










Servicios Personalizados
Revista

Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO  uBio
uBio
Compartir

 Permalink
PermalinkBoletín de la Sociedad Argentina de Botánica
versión On-line ISSN 1851-2372
Bol. Soc. Argent. Bot. v.41 n.1-2 Córdoba ene./jul. 2006
Las Estípulas como Caracteres Sistemáticos Confiables1
Focko Weberling2
Resumen: Generalmente la presencia o ausencia de estípulas es importante en la caracterización de familias de dicotiledóneas y en la estimación de sus relaciones de parentesco. Esto significa, que las estípulas en general son estructuras muy constantes. Tan solo debemos recordar las Rubiaceae con sus estípulas interfoliares, que en casos de material vegetativo pueden permitir la identificación como miembros de esta familia, o, en otro caso la distinción entre Rosaceae y Ranuculaceae en claves analíticas. ¿Cuál es la causa de esta constancia? ¿Esta cualidad también se refiere a estípulas rudimentarias? ¿Cuáles son las relaciones morfológicas entre formas diferentes de estípulas y sus funciones? ¿Cómo se pueden distinguir estípulas de otros apéndices basales de las hojas, que no son equivalentes? En la mayoría de los casos la aplicación de los métodos de la morfología comparada con la observación crítica de sus reglas no solamente puede llevar a la respuesta de las preguntas, sino que en combinación con métodos genéticos también, probablemente, puede abrir nuevos aspectos en cuanto a la formación de estructuras vegetales.
Palabras clave: Aparición temprana; Desarrollo precoz; Aurículas; Lóbulos de la vaina pseudoestípulas; Reducción.
Summary: The stipules as reliable systematical characters. Stipules are reckoned among the most constant vegetative characters in Dicotyledons. They are formed as marginal outgrowths of the leaf base, at the border of the two integral parts of the leaf, the leaf base ("Unterblatt") and the "Oberblatt", the distal part of the leaf, which forms the lamina and the petiole. Their ontogeny is characterized by an early, often extremely early, appearance and a following precursive development. Therefore the stipules can temporarily exceed the other parts of the leaf, which enables them to function as protecting organs for the terminal bud of a shoot and even for the same leaf. The protecting function is not the only function of stipules. They can also work as assimilation organs or can be developed as spines or glands. In pinnate leaves with basipetal segmentation the stipules often have finished their growth whereas the segmentation of the lamina still is progressing. This confirms the opinion of Eichler (1861) who regarded "Unterblatt" (leaf base) and "Oberblatt" (the distal part of the leaf) as relatively independent components of the leaf. The early appearance of the stipules in the ontogeny of the leaf certainly is responsible for their high constancy and consequently their high systematic value. Form and position of the stipules can vary according to the "principle of variable proportions". In such cases, however, the study of their ontogeny can prove the morphological identity and elucidate the relations between different forms. Often stipules are confused with non homologous structures like basal auricules or pseudostipules, basal leaflets which are more or less transformed. Usually these structures are not as constant elements as the sitpules and they can be easily identified by comparative methods including the study of their ontogeny. On the other hand, stipules often appear to be absent because they are caducous organs or rudimentary. The distribution of stipules within the system of Dicotyledons and some comparative indications point to the possibility that stipules may have evolved several times and independently.
Key words: Early appearance; Precursive growth; Auricules; Vaginal lobes; Pseudostipules; Rudimentation.
INTRODUCCIÓN
Las Etípulas como caracteres Sistemáticos
La presencia o ausencia de las estípulas es mencionada usualmente en las descripciones sistemáticas de familias y órdenes de Angiospermas. La causa es, que normalmente estos apéndices basales de las hojas son órganos característicos y constantes. Ocurren en un grandísimo número de familias y ayudan a identificar plantas. Además pueden ser interpretados como indicios de parentesco.
Ejemplos bien conocidos son las Rosaceae o las Leguminosas. En muchas Rubiaceae frecuentemente ya se puede comprobar la pertenencia a esta familia en material solamente vegetativo por la presencia de estípulas "interfoliares" situadas entre las hojas opuestas. No obstante, la presencia o ausencia de estípulas no siempre es mencionada con la certeza suficiente, quizás porque no se han investigado bien, quizás porque las estípulas caducas ya se habían caído o solamente existen estípulas rudimentarias y poco visibles. Por eso Ridsdale ya en 1974, escribió una nota reclamando: "Stipules, stipules, more stipules!".
Aparte de eso, hay apéndices basales de las hojas vegetativas, que no son homólogos con estípulas, es decir: que no son elementos comparables en el sentido de la morfología comparada. Esto no sólo está indicado por su constancia menor, sino también por otras cualidades, que se discutirán más adelante. Esto muestra, que es necesario definir los órganos designados como estípulas con precisión en base a la morfología comparada.
RESULTADOS
Definición y Propiedades de las Estípulas, Criterios de Homología
Las estípulas son apéndices de la parte basal de las hojas (Unterblatt) situadas a ambos lados de la inserción de la misma. Su aparición es al inicio de la ontogenia de la hoja y tienen un desarrollo proléptico. Según el principio de las proporciones variables (Goethe, véase Troll, 1949) los elementos del plan estructural de un grupo de organismos presentan variaciones en las proporciones relativas que integran su estructura. En este sentido las estípulas también pueden presentarse en muchas formas diferentes y su posición puede ser diversa. En casos dudosos es necesario examinar los caracteres de los apéndices basales siguiendo los criterios de la morfología comparada (criterios de homología, Remane, 1956). De los cuales se tendrán en cuenta los siguientes:
1. el criterio de la posición equivalente en sistemas estructurales equivalentes
2. el criterio de series de formas de transición
3. el criterio de la calidad especial (aplicable en algunos casos).
Estos criterios se pueden aplicar a estructuras anatómicas complicadas y en algunos rasgos característicos de la ontogenia de los órganos, como por ejemplo en las diferentes maneras de la formación de las corolas gamopétalas por una activación temprana o tardía de meristemas (Erbar, 1996) y el desarrollo precoz de las estípulas como se explica a continuación.
Para examinar si apéndices basales pueden ser interpretados como estípulas o no, la comparación de las formas diversas y su ontogenia ya nos brindan una base confiable, porque en el caso de formas raras o todavía no conocidas muchas veces se pueden hallar plantas que de alguna manera presentan formas transicionales que pueden relacionar estas formas inusuales con la forma usual (criterio de series de transición). Muchas veces el estudio de la ontogenia da buenos resultados, como en caso de órganos que en su forma adulta parecen muy diferentes, pero usualmente son más o menos similares en distintos estadios de su ontogenia temprana (en el "punto de contacto", Remane, 1956, 46). De esta manera el estudio de la ontogenia puede completar el criterio de series de formas de transición o sustituir formas intermedias. La condición indispensable para aplicar el criterio de series de transición es, que solamente podemos comparar organismos ligados por parentesco, lo cual debe ser comprobado por la coincidencia en caracteres independientes de los organismos en cuestión.
Usualmente las estípulas están situadas a ambos lados de la inserción de la hoja y en su consistencia foliácea pueden ser parecidas a la lámina de hojas vegetativas (por ejemplo: Fagonia cretica L., Fig. 3); en cuyo caso generalmente son más o menos asimétricas en su contorno . En la lengua Alemana esta posición ha llevado a la designación "Nebenblätter", que significa hojas adyacentes (o accesorias). Sin embargo A. W. Eichler en su tesis sobre la ontogenia de las hojas (1961) ya ha comprobado, que las estípulas son partes basales de la hoja. Esto coincide con los resultados de Colomb (1887), donde se observa que la innervación de las estípulas es realizada por los hacecillos conductores laterales del rastro foliar (traza foliar), como explican las Figs. 1 A , B.
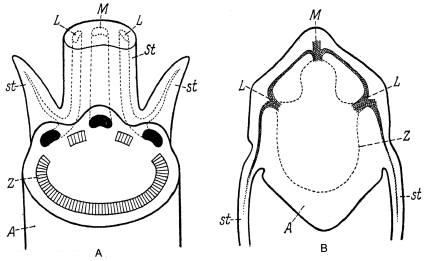
Fig. 1. A Diagrama para explicar la inervación de estípulas, A: eje; Z: cilindro central (corte transversal), trilacunar; los hacecillos vasculares del rastro foliar (negro) entran a la base de la hoja, como hacecillo mediano (M) y laterales (L). Los hacecillos laterales proveen las estípulas (st) con un hace lateral del segundo orden; St: pecíolo de la hoja. B Viola tricolor , transcorte de un nodo, designaciones como en A . A según Sinnot & Bailey, B según Colomb.
Entre los criterios de homología el más importante es el criterio de la posición equivalente entre el mismo plan de organización biológica (plan estructural, véase Remane, 1956), como explican los dos esquemas en Fig. 2, en los cuales las posiciones relativas de los puntos han cambiado, pero las conexiones han persistido ("principe des connexions"; St. Hilaire, 1830).

Fig. 2. Diagrama para explicar el criterio de la posición equivalente: dislocación de puntos distintos ( a, b, c - a 1 , b 1 , c 1 ) bajo de las mismas conexiones. No obstante el cambio de las posiciones los puntos pueden ser interpretados como elementos homólogos (según Remane 1956).

Fig. 3 . Fagonia cretica L., A , B : estadios de la ontogenia de las hojas vegetativas, C : hojas adultas bajo de la flor terminal, con el eje de un brote axilar (de Weberling 1956c).
Existen formas muy diferentes, en la posición de las estípulas en relación a la lámina y el tallo, que también pueden ser explicadas por la existencia de formas de transición y por la observación de su ontogenia. Estípulas libres se presentan como insertadas a los lados de la inserción del pecíolo de la hoja, cuando la vaina ("Unterblatt") queda corta (Fig. 4 F , Fagonia cretica, Fig. 3). Por otro lado la posición de estípulas adnatas resulta cuando toda la zona basal de la hoja ("Unterblatt") se extiende por un crecimiento longitudinal formando una vaina (Fig.4 E entre x y x'), lo que sucede después de la formación temprana de las estípulas. En el estadio adulto dan la impresión de que están fundidas al pecíolo, (por ejemplo en las hojas de Rosa ). En contraposición a las estípulas externas (Fig. 4 A , B ) la posición de las estípulas internas resulta, cuando los márgenes meristemáticos de la base de la hoja confluyen en la parte proximal del pecíolo, formando un pecíolo unifacial (Fig. 4 C , D). En estas condiciones también es posible que los órganos laterales puedan estar unidos como una estípula intrapeciolar (axilar), Melianthus major, (Fig. 4 M ) . La posición contraria se encuentra en la estípula opuesta: debido a la dilatación del meristema marginal en los bordes de la base foliar encerrando el tallo Fig. 4 G , H), las estípulas pueden colocarse en una posición contrapuesta a la inserción del pecíolo y estar más o menos unidas, como en Coronilla montana (Fig. 4 I). En muchas plantas las estípulas son foliáceas como la lámina y sirven para la asimilación de CO2. Las estípulas externas frecuentemente forman escamas gemarias y debido a su desarollo precoz (vease más bajo) aun pueden envolver y proteger no solamente la yema axilar sino también la lámina propia. Esto se observa en muchas Fagaceae, Betulaceae (Fig. 5, Carpinus betulus L.), Salicaceae o en otras plantas leñosas. No es raro, que la forma de ambas estípulas de una misma hoja pueda ser muy diferente como muestran las especies del género Azara (Flacourtiaceae) . En Crataegus oxyacantha L . (Rosaceae) como en otras especies de este género , las estípulas de las hojas de los braquiblastos y macroblastos difieren no solamente en su tamaño sino también en su forma (Fig. 17 D, E). Ambas formas de estípulas son asimétricas, pero en las estípulas grandes de los macroblastos el crecimiento en el lado exterior de la base continúa por un espacio de tiempo, a consecuencia de más divisiones celulares o un alargamiento de las células; lo cual produce un ensanchamiento en las zonas entre los nervios simpodiales. Esto resulta en la formación de una parte amplexicaule, similar a una aurícula (Fig. 17 E). En otros casos las estípulas están transformadas en espinas como en Robinia o la mayoría de las Mimosaceae (estípulas espinosas). En muchos taxones las estípulas están más o menos reducidas formando estípulas rudimentarias (muchas Mirtales, las Brassicaceae, Resedaceae y otras familias de las Capparidales , véase la disposición en Tabla 1). El valor taxonómico de las estípulas deriva de su constancia entre parentescos lejanos. Esto es una consecuencia de la aparición temprana durante la ontogenia de la hoja:

Fig. 4. Posiciones diferentes de estípulas (st). A , B : estípulas exteriores de la vista lateral; C , D : estípulas interiores de la vista ventral ( D ), C explicando la posición de las estipulas x x en los márgenes basales de la hoja; E , F : esquemata de estípulas adnatas ( E ) y libres ( F ); G , H : diagramas para explicar el cambio de la posición hacia la formación de una estípula opuesta como en el caso de Coronilla montana ( I ); M : Melianthus major , hoja pinnada con estípula axilar; St: peciolo, st estípula (todos según W. Troll, por partes modificado).

Fig. 5. Carpinus betulus L., A : yema hibernante en el eje madre, en estadio de brotadura; B: cicatriz de la hoja tectriz. B : hoja joven, todavía plegada, con las estípulas libres, que han servido como catáfilos (como los pares de estípulas st 1 , st 2 , st 3 en A ), L 1 , L 2 , L 3 los ápices de hojas (según W. Troll).
Tabla 1. Presencia o confirmación de la existencia de estípulas en familias de dicotiledóneas. Abreviaturas = estípulas presentes: +, estípulas ausentes: -, estípulas pequeñas: peq., estípulas rudimentarias: r, confirmado: conf.

Las estípulas aparecen inmediatamente después de la primera subdivisión de un primordio formando la parte basal, el 'Unterblatt' (Eichler, 1861) y la parte distal, el 'Oberblatt', que usualmente produce la lámina y el pecíolo. Muchas veces presentan el primer signo de la subdivisión del primordio (Fig.6 A, 8 A , 10 A). Durante las fases siguientes, las estípulas se desarrollan muy rápido adelantándose a todas las otras partes de la hoja y no raramente aventajando a la propia hoja (Fig . 6 E, 3 A ). La aparición temprana y este desarrollo precoz ('proléptico') es muy característico para estípulas y puede ser tratado como un ejemplo del criterio de la calidad especial. En muchas hojas pinnadas, en las cuales la formación de los segmentos de la lámina se realiza basípetamente, las estípulas ya están bien desarrolladas, mientras que la segmentación basípeta de la lámina todavía continúa (Fig. 10 B-D, 13 B-E). Según los ejemplos investigados, el crecimiento de las estípulas comienza en forma apical y más tarde está seguido por un crecimiento intercalar en su base (Bolze, 1971, véase también Reitberger, 1953). Probablemente, la aparición temprana durante la ontogenia es la causa de la constancia de las estípulas (Weberlin G , 1958). Esto también vale cuando éstas son rudimentarias como - entre otros - es el caso de la mayoría de las Capparales y las Mirtales (Weberling, 1956, 1957, 1958a, b, 1960).
Estípulas Rudimentarias
Un buen ejemplo que nos muestra la aparición temprana de las estípulas y su crecimiento precoz, es la ontogénesis de las hojas de Reseda alba Lag. (Fig. 7). Donde se observa además, que la formación de los segmentos de la lámina pinnada se realiza basípetamente (como Eichler, 1861, ya había constatado para las Resedaceae y otras familias) y todavía progresa, después que las estípulas en la base ya han terminado su desarrollo. Esto confirma, que Eichler tenía razón, cuando ha definido la parte basal, 'Unterblatt', como parte esencial de la hoja, que forma una zona de desarrollo autónoma. Lo mismo se aplica a Capsella bursa-pastoris (L.) Med. (Fig. 8), la cual al igual que la mayoría de las Brassicaceae y muchas Capparales tiene estípulas rudimentarias con la función de glándulas productoras de mucílago. Esta sustancia mantiene los primordios de las hojas humedecidas durante la fase sensible de su ontogenia. En el estadio adulto de las hojas sus estípulas ya han terminado su actividad y frecuentemente están muertas.

Fig. 6 . Estípulas de Onagraceas, A - D : Lopezia coronat a Andr., A cono vegetativo con dos primordios, en el segundo ya han aparecido las estípulas; B : hoja con estípulas en un estadio poco más tarde que A ; C , D : parte del eje con dos bases de hojas, de la hoja enfrente han cortado el parte distal de la hoja y el brote axilar, en D las estípulas han caído, véase las estrías que bajan de las inserciones de las estípulas; E , F : Circaea lutetiana L., dos primordios de hojas, en el primero las estípulas ya han aventajado considerablemente el primordio del 'Oberblatt'; G : Jussieua grandiflora Ruiz & Pav., eje con las zonas basales de dos hojas con estípulas; H : Fuchsia virgata Sw., corte transversal por una estípula ( H según W. Troll, otros de Weberling 1956a).

Fig. 7. A - F : Reseda alba Lag., estadios sucedentes de la ontogenia de las hojas foliáceas pinnadas, mostrando la sucesión ontogenética basipétala de los foliolos (de Weberling 1968a).

Fig. 8. Capsella bursa-pastoris (L.) Med. , A - E : estadios de la ontogenia de la hoja pinnada y sus estípulas; E : hoja juvenil con estípulas (de Weberling 1955a).
G. Krause (1846) probablemente fue el primero, que ha mencionado estípulas en algunas Crucíferas. En un estudio profundo y amplio J. M. Norman (1857) ha indicado la presencia de estípulas para más de 130 especies de 69 géneros de las Cruciferae (según el sistema de O. E. Schulz 1936). Sus resultados fueron citados por Glück (1919). Lubbock (1895, p. 469) encontró estípulas en 6 especies. Nuestras investigaciones han podido confirmar y ampliar los resultados de Norman para 190 especies de 115 géneros con estípulas (en 7 géneros no los hemos hallado) en base a un estudio morfológico comparativo de la base de las hojas (Weberling, 1955). No obstante, la presencia de estípulas rudimentarias en estas familias todavía ha pasado inadvertida en algunos tratados de sistemática. Todavía en 1936 O. E. Schulz en su tratado de las Cruciferae en la obra de Engler-Prantl-Harms (Die natürlichen Pflanzenfamilien, Bd. 17b, 227 ff.) escribió: "Nebenblätter fehlend oder in verkümmerter Form nachweisbar","Las estípulas faltan o pueden ser comprobadas en forma rudimentaria" aunque más tarde menciona estípulas para Hesperis matronalis y Biscutella laevigata. A veces estaban clasificadas solamente como 'glándulas', especialmente las estípulas de las Brassicaceae (Thellung en Hegi, 1919, y otro autores). Recién, en su tratado de la familia por Appel & Al-Shehbaz (2003) en "The Families and Genera of Vascular Plants" vol. V3 constataron: "Stipules are apparently absent from mature leaves, but papillae and small glands at the leaf base are ubiquitous in the Cruciferae (Weberling, 1956). These have been interpreted as stipules by Krause (1846) and Glück (1919)". Dejando aparte, que los autores han olvidado mencionar el trabajo fundamental de Norman (1857) el párrafo citado no toma en cuenta que yo explícitamente he confirmado que la mayoría de las Cruciferae tienen estípulas rudimentarias, que todavía están presentes en las hojas adultas (Fig. 8 E) (lo mismo dice el encabezamiento de la tabla sobre mis resultados).
Las Crucíferas presentan una serie de formas que se extiende desde estípulas grandes y verdes ( Hesperis matronalis L., Notoceras bicorne Am. y Lunaria annua L.) hasta estípulas muy reducidas aleznadas o de otras formas en muchas otras especies. Las estípulas de Alliaria officinalis Andrz., Eruca sativa Lam. y otras, llevan pelos. En las hojas basales de Armoracia rusticana P. Gaertn. & B. Mey . son triangulares y tienen 3 mm de largo, en Matthiola valesiaca Gay (Fig. 9) están densamente cubiertas con pelos estrellados y emergencias glandulosas (¿entonces son glándulas que llevan glándulas?). Las estípulas de Ochradenus baccatus Del. (Resedaceae) alcanzan una longitud de ca. 0,5 cm , y en las Capparidaceae hay estípulas más grandes (por ejemplolas estípulas espinosas) al lado de las estípulas rudimentarias.

Fig. 9. Matthiola valesiaca Gay, A : parte proximal de una hoja tectriz (D) y estípula (St); H: eje primario; A: eje de un brote axilar, ß: un de sus profilos; B: enseña, que la estípula (como toda la planta) esta cubierta con pelos estrellados y emergencias glandulosas (de Weberling 1955).
Como muestra la Fig. 9, Matthiola valesiaca tiene estípulas partidas en dos o tres lacinias que se originan de una base común (que frecuentemente es indicio de la reducción). No es raro, que en lugar de una estípula más o menos aleznada se encuentren dos o tres o más lacinias, como en Fibigia clypeata (L.) Medik. Las hojas basales de esta planta llevan una estípula larga en ambos lados, mientras que las hojas superiores llevan dos o tres lacinias que se originan de una base común, las brácteas llevan lacinias simples en ambos lados así como en las hojas basales.
En muchos casos la posición a ambos lados de la inserción de la hoja, que ocupan las estípulas rudimentarias al inicio de la ontogenia, cambia en el estadio adulto a una posición más o menos axilar. Esto resulta de un crecimiento lateral de la base de la hoja, causando que el margen primario sobresalga sobre el margen secundario, como muestran las Figs. 11 A* y B* de Rapistrum rugosum All. Si este "desplazamiento" ocurre en combinación con la multiplicación de las lacinias estipulares ya descrita arriba, éstas pueden formar dos series axilares, como muestra la ontogenia de las hojas de Morisia hypogaea J. Gay (Fig. 10).

Fig. 10 . Morisia hypogaea J. Gay, a - e : ontogenia de las hojas; k : hoja adulta; f base de una hoja siguiente d ; g : base de la hoja situado antes de la hoja e ; h base de una hoja siguiente e , con el numero de las lacinias estípulares aumentadas; i : base de la hoja k con yema axilar y lacinias estípulares más aumentadas y distendidas por el crecimiento lateral de la base de la hoja. (De Weberling, 1955).
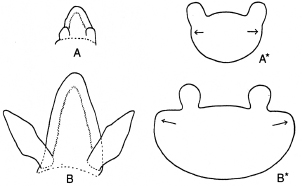
Fig. 11 . Rapistrum perenne All. A , A * y B , B *: dos estadios siguientes de la ontogenia de la hoja, mostrando el desarollo precoz y el cambio de la posición lateral de las estípulas ( A, A*) a una posición axilar ( B, B* ), debido a un crecimiento lateral de la base de la hoja, causando un curso secundario del margen (de Weberling 1955).
Solamente la vecindad de la yema axilar queda libre de lacinias. Lo mismo se puede observar en especies de Cheiranthus, Erysimum, Diplotaxis, Crambe, Neslea y otras. En todos los casos la ontogenia no deja lugar a duda, de que se trata de estípulas! Esta interpretación resulta de un espectro continuo de las formas. Esta afirmación no solamente se refiere a las estípulas de las Capparales (Capparaceae, Resedaceae, Brassicaceae, Tovariaceae, Moringaceae, también en las Koeberliniaceae , Weberling, 1975b), sino también a muchas otras familias u órdenes para los cuales hemos podido descubrir o confirmar estípulas, a veces exclusivamente rudimentarias, como caracteres constantes (véase Tabla 1).
El Fenomeno de Reproducción en la Morfologia Comparada
Uno de los argumentos repetidas veces usado contra la interpretación de apéndices basales como estípulas rudimentarias es la ausencia de hacecillos conductores, que ha motivado a Lothelier (1893) a poner en duda la interpretación de las espinas de Cleome spinosa L. Si esta argumentación, que ya fue emitida por Troll (1937), fuera concluyente, muchas brácteas no serían homólogas de hojas, como las brácteas de algunas especies de Rivina, que carecen de hacecillos (vease Napp-Zinn 1974, 1960). Generalmente las brácteas son caracterizadas como hojas que están inhibidas en su desarrollo completo (hojas más o menos simplificadas). Así se puede explicar la ausencia de hacecillos como consecuencia de la reducción.
Además muchos elementos rudimentarios perdieron su valor para conclusiones sistemáticas. Por ejemplo: en las flores femeninas de Tonina fluviatilis Aublet (Eriocaulaceae) los pétalos están reducidos a lóbulos pequeños. Las flores masculinas de Lachnocaulon carecen de pétalos, mientras que en las flores femeninas están reducidos a mechones de pelos, que por lo menos indican la ubicación de los pétalos (Stützel, 1985).
En los pétalos de las especies de Paepalanthus de flores pequeñas no hay hacecillos (Stützel, 1986). En las Typhaceae el perigonio de la flor femenina de Sparganium consta de tépalos homogéneos. En Typha el perigonio de la flor está reemplazado por cerca de tres ciclos de pelos largos, sin ningún vestigio de hacecillos, mientras que en las flores masculinas el perigonio todavía consiste en escamas más o menos laciniadas. Sin duda la posición equivalente de los anillos de pelos y del perigonio muestra que los tépalos homogéneos de las flores femeninas de Sparganium son homólogos a los tres ciclos de pelos largos en las flores femeninas y a las escamas laciniadas masculinas de Typha . (D. Müller- Doblies, 1970, 546, Fig. 41-68).
En relación a este tema, se debe tener en cuenta los comentarios de A. W. Eichler (1875, p. 52), quién expresa que la evaluación de una estructura como un órgano rudimentario o no, esencialmente resulta de la comparación entre taxones diversos. Para ilustrar el fenómeno de reducción de estípulas se mostrarán algunas observaciones en Viola tricolor L . que pueden ser instructivas. Como es bien conocido, las hojas de esta planta tienen estípulas foliáceas grandes (Fig. 12 A-C), que especialmente en la zona de la inflorescencia son pinnadas y repiten aproximadamente la forma de la lámina con su pecíolo. En las láminas como en las estípulas entre todos los dientes hay una emergencia glandulosa en forma de maza (Fig. 12 G). Las hojas tan grandes y foliáceas son precedidas por una serie de hojas primarias con un pecíolo largo y una lámina pequeña. Las hojas distales de esta serie llevan estípulas más pequeñas y aleznado-lanceoladas (Fig. 12 E, E*), no obstante estas también llevan una glándula en su cima, la que todavía se encuentra en las hojas mas bajas, en las cuales solamente hay una estípula muy corta (Fig. 12 G). ¡En las hojas precedentes en el lugar de la estípula solamente ha quedado la glándula! (Fig. 12 D, D*). En los primeros 2 o 3 miembros de la serie de hojas primarias usualmente faltan los vestigios de las estípulas, no obstante, a veces tienen una estípula glandulosa (Weberling, 1955a, p. 92).
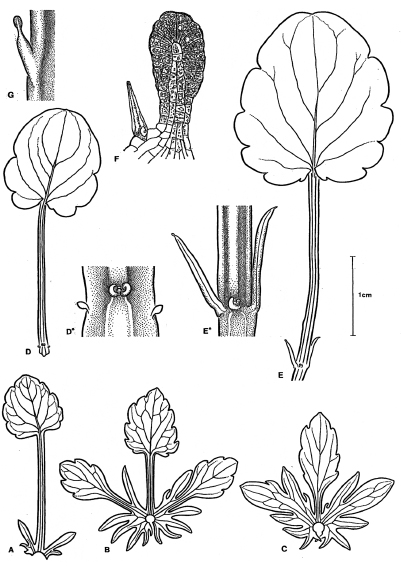
Fig. 12. Viola tricolo r L., A - C : secuencia de hojas, A : basal; B , C : hojas caulinares, B : intermedia y C : superior (según W, Troll). D , E : tercero (D) y cuarto (E) hoja primaria, D *, E *: las bases de estas hojas con sus estípulas en escala más aumentada; F : glándula como se encuentran generalmente en la cima y en los dientes de las estípulas de la forma A - C, al lado de la estípula un pelo simple; G : estípula de la quinta hoja primaria que termina en una glándula terminal como las estípulas en E / E *. (G según Strasburger, 5. ed. 1902, D - E orig.).

Fig. 13. Myriophyllum pinnatum (Walt.) Britton. A : hoja adulta; B - E : estadios de la ontogenia; F estípula más aumentada (de Weberling 1956a).
Sin duda podemos identificar a la glándula como una estípula rudimentaria, debido a que este residuo está conectado con las estípulas foliáceas más grandes y pinnadas por una serie continua de formas que ocurren en las hojas del mismo individuo.
Valor Taxonómico y Diagnostico de las Estípulas Rudimentarias
Ya hemos mencionado el valor taxonómico muy alto de las estípulas. Este valor está reforzado cuando familias u órdenes están caracterizados por estípulas rudimentarias, como en el caso de las Mirtales o las Capparales (entonces la reducción de las estípulas puede ser evaluada según el criterio de la calidad especifica). Por otro lado, para distinciones diagnósticas pueden resultar dificultoso aplicar este carácter, porque las estípulas rudimentarias frecuentemente no son visibles. Por eso en claves se recomienda una nota: 'estípulas rudimentarias, en la mayoría no visibles sin lupa'.
Otros Apéndices Basales de las Hojas
Además de las estípulas debemos distinguir otros apéndices foliáceos formados en la base de la hoja, llamados aurículas (aurículas basales) que se observan frecuentemente en las Compositae . Estos pueden resultar de la amplificación de la vaina en las hojas (Fig. 14, Fig. 17 H - L) o por un ensanchamiento basal semejante a la lámina que forma apéndices foliares (Fig. 15, Fig. 17 M - P).

Fig. 14 . Ligularia clivorum Maxim., A : hoja de la base del eje, B: su base más ampliada.; C : hoja del medio del tallo, D:su base más ampliada (de Weberling 1955).
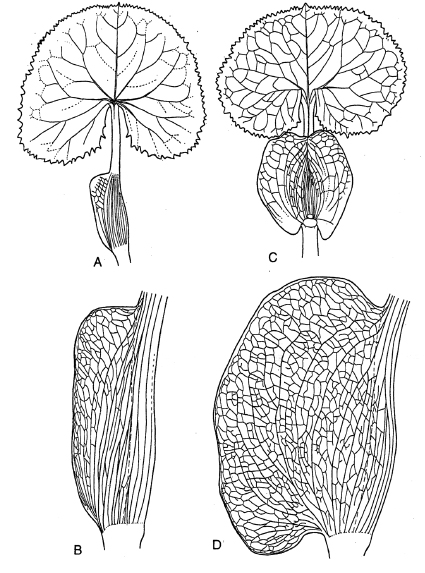
Fig. 15 : Adenostyles albifrons Rchb. A - D : bases de una secuencia de las hojas de la base de la planta y hojas caulinares, E - H : secuencia de hojas caulinares superiores hasta la formación de brácteas (de Weberling 1955).
Las aurículas basales que ocurren en algunas Caprifoliaceae (Weberling, 1957), antes fueron interpretadas como estípulas (especies de Lonicera, Leycesteria ). En Lonicera ferdinandii Franch. (Fig. 16 C) las aurículas son muy parecidas a estípulas interfoliares, pero al contrario de las estípulas interfoliares de las Rubiaceae (Fig. 16 D, E) muestran una ontogenia tardía (Fig. 16 A , B) y no son elementos constantes. Entre las Plumbaginaceae, Plumbago auriculata Lam. (también, aunque menos pronunciado, y en pocos otros taxones) se caracteriza por tener aurículas basales (Weberling, 1956b).

Fig. 16 . A - C Lonicera ferdinandii Franch., verticilo de dos hojas foliáceas, mostrando la ontogenia tardía de las auriculas interfoliares; D : Coffea arabica L. ( Rubiaceae ), verticilo de hojas con una estípula interfoliar; - E , F : Manettia reclinata L. ( Rubiaceae ), verticilo de hojas con una estípula interfoliar, que - al contrario a las aurículas - aparece muy temprano; las estípulas de ambas hojas ya unidas en una base común ( E ), en F la estípula interfoliar adulta lleva una serie de glándulas (A - C de Weberling 1957, D - E de S. Peters, 1962/63).
Además tenemos que mencionar las pseudoestípulas, que son folíolos (pinnas) de la lámina que ocupan la posición de estípulas y más o menos tienen el aspecto de éstas como en muchas Burseraceae, Simarubaceae4 , Meliaceae, Anacardiaceae, Sapindaceae (Weberling & Leenhouts, 1966; Weberling, 1976a) y en algunas Caprifoliaceae ( Sambucus5 , Viburnum ; Weberling, 1957). La homologación como pinnas resulta del estudio de la ontogenia. No obstante en algunas Burseraceae, especialmente del género Canarium hay apéndices que no solamente en su forma, sino también en su desarrollo no difieren de las estípulas, aunque series de las formas indican que son equivalentes a las pinnas. Por eso en este caso han recibido la designación de 'Meta-Estípulas' (Lam 1932).
En un sentido más amplio las aurículas frecuentemente están incluidas bajo el término 'pseudoestípulas' .
En plantas que tienen una vaina bien desarrollada se pueden observar lóbulos del margen superior que están más o menos destacados en Ranunculus repen s L. Al contrario de la ontogenia de las estípulas, éstos deben su origen a un crecimiento marginal en un estadio tardío de la ontogenia de la hoja y se destacan más y más en las hojas. Por eso, con algunas excepciones, la aparición de estos lóbulos de la vaina (Fig. 17 K) no es constante. En las Apiaceae, Ranunculaceae, Berberidaceae y otras familias hay formas transicionales en las cuales la vaina se acorta durante la secuencia de las hojas de un tallo y los lóbulos aparecen más y más temprano desarrollándose también precozmente como las estípulas. Así, los apéndices basales de las hojas deRanunculus aquatilis L . y las hojas caulinares (véase FONT QUER, 1975, p. 195) de Isopyrum fumarioides L . (Ranunculaceae) como en las Apiaceae Hydrocotyle, Bowlesia y Drusa glandulosa (Poir.) Bornm. son estípulas. Estos pasos del desarrollo de estípulas de una vaina se puede imaginar como un camino filogenético de estípulas (Weberling, 1973, 1975a). Domin (1911, p. 151), que particularmente se ha dedicado al estudio de las hojas de monocotiledóneas y ya tenía un concepto similar al nuestro sobre el camino filogenético de "estípulas".
Comentarios a la Filogenia de las Estípulas
La pregunta si la presencia de estípulas es un carácter plesiomórfico, todavía no tiene respuesta. Takhtajan (1959, 61, 65/66), refiriéndose a Hallier (1912), Sinnott & Bailey (1914) y otros autores, opinó que sí, partiendo del supuesto que las estípulas estaban presentes en todas las Angiospermas primitivas y destacando la presencia de estípulas en muchas Magnoliaceae , mientras que la ausencia en otras familias (Degeneriaceae, Himantandraceae, Winteraceae, Schisandraceae, Trochodendraceae) está considerada como resultado de la reducción. El mismo autor remite también a la estructura trilacunar de los nudos en la mayoría de las familias mencionadas, que según Sinnott & Bailey (1914) está correlacionada con la presencia de estípulas, mientras que muchos grupos caracterizados por nudos unilacunares aparecen más adelantados en todas sus estructuras y carecen de estípulas. Su conclusión general es que el tipo de la hoja más primitivo de las Angiospermas recientes es la hoja simple, entera, siempreverde, con nervadura pinnada y con dos estípulas grandes. En este sentido la ausencia de estípulas sería un resultado de reducción.

Fig . 17. Vista general de las posibilidades de las formaciones de la base de hojas ("Unterblatt") de plantas dicotiledóneas (de Weberling 1958b).
No obstante, a pesar de la ampliación de los conocimientos referentes a Angiospermas primitivas, no tenemos aún una respuesta definitiva6 . Además, si consideramos la distribución de las estípulas entre los grupos de las Angiospermas no podemos excluir, que la evolución de estípulas ha ocurrido varias veces e independientemente en diferentes -aun cuando pocos- parentescos de Angiospermas. Por lo menos, las formas transicionales entre lóbulos de la vaina y estípulas ya mencionadas como casos excepcionales entre las Ranunculaceae y Apiaceae favorecen este punto de vista.
La Biologia Molecular y la Morfologia Comparada
Como las estípulas son órganos muy constantes y su aparición significa uno de los primeros pasos durante el desarrollo de una hoja, sería muy interesante, investigar la acción de los genes codificando su formación. Es de esperar una secuencia jerárquica parecida a la secuencia de genes que determinan el desarrollo de la flor, que en este caso comienza con la separación del "Unterblatt" y "Oberblatt", y de esta manera se podría confirmar el concepto morfológico de Eichler (1861).
Entre tanto hay marcadores de genes que se refieren a las estípulas. Por investigaciones de la ontogenia de las hojas de Arabidopsis thaliana (Tsukaya & Uchimiya, 1997) se ha demostrado que el desarrollo de la hoja es basípeto. Para los morfólogos este resultado no es nada extraordinario, ni nuevo, - la segmentación basípeta fue descrita en general para las Resedaceae por Eichler (1861). Pero su obra no fue mencionada, debido a que Eichler publicaba sus resultados en alemán y no en inglés. También Tsukaya & Uchimiya (1997) han observado que el desarrollo de las estípulas no es influenciado por el desarrollo basípeto de la lámina - como yo había descrito para las Cruciferae Brassicaceae, Resedaceae, Capparaceae y otras familias relacionadas (Weberling, 1955) (Fig. 8, 10) Entonces los resultados de la biología molecular se corresponden bien con nuestros puntos de vista y los de Eichler (1861): las estípulas son apéndices de la base de la hoja, el "Unterblatt" una parte relativamente autónoma Como hay más ejemplos de este tipo (por ejemplo: los resultados de Erbar, 1996, ya mencionados), nos parece muy aconsejable que la biología molecular tome más nota de los entre la biología molecular y la morfología comparada podría ser muy efectiva.
NOTAS
1Conferencia pronunciada en las XXX Jornadas Argentinas de Botánica, Rosario, 6-10 Noviembre 2005.
2Arbeitsstelle Biosystematik, Universität Ulm, Alemania. Email: focko.weberling@extern.uni-ulm.de
3En la "Introduction to Capparales" el editor, K. K UBITZKI , escribió: "Stipules are present in all Capparales families except Setchellanthaceae and Koeberliniaceae. In Capparaceae their presence is variable, and in Resedaceaeand Cruciferae they are often rudimentary."(p.9).
4Recchia y Cadellia (Recchioideae) con estípulas (Weberling et al. , 1980).
5 Especialmente en Sambucus ebulus las pinnas (foliolos) basales frecuentemente son erradamente considerado y descrito com ,estípulas foliosas'. En este caso también en claves analíticas se recomienda una nota: "las hojas con foliolos basales que tienen la apariencia de estípulas".
6Nota del Editor: Debe señalarse que en filogenias recientes (APG II, 2003, Bot. J. Linn. Soc. 141: 399-436) se señala a Amborellaceae, Nymphaeaceae y Austrobaileyales (Illiciaceae + Austrobaileyaceae + Trimeniaceae) como los grupos más ancestrales de Angiospermas, los cuales carecen de estípulas.
BIBLIOGRAFÍA
BATEMAN, R. M. 1999. Integrating molecular and morphological evidence of evolutionary radiations . In: HOLLINGSWORTH, P.M., BATEMAN, R.M. & GORNALL, R.J. (eds.), Molecular Systematics and Plant Evolution . Taylor & Francis, London & New York. [ Links ]
BOLZE, A. 1971. Morphologisch - histologische Untersuchungen über die Entwicklung von Stipeln und Scheidenlappen. Unveröff. Diplomarbeit. Naturw. Fakultät Univ, Gießen. [ Links ]
CHASE, M. W. et al . 1993. Phylogenetics of seed plants: an analysis of nucleotide sequences from the plastid gene rcbL. Ann. Missouri Bot. Gard . 80: 528-580. [ Links ]
COEN, E. S. & E. M. MEYEROWITZ1. 1991. The war of whorls: Genetic interactions controlling flower development. Nature 353: 31-37. [ Links ]
COLOMB, G. 1887. Recherches sur les stipules. Ann. sc. nat., 7. sér., Bot . 6 : 1. [ Links ]
DOEBLEY, J. 1993. Genetics, development and plant evolution. Curr. Opinion Genet. Develop. 3: 865-872. [ Links ]
DOMIN, K. 1911. Morphologische und phylogenetische Studien über die Stipularbildungen. Ann.Jard. Bot. Buitenzorg 24: 117-326. [ Links ]
ECKARDT, TH. 1964. Das Homologieproblem und Fälle strittiger Homologien. Phytomorphology 14: 79-92. [ Links ]
EICHLER, A. W. 1861. Zur Entwicklungsgeschichte des Blattes mit besonderer Berücksich-tigung der Nebenblatt- Bildungen. Diss. Marburg. [ Links ]
EICHLER, A. W. 1875/78. Blüthendiagramme I (1875) y II (1878) . Engelmann, Leipzig. Reprint 1954, O. KOELTZ, Eppenhain. [ Links ]
ERBAR, C. 1996. Distribution of Character States "Early Sympetaly" and "Late Sympetaly"Within the "Sympetalae Tetracyclicae" and Presumably Allied Groups . Botanica Acta 109: 427-440. [ Links ]
GLÜCK, H. 1919. Blatt- und blütenmorphologische Studien. G. FISCHER, Jena. [ Links ]
KRAEMER, H. 1897. Viola tricolor in morphologischer, anatomischer und biologischer Beziehung. Diss. Marburg 1897. [ Links ]
KRAUSE, G. 1846. Einige Bemerkungen über den Blumenbau der Fumariaceae und Cruciferae . Bot. Ztg., hrsg. MOHL & SCHLECHTENDAHL, 1846, 121-125, 137-150, p. 142-147. [ Links ]
LAM, H. J. 1932 . Beiträge zur Morphologie der Burseraceae, insbesondere der Canarieae II . Ann. Jard. bot. Buitenzorg 42 : 97-226. [ Links ]
LOTHELIER, A. M. 1893. Recherches anatomiques sur les épines et les aiguillons des plantes . Thèse, Lille. [ Links ]
LUBBOCK, Sir J. 1891/95. On Stipules, their Form and Function. Part I. J. Linnean Soc . 28:217-243 (1891), Part II, 30: 463-532 (1895). [ Links ]
MÜLLER-DOBLIES, D. 1970. Über die Verwandtschaft von Typha und Sparganium in Infloreszenz- und Blütenbau. Bot. Jahrb. Syst . 89: 451- 562. [ Links ]
MÜLLER-DOBLIES, U. 1970. Über die Blütenstände und Blüten sowie zur Embryologie von Sparganium . Bot. Jb. Syst . 89: 359-450. [ Links ]
NAPP-ZINN, K. 1974. Anatomie des Blattes, 2. ed., II. Blattanatomie der Angiospermen . Gebr. BORNTRAEGER, Berlin, Stuttgart. [ Links ]
NORMAN, J. M. 1857. Quelques observations de morphologie végétale. Programme de l'Université. Christiania 1857. Abgedruckt ohne Beigabe der Abbildungen in: Ann. sc. nat., sér. 4, Bot . 9: 105. [ Links ]
PATTERSON, C. (ed.). 1985. Molecules and Morphology in Evolution: Conflict or Compromise? I. Internat. Congress of Systematic and Evolutionary Biology 3, Introduction: 1-27.Brighton. [ Links ]
PETERS, S. 1962/63. Vergleichende und entwicklungsgeschichtliche Untersuchungen über Stipularbildungen bei Rubiaceen. Unveröff. Staatsexamensarbeit, Univ. Mainz. [ Links ]
REMANE, A. 1956. Die Grundlagen des natürlichen Systems, der vergleichende Anatomie und der Phylogenetik. 2. ed. Akad. Verlagsgesellschaft [ Links ]
REMANE, A. 1956. Die Grundlagen des natürlichen Systems, der vergleichende Anatomie und der Phylogenetik. 2. ed. Akad. Verlagsgesellschaft GEEST & PORTIG K.-G. Leipzig. [ Links ]
RIDSDALE, C. E. 1974. Stipules, stipules, more stipules! Flora Malesiana Bull. 27: 2207. [ Links ]
SCHULZ, O. E. 1936. Cruciferae . In: ENGLER-PRANTLHARMS, Die natürlichen Pflanzenfamilien , 2. Aufl., Bd. 17b: 226-658. [ Links ]
SINNOTT, E. W. & I. W. BAILEY. 1914. Investigations on the Phylogeny of Angiosperms. 3. Nodal Anatomy and the Morphology of Stipules. Amer. J. Bot . 1: 441-453. [ Links ]
SOLTIS, D. E. et al . 1997. Angiosperm Phylogeny Inferred from 18S Ribosomal DNA Sequences. Ann. Missouri Bot. Gard . 84: 1-49. [ Links ]
SOLTIS, P. S., M.W.CHASE, D. C. ALBACH, M. E. MORT, V. SAVOLAINEN & M. ZANIS. 1998. Molecular phylogeny of angiosperms: congruent patterns inferred from three genes. Amer. J. Bot. 85 (suppl.): 157-158. [ Links ]
SOLTIS, P. S. & J. J. DOYLE. 1998. Molecular Systematics II . Chapman & Hall, New York. [ Links ]
STÜTZEL, Th. 1985. Die Bedeutung monothecatbisporangiater Antheren als systematisches Merkmal zur Gliederung der Eriocaulaceen. Bot. Jahrb. Syst . 105: 433- 438. [ Links ]
STÜTZEL, Th. 1986. Die epipetalen Drüsen der Gattung Eriocaulon (Eriocaulaceae). Beitr. Biol. Pflan. 60: 271- 276. [ Links ]
TAKHTAJAN, A. 1959. Die Evolution de Angiospermen. Translated from Russian by W. HÖPPNER. G. FISCHER, Jena. [ Links ]
THE ANGIOSPERM PHYLOGENY GROUP. 1998. An ordinal classification for the families of flowering plants. Ann. Missouri Bot. Gard . 85: 531-553. [ Links ]
THEISSEN, G., A. BECKERWINKLER, K. U. MÜNSTER, T. G. KIRCHNER & H. SAEDLER. 2002. How the Landplants learned their floral ABCs: the role of MADSbox genes in the evolutionary origin of flowers. In: Cronk et al . (eds.): Developmental Genetics and Plant Evolution. [ Links ]
TAYLOR & FRANCIS, London. THELLUNG, A. 1919. Cruciferae. In: G. Hegi, Illustrierte Flora von Mitteleuropa IV/1, 51-491. [ Links ]
TRECUL, A. 1853. Mémoire sur la formation des feuilles. Ann. sc. Nat., 3.sér., Bot . 19: 235-314. [ Links ]
TROLL, W. 1937. Vergleichende Morphologie der höheren Pflanzen, Vol. I/1 . Gebr. Borntraeger, Berlin. [ Links ]
TROLL, W. 1949. Die Stiel-Spreiten-Relation als Ausdruck des Prinzips der variablen Proportionen. Naturwissenschaften 36: 333?-338. [ Links ]
TSUKAYA, H. & A. H. UCHIMIY. 1997. Genetic analyses of the formation of the serrated margin of leaf blades in Arabidopsis : combination of a mutational analysis of leaf morphogenesis with the characterization of a specific marker gene expressed inn hydathodes and stipules. Molec. Gen. Genet . 256: 231-238. [ Links ]
WEBERLING, F. 1955a. Morphologische und entwicklungsgeschichtliche Untersuchungen über die Ausbildung des Unterblattes bei dikotylen Gewächsen. (Diss.1953) Beitr. Biol. Pflan. 32: 27-105. [ Links ]
WEBERLING, F. 1955b. Die Stipularbildungen der Coriariaceae . Flora 142: 629-630, Taf. IX. [ Links ]
WEBERLING, F. 1956a. Untersuchungen über rudimentäre Stipeln bei den Myrtales . Flora 143: 201-218. [ Links ]
WEBERLING, F. 1956b. Weitere,Untersuchungen zur Morphologie des Unterblattes bei den Dikotylen. I. Balsaminaceae; II Plumbaginaceae. Beitr. Biol. Pflanzen 33: 17-32. [ Links ]
WEBERLING, F. 1956c. Weitere Untersuchungen zur Morphologie des Unterblattes bei den Dikotylen. III. Convolvulaceae; IV. Zygophyllaceae. Beitr. Biol. Pflanzen 33: 149-161. [ Links ]
WEBERLING, F. 1957. Morphologische Untersuchungen zur Systematik der Caprifoliaceen. Abh. Akad. Wiss. Lit. Mainz. math.-naturw . Kl. Nr. 1. [ Links ]
WEBERLING, F. 1958a. Über das Vorkommen rudimentärer Stipeln bei den Lecythidaceae (s.l.) und Sonneratiaceae. Flora 145: 72-77. [ Links ]
WEBERLING, F. 1958b. Die Bedeutung blattmorphologischer Untersuchungen für die Systematik (dargestellt am Beispiel der Unterblattbildungen). Bot. Jahrb . 77: 458-468. [ Links ]
WEBERLING, F. 1960. Weitere Untersuchungen über das Vorkommen rudimentärer Stipeln bei den Myrtales ( Combretaceae, Melastomataceae ). Flora 149: 189-205. [ Links ]
WEBERLING, F. 1963. Ein Beitrag zur systematischen Stellung der Geissolomataceae, Penaeaceae und Oliniaceae sowie der Gattung Heteropyxis (Myrtaceae). Bot. Jahrb . 82: 119-128. [ Links ]
WEBERLING, F. 1968a. Über die Rudimentärstipeln der Resedaceae. Acta Bot. Neerl . 17: 360-372. [ Links ]
WEBERLING, F. 1968 b. Bemerkungen über das Vorkommen rudimentärer Stipeln. I. Cyrillaceae und die Gattung Cyrillopsis KUHLM. II Alzatea RUIZ & PAV. (Lythraceae) und Tristania R. B R . Acta Bot. Neerl . 17: 282-287. [ Links ]
WEBERLING, F. 1970a. Die vermeintlichen Stipulardornen bei Zanthoxylum und Fagara (Rutaceae) sowie bei Acanthopanax M IQ . (Araliaceae). Ber. Oberhess. Ges. f. Natur- u. Heilkunde 17: 141-147. [ Links ]
WEBERLING, F. 1970b. Weitere Untersuchungen zur Morphologie des Unterblattes bei den Dikotylen. V. Piperales. Beitr. Biol. Pflanzen 46: 403-434. [ Links ]
WEBERLING, F. 1970c. Weitere Untersuchungen zur Morphologie des Unterblattes bei den Dikotylen. VI. Polygonaceae. Beitr. Biol. Pflan. 47: 127-140. [ Links ]
WEBERLING, F. 1973. Über die Beziehungen zwischen Vaginalloben und Stipeln. (Symposium Morphology and Anatomy, Nijmegen 1973). Acta Bot. Neerl . 22: 247. [ Links ]
WEBERLING, F. 1975a. Über die Beziehungen zwischen Scheidenlappen und Stipeln. Bot. Jahrb . 96: 471-491. [ Links ]
WEBERLING, F. 1975b. Weitere Untersuchungen zur Morphologie des Unterblattes bei den Dikotylen. VII. Polygalales; VIII. Koeberlinia Zucc. Beitr. Biol. Pflanzen 50: 277-289. [ Links ]
WEBERLING, F. 1976a. Die Pseudostipeln der Sapindaceae. Abh. Akad. Wiss. Lit. Mainz, math.-naturw. Kl. Nr. 22 (1976). [ Links ]
WEBERLING, F. 1976b. Weitere Untersuchungen zur Morphologie des Unterblattes bei den Dikotylen. IX. Saxifragaceae s. l., Brunelliaceae und Bruniaceae. Beitr. Biol. Pflan. 52: 163-181. [ Links ]
WEBERLING, F. 1983. Weitere Untersuchungen zur Morphologie des Unterblattes bei den Dikotylen. X. Zur Ausbildung von Stipeln bei den Salvadoraceae und bei Phylica stipularis (Rhamnaceae). Beitr. Biol. Pflanzen 58: 149-155. [ Links ]
WEBERLING, F. 1984. Stipular structures in Myrtales and suspectedly related families. In: DAHLGREN, R. & R. F. THORNE: The order Myrtales: Circumscription, Variation and Relationships. Ann. Missouri Bot. Gard . 71: 633- 699. [ Links ]
WEBERLING, F. 2000. Tepualia stipularis (Myrtaceae), una planta con apéndices stipuliformes y estípulas rudimentarias. Kurtziana 28: 1-10. [ Links ]
WEBERLING, F. 2005. Nebenblätter und Öhrchen - was man an einheimischen Pflanzen noch entdecken kann. Mitteilungen d. Vereins f. Naturwiss. u. Mathematik Ulm/ Donau, Heft 40/41: 1. [ Links ]
WEBERLING, F. & P. W. LEENHOUTS. 1966. Systematischmorphologische Studien an Terebinthales- Familien (Burseraceae, Simaroubaceae, Meliaceae, Anacardiaceae, Sapindaceae). Abh. Akad. Wiss. Lit. Mainz, math.-naturw. Kl. Nr . 10 (1965). [ Links ]
WEBERLING, F., H. LÖRCHER & E. BÖHNKE. 1980. Die Stipeln der Irvingioideae und Recchioideae und ihre systematische Wertung nebst Bemerkungen zur Holzanatomie und Palynologie. Plant Syst. Evol . 133: 261-283. [ Links ]
Recibido el 23 de Noviembre de 2005
Aceptado el 17 de Abril del 2006.

