: la o las variables independientes seleccionadas, la ordenada al origen y el coeficiente (coef.) de la variable independiente, el valor del estadístico t de Student para ese coeficiente (para todos ellos p ≤ 0,001), y el coeficiente de determinación de la función (R2).

DISCUSIÓN
Dinámica de alargamiento de los brotes de N. dombeyi.
Los datos aportados por el presente estudio se oponen a la hipótesis que sugiere la posibilidad de crecimiento continuo de los ejes del coihue. La alternancia de fases de crecimiento y fases de reposo durante el desarrollo del eje principal de plantas jóvenes de esta especie se produce aún bajo condiciones constantemente favorables para su crecimiento. Alternativamente, podría sugerirse que las fluctuaciones ambientales que ocurrieron en el interior de la cámara de crecimiento habrían sido suficientes como para determinar la alternancia de fases de crecimiento primario y fases de reposo en esta especie. Bajo esta perspectiva, sin embargo, la falta de sincronismo entre individuos en la alternancia de fases sería de difícil explicación.
En otras especies cuyo crecimiento ha sido calificado como rítmico, la duración de cada ciclo constituido por una fase de crecimiento y una de reposo, depende de la especie considerada y es afectado en forma significativa solo por condiciones ambientales extremas (Hallé & Martin, 1968; Champagnat, 1989; Collin et al., 1996; Caraglio & Barthélémy, 1997). Sin embargo, el patrón de crecimiento de los brotes parece ser sumamente variable entre individuos de coihue de igual edad, en especial si se considera que los individuos medidos en el presente estudio procedían de la misma población natural y se desarrollaron bajo condiciones ambientales semejantes, tanto antes como durante el período de mediciones. Dadas las diferencias entre individuos encontradas en este trabajo, el patrón de crecimiento de N. dombeyi podría describirse como "ritmo atípico" o "crecimiento episódico", semejante al exhibido por Betula pubescens (Lavarenne et al., 1971; Caraglio & Barthélémy, 1997). Independientemente de su patrón de crecimiento, el ápice de un brote de N. dombeyi entra en una fase de reposo prolongado o dormancia (siguiendo la definición de Champagnat, 1989), que marca el final del alargamiento de ese brote.
El agrupamiento de los individuos de coihue del presente estudio en base al patrón de crecimiento del primero de los brotes medidos fue complicado por las variaciones en cuanto a la presencia, momento de ocurrencia y duración de períodos de reposo (de dos o tres semanas a más de dos meses) entre fases de alargamiento. Aunque en cierta medida arbitrario, este agrupamiento permite arribar a la siguiente conclusión. Los brotes que desarrollan hojas sin interrupción intermedia y por período mayor a ocho meses alcanzan mayor tamaño y presentan tasa máxima de producción de hojas mayor que brotes que se desarrollan por un período más corto o que presentan períodos intermedios de reposo. Este resultado se contrapone con observa ciones sobre otras especies arbóreas en las cuales los brotes desarrollados en varios eventos separados por períodos de reposo alcanzan mayor tamaño que aquellos desarrollados en un solo evento ininterrumpido (Walter & Soos, 1963; Collin et al., 1996).
El término "policiclismo" suele aplicarse a aquellos brotes en los cuales existe por lo menos una fase de detención del crecimiento con formación de yema terminal entre dos fases de crecimiento dentro de la misma estación de crecimiento (Caraglio & Barthélémy, 1997). Los brotes policíclicos son comunes en ejes vigorosos tanto de Pinus spp. como de Fagus spp. y Quercus spp. (estos dos últimos géneros emparentados con Nothofagus; Caraglio & Barthélémy, 1997). No existe acuerdo con respecto a la duración mínima del reposo entre dos picos de crecimiento a partir de la cual podría calificarse a un brote como policíclico (Caraglio & Barthélémy, 1997). No obstante, podrían considerarse policíclicos aquellos brotes medidos en este trabajo (los del grupo 2) con fases intermedias de reposo más prolongadas que aquellas registradas para brotes policíclicos de Pinus, Fagus y Quercus desarrollados a campo (Kozlowski & Ward, 1961; Walter & Soos, 1963; Nicolini, 1998; Collin et al., 1996). Estudios sobre el crecimiento y la arquitectura de especies de Nothofagus de Sudamérica han sugerido que el policiclismo sería excepcional entre estas especies (Barthélémy et al., 1999; Puntieri et al., 1998, 1999, 2000, 2002; Stecconi et al., 2000). En un estudio realizado sobre árboles jóvenes de coihue creciendo en condiciones naturales se registraron, para los brotes de mayor longitud, fases de alargamiento rápido y fases de alargamiento lento, pero no se identificaron fases de reposo de varias semanas como en el presente estudio (Puntieri et al., 1998). Aunque la existencia de brotes anuales policíclicos en plántulas de Nothofagus había sido mencionada (Thiébaut et al., 1997), no se disponía, hasta ahora, de datos cuantitativos que sustentaran esas observaciones.
En el presente estudio se observó que el inicio del alargamiento de un nuevo brote en N. dombeyi se produce en forma sincrónica luego de un período bajo condiciones menos favorables para el crecimiento (temperaturas bajas y/o fotoperíodo corto), lo cual coincide con lo encontrado en estudios a campo sobre esta especie (Puntieri et al., 1998, 2002), así como para otras especies de climas templados con crecimiento rítmico (Champagnat, 1989; Champagnat et al., 1986).
Para todas las plantas medidas en este estudio, el segundo de los dos brotes medidos fue más corto y desarrolló menos hojas que el primero, alargándose en un solo evento ininterrumpido (es decir, sin que mediara una fase de reposo ostensible). A pesar de las diferencias de tamaño entre el primero y segundo brotes, las tasas de producción de hojas y de alargamiento, tanto medias como máximas, fueron, en promedio, semejantes para ambos brotes, lo que indica que las diferencias de tamaño se debieron al mayor período efectivo de crecimiento (excluyendo los períodos intermedios de reposo) del primer brote en comparación con el segundo. De manera que el eje principal del coihue pasaría de un estadio ontogenético inicial, quizás expresado solamente en plántulas, en el cual se podrían desarrollar brotes policíclicos, a un estadio en el cual se desarrollarían exclusivamente brotes monocíclicos, como se ha observado para otras especies leñosas (Nicolini, 1998; Heuret et al., 1999).
Se demuestra en este estudio que la tasa de producción de hojas de un brote de coihue se puede relacionar con igual tasa del brote previo del mismo individuo, independientemente de la modalidad de crecimiento del primero de los brotes (Tabla 2). La relación entre el desarrollo de un eje en un período de crecimiento determinado y su desarrollo en el período siguiente había sido demostrada en un estudio sobre la lenga, N. pumilio (Passo et al., 2002). Estos resultados podrían sugerir, por un lado, la intervención de un importante componente genético en la tasa de crecimiento de los ejes de un árbol, y por otro, el establecimiento de un efecto de cascada por el cual la acción de un factor ambiental sobre el desarrollo de un brote repercutiría sobre el desarrollo de los brotes siguientes del mismo eje (Passo et al., 2002).
Desde hace varias décadas se tiene conocimiento acerca de las variaciones morfológicas y de la dinámica de alargamiento de los ejes foliados según la posición de los mismos en la planta (por ej. Kozlowki & Ward, 1961; Walters & Soos, 1963) o según la edad de la planta (Njoku, 1963, 1964; von Wühlisch & Muhs, 1986). Más recientemente, se han interpretado tales variaciones haciendo referencia a cambios fisiológicos asociados a gradientes ontogenéticos propios de cada especie (Barthélémy et al., 1997; Reffye & Houllier, 1997; Sabatier & Barthélémy, 1999; Sabatier et al., 1998). Siguiendo estas ideas, podría sugerirse como hipótesis que las diferentes modalidades de alargamiento de los ejes de coihue constituirían parte de una secuencia expresada a lo largo de la ontogenia de esta especie y que las plantas incluidas en el presente estudio habrían expresado diferentes fases de esa secuencia. Una hipótesis alternativa sería que las variaciones encontradas responderían a diferencias genéticas entre los individuos. El esclarecimiento de este punto requeriría del estudio simultáneo, en un número elevado de árboles de coihue, de las características genéticas y de la dinámica de crecimiento de brotes a lo largo de varios años (o de un análisis retrospectivo del crecimiento como el realizado por Passo et al., 2002).
Marcadores morfológicos de la dinámica de alargamiento
La alternancia de fases de alargamiento y de reposo registrada en el presente estudio se manifestó morfológicamente en variaciones de longitud de los entrenudos. A grandes rasgos, las porciones proximales y distales de los brotes presentan entrenudos más cortos que las porciones intermedias. Esto explica que la posición de un entrenudo relativa al número total de nudos del brote correspondiente (con términos cuadrático y linear) fuera la variable que más explicó la variación en la longitud de los entrenudos (~ 30%), lo cual valida el empleo de la presencia de entrenudos muy cortos como marcador morfológico del límite entre brotes en esta especie. El número de hojas del brote contribuyó en forma significativa: brotes con mayor número de hojas presentan entrenudos más largos que brotes con menos hojas. La influencia significativa de la semana de desarrollo de un entrenudo sobre su longitud podría relacionarse con la posibilidad de neoformación que presentan los brotes de esta especie, la cual se desarrolla, necesariamente, luego del alargamiento de la preformación (Puntieri et al., 2000, 2002). Los entrenudos correspondientes a la porción neoformada podrían presentar diferente longitud que aquellos preformados, como se ha observado para otras especies (Critchfield, 1960). Cerca del 60% de la variación en la longitud de los entrenudos no fue explicada por las variables tomadas en cuenta en este estudio. La existencia de fluctuaciones en la tasa de crecimiento de los brotes, de ocurrencia asincrónica en los individuos medidos, tendría una influencia significativa en la longitud de los entrenudos.
En las especies con crecimiento rítmico, los límites entre brotes anuales o entre unidades de alargamiento de un brote anual policíclico, se suelen evidenciar no solo por la presencia de entrenudos cortos sino por los catafilos asociados a ellos (Champagnat, 1989; Nicolini, 1996; Collin et al., 1996; Chaar et al., 1997; Caraglio & Barthélémy, 1997). Aunque las especies de Nothofagus estudiadas hasta el momento presentan catafilos en la base de los brotes derivados de yemas axilares (Barthélémy et al., 1999; Puntieri et al., 1999), este tipo de hojas no se desarrolla en la base de brotes o unidades de alargamiento derivadas de yema terminal. Esto indica que el mecanismo sugerido por Champagnat et al. (1986; Champagnat, 1989) según el cual la alternancia de fases de alargamiento y fases de reposo estaría regulada por interacciones entre la actividad de distintos tipos de hojas (nomofilos con lámina bien desarrollada, nomofilos con lámina abortiva y catafilos) y la actividad del meristema terminal, no tendría lugar en N. dombeyi. Thiébaut et al. (1997) hacen referencia a la presencia de hojas de lámina abortiva o tempranamente caduca en los límites entre unidades de alargamiento del mismo brote anual o entre brotes anuales sucesivos para los primeros años de desarrollo de varias especies de Nothofagus de Sudamérica. En el presente estudio no se observaron diferencias notables de desarrollo o persistencia de la lámina entre hojas cercanas y hojas alejadas de los límites. Observaciones a campo de esta y otras especies de Nothofagus (Barthélémy et al., 1999; Puntieri et al., 2002) indican que, una vez finalizado el crecimiento estacional de un eje, la hoja más proximal de la yema terminal del eje presenta, con frecuencia, una lámina parcialmente desarrollada. Durante el período desfavorable del año, y según el grado de desarrollo que haya alcanzado esa lámina, puede producirse su desecación y muerte. La observación del mismo eje en la siguiente brotación puede sugerir erróneamente la presencia de un verdadero catafilo entre ambos brotes, cuando en realidad se trata de un nomofilo cuya lámina degeneró en forma secundaria. En las plantas del presente estudio, desarrolladas bajo condiciones menos extremas de temperatura, la lámina de ese nomofilo intermedio completó su desarrollo y permaneció en pie, lo cual sugiere la intervención de factores ambientales en la desecación de la lámina entre dos brotes anuales sucesivos.
Hallé & Martin (1968) consideraron que el desarrollo de una yema terminal con hojas externas protectoras (catafilos) en ejes con crecimiento rítmico habría constituido una pre-adaptación que permitió el desarrollo de las plantas leñosas en climas con períodos desfavorables para el crecimiento. Estos autores propusieron que el ajuste del ritmo de crecimiento a la alternancia de períodos climáticamente favorables y desfavorables habría evolucionado con posterioridad. Las características morfológicas y la dinámica de alargamiento de los brotes de N. dombeyi indican que, por el contrario, un ritmo de crecimiento acorde con la estacionalidad climática de una región puede presentarse sin un alto grado de especialización morfológica de las hojas.
AGRADECIMIENTOS
A J. López, L. Mansilla, A. Inchaurza por sus contribuciones en el diseño de la cámara de crecimiento y a D. Barthélémy y un revisor anónimo por sus críticas constructivas. Este estudio fue posible gracias al apoyo económico de la Secretaría de Investigación de la Universidad Nacional del Comahue (Proyecto B116) y del CONICET (PEI 0800/99).
BIBLIOGRAFÍA
BARTHÉLÉMY, D., Y. CARAGLIO & E. COSTES. 1997. Architecture, gradients morphogénétiques et age physiologique chez les végétaux. En: Bouchon, J., P. de Reffye & D. Barthélémy (eds.), Modélisation et simulation de l'architecture des végétaux, pp. 89-136. Institut National de la Recherche Agronomique. Science Update, París. [ Links ]
BARTHÉLÉMY, D., J. PUNTIERI, C. BRION, E. RAFFAELE, J. MARINO & P. MARTINEZ. 1999. Morfología de las unidades estructurales y modo de desarrollo básico de especies patagónicas de Nothofagus (Fagaceae), Bol. Soc. Argent. Bot. 34: 29-38. [ Links ]
CARAGLIO, Y. & D. BARTHÉLÉMY. 1997. Revue critique des termes relatifs à la croissance et à la ramification des tiges des végétaux vasculaires. En: Bouchon, J., P. de Reffye & D. Barthélémy (eds.), Modélisation et simulation de l'architecture des végétaux, pp. 11-87. Institut National de la Recherche Agronomique. Science Update, Paris. [ Links ]
CHAAR, H., F. COLIN & C. COLLET. 1997. Effects of environmental factors on the shoot development of Quercus petraea seedlings. A methodological approach. For. Ecol. & Management 97: 119-131. [ Links ]
CHAMPAGNAT, P. 1989. Rest and activity in vegetative buds of trees. Ann. Sci. For. 46: 9-26. [ Links ]
CHAMPAGNAT, P., P. BARNOLA & S. LAVARENNE. 1986. Quelques modalités de la croissance rythmique endogène des tiges chez les végétaux ligneux. Naturalia Monspeliensia, Colloque International sur l'Arbre: 279-302. [ Links ]
COLLIN, P., P. M. BADOT & B. MILLET. 1996. Croissance rythmique et développement du chêne rouge d'Amerique, Quercus rubra L., cultivé en conditions contrôlées. Ann. Sci. For. 53: 1059-1069. [ Links ]
CONTI, H. A. 1998. Características climáticas de la Patagonia. En: M. N. Correa (ed.), Flora Patagónica VIII (I), pp. 31-47. INTA, Buenos Aires. [ Links ]
CORREA, M. N. 1984. Fagaceae. En: M. N. Correa (ed.), Flora Patagónica VIII (IV), pp. 4-11. INTA, Buenos Aires. [ Links ]
CRITCHFIELD, W. D. 1960. Leaf dimorphism in Populus trichocarpa. Amer. J. Bot. 47: 699-711. [ Links ]
GILL, A. M. 1971. The formation, growth and fate of buds of Fraxinus americana L. in Central Massachusetts. Harvard For. Paper 20: 1-16. [ Links ]
GROSFELD, J. 2002. Análisis de la variabilidad morfológica y arquitectural de Austrocedrus chilensis (D. Don) Pic. Serm. et Bizarri, Fitzroya cupressoides (Molina) I. M. Johnst., Pilgerodendron uviferum (D. Don) Florin y Cupressus sempervirens L. (Cupressaceae). Tesis Doctoral, Universidad Nacional del Comahue, Argentina. [ Links ]
GROSFELD, J. & D. BARTHÉLÉMY. En Prensa. Primary growth and morphological markers of inter-annual growth limits in Cupressaceae from Patagonia. Bot. J. Linn. Soc. [ Links ]
HALLÉ, F. & R. MARTIN. 1968. Etude de la croissance rythmique chez l'Hévéa (Hevea brasiliensis Müll. Arg., Euphorbiacées - Crotonoïdées). Adansonia 8: 475-504. [ Links ]
HEURET, P., D. BARTHÉLÉMY, E. NICOLINI & C. ATGER. 1999. Analyse des composantes de la croissance en hauteur et de la formation du tronc chez le chene sessile, Quercus petraea (Matt.) Liebl. (Fagaceae) en sylviculture dynamique. Canad. J. Bot. 78: 361-373. [ Links ]
HILL, R. S. 1992. Nothofagus: evolution from a southern perspective. Trends Ecol & Evol. 7: 190-194. [ Links ]
HILL, R. S. & M. E. DETTMANN. 1996. Origin and diversification of the genus Nothofagus. En: Veblen, T. T., R. S. Hill & J. Read (eds.), The ecology and biogeography of Nothofagus forests, pp. 11-24. Yale University Press, Yale. [ Links ]
KOZLOWSKI, T. T. & R. C. WARD. 1961. Shoot elongation characteristics of forest trees. For. Sci. 7: 357-368. [ Links ]
LAVARENNE-ALLARY, S. 1965. Recherche sur la croissance des bourgeons du chêne et de quelques espèces ligneuses. Ann. Sci. For. 22: 1-203. [ Links ]
LAVARENNE, S., P. CHAMPAGNAT & P. BARNOLA. 1971. Croissance rythmique de quelques végétaux ligneux de régions tempérées cultivés en chambres climatisées à température elevée et constante et sous diverses photopériodes. Bull. Soc. Bot. Fr. 118: 131-162. [ Links ]
MANOS, P. S. 1997. Systematics of Nothofagus (Nothofagaceae) based on rDNA spacer sequences (ITS): Taxonomic congruence with morphology and plastid sequences. Amer. J. Bot. 84: 1137-1155. [ Links ]
MINITAB. 1989. Minitab Data Analysis Software, standard version 7.1. State College Pensylvania 16801, USA. [ Links ]
NICOLINI, E. 1996. Développement architectural et installation de la fourche chez le hêtre europeén Fagus sylvatica L. Forêt Entreprise 110: 37-46. [ Links ]
NICOLINI, E. 1998. Architecture et gradients morphogénétiques chez de jeunes hêtres (Fagus sylvatica L. Fagaceae) en milieu forestier. Canad. J. Bot. 76: 1232-1244. [ Links ]
NITTA, I. & W. OHSAWA. 1998. Leaf dynamics and shoot phenology of eleven warm-temperate evergreen broad-leaved trees near their northern limit in central Japan. Plant Ecol. 130: 71-88. [ Links ]
NJOKU, E. 1963. Seasonal periodicity in the growth and development of some forest trees in Nigeria. I. Observation on mature trees. J. Ecol. 51: 617-624. [ Links ]
NJOKU, E. 1964. Seasonal periodicity in the growth and development of some forest trees in Nigeria. II. Observation on seedlings. J. Ecol. 52: 19-26. [ Links ]
PASSO, A., J. PUNTIERI & D. BARTHÉLÉMY. 2002. Trunk and main-branch development in Nothofagus pumilio (Nothofagaceae): a retrospective analysis of tree growth. Canad. J. Bot. 80: 763-772. [ Links ]
PUNTIERI, J., D. BARTHÉLÉMY, P. MARTINEZ, E. RAFFAELE. & C. BRION. 1998. Annual shoot growth and branching patterns in Nothofagus dombeyi (Fagaceae). Canad. J. Bot. 76: 673-685. [ Links ]
PUNTIERI, J., E. RAFFAELE, P. MARTINEZ, D. BARTHÉLÉMY & C. BRION. 1999. Morphological and architectural features of young Nothofagus pumilio (Poepp. & Endl.) Krasser (Fagaceae). Bot. J. Lin. Soc. 130: 395-410. [ Links ]
PUNTIERI, J., M. S. SOUZA, D. BARTHÉLÉMY, C. BRION, M. NÚÑEZ & C. MAZZINI. 2000. Preformation, neoformation and shoot structure in Nothofagus dombeyi (Nothofagaceae). Canad. J. Bot. 78: 1044-1054. [ Links ]
PUNTIERI, J., D. BARTHÉLÉMY, C. MAZZINI & C. BRION. 2002. Periods of organogenesis in shoots of Nothofagus dombeyi (Mirb.) Oersted (Nothofagaceae). Ann. Bot. 89: 115-124. [ Links ]
REFFYE, P. de & F. HOULLIER. 1997. Modelling plant growth and architecture: some recent advances and applications to agronomy and forestry. Current Sci. 73: 984-992. [ Links ]
SABATIER, S. & D. BARTHÉLÉMY. 1999. Growth dynamics and morphology of annual shoots, according to their architectural position, in young Cedrus atlantica (Endl.) Manetti ex Carrière (Pinaceae). Ann. Bot. 84: 387-392. [ Links ]
SABATIER, S., D. BARTHÉLÉMY, I. DUCOUSSO & E. GERMAIN. 1998. Modalités d'allongement et morphologie des pousses annuelles chez le noyer commun, Juglans regia L. 'Lara' (Juglandaceae). Canad. J. Bot. 76: 1253-1264. [ Links ]
STECCONI, M., J. PUNTIERI & D. BARTHÉLÉMY. 2000. Annual shoot-growth in Nothofagus antarctica (G. Forster) Oersted (Nothofagaceae) from northern Patagonia. Trees 14: 289-296. [ Links ]
SOKAL, R. R. & F. J. ROHLF. 1981. Biometry. Second edition. Freeman and Co., New York. [ Links ]
THIÉBAUT, B., I. SEREY, J. -L. DRUELLE, J. LI, A. BODIN & J. RECHAIN. 1997. Forme de la plantule et architecture de quelques hêtres, chiliens (Nothofagus) et chinois (Fagus). Canad. J. Bot. 75:640-655. [ Links ]
WALTERS, J. & J. SOOS. 1963. Shoot growth patterns of some Brish Columbia conifers. For. Sci. 9: 73-85. [ Links ]
WÜHLISCH, G. von & H. -J. MUHS. 1986. Influence of age on sylleptic and proleptic free growth of norway spruce seedlings. Silvae Genet. 35: 42-48. [ Links ]
Recibido el 07 de Febrero de 2005
Aceptado el 05 de Abril de 2005.

