










Servicios Personalizados
Revista

Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO  uBio
uBio
Compartir

 Permalink
PermalinkBoletín de la Sociedad Argentina de Botánica
versión On-line ISSN 1851-2372
Bol. Soc. Argent. Bot. v.40 n.3-4 Córdoba ago./dic. 2005
Micorrizas arbusculares del tipo 'Arum' y 'Paris' y endófitos radicales septados oscuros en Miconia ioneura y Tibouchina paratropica (Melastomataceae)
Carlos Urcelay1, 2 , Paula A. Tecco1 y Franco Chiarini1
1Instituto Multidisciplinario de Biología Vegetal y FCEFyN, Universidad Nacional de Córdoba - CONICET. Casilla de Correo 495, 5000 Córdoba, Argentina.
2 E-mail: curcelay@imbiv.unc.edu.ar
Resumen: Se estudió la colonización de las raíces por simbiontes fúngicos en Miconia ioneura y Tibouchina paratropica (Melastomataceae). Se observaron y describen estructuras micorrícicas arbusculares pertenecientes a los tipos 'Paris' (hifas y rulos intracelulares) y 'Arum' (hifas intercelulares y arbúsculos). Además se observaron endófitos septados oscuros (hifas y esclerocios). Se registra por primera vez la ocurrencia de ambos tipos micorrícicos arbusculares y de endófitos septados oscuros en raíces de especies pertenecientes a la familia Melastomataceae. Se discuten las implicancias ecológicas y evolutivas de la ocurrencia simultánea de los distintos tipos de colonización micorrícica en raíces de la misma especie.
Palabras clave: Tipos micorrícicos; Hongos septados ocuros; Yungas; Pastizales de altura.
Summary: 'Arum' and 'Paris' arbuscular mycorrhizal types and dark septate root endophytes in Miconia ioneura and Tibouchina paratropica (Melastomataceae). The roots of Miconia ioneura and Tibouchina paratropica (Melastomataceae) were studied for fungal symbionts colonization. Typical structures of 'Paris' (intracellular hyphae and coils) and 'Arum' (intercellular hyphae and arbuscules) arbuscular mycorrhizal types were observed and are described here. Dark septate fungi (hyphae and sclerotia) were also observed. The occurrence of both types of arbuscular mycorrhizal colonisation and dark septate fungi in the roots of species belonging to Melastomataceae is reported for the first time. The possible ecological and evolutionary implications of the co-occurrence of these mycorrhizal colonisation types in the same species are discussed.
Key words: Mycorrhizal types; Dark septate fungi; Yungas; Altitudinal grassland.
INTRODUCCIÓN
Las micorrizas son asociaciones simbióticas entre hongos y plantas. Esta simbiosis se forma en las raíces de la mayoría de las plantas terrestres donde los hongos desarrollan estructuras específicas que intervienen principalmente en el intercambio de nutrientes entre ambos simbiontes (Brundrett, 2002). La planta provee al simbionte fúngico de carbono que proviene de la fotosíntesis mientras que éste puede facilitar la provisión de nutrientes para la planta. De acuerdo a los simbiontes fúngicos involucrados en la asociación se reconocen varios tipos de micorrizas (Smith & Read, 1997). De estos tipos, sólo dos se encuentran ampliamente distribuidos: las micorrizas arbusculares y las ectomicorrizas. Los hongos micorrícicos arbusculares (HMA) colonizan alrededor del 80 % de las plantas terrestres y se relacionan principalmente con la provisión de fósforo (Smith & Read, 1997). Si bien tienen una distribución muy amplia, las micorrizas arbusculares son características de suelos donde el fósforo es el principal nutriente limitante del crecimiento de las plantas, tales como aquellos que se encuentran en regiones tropicales y subtropicales (Read & Pérez-Moreno, 2003). Las ectomicorrizas, en cambio, se relacionan principalmente con la adquisición de nitrógeno a partir de residuos orgánicos y colonizan sólo alrededor del 3%de las especies de plantas.
Recientemente, se ha observado asociación con hongos ectomicorrícicos en la especie tropical de América del Sur Graffenrieda emarginata (Ruiz & Pav.) Triana, perteneciente a la familia Melastomataceae (Haug et al., 2004). Esta familia está conformada por especies tanto leñosas como herbáceas de distribución pantropical, principalmente en América (Boelcke, 1992). Previo al antecedente de Haug et al. (2004), era considerada una típica familia asociada a micorrizas arbusculares con colonización del tipo "Paris' (Smith & Smith, 1997). Este tipo micorrícico se caracteriza por la formación de abundantes rulos intracelulares a partir de los cuales pueden desarrollarse pequeños arbúsculos en contraste con el tipo "Arum' caracterizado por desarrollar hifas intercelulares y arbúsculos intracelulares (Smith & Smith, 1997).
En nuestro país se encuentran 23 especies de Melastomataceae (2 endémicas), distribuidas principalmente en el litoral y en las selvas del noroeste (Zuloaga & Morrone, 1999). Tibouchina paratropica (Griseb.) Cogn. constituye una excepción ya que presenta una distribución disyunta, encontrándosela en el noroeste Argentino y, fuera de esos ecosistemas tropicales o subtropicales, en los pastizales de altura de las Sierras Grandes de Córdoba.
En Argentina no existen estudios relacionados a las asociaciones micorrícicas en especies pertenecientes a la familia Melastomataceae. En el presente trabajo nos propusimos estudiar y describir las estructuras micorrícicas que desarrollan: a) Miconia ioneura Griseb., especie arbustiva endémica de las Yungas en el Noroeste Argentino y b) Tibouchina paratropica, especie subarbustiva que habita en las Yungas del noroeste argentino y en los pastizales de altura del centro de Argentina.
MATERIALES Y MÉTODOS
Área de estudio y recolección de muestras
En las Yungas el clima es cálido y húmedo, las precipitaciones varían entre 900 y 2500 mm y la temperatura media entre 14°C y 26°C (Cabrera, 1971). Los suelos donde se colectaron las plantas tienen un pH que varía entre 3.9 y 5.3 (T. Easdale datos no publicados). En Pampa de Achala las precipitaciones varían entre 750 y 970 mm y la temperatura media es de 8°C con ocurrencia de heladas todo el año. Los suelos tienen un pH que varía entre 4.6 y 6.6 (Cabido et al., 1987).
Se estudiaron las raíces de dos individuos de M. ioneura colectados el día 29.XI.2004, en Salta, Dpto. Santa Victoria, a 3 km. desde el río Lipeo en dirección a Los Toldos, (Provincia fitogeográfica de la Yunga, Distrito de las Selvas Montanas, sensu Cabrera, 1971). Además se estudiaron las raíces de cinco individuos de T. paratropica colectados el día 20.XII.04 en Pampa de Achala, Copina, Dpto. Punilla, Córdoba (Provincia fitogeográfica del Chaco, distrito Chaqueño Serrano, sensu Cabrera, 1971) y de un individuo colectado el día 27.XI.2004, en Abra de las Cañas, Depto. Ledesma, Jujuy (Provincia fitogeográfica de la Yunga, Distrito de las Selvas Montanas, sensu Cabrera, 1971). Materiales de referencia fueron depositados en el Herbario del Museo Botánico: Tibouchina paratropica, Barboza et al. 1075. Prov. Jujuy. Dpto. Ledesma: Parque Nacional Calilegua, Abra de Cañas, 1750 m.s.m., 23º 40,52' 1'' S, 64º 54,05' 0''W , 27-XI-2004. Miconia ioneura, Barboza et al. 1103. Prov. Salta. Dpto. Santa Victoria, aprox. 3 km desde el Río Lipeo rumbo hacia Los Toldos, 1680 msm., 29-XI-2004.
Tratamiento de las raíces
Las raíces de los distintos individuos pertenecientes a ambas especies fueron separadas y lavadas. Se eligieron las raíces delgadas (generalmente < 2 mm de diámetro, no lignificadas en apariencia) y se clarificaron en hidróxido de potasio (KOH) al 20% durante 30 minutos a 90 °C. Después fueron lavadas con agua corriente y acidificadas en ácido clorhídrico (HCL) al 10% por 10 minutos a 90°C para luego ser teñidas con azul de anilina al 0,05% en ácido láctico 36-48hs (Grace & Stribley, 1991, Brundrett et al., 1996) a temperatura ambiente. Finalmente fueron lavadas y montadas en alcohol polivinílico (Omar et al., 1979).
Las raíces teñidas y montadas fueron observadas usando microscopio óptico Kyowa, con objetivos estándar (x10-x40-x100). Las fotografías fueron tomadas usando un microscopio Axiophot Zeiss con objetivos de contraste de interferencia diferencial.
Cortes vegetales de referencia fueron herborizados (preparados semi-permanentes) y depositados en el Herbario del Museo Botánico de la Facultad de Ciencias Exactas, Físicas y Naturales de la Universidad Nacional de Córdoba (CORD).
RESULTADOS
Ambas especies mostraron simultáneamente colonización micorrícica del tipo Arum y Paris formados por hifas de distinto grosor y colonización por "endófitos septados oscuros" en sus raíces. No se observaron estructuras ectomicorrícicas en ninguna de las dos especies.
Miconia ioneura
Esta especie mostró colonización por hifas intracelulares de mayor grosor (7-9 µm) e intercelulares más delgadas (1,5-4 µm). En algunos sectores se observaron rulos intracelulares (Fig. 1 A) propios del tipo 'Paris'', mientras que en otros se observaron arbúsculos laterales (Fig. 1 B)y terminales y sólo algunas escasas vesículas intracelulares propias del tipo 'Arum'. Los "endófitos septados oscuros" mostraron colonización por hifas septadas castañas y esclerocios intracelulares (Fig. 1 C).
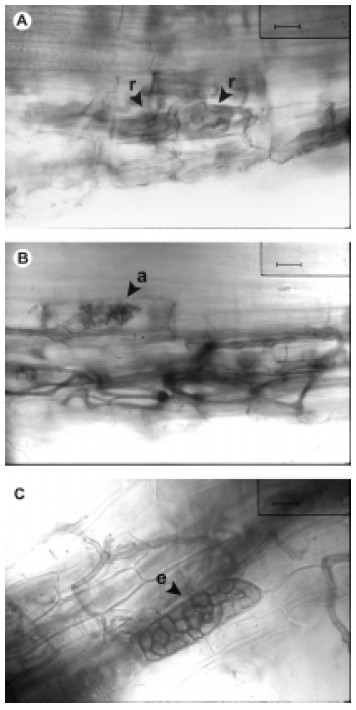
Fig. 1. Miconia ioneura A, colonización micorrícica por rulos intracelulares muy enrollados en células contiguas (tipo ‘Paris’); Barra = 12.5 µm. B, colonización micorrícica por hifas intercelulares y arbúsculo lateral (tipo ‘Arum’); Barra = 12.5µm. C, colonización por hifas y esclerocios de ‘hongos septados oscuros’; Barra = 12.5 µm. Abreviaturas: r, rulo intracelular; a, arbúsculo; e, esclerocio.
Tibouchina paratropica
Esta especie presentó colonización por abundantes rulos intracelulares, algunos muy enrollados, propios del tipo 'Paris' (Fig. 2 A). Además se observaron hifas intercelulares (Fig. 2 B), arbúsculos y vesículas intracelulares que indican colonización del tipo 'Arum'. También se distinguieron dos tipos de hifas: unas de mayor grosor (7-9 µm) que forman los rulos intracelulares y otras delgadas intercelulares (1,5-2 µm). En esta especie también se observaron hifas castañas pertenecientes a "endófitos septados oscuros" y esclerocios similares a los observados en M. ioneura.
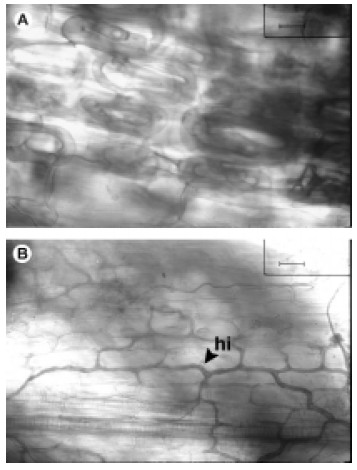
Fig. 2. Tibouchina paratropica A, colonización micorrícica por rulos intracelulares (tipo ‘Paris’); Barra = 12.5 µm. B, colonización micorrícica por hifas intercelulares (tipo ‘Arum’); Barra = 25 µm. Abreviatura: hi, hifa intercelular.
No se observaron diferencias en las estructuras micorrícicas de los individuos provenientes de los pastizales de altura en Pampa de Achala con respecto al individuo colectado en el bosque subtropical de las Yungas.
DISCUSIÓN
En este estudio se cita y describe por primera vez la presencia de simbiontes micorrícicos en las raíces de M. ioneura y T. paratropica. Previamente, Melastomataceae había sido caracterizada como una familia que presenta micorrizas arbusculares del tipo 'Paris' (Smith & Smith 1997, Haug et al., 2004) y ectomicorrizas en el caso de Graffenrieda emarginata (Haug et al., 2004). Sin embargo, en este estudio se observó que M. ioneura y T. paratropica presentaron en forma simultánea una colonización micorrícica arbuscular de los tipos 'Arum' y 'Paris'. Si bien este fenómeno ha sido observado en ciertas familias (Smith & Smith, 1997) y especies (Kubota et al., 2005), el presente estudio representa el primer antecedente de co-ocurrencia de ambos tipos de colonización en especies de la familia Melastomataceae.
De acuerdo a estudios filogenéticos basados en datos moleculares (Clausing & Renner, 2001), la tribu Merianieae, dentro de la que se ubica el género Graffenrieda, y la tribu Miconieae, que incluye a Miconia, constituirían clados basales, mientras que la tribu Melastomeae, en donde se encuentra Tibouchina, conformaría un clado con características derivadas. Si bien la filogenia se relaciona estrechamente con el tipo de asociación micorrícica presente en las distintas familias de plantas (Trappe, 1987), los resultados del presente estudio sugerirían que a nivel específico, otros factores tales como historia de vida y estrategias en el uso de los recursos influirían en el tipo micorrícico presente en las especies que componen determinada familia. En este sentido, M. ioneura presenta estructuras y características de la colonización micorrícica similares a las de T. paratropica siendo arbustivas y subarbustivas respectivamente, mientras que G. emarginata, especie arbórea, presenta micorrizas arbusculares del tipo 'Paris' y ectomicorrizas.
El rol de las plantas y los simbiontes fúngicos en la determinación del tipo asociaciones micorrícicas es un tema que ha adquirido renovada atención (Dickson, 2004). Ha sido propuesto que el tipo de colonización micorrícica arbuscular- 'Arum' y/o 'Paris' - lo determina la planta hospedante (Smith & Smith, 1997; Bedini et al., 2000). Sin embargo, estudios recientes han mostrado que el tipo micorrícico presente en las raíces de las plantas estaría determinado por la interacción entre el genotipo de la planta y el genotipo del hongo micorrícico (Cavagnaro et al., 2001; Wu et al., 2004; Kubota et al., 2005). Más aun, Dickson (2004) observó que además de los tipos 'Arum' y 'Paris' se pueden encontrar tipos intermedios, que poseen características de ambos, formados por la misma especie de HMA y que la formación de las distintas estructuras micorrícicas en las raíces estaría determinada por la interacción entre las especies de plantas y hongos involucrados. Las implicancias ecológicas de tener ambos tipos de MA ('Paris' y 'Arum') no están del todo claras porque aún no existen evidencias consistentes sobre diferencias funcionales entre estos tipos micorrícicos (Brundrett & Kendrick, 1990). Recientemente, Ahulu et al. (2005) propusieron que el tipo 'Arum' se relacionaría con especies de plantas pioneras mientras que el tipo 'Paris' estaría asociado con especies de etapas sucesionales tardías. En concordancia con esta proposición, el tipo 'Arum' ha sido relacionado con especies de crecimiento rápido (Smith & Smith, 1997) mientras que el 'Paris' a especies tolerantes de la sombra y crecimiento lento (Brundrett & Kendrick, 1990; Yamato & Iwasaki, 2002). La ocurrencia simultánea de ambos tipos en T. paratropica que crece en claros de selva en las Yungas (Meyer, 1963) y pastizales de altura de las sierras de Córdoba sobre las márgenes de los arroyos y en M. ioneura que habita en el sotobosque de las Yungas no apoyarían dichas proposiciones.
De manera similar a lo observado en este estudio, la presencia de "endófitos septados oscuros" en especies que crecen en los pastizales de altura de las sierras de Córdoba ha sido previamente notada (Urcelay, 2002) y sería frecuente en especies del bosque serrano de Córdoba (Battistella, 2003). Los "endófitos septados oscuros" son un grupo heterogéneo de Ascomycetes anamorfos que pueden ser desde patógenos a beneficiosos para las plantas dependiendo de la especie vegetal y el genotipo fúngico involucrados en la asociación (Jumpponen, 2001). No obstante, no hay estudios que hayan evaluado el efecto de estos simbiontes fúngicos sobre plantas nativas de nuestro país.
En conclusión, la co-ocurrencia de los tipos micorrícicos arbusculares 'Arum' y 'Paris' y "endófitos septados oscuros" en las dos especies de Melastomataceae observadas aquí conjuntamente a la formación de ectomicorrizas observadas recientemente (Haug et al., 2004), sugieren que el espectro de asociaciones con simbiontes fúngicos en especies de la familia Melastomataceas sería más amplio de lo que se creía y abre nuevos interrogantes acerca de las relaciones filogenéticas y ecológicas en las asociaciones micorrícicas en especies de esta familia.
AGRADECIMIENTOS
Agradecemos a Tomás Easdale por suministrar generosamente datos de suelo de Los Toldos. Dos revisores anónimos realizaron sugerencias que beneficiaron la versión final del manuscrito. CONICET y SECyT-UNC brindaron apoyo financiero para realizar el presente trabajo.
BIBLIOGRAFÍA
AHULU, E. M., M. NAKATA & M. NONAKA. 2005. Arum- and Paris-type arbuscular mycorrhizas in mixed pine forest on sand dune soil in Niigata Prefecture, central Honshu, Japan. Mycorrhiza 15: 129-136. [ Links ]
BATTISTELLA, R. 2003. Condición y colonización micorrícica en especies pertenecientes a diferentes tipos funcionales de plantas de un arbustal serrano de la provincia de Córdoba. Tesina de Grado, F.C.E.F.y N., Universidad Nacional de Córdoba. [ Links ]
BEDINI, S., A. MAREMMANI & M. GIOVANNETTI M. 2000. Paris-type mycorrhizas in Smilax aspera L. growing in a Mediterranean scherophyllous wood. Mycorrhiza 10: 9-13. [ Links ]
BOELCKE, O. 1992. Plantas vasculares de la Argentina. Editorial Hemisferio Sur, Buenos Aires. [ Links ]
BRUNDRETT, M. C. 2002. Coevolution of roots and mycorrhizas of land plants. New Phytol. 154: 275-304. [ Links ]
BRUNDRETT, M. C. & W. C. KENDRICK. 1990. The roots and mycorrhizae of herbaceous woodland plants. II. Structural aspects of morphology. New Phytol. 114: 469-479. [ Links ]
BRUNDRETT, M. C., N. BOUGHER, B. DELL, T. GROVE & N. MALAJCZUK. 1996. Working with Mycorrhizal in Forestry and Agriculture. ACIAR Monograph 32. Camberra. [ Links ]
CABIDO, M., R. BREIMER & G. VEGA. 1987. Plant communities and associated soil types in high plateau of the Córdoba Mountains, Central Argentina. Mt. Res. Dev. 7:25-42. [ Links ]
CABRERA, A. L. 1971. Fitogeografía de la República Argentina. Bol. Soc. Argent. Bot. 14: 1-42. [ Links ]
CAVAGNARO, T. R., L-L. GAO, F. A. SMITH & S. E. SMITH. 2001. Morphology of arbuscular mycorrhizas is influenced by fungal identity. New Phytol. 151: 469-475. [ Links ]
CLAUSING, G. & S.S.RENNER. 2001. Molecular phylogenetics of Melastomataceae and Memecylaceae: implications for character evolution. Amer. J. Bot. 88: 486-498. [ Links ]
DICKSON, S. 2004. The Arum-Paris continuum of mycorrhizal symbioses. New Phytol. 163: 187-200. [ Links ]
GRACE, C. & D. P. STRIBLEY. 1991. A safer procedure for routine staining of vesicular-arbuscular mycorrhizal fungi. Mycol. Res. 95:1160-1162. [ Links ]
HAUG I., J. LEMPE, J. HOMEIER, M. WEISS, S. SETARO, F. OBERWINKLER & I. KOTTKE. 2004. Graffenrieda emarginata (Melastomataceae) forms mycorrhizas with Glomeromycota and with a member of Hymenoscyphus ericae aggregate in the organic soil of neotropical mountain forest. Canad. J. Bot. 82: 340-356. [ Links ]
JUMPPONEN, A. 2001. Dark septate endophytes - are they mycorrhizal. Mycorrhiza 11: 207-211. [ Links ]
KUBOTA, M., T. P. MCGONIGLE, M. HYAKUMACHI. 2005. Co-occurrence of Arum and Paris-type morphologies of arbuscular mycorrhizae in cucumber and tomato. Mycorrhiza 15: 73-77. [ Links ]
MEYER, T. 1963. La Selva de mirtáceas de "Las Pavas". Op. Lilloana 10: 1-144. [ Links ]
OMAR, M. B., L. BOLLAND & W. A. HEATHER. 1979. A permanent mounting medium for fungi. Bull. Brit. Mycol. Soc.13:31-32. [ Links ]
READ, D. J. & J. PÉREZ-MORENO. 2003. Mycorrhizas and nutrient cycling in ecosystems - a journey towards relevance? New Phytol. 157: 475-492. [ Links ]
SMITH, F. A. & S. E. SMITH. 1997. Structural diversity in (vesicular)-arbuscular mycorrhizal symbioses. New Phytol. 137: 373-388. [ Links ]
SMITH, S. E. & D. J. READ. 1997. Mycorrhizal Symbiosis. Second edition. Academic Press, San Diego. [ Links ]
TRAPPE J.M. 1987. Phylogenetic and ecological aspects of mycotrophy in the angiosperms from an evolutionary standpoint. In: Zafir G. R. Ed. Ecophysiology of VA mycorrhizal plants. CRC Press, Boca Raton, Fl. Pp. 5-25. [ Links ]
URCELAY, C. 2002. Co-ocurrence of three fungal root symbionts in Gaultheria poeppiggi DC in Central Argentina. Mycorrhiza 12: 89-92. [ Links ]
WU, B., K. ISOBE & R. ISHII. 2004. Arbuscular mycorrhizal colonization of the dominant plant species in primary successional volcanic deserts on the Southeast slope of Mount Fuji. Mycorrhiza 14: 391-395. [ Links ]
YAMATO, M. & IWASAKI M. 2002. Morphological types of arbuscular mycorrhizal fungi in roots of understory plants in Japanese deciduous broadleaved forests. Mycorrhiza 12: 291-296. [ Links ]
ZULOAGA, F.O. & O. MORRONE (eds.). 1999. Catálogo de las Plantas Vasculares de la República Argentina. II. Monogr. Syst. Bot. Missouri Bot. Gard. 74: 1-1269. [ Links ]
Recibido el 28 de Agosto de 2005Aceptado el 21 de Octubre de 2005.

